Molecular Typing and Virulence Analysis among Serotype K1 Klebsiella pneumoniae Isolated from Liver Abscess Patients and Stool Carriage from
Non-Infectious Subjects in Hong Kong, Singapore and Taiwan
L. Kristopher Siu1,2, Chang-Phone Fung3, Feng-Yee Chang,4, Nelson Lee5, Kuo-Ming
Yeh4, Tse-Hsien Koh6*, Margaret Ip7*
Division of Infectious Diseases, National Health Research Institutes1; Graduate Institute of Basic Medical Science, China Medical University, Taichung2; Section of
Infectious Diseases, Department of Medicine, Taipei Veterans General Hospital and National Yang-Ming University3, Taipei; Division of Infectious Diseases and Tropical
Medicine, Department of Internal Medicine, Tri-Service General Hospital, Taipei4, Taiwan; Department of Pathology, Singapore General Hospital6, Singapore; Department of Medicine and Therapeutics5, and Department of Microbiology7, Faculty of Medicine, Chinese University of Hong Kong, Hong Kong SAR, People’s
Republic of China
*Corresponding authors:
T.-H. Koh: Department of Pathology, Singapore General Hospital, Singapore. Phone: 65-98259566. Fax: 65-62226826. E-mail: [email protected].
M-Ip: Department of Microbiology, Faculty of Medicine, Chinese University of Hong Kong, Hong Kong SAR, P.R.C. Phone: 852-26321265 Fax: 852-26473227. E-mail: [email protected]
Key words: Liver abscess; Klebsiella pneumoniae; MLST Running Title: Klebsiella pneumoniae Serotype K1 in Asia
Serotype K1 Klebsiella pneumoniae with Multi-Locus Sequence Type, ST23, has been strongly associated with liver abscess in Taiwan. Little data is available regarding strain types and virulence of this serotype from other Asian countries. Serotype K1 K. pneumoniae strains, isolated from liver abscess and stool from subjects hospitalized in Hong Kong, Singapore and Taiwan hospitals were examined. Twenty-eight and 21 serotype K1 isolates were identified respectively from liver abscess and stool carriage. Seven sequence types were identified; 85.1% (40 of 47) belonged to ST23, 1 isolate belonged to ST163 (a single locus variant of ST23), and 2 isolates were ST249 (a 3-locus variant of ST23). New STs, namely ST367, 425, and 426 were allocated to 3 of 4 isolates from stool carriage. The virulence of these strains was determined by neutrophil phagocytosis and mice infection models. Except for two ST23 isolates, all KPs were resistant to phagocytosis. Resistance to serum killing varied in isolates of ST23 while non-ST23 strains were all susceptible to serum killing except one with ST249 from liver abscess. Hyper-virulence isolates, LD50<102 cfu,
were all from ST23 and were resistant to phagocytosis, serum killing and also carried both virulence associated genes rmpA and aerobactin. MLST genotype 23 in serotype K1 K. pneumoniae was strongly associated with liver abscess in Asia Pacific regions. ST23 is also a prevalent strain type isolated from stool carriage. Serotype K1 K.
pneumoniae with capsule expression leading to phagocytic resistance and the carrying
Introduction
Klebsiella pneumoniae is an opportunistic nosocomial pathogen frequently
encountered in urinary tract infections, pneumonia and septicaemia (17). However, community-acquired K. pneumoniae liver abscess (KP-LA) has been frequently observed in Taiwan in the past two decades and seems to be endemic in this locality (6). Recent epidemiologic surveys have also shown that K. pneumoniae as a single agent, rather than polymicrobial infection, is the most common causative agent for liver abscess in Singapore and Korea (4,22). Individual cases have also been reported from US, Japan, Canada and Thailand (10,11,21).
K. pneumoniae serotype K1 has been identified as the major cause of KP-LA and
bacteraemia. Complications of endophthalmitis and meningitis are often seen in KP-LA especially in patients with diabetes mellitus (DM) (6). DM is prevalent in all three continents while alcoholism, as a predisposing factor, has only been reported in Korea (4). KP-LA isolates have been associated with capsule hypermucoviscosity and the presence of the virulences gene, magA and rmpA. However, it is now clear that
magA isa capsular polymerase, wzyKPK1, specific for serotype K1 capsule formation
(23).
Molecular typing of isolates from KP-LA in Taiwan has shown that a portion of isolates in one report were clustered by pulsed-field gel electrophoresis (PFGE) while non-clonal isolates were also observed in other study (1). Turton et al., has identified the genetic similarity of KP-LA isolates from three continents by multilocus sequence typing (MLST) (20). To our knowledge, no data is available on the molecular types and characteristics of serotype KI isolates from the stool of healthy carriers in the
general population, nor had this been compared to isolates from liver abscess. In the present study, we have performed MLST for serotype K1 isolates from liver abscess as well as isolates from carriers without a history of KP-LA and assessed the virulence of isolates with different MLST types from Hong Kong, Singapore and Taiwan.
MATERIAL AND METHODS Bacterial strains.
K. pneumoniae strains that were isolated from liver abscess and stool from healthy
subjects, hospitalized patients without history of liver abscess or patients admitted with noninfectious diseases were collected at Prince of Wales hospital in Hong Kong, Singapore General Hospital, National University Hospital in Singapore and Tri-Service General Hospital in Taiwan from 2002 to 2009. The diagnosis of liver abscess was confirmed by abdominal ultrasonography and/or computerized tomography. Identification of the isolates was according to standard clinical microbiologic methods.
Serotyping, rmpA and aerobactin gene detection by PCR.
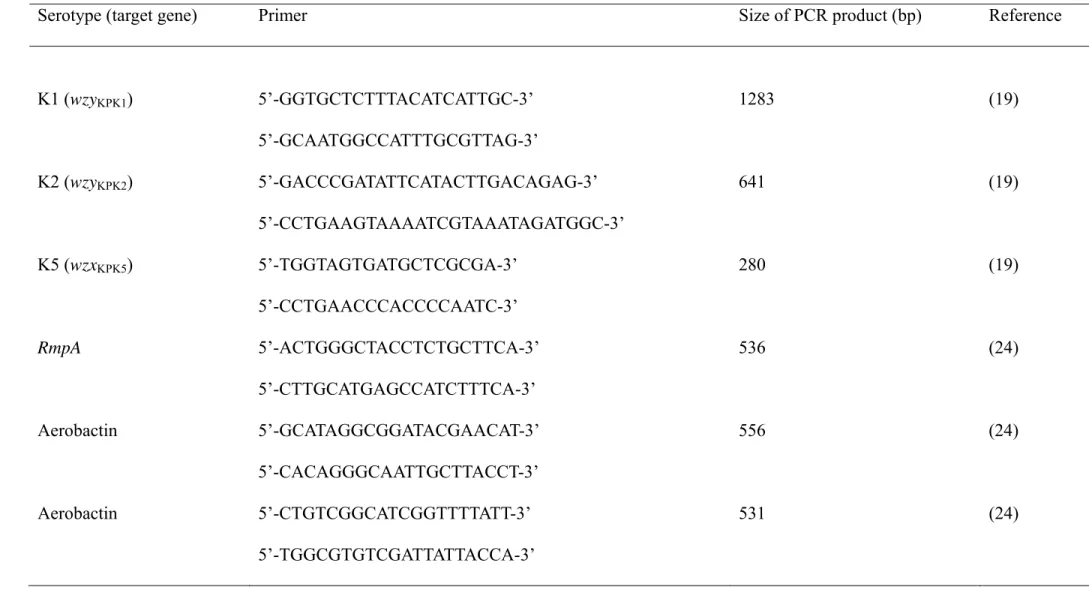
Isolates were serotyped using the capsule swelling reaction with antisera obtained from the Health Protection Agency in UK and by PCR as previously described (6). PCRs to determine the presence of the specific genes for serotype K1, K2 and K5,
rmpA and the aerobactin gene. were performed using primers as listed in Table 1
(19,24). A bacterial colony from an overnight-culture was added to 300l water and boiled for 15 min to release DNA template. The reaction mixture was kept at 95°C for 5 min, followed by 40 temperature cycles of 95°C for1 min, 50°C for 1 min, and 72°C for 2 min, and 72°C for 7 min. The expected PCR products were 1283bp for wzyKPK1,
535bp for rmpA and either 556 or 531bp for aerobactin (Table 1).
Pulsed field gel electrophoresis (PFGE).
Total DNA was prepared and PFGE was performed as described previously (22). The restriction enzyme XbaI (New England Biolabs, Beverly, MA, USA) was used at the manufacturer’s suggested temperature. Restriction fragments were separated by PFGE in 1% agarose gel (Bio-Rad, Hercules, CA, USA) in 0.5×TBE buffer (45 mM Tris, 45 mM boric acid, 1.0 mM EDTA, pH8.0) for 22 h at 200 V at a temperature of 14oC, with ramped times of 2 to 40 s using the Bio-Rad CHEF MAPPER apparatus (Bio-Rad Laboratories, Richmond, CA, USA). Gels were then stained with ethidium bromide and photographed under ultraviolet light. The resulting genomic DNA profiles, or “fingerprints”, were interpreted according to established guidelines (18).
Multilocussequencetyping(MLST)
MLST were performed according to Turton et al., (20). Sequences of seven housekeeping genes were obtained for isolates from liver abscess patients and carriers. Sequence information was compared with that available from the MLST website (http://pubmlst.org/kpneumoniae/) developed by Keith Jolley (8). Alleles and sequence types (STs) were assigned accordingly. Sequences of any alleles that were not in the database were submitted to the curator and a new allele number obtained. A difference in two or more alleles was considered to indicate that the sequence types being compared were unrelated.
Fluorescence labeling of bacteria. Labeling was performed as previously described
overnight at 370C. The concentration was approximated using photospectrometry (Olympus, US). The percentage of bacterial viability in an aliquot of each population was determined by quantitative plate counting. Populations were then heat killed for 60 min in a 700C water bath, and quantitative colony count determination of the population viability was performed again. The bacteria were washed with PBS and labeled with fluorescein isothiocyanate (FITC) by incubation with 0.1 g/mL FITC (Sigma Chemical Co., St. Louis, MO) in 0.10 M NaHCO3, pH 9.0, for 60 min at 25 0C.
Bacteria were washed of unbound fluorochrome with PBS by three cycles of centrifugation (13,000 rpm, 10 min). The FITC-labeled bacteria were resuspended at a concentration of 2×108 cells/mL in PBS, divided into equal volumes, and stored at –700C. Aliquots were thawed just prior to use.
Phagocytosis assay. Phagocytosis was measured using a standard assay. Normal
human serum pooled from healthy volunteers was divided into equal volumes and stored at –700C. Serum was thawed immediately prior to use and stored on ice until added to the phagocytosis assay. Briefly, for the assay, 100 L of a neutrophil suspension (representing 1 ×106 cells), 100L of freshly thawed pooled normal human serum (10% v/v opsonization), and 600 L PBS was added to sealable 10 × 75 mm Falcon™ polypropylene tubes (BD, Franklin Lakes, NJ). The suspension was pre-warmed with shaking for 5 min at 370C. Multiple volumes of 200 μL FITC-labeled bacteria (representing 4×107 colony forming units [cfu]/mL) were added to 800 μL to produce a final volume of 1.0 mL. Each tube was capped and incubated in a shaking water bath at 370C with continuous agitation for 15 min. An unincubated tube served as the 0-min time point. At each designated time, samples were removed and placed in an ice bath. The cells in each suspension were removed
by centrifugation at 250 g for 6 min, and the cell pellet was resuspended in 1.0 mL of ice-cold PBS and maintained at 4 0C. A 600 μL volume of the suspension was transferred into a new tube, and ethidium bromide was added to a final concentration of 50 mg/L before measurement. Excess ethidium bromide was used to suppress the extracellular fluorescence. Bacteria that were not localized in neutrophils appeared red in color upon microscopic examination (see below).
Phagocytosis assay using flow cytometry. A FACScan, emitting an argon laser beam
at 488 nm (Becton Dickinson Immunocytometry Systems, San Jose, Calif.), was used to detect FITC fluorescence. The sideway scatter (SSC) threshold was 52. The detector was set at E00, 350, and 427 for forward scatter (FSC), SSC, and fluorescence 1 (FL1-H, green), respectively. Fluorescence values were collected after gating the detector on the FSC and SSC combination. A total of 10,000 cells were processed using the Cellquest version 1.0 software (Becton Dickinson Immunocytometry Systems). Fluorescence distribution data collected using a logarithmic amplifier was displayed as single histograms for FL1-H. By processing unstained and FITC-stained bacterial phagocytosis mixtures, the boundary of positive and negative fluorescence was determined. The percentage of ingested bacteria was assessed after the addition of ethidium bromide.
Susceptibility to serum killing. Serum bactericidal activity was measured using the
method of Hughes et al. (7) as modified by Podschun et al..(16). Bacteria grown in nutrient broth were collected during the early logarithmic phase. The viable bacterial concentration was adjusted to 1×106 colony forming units/mL. Twenty-five microliters of bacteria were added to 75 µL of pooled human sera contained in a
10×75 mm Falcon polypropylene tube (BD Biosciences, Franklin Lakes, New Jersey). Tubes were agitated for 0, 60, 120, or 180 min. To determine the number of viable bacteria after exposure to serum, an aliquot of each bacterial suspension was removed at the designated time point, diluted 10-fold by addition of Mueller-Hinton broth, plated on Mueller-Hinton agar, and assayed as described immediately below.
Results were expressed as percentage of inoculums, and responses in terms of viable counts were graded from 1–6 as described previously (16). Grade 1 represented viable counts <10% of the inoculum after 1 and 2 h, and <0.1% after 3 h. Grade 2 represented viable counts 10–100% of the inoculum after 1 h and <10% after 3 h. Grade 3 represented viable counts that exceeded those of the inoculum after 1 h, but <100% after 2 and 3 h. Grade 4 represented viable counts >100% of the inoculum after both 1 and 2 h, but <100% after 3 h. Grade 5 represented viable counts >100% of the inoculums 1, 2, and 3 h, but which decreased during the third hour. Finally, grade 6 represented viable counts that exceeded those of the inoculum after 1, 2, and 3 h, and which increased throughout this time period. Each strain was tested at least three times. A strain was considered serum resistant or serum sensitive if the grading was the same in all experiments. Each isolate was classified as highly sensitive (grades 1 or 2), intermediately sensitive (grades 3 or 4), or resistant (grades 5 or 6).
Mice lethality test
In determination of LD50 in mice, six mice were used as a sample population for each
bacterial concentration. Bacterial concentration was calculated by cell forming unit (cfu). Intraperitoneal (i.p.) injection was used to assess virulence. Symptoms and signs of infection were observed for 14 days. Survival of the inoculated mice was recorded and the LD50 was calculated using SigmaPlot version 7.0 from SPSS Inc. (Chicago,
IL).
Results
MLST profiles of isolates and detection for virulence associated rmpA and aerobactin genes from Hong Kong, Singapore and Taiwan
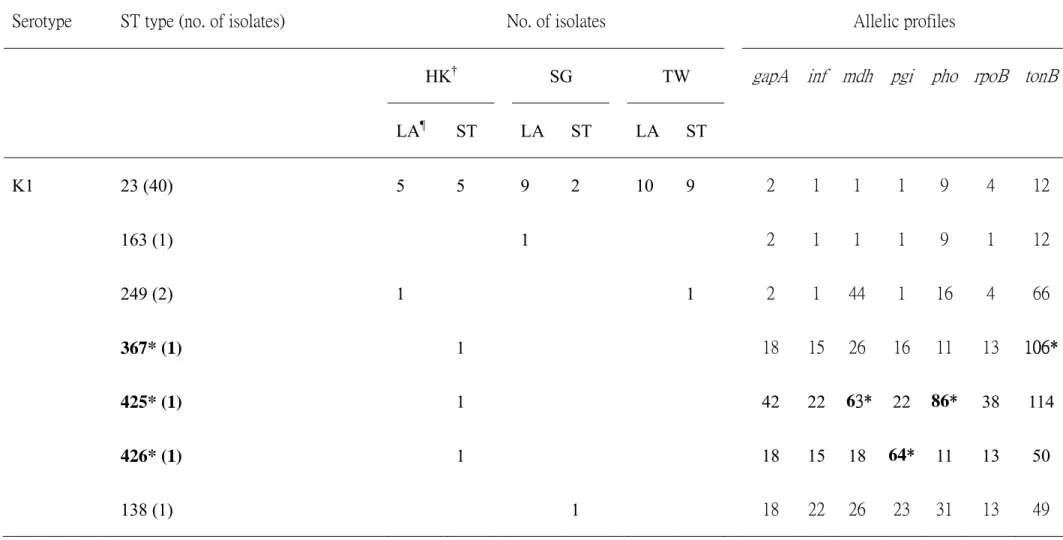
A total of 47 serotype K1 isolates were confirmed by serotyping and PCR and selected for this study. Twenty-six and 21 KPs were isolated from liver abscess patients and stool of uninfected subjects (Table 2). Forty of 47 (85.1%) of these isolates belonged to ST23. One liver abscess isolate had ST163 type. ST23 and ST163 were clonally related, the latter being a single locus variant with allelic difference in the rpoB gene. Two isolates had ST249 including one isolated from liver abscess and another from stool carriage. There was one isolate each belonging to ST 367, 425, 426 and 138 and all these isolates were from stool carriage. ST 367, 425 and 426 were new ST types found in this study. Except ST23 and 163, all other ST types were non-clonally related (Table 2). PCR results revealed that all isolates were carrying
rmpA. In addition, all ST23 and SLV, ST163 were carrying both rmpA and aerobactin.
All the remaining isolates, except 1 strain of ST367, were negative for aerobactin.
Neutrophil phagocytosis and serum resistance of K. pneumonaie with different MLST types.
Overall, there was no statistical difference in resistance to neutrophil phagocytosis among strains of different MLST types in this study. Two ST23 isolates, one from liver abscess and the other from stool carriage were relatively susceptible to neutrophil phagocytosis and these two isolates were both from Hong Kong. The remainder of isolates demonstrated resistance to neutrophil phagocytosis as compared
to the capsule deficient control strain (Figure 1).
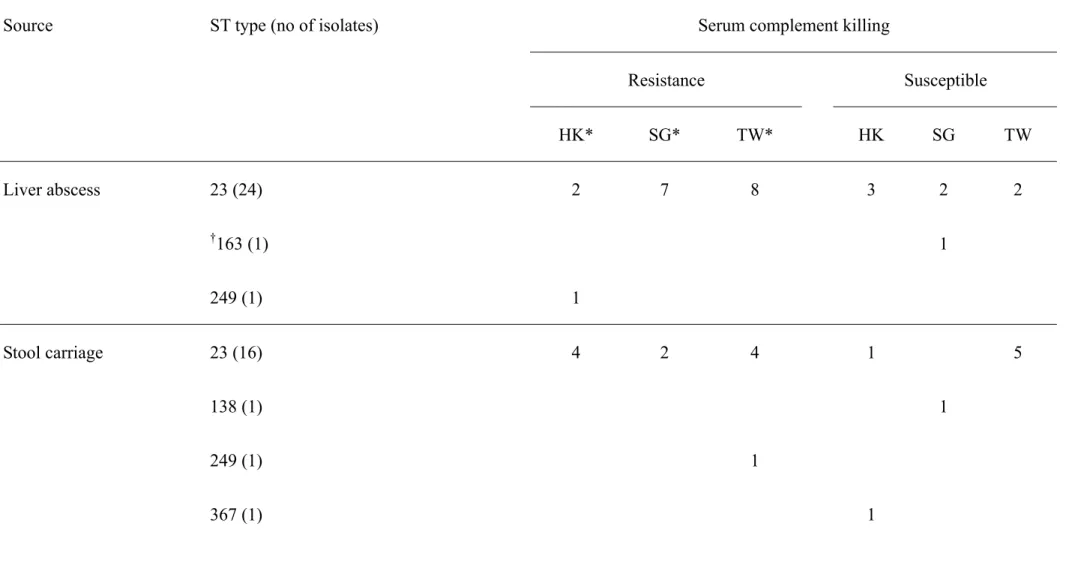
Except for two ST249 isolates, isolates with STs other than 23, were all susceptible to serum complement killing. The majority of ST23 isolates, 10 of 16 from carriage and 17 of 24 from liver abscess, demonstrated intermediate resistance or resistance (of grade 4 or 5) to serum complement killing. Serum killing susceptible strains with ST23 were found in all three geographical regions and these strains were isolated from liver abscess patients and stool carriage from uninfected subjects. Although ST163 was clonally related to ST23, the isolate with ST163 was susceptible to serum complement killing (Table 3).
PFGE for selected serotype K1 isolates with different MLSTs and susceptibility to neutrophil phagocytosis and serum killing
Eight isolates in the ST23 group were further pulse-field typed, according to their phagocytic and serum susceptibilities; (1) susceptible to both phagocytosis and serum complement killing; (2) susceptible to phagocytosis but resistant to serum complement killing; and (3) resistant to both phagocytosis and serum complement killing. In addition, 2 isolates with ST249 were also included. Except for two isolates from group (1) with indistinguishable pulsed field patterns all the other isolates had PFGE profiles that were unrelated.
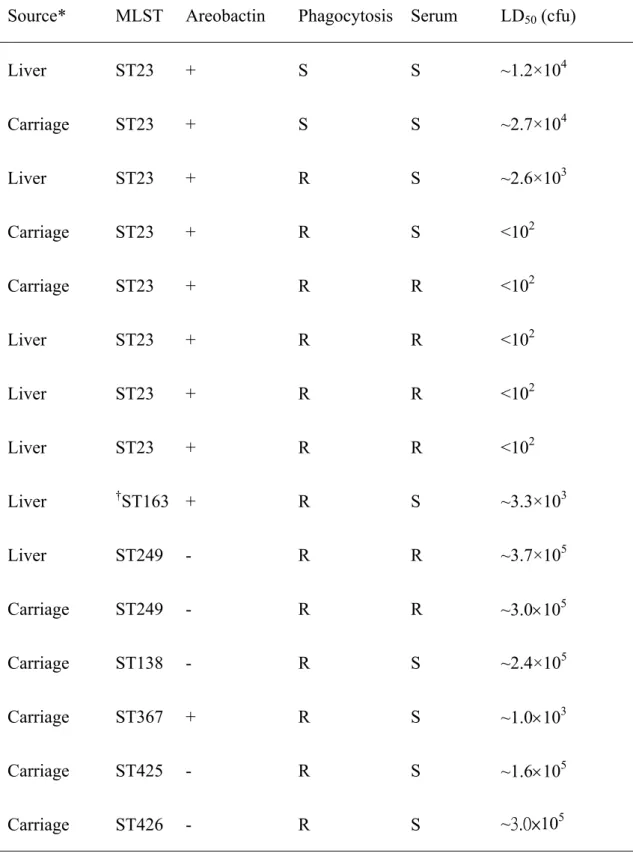
Mice lethality by intra-peritoneal injection of serotype K1 isolates with different MLSTs, susceptibility to neutrolphil phagocytosis and serum killing and virulence associated aerobactin gene
selected for mice lethality studies. Isolates had LD50 that varied from <102 to 3.0105
cfu (Table 4). Hyper-virulent isolates, LD50<102 cfu, were all from ST23 and were
represented by both stool carriage and liver abscess isolates. All these isolates were resistant to phagocytosis and serum killing and also possessed both virulence associated genes of rmpA and aerobactin. Isolates susceptible to both neutrophil phagocytosis and serum killing assays were relatively less virulent, with LD50, ~104.
Although one ST-23 isolate showed susceptibility to serum killing, it was as virulent as ST-23 with both resistance to phagocytosis and serum killing (Table 4). Strains that were negative for the aerobactin gene showed relatively low virulence, LD50, ~105.
Comparing isolates with similar resistance to phagocytosis and serum killing but with or without aerobactin, ST23 and ST249 isolates showed different mice lethality implicating aerobactin as an independent virulence factor. In vitro parameters of neutrophil phagocytosis, serum resistance and the presence of aerobactin and rmpA for virulence assessment were correlated to in vivo mice lethality.
Discussion
Turton et al., were the first group to identify a high prevalence of ST23 in liver abscesses in Taiwan and revealed the clonal relationship of strains in her study using PFGE (20). In our study, we demonstrated that ST23 was also predominant in serotype K1 K. pneumoniae causing liver abscess and carried in stool in uninfected subjects in Hong Kong, Singapore and Taiwan. In contrast to Turton et al.,(20), our ST23 isolates from KP-LA and KP-ST were not related by PFGE indicating KP-LA may not caused by a specific clone. The diversity in our isolates might be explained by the fact that they were isolated from two patient sources (liver abscess and stool) and from three different regions in Asia. In general, non-ST23 isolates were mostly isolated from stool carriage and appeared relatively less virulent, although one case of KP-LA was due to ST249. This might reflect that non-ST23 is relatively uncommon in stool carriage in Asia Pacific regions, or that other host factors might also play a role in KP-LA. Further studies may elucidate the background prevalence of serotype K1 KPs in the Asia Pacific region, and the significance of non-ST23 in KP-LA.
In this study, no significant difference in virulence was identified between ST23 isolated from LA and from stool of carriers. The determinants and virulence of the strains were clearly demonstrated by the presence of the aerobactin gene, and the low LD50 in the mice lethality model. Previous studies had specifically mentioned
hypermucovisity as an in vitro parameter for the virulence phenotype (5,9,13). However, the results obtained from this study showed all serotype K1 isolates were hypermucoviscous but varied in resistance to phagocytosis, serum and mice lethality (Table 4). This result was in-line with the observation from Turton et. al.,(20). Non-hypermucoviscous strains could be as virulent as hypermucoviscous strains.
Since all K1 isolates in this study are carrying the rmpA, we are unable to assess the contribution of this gene in virulence. Previous studies have confirmed that rmpA is regulator of capsule formation (2). Lost of this regulator will down regulate the capsule synthesis leading to the lost of phagocytic resistance and mucoid phenotype. The LD50 of this type of deficiency strain is at 107 cfu implicating the importance of
presence of capsule to virulence (14,22). In our study, the LD50 of selected isolates fell
between <102 all 105 cfu indicating that normal expressed K1 capsule is not a solely factor for hyper-virulence. Our study observed that phagocytic resistance or carriage of the aerobactin gene were two independent determinants contributing to mice lethality. Lack of any one of these factors would reduce at least 100 fold the bacterial concentration for LD50. Hypervirulent strains, LD50 <102 cfu, were all resistant to
phagocytosis and possessed the aerobactin gene. One isolate, ST367, with resistance to phagocytosis and carrying the aerobactin gene had an LD50 of approximately 103
indicating other factor(s) may also play a role in hyper-virulence. Previous studies have also documented other virulence factors including kfu (an iron uptake system) and allS (a gene associated with allantoin metabolism) that may contribute to virulence in KP-LA (3,15). Unfortunately, the previous study also indicated that all serotype K1 isolates contained these two determinants (24). Furthermore, virulence of non-K1 isolates contained these two virulence determinants could be as low as LD50
>107 cfu. Thus, future studies comparing these two strain types, ST367 and ST23 may shed further light into other determinant(s) that may contribute to virulence.
Our present data demonstrated the prevalence of K. pneumoniae serotype K1 ST23 in KP-LA in the Asia region. Outside Asia, KP-LA has been reported more frequently in patients of Asian descent than non-Asians (11,12). Most of these cases had no travel
history before disease occurred. Whether Asians are genetically more susceptible to LA or there are other factors that contribute to the increased prevalence in Asians is yet unknown. Our present data could not answer this question but have given an indication that hyper-virulence may not be the sole factor in serotype K1 leading to disease in the Asia Pacific region. In conclusion, MLST genotype 23 in serotype K1 K.
pneumoniae was strongly associated with high carriage prevalence and liver abscess
in Asia Pacific regions. A few virulence determinants are demonstrated, but other genetic factors of these strains and host factors may play a role in the pathogenesis of LA and remain to be elucidated.
Acknowledgement
This work was supported by grants from the National Science Council and the National Health Research Institutes, Taiwan.
References
1. Cheng, H. P., F. Y. Chang, C. P. Fung, and L. K. Siu. 2002. Klebsiella pneumoniae liver abscess in Taiwan is not caused by a clonal spread strain. J.Microbiol.Immunol.Infect. 35:85-88.
2. Cheng, H. Y., Y. S. Chen, C. Y. Wu, H. Y. Chang, Y. C. Lai, and H. L. Peng. 2010. RmpA regulation of capsular polysaccharide biosynthesis in Klebsiella pneumoniae CG43. J.Bacteriol. 192:3144-3158.
3. Chou, H. C., C. Z. Lee, L. C. Ma, C. T. Fang, S. C. Chang, and J. T. Wang. 2004. Isolation of a chromosomal region of Klebsiella pneumoniae associated with allantoin metabolism and liver infection. Infect.Immun. 72:3783-3792. 4. Chung, D. R., S. S. Lee, H. R. Lee, H. B. Kim, H. J. Choi, J. S. Eom, J. S.
Kim, Y. H. Choi, J. S. Lee, M. H. Chung, Y. S. Kim, H. Lee, M. S. Lee, and C. K. Park. 2007. Emerging invasive liver abscess caused by K1 serotype
Klebsiella pneumoniae in Korea. J.Infect. 54:578-583.
5. Fang, C. T., Y. P. Chuang, C. T. Shun, S. C. Chang, and J. T. Wang. 2004. A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J.Exp.Med. 199:697-705.
6. Fung, C. P., F. Y. Chang, S. C. Lee, B. S. Hu, B. I. Kuo, C. Y. Liu, M. Ho,
and L. K. Siu. 2002. A global emerging disease of Klebsiella pneumoniae liver
abscess: is serotype K1 an important factor for complicated endophthalmitis? Gut 50:420-424.
7. Hughes, C., R. Phillips, and A. P. Roberts. 1982. Serum resistance among Escherichia coli strains causing urinary tract infection in relation to O type and the carriage of hemolysin, colicin, and antibiotic resistance determinants. Infect.Immun. 35:270-275.
8. Jolley, K. A., M. S. Chan, and M. C. Maiden. 2004. mlstdbNet - distributed multi-locus sequence typing (MLST) databases. BMC.Bioinformatics. 5:86. 9. Keynan, Y., J. A. Karlowsky, T. Walus, and E. Rubinstein. 2007. Pyogenic
liver abscess caused by hypermucoviscous Klebsiella pneumoniae. Scand.J.Infect.Dis. 39:828-830.
10. Kohayagawa, Y., K. Nakao, M. Ushita, N. Niino, M. Koshizaki, Y.
Yamamori, Y. Tokuyasu, and H. Fukushima. 2009. Pyogenic liver abscess
caused by Klebsiella pneumoniae genetic serotype K1 in Japan. J.Infect.Chemother. 15:248-251.
11. Lederman, E. R. and N. F. Crum. 2005. Klebsiella liver abscess: a coast-to-coast phenomenon. Clin.Infect.Dis. 41:273.
12. Lederman, E. R. and N. F. Crum. 2005. Pyogenic liver abscess with a focus on Klebsiella pneumoniae as a primary pathogen: an emerging disease with unique clinical characteristics. Am.J.Gastroenterol. 100:322-331.
13. Lee, H. C., Y. C. Chuang, W. L. Yu, N. Y. Lee, C. M. Chang, N. Y. Ko, L. R.
Wang, and W. C. Ko. 2006. Clinical implications of hypermucoviscosity
phenotype in Klebsiella pneumoniae isolates: association with invasive syndrome in patients with community-acquired bacteraemia. J.Intern.Med.
14. Lin, J. C., F. Y. Chang, C. P. Fung, J. Z. Xu, H. P. Cheng, J. J. Wang, L. Y.
Huang, and L. K. Siu. 2004. High prevalence of phagocytic-resistant capsular
serotypes of Klebsiella pneumoniae in liver abscess. Microbes.Infect.
6:1191-1198.
15. Ma, L. C., C. T. Fang, C. Z. Lee, C. T. Shun, and J. T. Wang. 2005. Genomic heterogeneity in Klebsiella pneumoniae strains is associated with primary pyogenic liver abscess and metastatic infection. J.Infect.Dis. 192:117-128.
16. Podschun, R., E. Teske, and U. Ullmann. 1991. Serum resistance properties of Klebsiella pneumoniae and K. oxytoca isolated from different sources. Zentralbl.Hyg.Umweltmed. 192:279-285.
17. Podschun, R. and U. Ullmann. 1998. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin.Microbiol.Rev. 11:589-603.
18. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray, D.
H. Persing, and B. Swaminathan. 1995. Interpreting chromosomal DNA
restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J.Clin.Microbiol. 33:2233-2239.
19. Turton, J. F., H. Baklan, L. K. Siu, M. E. Kaufmann, and T. L. Pitt. 2008. Evaluation of a multiplex PCR for detection of serotypes K1, K2 and K5 in Klebsiella sp. and comparison of isolates within these serotypes. FEMS Microbiol.Lett. 284:247-252.
serotype K1 causing liver abscesses in three continents. J.Med.Microbiol.
56:593-597.
21. Wiwanitkit, V. 2005. Causative agents of liver abscess in HIV-seropositive patients: a 10-year case series in Thai hospitalized patients. Trop.Doct.
35:115-117.
22. Yeh, K. M., A. Kurup, L. K. Siu, Y. L. Koh, C. P. Fung, J. C. Lin, T. L. Chen,
F. Y. Chang, and T. H. Koh. 2007. Capsular serotype K1 or K2, rather than
magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in Singapore and Taiwan. J.Clin.Microbiol. 45:466-471.
23. Yeh, K. M., J. C. Lin, F. Y. Yin, C. P. Fung, H. C. Hung, L. K. Siu, and F. Y.
Chang. 2010. Revisiting the importance of virulence determinant magA and its
surrounding genes in Klebsiella pneumoniae causing pyogenic liver abscesses: exact role in serotype K1 capsule formation. J.Infect.Dis. 201:1259-1267.
24. Yu, W. L., W. C. Ko, K. C. Cheng, C. C. Lee, C. C. Lai, and Y. C. Chuang. 2008. Comparison of prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with capsular K1/K2 and non-K1/K2 serotypes. Diagn.Microbiol.Infect.Dis. 62:1-6.
Figure Legend
Figure 1. Effect of all isolates on phagocytosis in according to their ST types and sites and region of isolation. The phagocytosis rate is calculated the percentage of the neutrophils ingested FITC-labeled bacteria at 15 minutes. Strains from liver abscess and carriage were labeled as open and filled symbol according to their isolated area. The isolates from Hong Kong, Singapore and Taiwan were labeled as circle, triangle, and square respectively: LA, liver abscess, Two ST23 laboratory capsule deficiency strains ( from previous study (23) were included as control for highly phagocytic susceptible strains. Except two ST23 isolates with phagocytic rate over 50% at 15 minutes, all isolates were highly resistant neutrophils phagocytosis.
Table 1. Specific primers used for amplification of the target genes of K. pneumoniae in this study.
Serotype (target gene) Primer Size of PCR product (bp) Reference
K1 (wzyKPK1) 5’-GGTGCTCTTTACATCATTGC-3’ 1283 (19) 5’-GCAATGGCCATTTGCGTTAG-3’ K2 (wzyKPK2) 5’-GACCCGATATTCATACTTGACAGAG-3’ 641 (19) 5’-CCTGAAGTAAAATCGTAAATAGATGGC-3’ K5 (wzxKPK5) 5’-TGGTAGTGATGCTCGCGA-3’ 280 (19) 5’-CCTGAACCCACCCCAATC-3’ RmpA 5’-ACTGGGCTACCTCTGCTTCA-3’ 536 (24) 5’-CTTGCATGAGCCATCTTTCA-3’ Aerobactin 5’-GCATAGGCGGATACGAACAT-3’ 556 (24) 5’-CACAGGGCAATTGCTTACCT-3’ Aerobactin 5’-CTGTCGGCATCGGTTTTATT-3’ 531 (24) 5’-TGGCGTGTCGATTATTACCA-3’
Table 2. MLST of serotype K1 isolates from liver abscess patients and from stool carriage of non-liver abscess subjects in Hong Kong, Singapore and Taiwan.
Serotype ST type (no. of isolates) No. of isolates Allelic profiles
HK† SG TW gapA inf mdh pgi pho rpoB tonB
LA¶ ST LA ST LA ST K1 23 (40) 5 5 9 2 10 9 2 1 1 1 9 4 12 163 (1) 1 2 1 1 1 9 1 12 249 (2) 1 1 2 1 44 1 16 4 66 367* (1) 1 18 15 26 16 11 13 106* 425* (1) 1 42 22 63* 22 86* 38 114 426* (1) 1 18 15 18 64* 11 13 50 138 (1) 1 18 22 26 23 31 13 49
*New ST types or alleles in this study.
†Isolates from: HK, Hong Kong; SG, Singapore; TW, Taiwan.
Table 3. Serotype K1 isolates from stool carriage and liver with different MLSTs in against serum complement killing (I would like to retain this table, perhaps, we may wait for reviewer’s suggestion).
Source ST type (no of isolates) Serum complement killing
Resistance Susceptible HK* SG* TW* HK SG TW Liver abscess 23 (24) 2 7 8 3 2 2 †163 (1) 1 249 (1) 1 Stool carriage 23 (16) 4 2 4 1 5 138 (1) 1 249 (1) 1 367 (1) 1
425 (1) 1
426 (1) 1
* No. of isolates from: HK, Hong Kong; SG, Singapore; TW, Taiwan.
Table 4. Virulence analysis by combining results obtained from phagocytosis, serum complement killing and mice lethality.
Source* MLST Areobactin Phagocytosis Serum LD50 (cfu)
Liver ST23 + S S ~1.2×104 Carriage ST23 + S S ~2.7×104 Liver ST23 + R S ~2.6×103 Carriage ST23 + R S <102 Carriage ST23 + R R <102 Liver ST23 + R R <102 Liver ST23 + R R <102 Liver ST23 + R R <102 Liver †ST163 + R S ~3.3×103 Liver ST249 - R R ~3.7×105 Carriage ST249 - R R ~3.0105 Carriage ST138 - R S ~2.4×105 Carriage ST367 + R S ~1.0103 Carriage ST425 - R S ~1.6105 Carriage ST426 - R S ~3.0×105
*All isolates were carrying rmpA.