A Quick Method to Identify Engraulid Fish Larvae in
the Tanshui River Estuary of Northern Taiwan
Yu-Tzu Wang and Wann-Nian Tzeng
Department of Zoology, College of Science, National Taiwan University, Taipei, Taiwan 10617, ROC
Abstract
Five species of larval engraulids, Engraulis japonica Schlegel, Encrasicholina punctifer Fowler, E. heteroloba (Ruppell), Stolephorus insularis Hardenberg, and Thryssa dussumieri (Valenciennes), were found in coastal waters off the Tanshui River Estuary, northern Taiwan during the period from May 1992 through November 1993. Their sizes ranged between 8 and 46 mm SL. Comparing the meristics, morphometrics, and pigmentations of the 5 species, we found pigment patterns on the ventral side of the larvae that can be used as a key character to discriminate these species.
Key words: larval engraulid, species identification, meristics, morphometrics, pigmenta-tion, Tanshui River Estuary.
INTRODUCTION
Engraulid larvae are the dominant compo-nents of the larval fish community in coastal waters of Taiwan (Chen 1985, 1986; Chen and Huang, 1985; Huang et al., 1985; Tzeng and Wang, 1986, 1992; Wang et al., 1991; Wang and Hwang, 1992). The larvae of engraulids and clupeids are named “bull-ard” in Taiwanese. They are harvested for local consumers and greatly contributing to coastal fisheries (Shen, 1971; Cheng, 1980; Chen, 1984; Young et al., 1992). The amount of engraulid larvae consti-tutes ca. 75% of the total bull-ard catch (Cheng, 1980; Wang and Tzeng, 1997). Due to its economic importance, several studies have been conducted on species composition (Liu and Shen, 1957; Shen, 1971; Cheng, 1980), feeding habits (Chern and Tzeng, 1993, 1994), growth (Huang and Chiu, 1996), fluctuation of the catch ( Tsai et al., 1996), and fisher y oceanography (e.g., Lee et al., 1990, 1995, 1996). However, due to the difficulty in classifi-cation of the larvae, the community structure
and population dynamics of engraulids have been little studied (Tzeng and Wang, 1992, 1993, 1997; Wang and Tzeng, 1997).
There are 5 genera and 12 species of engraulids in the coastal waters of Taiwan are revised (Young et al., 1994). Species of adult engraulids are classified mainly based on the numbers of pre- or post-pelvic scutes (Shen, 1984; Chen and Yu, 1986; Whitehead et al., 1988; Nakabo, 1993; Shen et al., 1993; Young et al., 1994). However, these characters are lack-ing in larvae and thus not applicable to the lar-val stage of engraulids.
Chen (1987), Yu and Chiu (1994), and Young et al. (1995) tried to identify the species of engraulid larvae by meristics, morphomet-rics, and pigmentation. Their works have a combination of these characters. However, counting meristics and measuring morpho-metrics are time consuming and difficult when dealing with large numbers of samples (Powles and Markle, 1984). This paper attempts to evaluate the merits and defects of the method of meristics, morphometrics, and pigmenta-Acta Zoologica Taiwanica 8(2): 103-119 (1997)
tion, and to establish a simple key to identify species of engraulid larvae.
MATERIALS AND METHODS Collection of fish larvae
Engraulid larvae were collected from the fishing grounds in coastal waters off the Tanshui River Estuary as in a previous study (Wang and Tzeng, 1997). A commercial set-net was set against the tidal current, to catch the larvae in the estuary during the fishing season from May 1992 to November 1993. The struc-ture and dimension of the net were the same as that of the previous study (Wang and Tzeng, 1997). About 10 g of wet-weight larvae were randomly selected from each daily catch in the fishing season. The larvae were preserved in 95% alcohol and used for species identifica-tion.
Identification of fish larvae
Engraulid larvae were separated from fishes of other families by meristic characters and external morphology (Leis and Rennis, 1983; Wang, 1987; Okiyama, 1988; Leis and Trnski, 1989). Then standard lengths of the larvae were measured and their melanophores were examined and photographed. To identify the species of engraulid larvae, approximately 600 specimens were stained to examine meristics and morphometrics. The measurements of melanophores include the items of head (MH), isthmus (MI), anterior part of gut (MFG), pos-terior of gut (MPG), belly (MB), anal fin base (MAFB), tail (MT), and caudal fin (MC) (Fig. 1a, b). Measurements of meristics including numbers of total vertebrae (TV), vertebrae before anus (VBA), interneurals (INT), and dorsal and anal fin rays (DFR and AFR) (Fig. Figure 1. Measurement of pigmentation, meristics, and morphometrics of engraulid larvae: (a) lateral
view, (b) ventral view. MAFB, MB, MC, MFG, MH ,MI, MPG, and MT denote the melanophores appearing on anal fin base, belly, caudal fin, dorsolateral side of fore gut, head, isthmus, dorsolateral side of posterior gut, and tail, respectively. AFR and DFR, INT, Pc, Pv, and Vert denote the anal and dor-sal fin rays, interneurals, pectoral fin, pelvic fin, and vertebrae, respectively. HL, PAL, PDL, SL, and TL denote the head, preanal, predorsal fin, standard, and total lengths, respectively.
1a) were counted under a stereomicroscope. Morphometric characters of larvae, including total length (TL), standard length (SL), head length (HL), predorsal fin length (PDL), and preanal length (PAL) (Fig. 1a), were measured to the nearest 0.1 mm by profile projector at 10× magnification.
The staining method of larvae is modified from that of Potthoff (1984). The staining process includes 5 steps. 1) Larvae were pre-served with alcohol and put in an acidified alcian blue solution to stain cartilage for 1 day. 2) Larvae were transferred to a saturated sodi-um borate solution for 12 h for neutralization. 3) After neutralization, the bones of larvae were stained with alizarin red and cleared with 1% KOH solution, lasting for only a few min-utes. 4) When vertebrae were nearly clearly seen, larvae were transferred to a solution of 60% glycerin and 40% of 1% KOH for few days to destain the muscle. 5) Finally, stained larvae were preserved in pure glycerin. The bleaching process described by Potthoff (1984) was neglected to maintain the pigments of the lar-vae.
Data analysis
The normality of frequency distribution of standard length was tested by χ2, and the sig-nificant test of difference in mean standard length and mean meristic count among species was conducted by Scheffe’s multiple compari-son test. The regressions of morphometric characters on standard length were fitted with
linear regression. The differences in slope and adjusted mean of the regression lines among species were determined by analysis of covari-ance (ANCOVA) (Steel and Torrie, 1980).
RESULTS
Length frequency distribution
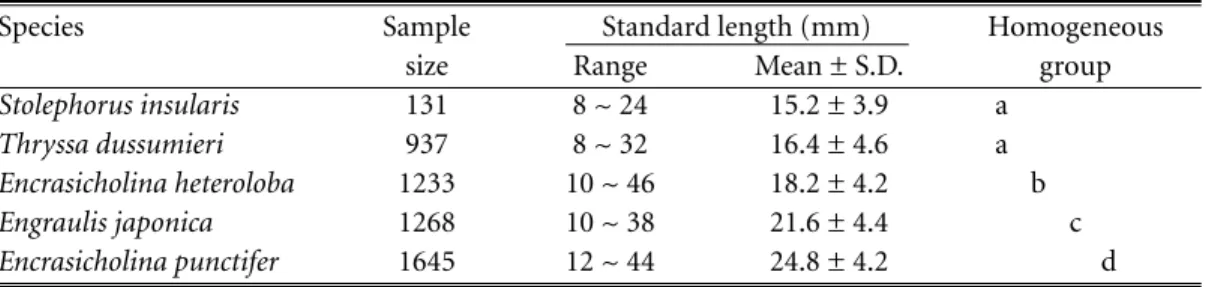
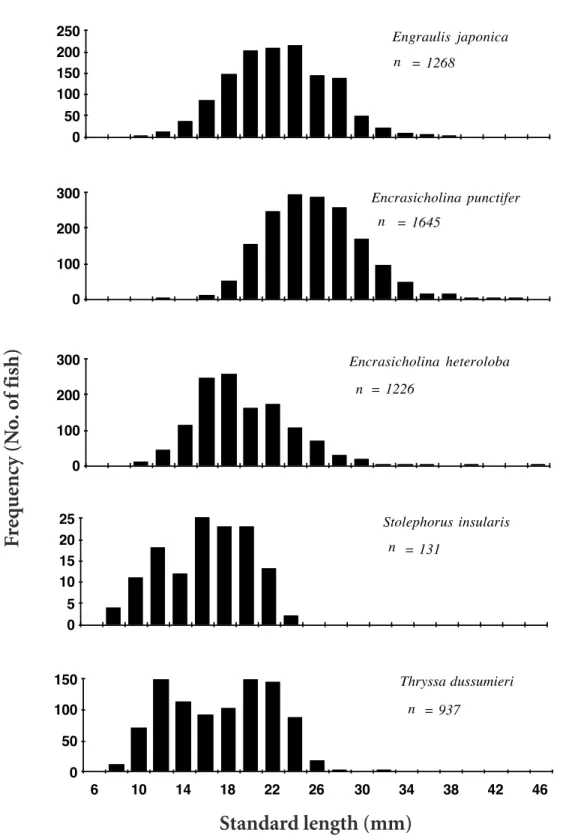
Five species of engraulid larvae were identi-fied, and their length frequency distributions are shown in Figure 2. Standard lengths of Engraulis japonica Schlegel ranged between 10 and 38 mm (21.6 ± 4.4 mm, mean ± standard deviation), Encrasicholina punctifer Fowler, 12 and 44 mm (24.8 ± 4.2 mm), E. heteroloba (Ruppell), 10 and 46 mm (18.2 ± 4.2 mm), Stolephorus insularis Hardenberg, 8 and 24 mm (15.2 ± 3.9 mm), and Thryssa dussumieri (Valenciennes), 8 and 32 mm (16.4 ± 4.6 mm). Scheffe’s multiple comparison test indicated that the differences of mean standard length among the 5 species were S. insularis = T. dus-sumieri < E. heteroloba < E. japonica < E.
punc-tifer (Table 1).
Morphological descriptions
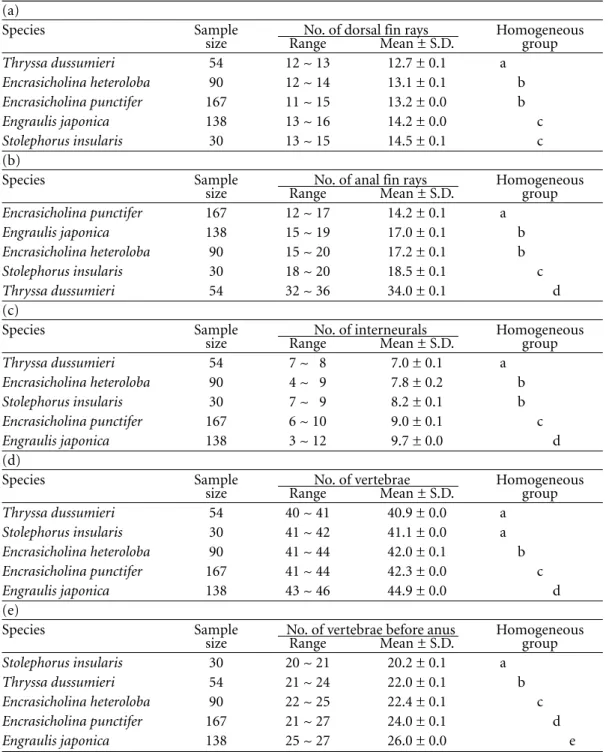
The degrees of ossification of the 5 species of engraulid larvae are shown in Figure 3. Vertebrae and fin rays, when ossified, were stained red. Dorsal, anal, and caudal fin bases and interneurals which remained cartilagenous were stained blue. Skulls appeared either red or blue or in combination, indicating that skulls were in the process of calcifying from cartilage to bone. The relative position of the dorsal fin Identification of Engraulid Larvae
Table 1. Homogeneity test for the standard lengths of the 5 species of engraulid larvae collected in coastal waters off the Tanshui River Estuary. Species sharing the same letter are in the same homoge-neous group.
Species Sample Standard length (mm) Homogeneous
size Range Mean ± S.D. group
Stolephorus insularis 131 8 ~ 24 15.2 ± 3.9 a
Thryssa dussumieri 937 8 ~ 32 16.4 ± 4.6 a
Encrasicholina heteroloba 1233 10 ~ 46 18.2 ± 4.2 b
Engraulis japonica 1268 10 ~ 38 21.6 ± 4.4 c
0 50 100 150 200 250 Engraulis japonica = 1268 n 0 100 200 300 Encrasicholina heteroloba = 1226 n 0 5 10 15 20 25 Stolephorus insularis = 131 n 0 50 100 150 6 10 14 18 22 26 30 34 38 42 46 Thryssa dussumieri = 937 n 0 100 200 300 Encrasicholina punctifer = 1645 n
Figure 2. Length frequency distributions of engraulid larvae collected by a commercial set-net in coastal waters off the Tanshui River Estuary, northern Taiwan, April 1992 to November 1993.
F
requency (N
o
. of fish)
(DF) and anal fin (AF) was apparently different among the 5 species. They did not overlap in Thryssa dussumieri (Fig. 3a), overlapped 4 ver-tebrae in Stolephorus insularis (Fig. 3b), over-lapped approximately 1-2 vertebrae in Encrasicholina punctifer (Fig. 3c), overlapped approximately 2-4 vertebrae in E. heteroloba (Fig. 3d), and approximately 1-3 vertebrae in Engraulis japonica but they seperated after grow th (Fig. 3e). After staining, the melanophores became brown in color, which appeared in the head (MH), isthmus (MI), anterior part of gut (MFG), posterior of gut (MPG), belly (MB), anal fin base (MAFB), tail (MT), and caudal fin (MC). The melanophore patterns, meristic counts, and relative growth of body parts in proportion to standard length were different among the 5 species.
Morphometric characters
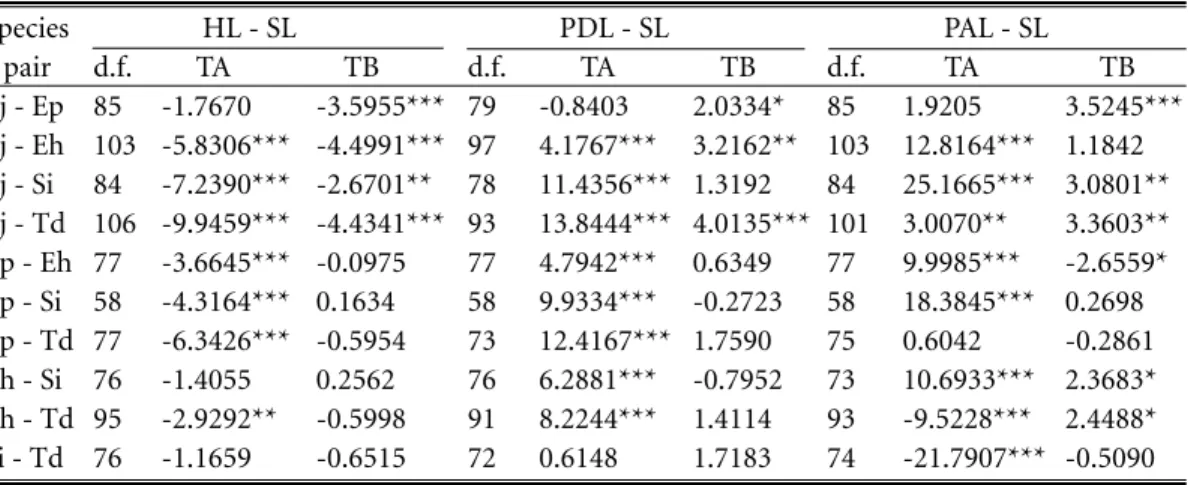
The regressions of head length (HL), pre-dorsal length (PDL), and preanal length (PAL) on standard length (SL) were significant for all 5 species of engraulid larvae (R2= 0.75 ~ 0.98).
Slopes and intercept of the regression lines of
the 5 species were calculated respectively (Table 2). ANCOVA analysis indicated that slopes and adjusted means of the regressions of HL, PDL, and PAL on SL were significantly different among the 5 species (Table 3).
Meristic characters
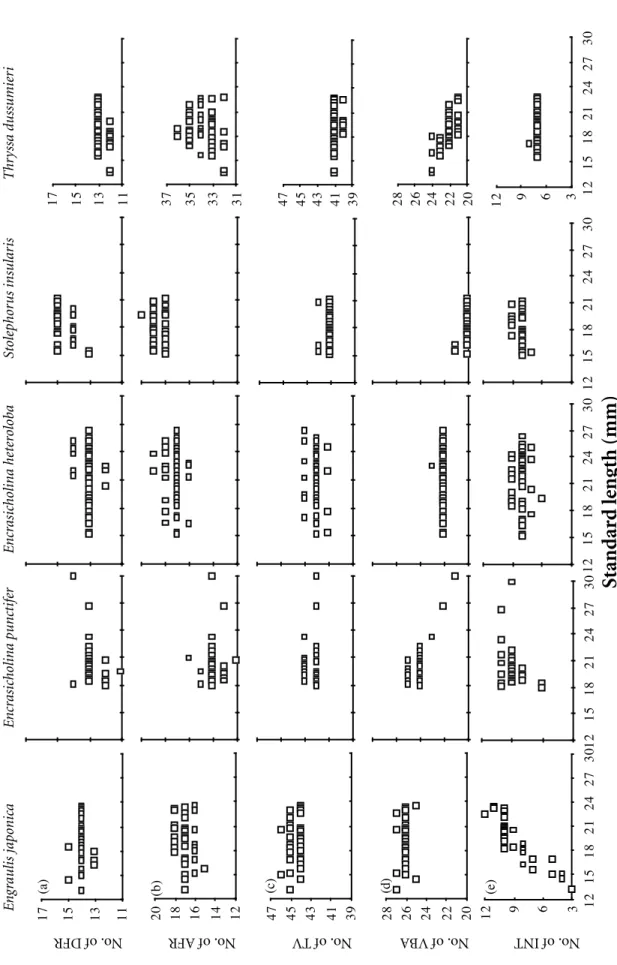
The 5 meristic characters of engraulid lar-vae, dorsal fin rays (DFR), anal fin rays (AFR), total vertebrae (TV), vertebrae before anus (VBA), and interneurals (INT), did not change with standard length beyond 12 mm SL, except the number of INT of E. japonica which increased with standard length but also became constant at the length larger than 18 mm SL (Fig. 4). This indicates that meristic characters were already stable in this study, except for INT of E. japonica. The difference of mean meristic counts among the 5 species were determined by Scheffe’s multiple comparison (Table 4). The number of DFR, T. dussumieri (Td) < E. heteroloba (Eh) = E. punctifer (Ep) < E. japoni-ca (Ej) = S. insularis (Si) (Table 4a); AFR, Ep < Ej = Eh < Si < Td (Table 4b); INT, Td < Eh = Si < Ep < Ej (Table 4c); TV, Td = Si < Eh < Ep Table 2. Regressions of head length (HL), predorsal fin length (PDL), and preanal length (PAL) on standard length (SL) of the 5 species of engraulid larvae. R2, square of correlation coefficient.
Species Sample size Regression R2
Engraulis japonica 57 HL = 0.0677 + 0.1904 SL 0.93 51 PDL = 2.3869 + 0.4593 SL 0.84 57 PAL = 1.6556 + 0.6165 SL 0.96 Encrasicholina punctifer 31 HL = -1.2268 + 0.2623 SL 0.93 31 PDL = 3.7925 + 0.3893 SL 0.91 31 PAL = 3.4004 + 0.5210 SL 0.98 Encrasicholina heteroloba 49 HL = -1.0977 + 0.2642 SL 0.93 49 PDL = 3.8225 + 0.3683 SL 0.89 49 PAL = 1.2696 + 0.5901 SL 0.96 Stolephorus insularis 30 HL= -0.8347 + 0.2579 SL 0.96 30 PDL = 2.4762 + 0.4020 SL 0.86 30 PAL = 1.8842 + 0.5010 SL 0.92 Thryssa dussumieri 49 HL = -1.1106 + 0.2752 SL 0.74 45 PDL = 3.9239 + 0.3221 SL 0.88 47 PAL = 3.1260 + 0.5293 SL 0.94
11 13 15 17 (a) 11 13 15 17 12 14 16 18 20 (b) 31 33 35 37 39 41 43 45 47 (c) 39 41 43 45 47 20 22 24 26 28 (d) 20 22 24 26 28 3 6 9 12 12 15 18 21 24 27 30 (e) 12 15 18 21 24 27 30 12 15 18 21 24 27 30 12 15 18 21 24 27 30 3 6 9 12 12 15 18 21 24 27 30 E n gr aulis j aponica E n cr asic holina punct ifer E nc rasicholina he te ro loba St ole phor us insular is Thr yssa dussumie ri No. of INT No. of VB A No. of TV No. of AFR No. of DFR F igur e 4 . C
ounts of the 5 mer
istic c har act ers vs. standar d length of the 5 sp ecies of eng raulid lar vae c o llect ed in c o astal wat ers off the T ansh ui Ri ve r Estuar y:
(a) dorsal fin r
ays, (b) anal fin r
ays, (c) v er tebr ae, (d) v er tebr ae befor e an
us, and (e) int
er ne ur als.
Standar
d length (mm)
< Ej (Table 4d); and VBA, Si < Td < Eh < Ep < Ej (Table 4e). Apparently, the number of VBA was most reliable to discriminate the 5 engraulid species.
Patterns of pigmentations
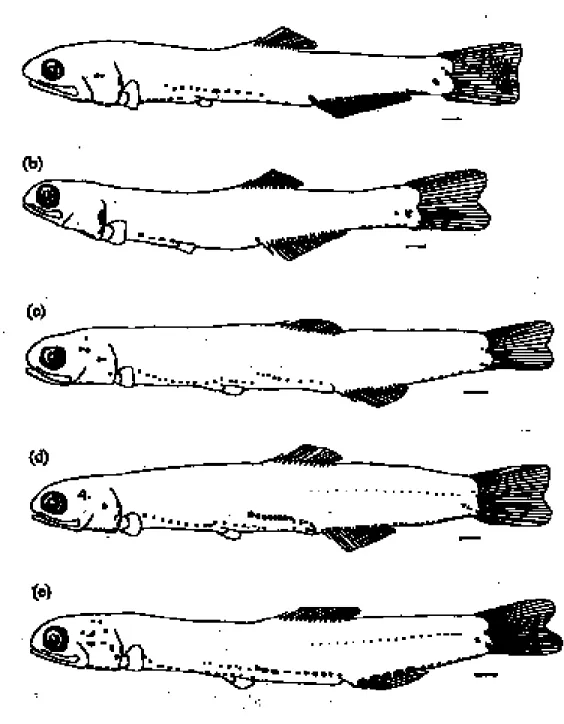
The pigment patterns of the engraulid lar-vae when viewed laterally were classified into 3 types (Fig. 5). Type I was for T. dussumieri, with several rows of melanophores on the top of the auditory vesicle and inner part of the opercles (MH), in the isthmus (MI), both dor-solateral side of fore- and post-gut (MFG and MPG), belly (MB), ventral part of the tail extending from the origin of anal fin to caudal fin bases (MAFB and MT), and lower part of the caudal fin extending to the peduncle (MC; Fig. 5a). Type II for S. insularis was similar to type I, except with fewer melanophores, MPG and MB absent, and MC not extending from the caudal fin to the peduncle (Fig. 5b). Type III included 3 species, E. punctifer, E. heteroloba, and E. japonica and was also similar to type I, but lacked MB (Fig. 5c-e). When the larvae of type III were greater than 20 mm TL, an additional row of melanophores appeared in the dorsal part of the gut and in the
mid-lat-eral part of the posterior half of the body, respectively. This means that the pigment pat-terns shown on the lateral side of the body can not easily distinguish E. punctifer, E. heteroloba, and E. japonica.
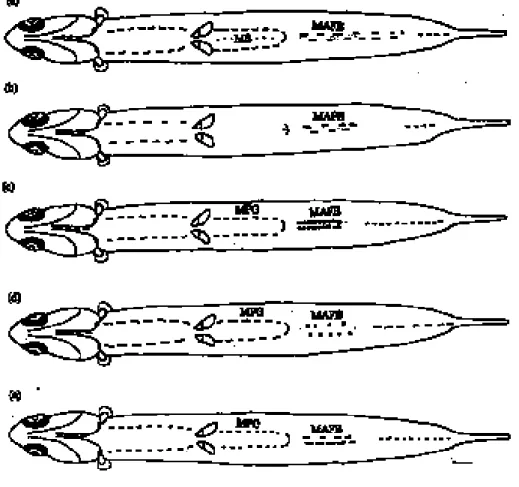
On the other hand, pigment patterns on the ventral side of the body were completely differ-ent among the 5 species and this enables them to be distinguished from one another. T. dus-sumieri presents a row of melanophores on the belly (MB), but the other 4 species lack MB (Fig. 6a). S. insularis lacks MB and melanophores in both dorsolateral sides of post-gut (MPG) (Fig. 6b). The melanophores in the anal fin base (MAFB) are symmetrical in E. punctifer (Fig. 6c), but asymmetrical in both E. heteroloba and E. japonica. The MAFB in the former of the 2 species is spot shaped (Fig 6d) and in the latter species is a bar-like shape (Fig. 6e). This indicates that pigment patterns appearing on the ventral side are more reliable and can be used as a distinct character to dis-criminate theses 5 larval engraulid species. A practical key for quick identification of these 5 larvae is presented below:
Table 3. ANCOVA of the regression of head length (HL), predorsal fin length (PDL), and preanal lengh (PAL) on standard length (SL) among 5 species of engraulid larvae. (d.f.: degree of freedom; TA, TB: t-values for testing the difference in intercepts and slopes between regression lines; Significance level, *: p < 0.05, **: p < 0.01, ***: p < 0.001). Species HL - SL PDL - SL PAL - SL pair d.f. TA TB d.f. TA TB d.f. TA TB Ej - Ep 85 -1.7670 -3.5955*** 79 -0.8403 2.0334* 85 1.9205 3.5245*** Ej - Eh 103 -5.8306*** -4.4991*** 97 4.1767*** 3.2162** 103 12.8164*** 1.1842 Ej - Si 84 -7.2390*** -2.6701** 78 11.4356*** 1.3192 84 25.1665*** 3.0801** Ej - Td 106 -9.9459*** -4.4341*** 93 13.8444*** 4.0135*** 101 3.0070** 3.3603** Ep - Eh 77 -3.6645*** -0.0975 77 4.7942*** 0.6349 77 9.9985*** -2.6559* Ep - Si 58 -4.3164*** 0.1634 58 9.9334*** -0.2723 58 18.3845*** 0.2698 Ep - Td 77 -6.3426*** -0.5954 73 12.4167*** 1.7590 75 0.6042 -0.2861 Eh - Si 76 -1.4055 0.2562 76 6.2881*** -0.7952 73 10.6933*** 2.3683* Eh - Td 95 -2.9292** -0.5998 91 8.2244*** 1.4114 93 -9.5228*** 2.4488* Si - Td 76 -1.1659 -0.6515 72 0.6148 1.7183 74 -21.7907*** -0.5090 Ej: Engraulis japonica Ep: Encrasicholina punctifer Eh: Encrasicholina heteroloba Si: Stolephorus insularis Td: Thryssa dussumieri
Identification of Engraulid Larvae
Table 4. Homogeneity test for the 5 meristic characters (a ~ e) of the 5 species of engraulid larvae collected in coastal waters off the Tanshui River Estuary. Species sharing the same letter are in the same homogeneous group.
(a)
Species Sample No. of dorsal fin rays Homogeneous
size Range Mean ± S.D. group
Thryssa dussumieri 54 12 ~ 13 12.7 ± 0.1 a Encrasicholina heteroloba 90 12 ~ 14 13.1 ± 0.1 b Encrasicholina punctifer 167 11 ~ 15 13.2 ± 0.0 b Engraulis japonica 138 13 ~ 16 14.2 ± 0.0 c Stolephorus insularis 30 13 ~ 15 14.5 ± 0.1 c (b)
Species Sample No. of anal fin rays Homogeneous
size Range Mean ± S.D. group
Encrasicholina punctifer 167 12 ~ 17 14.2 ± 0.1 a Engraulis japonica 138 15 ~ 19 17.0 ± 0.1 b Encrasicholina heteroloba 90 15 ~ 20 17.2 ± 0.1 b Stolephorus insularis 30 18 ~ 20 18.5 ± 0.1 c Thryssa dussumieri 54 32 ~ 36 34.0 ± 0.1 d (c)
Species Sample No. of interneurals Homogeneous
size Range Mean ± S.D. group
Thryssa dussumieri 54 7 ~ 8 7.0 ± 0.1 a Encrasicholina heteroloba 90 4 ~ 9 7.8 ± 0.2 b Stolephorus insularis 30 7 ~ 9 8.2 ± 0.1 b Encrasicholina punctifer 167 6 ~ 10 9.0 ± 0.1 c Engraulis japonica 138 3 ~ 12 9.7 ± 0.0 d (d)
Species Sample No. of vertebrae Homogeneous
size Range Mean ± S.D. group
Thryssa dussumieri 54 40 ~ 41 40.9 ± 0.0 a Stolephorus insularis 30 41 ~ 42 41.1 ± 0.0 a Encrasicholina heteroloba 90 41 ~ 44 42.0 ± 0.1 b Encrasicholina punctifer 167 41 ~ 44 42.3 ± 0.0 c Engraulis japonica 138 43 ~ 46 44.9 ± 0.0 d (e)
Species Sample No. of vertebrae before anus Homogeneous
size Range Mean ± S.D. group
Stolephorus insularis 30 20 ~ 21 20.2 ± 0.1 a
Thryssa dussumieri 54 21 ~ 24 22.0 ± 0.1 b
Encrasicholina heteroloba 90 22 ~ 25 22.4 ± 0.1 c
Encrasicholina punctifer 167 21 ~ 27 24.0 ± 0.1 d
Figure 5. Schemata of the lateral-view pigment patterns of the 5 species of engraulid larvae: (a) Thryssa dussumieri, (b) Stolephorus insularis, (c) Encrasicholina punctifer, (d) E. heteroloba, and (e) Engraulis japonica. Scale bar = 1 mm.
Identification of Engraulid Larvae
Figure 6. Schemata of the ventral-view pigment patterns of the 5 species of engraulid larvae: (a) Thryssa dussumieri, (b) Stolephorus insularis, (c) Encrasicholina punctifer, (d) E. heteroloba, and (e) Engraulis japonica. Scale bar = 1 mm.
DISCUSSION
Five species of larval engraulids, Engraulis japonica, Encrasicholina punctifer, E. heteroloba, Stolephorus insularis, and Thryssa dussumieri, were found in coastal waters off the Tanshui River Estuary, northern Taiwan. The mean meristic counts are significantly different among the 5 species (Table 2), indicating that these species can be discriminated by meristic characters. Similar results were also reported by McGowan and Berry (1984), Chen (1987), Yu and Chiu (1994), and Young et al. (1995). However, we found that meristic counts largely overlapped among the 5 species of engraulid lar vae as did results by Shen (1959). Meanwhile, meristic counts are unstable dur-ing the early developmental stage, e.g., interneurals of E. japonica (Fig. 4). Accordingly, meristic counts alone can not be used to clarify the species status of engraulid larvae. In addition, internal meristic charac-ters, such as vertebrae, can not be seen without staining. Numbers of vertebrae before the anus were different among the 5 species.
The slope and adjusted mean of the linear regressions of morphometric characters rela-tive to standard length were significantly differ-ent among the 5 species of engraulid larvae
(Table 4). This indicates that the species can be
discriminated by morphometric characters. However, variation in morphometric
parame-ters in larval fishes is high because of ontoge-netic changes in body shape, and damage or distortions caused during collection, and shrinkage (Leis and Rennis, 1983). A signifi-cant shift of dorsal and anal fins in the engraulid larvae was reported, and this may lead to the inconsistency in body proportions during different development stages. This phe-nomenon was named “iwashi type metamor-phosis” (Okiyama, 1979b). Thus, morphomet-rics is a conditional character in the discrimi-nation of engraulid larvae.
Pigment patterns on the ventral side of the body are quite different among the 5 species of engraulid larvae (Fig. 3). Pigments on fish body constitute a more versatile taxonomic character than meristics because pigmentation can be used over a greater range in larval size and can be easily recognized (Okiyama, 1979a; Kendall et al., 1984). Several successful applica-tions have been reported on larval identifica-tion with pigmentaidentifica-tion, e.g., notacanthiforms and anguilliforms (Smith, 1979; Castle, 1984), clupeids (Ditty et al., 1994), melanostomiids, gonostomatids, synodontids, paralepidids (Ozawa, 1986), exocoetids (Chen, 1987), tunas (Ueyanagi, 1969), and istiophorids (Ueyanagi, 1974). However, due to the greater variation in pigmentation during metamorphosis in apogonids and gobiids, or pigments easily lost during fixation, such as in labrids and bothids (Leis and Rennis, 1983; Okiyama, 1988; Leis Key to species of engraulid larvae
1a With melanophores on the belly (MB)
D. 12-13, A. 32-36, Vertebrae 40-41 ...Thryssa dussumieri 1b Without melanophores on the belly ...2 2a Without melanophores on both dorsolateral sides of post gut (MPG)
D. 13-15, A. 18-20, Vertebrae 41-42 ...Stolephorus insularis 2b With melanophores on both dorsolateral sides of post gut...3 3a Melanophores on both sides of anal fin base (MAFB) are symmetrical
D. 11-15, A. 12-17, Vertebrae 41-44...Encrasicholina punctifer 3b Melanophores on both sides of anal fin base asymmetrical ...4 4a Melanophores on both sides of anal fin base small spots
D. 12-14, A. 15-20, Vertebrae 41-44 ...Encrasicholina heteroloba 4b Melanophores on both sides of anal fin base a bar-like shape
and Trnski, 1989), the uses of pigmentation need to be confirmed before being applied.
In conclusion, the 5 species of engraulid lar-vae in coastal waters off the Tanshui River Estuary can be quickly identified by their ven-tral-view pigment patterns.
ACKNOWLEDGMENTS
This study was financially supported by the National Science Council of the Republic of China (NSC80-0421-B002-05z & NSC85-2311-B002-032). We are grateful to Mr. F.L Chen and Mr. Y.C. Chen for their help in col-lecting samples, and Dr. S.C. Lee and 2 anony-mous reviewers for helpful comments to the manuscript.
REFERENCES
Castle, P. H. J. (1984) Notacanthiformes and anguilliformes: development. In Ontogeny and systematics of fishes. H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, Jr. and S. L. Richardson, eds. Spec. Publ. No. 1, Amer. Soc. Ichthyol. Herpetol. pp. 62-93.
Chen, C. C. and C. C. Huang (1985) Reports on fish larvae research. COA Fish. Ser. 2, 279 pp. (In Chinese)
Chen, C. H. (1987) Studies of the early life his-tory of flying fishes (family Exocoetidae) in the northwestern Pacific. Taiwan Mus. Spec. Publ. Ser. 7, 203 pp. (in Chinese)
Chen, C. S. (1985) Occurrence of fish larvae in the Kau-Ping River Estuary. J. Fish. Soc. Taiwan 12: 1-20. (In Chinese)
Chen, C. S. (1986) Occurrence of fish larvae in the estuary of Jwo-Shoei River. J. Fish. Soc. Taiwan 13: 11-20. (In Chinese)
Chen, J. T. F. and M. J. Yu (1986) A synopsis of the vertebrates of Taiwan. Taiwan Commercial Book Co., Taipei, pp. 267-277. (In Chinese)
Cheng, T. S. (1980) Study and investigation of bull-ard and anchovy fisheries in coastal
waters of Taiwan. Bull. Taiwan Fish. Res. Inst. 32: 219-233. (In Chinese)
Chen, T. S. (1984) The suitable fishing season of larval anchovy in Fang-Liao and Lin-Yuan by studying on maturity and spawning of anchovy Stolephorus zollingeri (Bleeker). Bull. Taiwan Fish. Res. Inst. 37: 59-66. (In Chinese)
Chen, T. S. (1987) Identification of Engraulid larval fish. Bull. Taiwan Fish. Res. Inst. 42: 77-89. (In Chinese)
Chern, Y. T. and W. N. Tzeng (1993) Feeding strategy of Encrasicholina punctifer and Stolephorus insularis larvae in the estuary of Tanshui River, Taiwan -I. Ontogenetic dietary shifts and morphological correla-tion. J. Fish. Soc. Taiwan 20: 313-328. (In Chinese)
Chern, Y. T. and W. N. Tzeng (1994) Feeding strategy of two lar val anchovies, Encrasicholina punctifer and Stolephorus insularis, in the Tanshui River estuary, Taiwan -II. Prey selectivity and interspecific feeding competition. J. Fish. Soc. Taiwan 21: 33-48. (In Chinese)
Ditty, J. G., E. D. Houde and R. F. Shaw (1994) Egg and larval development of Spanish sar-dine, Sardinella aurita (Family Clupeidae), with a synopsis of characters to identify clu-peid larvae from the northern Gulf of Mexico. Bull. Mar. Sci. 54: 367-380.
Huang, C. C., W. N. Tzeng and S. C. Lee (1985) Preliminary survey on larval fishes of Yen-Liao Bay, northeastern Taiwan. Bull. Inst. Zool., Academia Sinica 24: 147-154.
Huang, J. B. and T. S. Chiu (1996) Daily growth and length distribution of larval Japanese anchovy (Engraulis japonica) in the neritic waters off northeastern Taiwan. Acta Zool. Taiwanica 7: 29-41.
Kendall, A. W. Jr., E. H. Ahlstrom and H. G. Moser (1984) Early life history stages of fishes and their characters. In Ontogeny and Identification of Engraulid Larvae
systematics of fishes. H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, Jr. and S. L. Richardson, eds. Spec. Publ. No. 1, Amer. Soc. Ichthyol. Herpetol. pp. 11-22.
Lee, M. A., K. T. Lee and H. C. Ou (1990) The larval anchovy fishing ground formation in relation to osmotic pressure changes of the coastal waters along southern Taiwan. J. Fish. Soc. Taiwan 17: 233-245
Lee, M. A., K. T. Lee and G. Y. Shiah (1995) Environmental factors associated with the formation of larval anchovy fishing grounds in the coastal waters of southwest Taiwan. Mar. Biol. 121: 621-625
Lee, M. A., S. C. Chou, K. T. Lee and P. C. Lee (1996) Distribution of fish larvae in relation to pycnocline in the coastal waters of Tan-Shui, Taiwan. J. Fish. Soc. Taiwan 23: 195-206
Leis, J. M. and D. S. Rennis (1983) The larvae of Indo-Pacific coral reef fishes. New South Wales Univ. Press, Kensington, 269 pp. Leis, J. M. and T. Trnski (1989) The larvae of
Indo-Pacific shorefishes. Univ. Hawaii Press, Honolulu, 371 pp.
Liu, F. H. and S. C. Shen (1957) A preliminary report on the activity of wen-fishes (her-ring-like fishes) along the coast of Taiwan. Rep. Inst. Fish. Biol., Natl. Taiwan Univ. 1: 24-38. (In Chinese)
McGowan, M. F. and F. H. Berr y (1984) Clupeiformes: development and relation-ships. In Ontogeny and systematics of fishes. H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, Jr. and S. L. Richardson, eds. Spec. Publ. No. 1, Amer. Soc. Ichthyol. Herpetol. pp. 108-126. Nakabo, T. (1993) Fishes of Japan with
pictori-al keys to the species. Tokai Univ. Press, Tokyo, 1474 pp. (In Japanese)
Okiyama, M. (1979a) Manuals for the larval fish taxonomy. (2) General aspects of larval characters that aid identification. Kaiyo to Seibutsu (Ocean and its life) 1(2): 53-59. (In Japanese)
Okiyama, M. (1979b) Manuals for the larval fish taxonomy. (3) Metamorphosis of the clupeoid fishes and problems of larval con-vergence. Kaiyo to Seibutsu (Ocean and its life) 1(3): 61-66. (In Japanese)
Okiyama, M. (ed) (1988) An atlas of the early stage fishes in Japan. Tokai Univ Press, Tokyo, 1154 pp. (In Japanese)
Ozawa, T. (1986) Studies on the oceanic ichthyoplankton in the western North Pacific. Kyushu Univ. Press, Fukuoka, 430 pp.
Potthoff, T. (1984) Clearing and staining tech-niques. In Ontogeny and systematics of fishes. H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, Jr. and S. L. Richardson, eds. Spec. Publ. No. 1, Amer. Soc. Ichthyol. Herpetol. pp. 35-37.
Powles, H. and D. F. Markle (1984) Identification of larvae. In Ontogeny and systematics of fishes. H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, Jr. and S. L. Richardson, eds. Spec. Publ. No. 1, Amer. Soc. Ichthyol. Herpetol. pp. 31-33.
Shen, S. S. (1959) Anchovies found in Taiwan. Rep. Inst. Fish. Biol., Natl. Taiwan Univ. 1: 24-33.
Shen, S. S. (1971) Preliminary report on the study of wen-yü resources of Taiwan. ACTA Oceanographica Taiwanica 1: 101-126. Shen, S. C. (1984) Synopsis of fishes of Taiwan.
Southern Materials Center Inc., Taipei, pp. 96-101.
Shen, S. C., S. C. Lee, K. T. Shao, H. K. Mok, C. T. Chen and C. H. Chen (1993) Fishes of
Taiwan. Dept. Zool., Natl. Taiwan Univ. 960 pp.
Smith, D. G. (1979) Guide to the leptocephali (Elopiformes, Anguilliformes and Notacanthiformes). NOAA Tech. Rep. NMFS Circ. 424.
Steel, R. G. D. and J. H. Torrie (1980) Principles and procedures of statistics. McGraw-Hill, Aukland, 633 pp.
Tsai, C. F., P. Y. Chen, M. A. Lee, K. Y. Hsia and K. T. Lee (1996) Effects of fishing effort on stock size and catch of larval anchovy in coastal waters of southwestern Taiwan. Fish. Res. 28: 71-83.
Tzeng, W. N. and Y. T. Wang (1986) Seasonal occurrence, abundance and diversity of lar-val fishes in the Shuang-hsi River estuary, northeastern Taiwan. Na. Sci. Council Monograph Ser. 14: 135-154. (In Chinese) Tzeng, W. N. and Y. T. Wang (1992) Structure,
composition and seasonal dynamics of the larval and juvenile fish community in the mangrove estuary of Tanshui River, Taiwan. Mar. Biol. 113: 481-490.
Tzeng, W. N. and Y. T. Wang (1993) Hydrography and distribution dynamics of larval and juvenile fishes in the coastal waters of Tanshui River estuary, Taiwan, with reference to estuarine larval transport. Mar. Biol. 116: 205-217.
Tzeng, W. N. and Y. T. Wang (1997) Movement of fish larvae with tidal flux in the Tanshui River Estuar y, northern Taiwan. Zool. Studies 36: 179-185
Ueyanagi, S. (1969) Observations on the distri-bution of tuna larvae in the Indo-Pacific Ocean with emphasis on the delineation of the spawning areas of albacore, Thunnus alalunga. Bull. Far Seas Fish. Res. Lab. 2: 177-256. (in Japanese)
Ueyanagi, S. (1974) On an additional diagnos-tic character for the identification of billfish
larvae with some notes on the variations in pigmentation. NOAA Tech. Rep. NMFS Circ. 675: 73-78.
Wang, S. B. and P. P. Hwang (1992) Occurrence of fish larvae and juveniles in Tan-Shui estuary, northern Taiwan. J. Fish. Soc. Taiwan 19: 173-182.
Wang Y. T. (1987) Studies on the eggs, larvae and juveniles of fishes in the estuaries of Tansui and Shuang-hsi Rivers, northern Taiwan. MSc Thesis, Inst. Mar. Sci., Chinese Culture Univ., 306 pp. (In Chinese)
Wang, Y. T., W. N. Tzeng and S. C. Lee (1991) A preliminary study on species composition and seasonal abundance of fish eggs and lar-vae from the coastal waters adjacent to the Tansui River estuary, Taiwan (1984-1985). J. Fish. Soc. Taiwan 18: 7-20.
Wang, Y. T. and W. N. Tzeng (1997) Temporal succession and spatial segregation of clu-peoid larvae in the coastal waters off the Tanshui River Estuary, northern Taiwan. Mar. Biol. 129: 23-32.
Whitehead, P. J. P., G. J. Nelson and T. Wongratana (1988) FAO species catalogue, Vol. 7 Clupeoid fishes of the world (subor-der Clupeoidei), Part 2 - Engraulididae. FAO, Rome, 579 pp.
Young, S. S., C. C. Chen and T. S. Chiu (1992) Resource characteristics of young herring-like fish in the I-Lan Bay area - Fishing sea-son, major species and size variation. J. Fish. Soc. Taiwan 19: 273-281.
Young, S. S., T. S. Chiu and S. C. Shen (1994) A revision of the family Engraulidae (Pisces) from Taiwan. Zool. Studies 33: 217-227. Young, S. S., T. S. Chiu and S. S. Shen (1995)
Taxonomic description and distribution of larval anchovy (Engraulidae) occurred in the waters around Taiwan. Acta Zoologica Taiwanica 6: 33-60.
Yu, P. T. and T. S. Chiu (1994) Fishery target species of larval anchovy fishery in the west-ern central Taiwan. J. Fish. Soc. Taiwan 21(3): 227-239.