238 Biochimica et Biophysica Acta, 950 (1988) 238-242 Elsevier
BBA 90113
BBA Report
C l o n i n g a n d s e q u e n c i n g o f b u l l f r o g g r o w t h h o r m o n e c o m p l e m e n t a r y D N A
F u - M i n g P a n a n d W e n - C h a n g C h a n g
Institute of Biochemical Sciences, National Taiwan University, and Institute of Biological Chemistry, Academic Sinica, Taipei (Taiwan, China)
(Received 15 December 1987) (Revised manuscript received 18 March 1988)
Key words: Growth hormone; eDNA sequence; Cloning; Amino acid sequence; Sequence homology; (Bullfrog)
Total mRNA was isolated from the pituitary glands of bullfrog (Rana catesbeiana), purified by affinity
chromatography with oligo(dT)-cellulose columns. The cDNA was synthesized and cloned in Escherichia coil
using EcoRl linkers and pBR322 as vector. The cDNA library was screened by hybridization with
32 P-labeled duck growth hormone (GH) cDNA. A positive clone was selected and sequenced. The full-length
bullfrog GH cDNA contains 950 nucleotide pairs with an open reading frame coding for the precursor GH
of 215 amino-acid residues. The partial amino-acid sequence from the protein confirms that derived from the
cDNA, with Phe as the first residue in the mature bullfrog GH preceded by a 25-residue hydrophobic signal
peptide. The bullfrog GH shares sequence homology with those of other vertebrate species in the following
order, duck (61% protein sequence homology; 67% cDNA homology), rat (56%; 61%), human (47%; 57%) and
salmon (42%; 50%).
The growth hormone (GH) of the vertebrate is synthesized and secreted by the anterior pituitary gland and required for preadult growth of the vertebrate [1,2]. It has been extensively investi- gated both at protein and gene levels in human and other mammals. These studies have been ex- tended to the lower vertebrates, such as avian [3-5] and fish [6-8]. For the bullfrog, a repre- sentative of the amphibia, the biosynthesis and gene expression of GH might be more com- plicated and interesting because it experiences a stage of metamorphosis in its life cycle.
The bullfrog GH has been purified and char- acterized as follows. Fresh pituitary glands of
Abbreviation: GH, growth hormone.
Correspondence: W.C. Chang, Institute of Biological Chem- istry, Academia Sinica, P.O. Box 23-106, Taipei, Taiwan, Re- public of China.
bullfrog
(Rana catesbemna)
were obtained from a local supplier. They were homogenized, and sep- arated on a Sephacryl S-200 column, followed by ion-exchange chromatography on a DEAE-cel- lulose column and identified as described before [9]. The purified bullfrog GH was sequenced in a pulsed-liquid protein sequencer (Applied Biosys- terns. Inc. 477A).The mRNA was purified and cloned as de- scribed previously [5]. Briefly, total RNA was extracted from the fresh pituitary glands by guanidinium isothiocyanate and hot phenol. Poly- adenylated RNA was selected by passing through the oligo(dT)-ceUulose column twice and used for the synthesis of eDNA [10]. The double-stranded eDNA was methylated with
EcoRI
methylase be- fore ligation withEcoRI
linkers and then digested withEcoRI
to generate the cohesive ends for insertion into theEcoRI-treated
pBR322 vector.The eDNA library was screened by hybridiza- tion with 32p-labeled duck GH-cDNA [5]. A posi- 0167-4781/88/$03.50 © 1988 Elsevier Science Publishers B.V. (Biomedical Division)
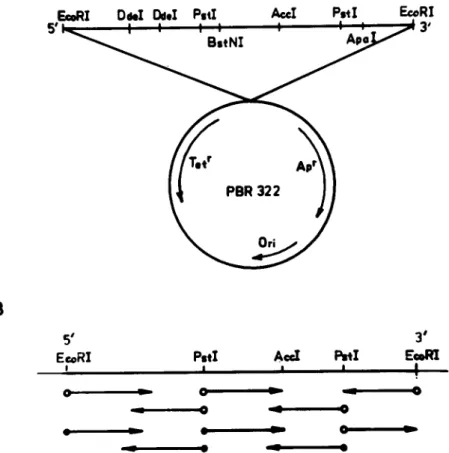
F.~RI D w I I ~ . I P.tI A = I Po*I E ~ R I 5 ' 1 ~ _ I I I I O 0 0 ~.-43'
A
B
# E~RI ! 0 u l 3 oP.tl
AccI
I ~ I
E m l ~ I I I !Fig. 1. Orientation and enzyme map of the bullfrog GH-cDNA in plasmid pBR322 (A) and sequencing strategy (B). The solid circles indicate the starting point of dideoxy sequencing, whereas the open ones are for the end-labeling sites for chemical sequencing.
tive clone c o n t a i n i n g the bullfrog G H - c D N A was selected a n d sequenced b y the chemical m e t h o d o f M a x a m a n d Gilbert [11] a n d the chain-termina- tion m e t h o d o f Sanger [12].
By using the d u c k G H - c D N A as probe, we were able to select a clone c o n t a i n i n g the full-sized bull-frog c D N A which was then sequenced b y the chemical a n d d i d e o x y m e t h o d as s h o w n in Fig. 1. It is evident that this bullfrog G H - c D N A c o n t a i n s an o p e n reading f r a m e e n c o d i n g the p r e c u r s o r G H o f 215 a m i n o - a c i d residues. T h e N - t e r m i n a l a m i n o - a c i d sequence of 30 residues derived f r o m the purified bullfrog G H indicates that the m a t u r e h o r m o n e starts with a Phe in its N - t e r m i n u s a n d c o m p l e t e l y agrees with the sequence derived f r o m the nucleotide sequence (Fig. 2). It c a n be seen that the p r e c u r s o r G H contains a signal p e p t i d e of 25 residues followed b y 190 residues in the m a t u r e
h o r m o n e . W h e n c o m p a r e d with several k n o w n sequences o f G H s f r o m representative vertebrate species (Fig. 3), the bullfrog G H shows significant h o m o l o g y with other G H s , in a c c o r d a n c e with the
TABLE I
SEQUENCE HOMOLOGY BETWEEN BULLFROG GROWTH HORMONE AND GROWTH HORMONES OF OTHER VERTEBRATE SPECIES
GH Protein a cDNA b (~) (%) Human [14] 47 57 Rat [15] 56 61 Duck [5] 61 67 Salmon [7] 42 50
a The precursor sequences are compared.
240 5O G G C G G A A C C C C A A T A T A C A T C A G G A A C A C T G C T A A C C T C A A T C A A T G G C T T C A G G G T T A G M A S G L G i00 G C T C A T C T C T T G T A C T C T T G G T T G T A A T A T G T C T T C A A A G C C C G C A A G G G T T C A A T G C C T S S L V L L V V I C L Q S P Q G F N A F 150 T T C C A C A G A T G T C T C T T T C C A A C C T C T T T A C A A A T G C T G T C A T T C G A G C T C A G C A C C T T C P Q M S L S N L F T N A V I R A Q H L H 200 A C C A G A T G G T T G C C G A T A C C T A C A G A G A C T A T G A A A G A A C A T A T A T A C C T G A A G A T C A G A Q M V A D T Y R D Y E R T Y I P E D Q R 250 300 G A T T C A A A C A A A C A C T T C T T A T T T C A G T G T A T T G C T A C T C A G A A A C C A T C C C A G C A C C A A F K Q T L L I S V Y C Y S E T I P A P T 350 C A G A C A A A G A C A A T A C T C A C C A G A A A T C A G A T A T A G A C C T C C T C C G C T T C T C C C T C A C A C D K D N T H Q K S D I D L L R F S L T L 4 0 0 T C C T G C A G T C A T G G A T G A C C C C T A T C C A A A T C G T C A A C A G A G T T T T T G G C A A C A A C C A G G L Q S w M T P I Q I V N R V F G N N Q v 4 5 0 T G T T T G G A A A C A T T G A C A G A G T T T A T G A C A G A C T G A G G G A C C T C G A T G A A G G A C T A C A C A F G N I D R V Y D R L R D L D E G L H I 500 T T C T C A T A A G A G A G C T G G A T G A T G G A A A T G T T C G G A A C T A C G G T G T G C T G A C T T T T A C A T L I R E L D D G N V R N Y G V L T F T Y 550 600 A T G A T A A G T T T G A T G T C A A T C T G C G C A G T G A A G A A G G C C G G G C G A A G A A T T A C G G T C T A C D K F D V N L R S E E G R A K N Y G L L 650 T C T C C T G C T T C A A G A A A G A C A T G C A T A A A G T A G A G A C C T A C C T G A A G G T G A T G A A A T G C C S C F K K D M H K V E T Y L K V M K C R 700 G G A G A T T T G T G G A G A G C A A C T G C A C C T T T T A A T C A A G A A A A A A A T G T T A C A T C C T C C A T A R F V E S N C T F 750 G T C A C C C C C A C C T C C T G C A G G A C A C C C G C T G C A T T T T T A T A T C T T C A G G A G G T A G G G C C C 8 0 0 A C T G A T A A T T G T C C C T T T T A A A C T T C C T G A C C A A T T G C A C A T T T T T T G C A G A T G T A T T T T 8 5 0 900 G C G T T G T G C T T T T G T T G T C T T T G T A T G T T G T T A A A A A A A T A A A G T A T A G G A G G C A T T T A C A T G A 4 6
B U L L F R O G H U M A N R A T D U C K S A L M O N : : T : S R T : : - - L : A F G L - L : : P W L : E G S : : : T I P : : R : : D : : M L : : H R : : : L A F : : : Q : : A D S Q T P W - - L : T F S L - L : : L W : : E A G : L : A : P : : S : : A : : : L : : : : : : : L A : : : : K : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : M : Q V F L : M P : : : : S - - - : F L : - : : A A I E N : R . . . . : : N I : : S : V : : : : L L A Q K M F N B U L L F R O G H U M A N R A T D U C K S A L M O N * * * * * * * * * * * D Y E R T Y I P E D Q R F : - K Q T L L I S V Y C Y S E T I P A P T D K D N T H Q K S D I - D L L R F S L T L L Q S E F : E A : : : K E : K Y S F L : N P Q T : L - : F : : S : : T : S N R E E : Q : : : N L - E : : : I : : L : I : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : F D G : L L : D E R : Q L N : I F : : D F - - : N : D S : V S : V : : H E : - : : : S V L K : : H I : F R : I E : B U L L F R O G H U M A N R A T D U C K S A L M O N * * * * * * * * * W M T P I Q - - - I V N R - - V F G N N Q V F G N I D R - V Y D R L R D L D E G L H I L I R E L D D G N V R . . . . : L E : V : - - - F L R S : - : : A : S L : Y : A S : S N : : : L : K : : E : : I Q T : M G R : E : : S P : . . . . : L G : V : - - - F L S : - - I : T : S L M : : T S : : - : : E K : K : : E : : I Q A : M Q : : E : : S P : . . . . : L : : V : - - - Y L S K - - : : T : : L : : : T S : : - : F E K : K : : E : : I Q A : M : : : E : R S P : . . . . : E Y : S : T L I : S : S L M : R N A : : . . . . :SE . . . . K : S : : K V : I N L : : T G S Q : : - : L S L D D B U L L F R O G H U M A N R A T D U C K S A L M O N , ** ** ** ** • ** • ** • • N Y G V L T F T Y D K F D V N L R S E E G R A K N Y G L L S C F K K D M H K V E T Y L K V M K C R R F V E S N C T F T G Q I F K Q : : S : : : T : S H N D D A L L : : : : : : Y : : R : : : D : : : : F : R I V Q : : S - : : G S : G : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : D S Q Q L P P : G N Y Y Q : : G G D G N V R R : : E : : A : : : : : : : : : : : : : T : A : : : K S L : A : : : L
Fig. 3. Comparison of amino-acid sequence of GH precursors of bullfrog (the present study), human [14], rat [15], duck [5] and salmon [7]. The colons indicate identical residues. Gaps are introduced to maximize homology. Asterisks indicate the invariant
residues for all sequences.
general concept that the G H is a well-conserved protein hormone in vertebrates. It also shows that 44 residues in the G H molecule are invariant through vertebrate evolution, suggesting that these residues may play important roles in maintaining the correct structure or conformation. A m o n g these invariant residues, the four cysteine residues are essential for the formation of two disulfide bridges; whereas the single T r p - l l 0 (numbering starting from the Met of bullfrog G H precursor) has been implied to form a hydrogen bond with a carboxylate ion in the hydrophobic interior of the G H molecule [13].
Comparison of amino-acid and nucleotide se- quences between bullfrog G H and G H s from other species is shown in Table I. It is interesting that the bullfrog G H sequence is closer to duck G H than to fish G H . This result would shed some light on the phylogenetic and evolutionary relationship of the hormone.
Several mammalian G H genes have been re- ported and the 5'-untranslated sequences corn-
pared [16]. These genomic sequences indicate that the 5'-untranslated region consists of around 60 base-pairs. It is obvious that this 5'-untranslated region has never been fully reverse transcribed from m R N A into cDNA. The bullfrog G H - c D N A contains 45 base-pairs in this region (Fig. 2), pre- sumably incomplete too, but it certainly provides the full-length coding sequence for the precursor G H .
It is also of interest to note that the translation initiator site of bullfrog G H - c D N A contains a T nucleotide at the - 3 position instead of a purine as predicted by the so-called Kozak rule [17]. All the other known G H - c D N A s conform to the Kozak rule for an optimal translation initiation sequence.
Acknowledgement
This work was financially supported by the National Science Council of Republic of China.
242
References
1 Miller, W.L. and Eberhardt, N.L. (1983) Endocrine Rev. 4, 97-130.
2 Chawla, P.K., Parks, J.S. and Rudman, D. (1983) Annu. Rev. Med. 34, 519-547.
3 Souza, L.M., Boone, T.C., Murdock, D., langley, K., Wypych, J., Fenton, D., Johnson, S., Lai, P.H., Everett, R., Hsu, R.Y. and Bosselman, R. (1984) J. Exp. Zool. 232, 465 -473.
4 Farmer, S.W., Papkoff, H. and Hayashida, T. (1974) Endo- crinology 95, 1560-1565.
5 Chen, H.T., Pan, F.M. and Chang, W.C. (1987) Biochim. Biophys. Acta 949, 249-253.
6 Farmer, S.W., Hayashida, T., Papkoff, H. and Polenov, A.L. (1981) Endocrinology 108, 377-381.
7 Sekine, S., Mizukami, T., Nishi, T., Kuwana, Y., Saito, A., sato, M., Itoh, S. and Kawauchi, H. (1985) Proc. Natl. Acad. Sci. USA 82, 4306-4310.
8 Agellon, L.B. and Chert, T.T. (1986) DNA 5, 463-471. 9 Pan, F.M. and Chang, W.C. (1986) J. Chin. Biochem. Soc.
15, 24-33.
10 Gubler, U. and Hoffmann, B.J. (1983) Gene 25, 263-269. 11 Maxam, A.M. and Gilbert, W. (1980) Methods Enzymol.
65, 499-560.
12 Sanger, F., Nicklen, S. and Coulson, A.R. (1977) Proc. Natl. Acad. Sci. USA 74, 5463-5467.
13 Bewley, T.A. and Li, C.H. (1984) Arch. Biochem. Biophys. 233, 219-227.
14 Martial, J.A., Hallewell, R.A., Baxter, J.D. and Goodman, H.M. (1979) Science 205, 602-607.
15 Seeburg, P.H., Shine, J., Martial, J.A., Baxter, J.D. and Goodman, H.M. (1977) Nature (London) 270, 486-494. 16 Vize, P.D. and Wells, J.R.E. (1987) Gene 55, 339-344. 17 Kozak, M. (1981) Nucleic Acids Res. 9, 5233-5252.

![Fig. 3. Comparison of amino-acid sequence of GH precursors of bullfrog (the present study), human [14], rat [15], duck [5] and salmon [7]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8780271.215474/4.853.80.753.82.515/comparison-amino-sequence-precursors-bullfrog-present-study-salmon.webp)
