0066-4804/11/$12.00 doi:10.1128/AAC.01777-10
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Loss of Heterozygosity of FCY2 Leading to the Development of
Flucytosine Resistance in Candida tropicalis
䌤
Yen-Ning Chen,
1Hsiu-Jung Lo,
2Chia-Chen Wu,
3Hui-Ching Ko,
3Te-Pin Chang,
2and Yun-Liang Yang
1,3*
Institute of Molecular Medicine and Bioengineering, National Chiao Tung University, Hsinchu,1Division of
Infectious Diseases, National Health Research Institutes, Miaoli,2and Department of
Biological Science and Technology, National Chiao Tung University, Hsinchu,3Taiwan
Received 20 December 2010/Returned for modification 28 January 2011/Accepted 3 March 2011
As fluconazole resistance becomes an emerging issue for treating infections caused by Candida tropicalis, searching for alternative becomes a prominent task. In the present study, 97 clinical isolates of C. tropicalis were tested for the susceptibilities to flucytosine (5FC) with the Etest method. Although only one isolate was resistant to 5FC, 30 susceptible isolates could produce resistant progeny after exposure to the drug. Interest-ingly, 22 of these 30 clinical isolates had a heterozygous G/T at the 145th position on FCY2, encoding purine-cytosine permease, whereas their progeny recovered from within the inhibitory ellipses had homozygous T/T, resulting in null alleles for both copies of the gene and produced only truncated proteins, effecting the 5FC resistance. Furthermore, we found that two major fluconazole-resistant clinical clones, diploid sequence type 98 (DST98) and DST140, had a homozygous G/G at the 145th position, and neither was able to produce 5FC-resistant progeny within the inhibitory ellipses. Hence, strains of C. tropicalis containing heterozygous alleles may develop 5FC resistance readily, whereas those with homozygous G/G wild-type alleles can be treated with 5FC. Subsequently, a combination of 5FC and another antifungal drug is applicable for treating infections of C. tropicalis.
The prevalence of invasive nosocomial Candida infections has increased significantly in association with the selective pressure of applying antibiotics, increased number of immu-nocompromised individuals, and invasive hospital procedures (38, 44). Although Candida albicans is the most prominent species causing candidemia in most situations, there has been a shift toward the more treatment-challenged non-albicans
Candida species (8, 28, 29, 36), of which the prevalences were
significantly different in various geographic areas (19).
Candida glabrata appears to be the most frequently isolated
non-albicans Candida species in Western countries, whereas, in Asia, it is Candida tropicalis (4, 7, 28, 36, 40, 41). In certain regions, it even surpassed C. albicans to become the most frequently isolated Candida species (4, 40). Furthermore, in-creasing prevalence of the resistance to fluconazole, the most commonly used antifungal in clinics, is an emerging issue (41, 42) in treating C. tropicalis infections.
Flucytosine (5FC) is one of the oldest antifungal drugs for treating human fungal infections such as candidiasis and cryp-tococcosis (33). Monotherapy with 5FC is limited because of the frequent development of resistance. Therefore, 5FC is mostly used in combination with another antifungal agent. Nevertheless, it may serve as an alternative for treating emerg-ing fluconazole-resistant C. tropicalis infections. Thus, it is in-teresting and important to determine the prevalence of 5FC resistance in clinically isolated C. tropicalis and the molecular mechanisms contributing to 5FC resistance.
FC is taken up by fungal cells and converted via 5-fluoro-uracil to 5-fluorouridine monophosphate (FUMP), which is then catalyzed by either one of two enzymes: cytosine deami-nase encoded by FCY1and uracil phosphoribosyl transferase (UPRT) encoded by FUR1. FUMP is in turn phosphorylated to 5-fluorouridine triphosphate (FUTP), which disturbs the protein synthesis by incorporation into RNA (30, 34). Alter-natively, the reduction of FUMP to 5-fluoro-2⬘-deoxyuridylate monophosphate (FdUMP) leads to the inhibition of the en-zyme thymidylate synthetase and thus DNA synthesis (15).
FCY2, encoding a purine-cytosine permease, is involved in the
uptake of 5FC, and URA3, encoding an orotidine 5⬘-phosphate decarboxylase, is involved in the metabolic pathway of uridyl-monophosphate (UMP) in nucleic acid synthesis. Mutations on
FCY1, FCY2, FUR1, or URA3 can result in 5FC resistance in
certain yeast species (5, 14, 16, 18, 23, 27, 33). Interestingly, 5FC-resistant clinical isolates have been reported to be genet-ically related (6, 14). Nevertheless, the mechanisms contribut-ing to 5FC resistance in C. tropicalis are not clear.
For diploid cells, the phenotypic change to display a reces-sive trait requires more than one step of genetic alteration. First of all, one of the two original alleles has to mutate to generate a genetic heterozygosity. Then, the other original allele is replaced by the newly mutated allele via mechanisms such as mitotic recombination, which leads to loss of heterozy-gosity (21). In contrast, if a diploid cell is heterozygous, then only one step is required to complete the loss of heterozygosity and display the mutant phenotype. Recently, Jacques et al. reported that a differential loss of heterozygosity in the diploid
Debaryomyces hansenii, phylogenetically related to C. albicans,
may result in the large genetic diversity found among isolates within this species (25). In vitro, it occurred in various popu-lations in the presence of antifungal drugs (2, 3, 13) and has
* Corresponding author. Mailing address: Department of Biological Science and Technology, National Chiao Tung University, 75 Pao-Ai Street, Hsinchu, Taiwan, Republic of China. Phone: 886 3 5712121, ext. 56920. Fax: 886 3 5729288. E-mail: [email protected].
䌤Published ahead of print on 21 March 2011.
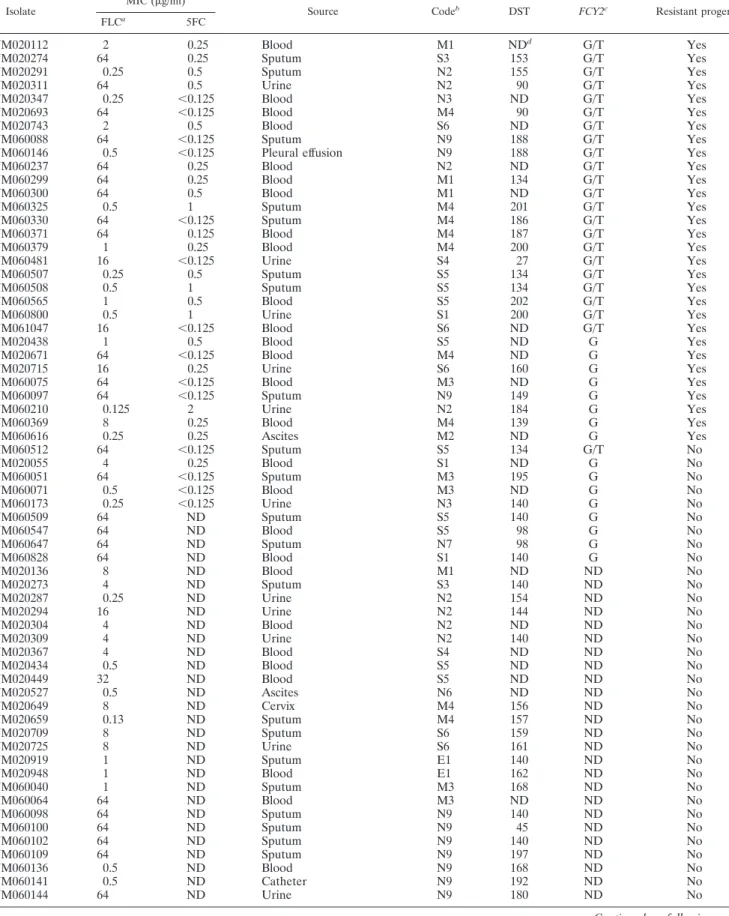
TABLE 1. Characterization of C. tropicalis clinical isolates collected from the TSARY studies
Isolate MIC (g/ml) Source Codeb DST FCY2c Resistant progeny
FLCa
5FC
YM020112 2 0.25 Blood M1 NDd G/T Yes
YM020274 64 0.25 Sputum S3 153 G/T Yes
YM020291 0.25 0.5 Sputum N2 155 G/T Yes
YM020311 64 0.5 Urine N2 90 G/T Yes
YM020347 0.25 ⬍0.125 Blood N3 ND G/T Yes
YM020693 64 ⬍0.125 Blood M4 90 G/T Yes
YM020743 2 0.5 Blood S6 ND G/T Yes
YM060088 64 ⬍0.125 Sputum N9 188 G/T Yes
YM060146 0.5 ⬍0.125 Pleural effusion N9 188 G/T Yes
YM060237 64 0.25 Blood N2 ND G/T Yes
YM060299 64 0.25 Blood M1 134 G/T Yes
YM060300 64 0.5 Blood M1 ND G/T Yes
YM060325 0.5 1 Sputum M4 201 G/T Yes
YM060330 64 ⬍0.125 Sputum M4 186 G/T Yes
YM060371 64 0.125 Blood M4 187 G/T Yes
YM060379 1 0.25 Blood M4 200 G/T Yes
YM060481 16 ⬍0.125 Urine S4 27 G/T Yes
YM060507 0.25 0.5 Sputum S5 134 G/T Yes
YM060508 0.5 1 Sputum S5 134 G/T Yes
YM060565 1 0.5 Blood S5 202 G/T Yes
YM060800 0.5 1 Urine S1 200 G/T Yes
YM061047 16 ⬍0.125 Blood S6 ND G/T Yes
YM020438 1 0.5 Blood S5 ND G Yes
YM020671 64 ⬍0.125 Blood M4 ND G Yes
YM020715 16 0.25 Urine S6 160 G Yes
YM060075 64 ⬍0.125 Blood M3 ND G Yes
YM060097 64 ⬍0.125 Sputum N9 149 G Yes
YM060210 0.125 2 Urine N2 184 G Yes
YM060369 8 0.25 Blood M4 139 G Yes
YM060616 0.25 0.25 Ascites M2 ND G Yes
YM060512 64 ⬍0.125 Sputum S5 134 G/T No YM020055 4 0.25 Blood S1 ND G No YM060051 64 ⬍0.125 Sputum M3 195 G No YM060071 0.5 ⬍0.125 Blood M3 ND G No YM060173 0.25 ⬍0.125 Urine N3 140 G No YM060509 64 ND Sputum S5 140 G No YM060547 64 ND Blood S5 98 G No YM060647 64 ND Sputum N7 98 G No YM060828 64 ND Blood S1 140 G No YM020136 8 ND Blood M1 ND ND No YM020273 4 ND Sputum S3 140 ND No YM020287 0.25 ND Urine N2 154 ND No YM020294 16 ND Urine N2 144 ND No YM020304 4 ND Blood N2 ND ND No YM020309 4 ND Urine N2 140 ND No YM020367 4 ND Blood S4 ND ND No YM020434 0.5 ND Blood S5 ND ND No YM020449 32 ND Blood S5 ND ND No YM020527 0.5 ND Ascites N6 ND ND No YM020649 8 ND Cervix M4 156 ND No YM020659 0.13 ND Sputum M4 157 ND No YM020709 8 ND Sputum S6 159 ND No YM020725 8 ND Urine S6 161 ND No YM020919 1 ND Sputum E1 140 ND No YM020948 1 ND Blood E1 162 ND No YM060040 1 ND Sputum M3 168 ND No YM060064 64 ND Blood M3 ND ND No YM060098 64 ND Sputum N9 140 ND No YM060100 64 ND Sputum N9 45 ND No YM060102 64 ND Sputum N9 140 ND No YM060109 64 ND Sputum N9 197 ND No YM060136 0.5 ND Blood N9 168 ND No YM060141 0.5 ND Catheter N9 192 ND No YM060144 64 ND Urine N9 180 ND No
been suggested to contribute to drug resistance in clinical C.
albicans isolates. For example, loss of heterozygosity was found
at and around ERG11, the target of azole drugs, to decrease susceptibility to fluconazole (22, 37). In addition, homozygosity for gain-of-function mutations in TAC1, an activator of ABC transporters, resulted in elevated levels of azole resistance (11). Similar phenomena have also been reported for mutations on multidrug resistance regulator (MRR1), a regulator for multidrug resistance, MDR1, an efflux pump contributing to azole resistance (17), and on GSC1 (FKS1), a glucan synthase catalytic subunit, involved in micafungin resistance (26). Loss of heterozygosity in C. tropicalis has also been reported (24). Nevertheless, whether loss of heterozygosity contributes to drug resistance in this species has not been reported.
In the present study, we screened and selected several 5FC-resistant C. tropicalis to show that isolates with a null mutation in one of the FCY2 allele, when exposed to 5FC, were readily to undergo loss of heterozygosity to effect the homozygous state with the mutant allele and lead to resistance. Hence, strains containing homozygous G/G wild-type alleles can be treated with 5FC, whereas those containing heterozygous G/T will require different medication. In light of the emerging flu-conazole-resistant C. tropicalis infection, a combination of 5FC
and another antifungal drug other than fluconazole is a rea-sonable choice for treatments.
MATERIALS AND METHODS
Strains and media. The C. tropicalis clinical isolates collected during the Taiwan Surveillance of Antimicrobial Resistance of Yeasts (TSARY) studies in 2002 and 2006 (43, 45) were used for screening the 5FC resistance strains listed in Table 1. Yeast-peptone-dextrose (YPD; 1% yeast extract, 2% peptone, and 2% dextrose) and synthetic dextrose (SD; 0.67% yeast nitrogen base without amino acid and 2% dextrose) were prepared as described previously (32). Cells were grown in either YPD or SD unless otherwise noted. The compounds for addition to media were from Difco unless otherwise noted.
Constructions of different FCY2 alleles.FCY2ORF, a 2,258-bp KpnI-XhoI fragment comprising of the entire FCY2 coding region and its flanking se-quences, was amplified from the genomic DNA of YM020192 by using the primers HJL1420 and HJL1424 (Table 2) and cloned into pSF2A containing the
SAT1 flipper cassette (31). FCY2d, a 528-bp SacII-SacI fragment complementary
to the 46 bp at the 3⬘ end sequence of FCY2 open reading frame (ORF), as well as its downstream region, was amplified by primers HJL1422 and HJL1423 and cloned into pSF2A containing the FCY2ORF fragment. LOB319 contained the G allele of FCY2, whereas LOB320 contained the T allele. The KpnI-SacI digested fragments of LOB319 containing homozygous G alleles were trans-formed into YM020291 and YM060800 to obtain YLO415 and YLO447, respec-tively. The KpnI-SacI digested fragments of LOB320 containing homozygous T alleles were transformed into YM020291 and YM060800 to obtained YLO417 and YLO440, respectively. The point mutation was generated with fusion PCR,
TABLE 1—Continued
Isolate MIC (g/ml) Source Codeb DST FCY2c Resistant progeny
FLCa 5FC YM060147 0.25 ND Catheter N9 198 ND No YM060172 0.5 ND Urine N3 171 ND No YM060175 64 ND Urine N3 179 ND No YM060177 64 ND Urine N3 149 ND No YM060184 4 ND Blood N3 ND ND No YM060185 64 ND Blood N3 ND ND No
YM060302 64 ND Pleural effusion M1 185 ND No
YM060310 64 ND Blood M1 ND ND No YM060327 64 ND Urine M4 140 ND No YM060342 0.25 ND Urine M4 196 ND No YM060354 1 ND Sputum M4 191 ND No YM060383 0.5 ND Blood M4 189 ND No YM060450 64 ND Sputum N5 98 ND No YM060451 64 ND Sputum N5 98 ND No
YM060500 0.5 ND Bronchoalveolar lavage S5 190 ND No
YM060529 64 ND Sputum S5 98 ND No YM060533 64 ND Ascites S5 ND ND No YM060541 64 ND Blood S5 ND ND No YM060559 64 ND Blood S5 183 ND No YM060590 64 ND Urine M2 181 ND No YM060607 64 ND Blood M2 ND ND No YM060689 64 ND Blood M6 ND ND No YM060767 64 ND Blood E1 ND ND No YM060776 64 ND Catheter E1 179 ND No YM060792 0.5 ND Urine S1 199 ND No YM060804 0.5 ND Urine S1 194 ND No YM060805 64 ND Urine S1 182 ND No YM060808 64 ND Blood S1 ND ND No YM060812 1 ND Blood S1 193 ND No YM060925 16 ND Blood M5 ND ND No YM060926 16 ND Blood M5 ND ND No
YM061045 64 ND Peritoneal fluid S6 ND ND No
YM061051 64 ND Ascites N5 ND ND No
a
FLC, fluconazole.
b
That is, the location of the collection source.
c
The nucleotide at position 145 of FCY2.
d
in which three separate PCRs were conducted as following. Primers HJL1420 and HJL2103 were used to amplified 5⬘ end of fragment from LOB319 and HJL2102 and HJL1424 were used to amplify 3⬘ end of the fragment from LOB319. The FCY2LOB319T fragment amplified by primers HJL1420 and HJL1424 using 5⬘ and 3⬘ end fragments as templates was used to replace the KpnI-XhoI fragment of LOB319 to generate LOB383. FCY2 5⬘ and 3⬘ end fragments were amplified from LOB320 by primer pairs HJL1420/HJL2101 and HJL2100/HJL1424, respectively. The FCY2LOB320G fragment, generated by amplification by the primers HJL1420 and HJL1424 and with the 5⬘ and 3⬘ end fragments as templates, then replaced the KpnI-XhoI fragment of LOB320 to generate LOB384. The KpnI-SacI-digested fragments of LOB383 and LOB384 were transformed into YM020291 competent cells by electroporation to generate
YLO468 and YLO466, respectively. Finally, the mutant isolates were confirmed by colony PCR and sequencing.
Antifungal susceptibility tests.Susceptibilities to 5FC of all C. tropicalis iso-lates collected in TSARY 2002 and 2006 (43, 45) were tested. The Etest assay was used to determine the susceptibilities to antifungal agents for C. tropicalis isolates. Homogenized colonies from an overnight YPD agar medium were transferred in 0.85% NaCl to achieve a density of 5⫻ 106
cell/ml. A sterile cotton swab was dipped into the inoculum suspension and used to swab the entire agar surface of the RPMI medium (Gibco-BRL) evenly. The 5FC (from 0.002 to 32 g/ml) drug strips (AB Biodisk, Solna, Sweden) were then applied onto the RPMI agar medium when the excess moisture was absorbed completely. Two colonies (when applicable) were selected within the inhibition ellipses of each of
FIG. 1. Isolation of flucytosine-resistant progeny from the clinical isolates using Etest. (a) Resistant isolate YM060607; (b) susceptible isolate with clear inhibitory ellipses YM020367; (c to j) susceptible isolates producing progeny within inhibitory ellipses YM060051 (c), YM020291 (e), YM060097 (g), and YM060800 (i) and their progeny YM060051-R1 (d), YM020291-R1 (f), YM060097-R1 (h), and YM060800-R1 (j). The results were photographed after 72 h (a, b, c, e, g, and i) or 48 h (d, f, h, and j) of incubation at 35°C. Arrows indicate small colonies.
TABLE 2. Primers used in this study
HJL designation Primer Sequence (5⬘–3⬘)a
Application
HJL1205 CtFCY1-f ATCATTAGTTCAGATGGTAAAGTCTTG PCR and sequencing
HJL1206 CtFCY1-r CCTTTTTAGTAACATGTCTATTCTCCA PCR and sequencing
HJL1211 CtFUR1-f TCATCAAAACCATGTCTGCTG PCR and sequencing
HJL1212 CtFUR1-r AAGTGTATGTAGTGATAATTGCTATGC PCR and sequencing
HJL1413 CtURA3-f ATTGGATAGTCCCTCTAAACTCACTACTA PCR and sequencing
HJL1414 CtURA3 AGCATTAGTTATATCACTCCACGATGAA Sequencing
HJL1415 CtURA3 TGCCGATATTGGAAATACAGTTA Sequencing
HJL1416 CtURA3-r AATCAACTATTCAAGTTGACCG PCR and sequencing
HJL814 CaSAT1-f CTCAACATGGAACGATCTAGC PCR
HJL1207 CtFCY2-f TGCCCATAAATTAAATGCAGAA Sequencing
HJL1208 CtFCY2-r GGAAGCAACAAACCCAAAAA Sequencing
HJL1209 CtFCY2-f TGCTGCCGATTATGTTGTTT Sequencing
HJL1210 CtFCY2-r GTGAAAACGAGCCAATCCAT Sequencing
HJL1420 CtFCY2-f (KpnI) ggtaccTCAACTCAACCCCAAAGT Fusion PCR and sequencing
HJL1421 CtFCY2-r (XhoI) ctcgagCCCAAGGAGAAAGTAGCA PCR
HJL1422 CtFCY2-f CGGATTCAATGTAGCCAG PCR
HJL1423 CtFCY2-r GTCATTCCATGTCGTGGT PCR
HJL1424 CtFCY2-r (XhoI) ctcgagGTCATTCCATGTCGTGGT Fusion PCR and sequencing
HJL1477 CtFCY2-r out of B (3⬘) CTGTTGCTCCAGGTGAATCA PCR
HJL1753 CtFCY2f TCGTTGCTTGTGTTGGTTGG Sequencing
HJL2100 CtFCY2-145Gf CATAAATTAAATGCAGAAACTAAAGGTATTG Fusion PCR
HJL2101 CtFCY2-145Gr CAATACCTTTAGTTTCTGCATTTAATTTATG Fusion PCR
HJL2102 CtFCY2-145Tf CATAAATTAAATGCATAAACTAAAGGTATTG Fusion PCR
HJL2103 CtFCY2-145Tr CAATACCTTTAGTTTATGCATTTAATTTATG Fusion PCR
HJL2104 CtFCY2-f CTTCTCCTTAACTACCTTTTCCTCC Sequencing
the 35 isolates and grown on YPD agar medium in the absence of drug for 2 days before they were kept in 50% glycerol at⫺80°C for further analysis.
The susceptibilities to 5FC of the 67 progeny within the inhibition ellipses of the 35 isolates along with their parental isolates were determined by the broth microdilution method according to the procedures in previous study (45), which is modified from the guidelines of Clinical and Laboratory Standards Institute (10). First, all isolates were grown on the YPD agar medium overnight. The RPMI medium 1640 (Gibco-BRL catalog no. 31800-022), which contains 0.2% glucose, was used for the testing. Strains from the American Type Culture Collection, including C. albicans (ATCC 90028), C. krusei (ATCC 6258), and C.
parapsilosis (ATCC 22019), were used as the standard controls. The
concentra-tion of 5FC and fluconazole ranged from 0.125 to 64g/ml. Cell growth was determined by using spectrophotometric measurement with a Biotrak II plate reader (Amersham Biosciences, Biochrom, Ltd., Cambridge, England) after a 48-h incubation at 35°C. For fluconazole, isolates with MICs ofⱖ64 g/ml were considered to be resistant, whereas those with an MICⱕ8 g/ml were suscep-tible. Isolates with MICs falling in between (16 to 32g/ml) were susceptible-dose dependent. For 5FC, isolates with MICsⱖ32 g/ml were considered resis-tant, whereas those with MICs ofⱕ4 g/ml were susceptible. Isolates with MICs falling in between (8 to 16g/ml) were intermediate. In addition, we used Etest to verify the susceptibilities of 5FC of at least one resistant progeny from each clinical parental isolate.
RESULTS AND DISCUSSION
Screening flucytosine-resistant isolates of C. tropicalis. A total of 27 and 70 C. tropicalis clinical isolates collected for the
TSARY studies in 2002 and 2006, respectively (43, 45), were tested for susceptibilities to 5FC by the Etest method (Table 2). Only one isolate, YM060607 (Fig. 1a), was resistant to 5FC. It was also resistant to fluconazole (Table 2). This low preva-lence of 5FC resistance may be due to the rare use of this drug in Taiwan. The inhibition ellipses of 61 isolates were clear, such as that of YM020367 (Fig. 1b). In contrast, colonies ap-peared within the inhibition ellipses of the remaining 35 iso-lates. Few had small colonies on the edges of the inhibition ellipses such as that of YM060051 (Fig. 1c), whereas others had colonies evenly distributed within the inhibition ellipses, such as those of YM020291 (Fig. 1e), YM060097 (Fig. 1g), and YM060800 (Fig. 1i). The 5FC susceptibilities were determined for the 67 isolates recovered from within the inhibitory ellipses, as well as their parental isolates by the broth microdilution method (Table 2). Of the 67 isolates 55 (82.1%), derived from 30 different clinical isolates, still displayed resistance to 5FC, whereas their parental isolates were susceptible. For the re-maining five clinical isolates, YM060512 produced a progeny, YM060512-1, with intermediate susceptibility to 5FC, and the progeny from the other four clinical isolates, YM020055, YM060051, YM060071, and YM060173, were still 5FC suscep-tible. The 5FC resistance phenotype of at least one progeny of
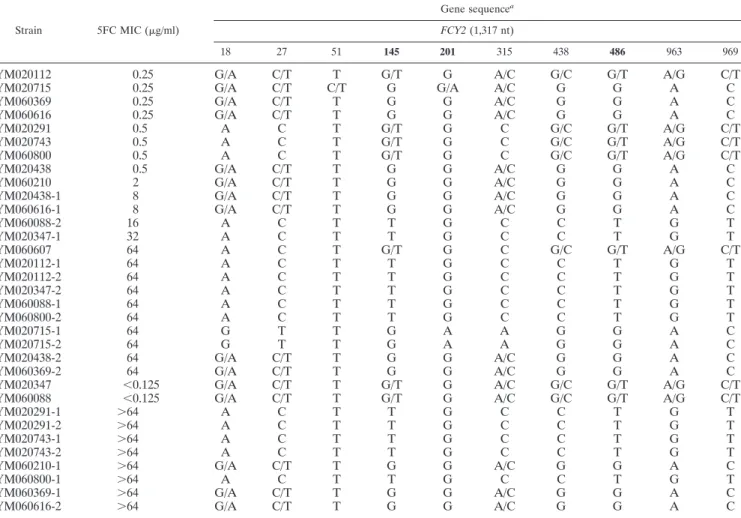
TABLE 3. Sequence of different genes of 33 C. tropicalis isolates
Strain 5FC MIC (g/ml)
Gene sequencea
FCY2 (1,317 nt)
18 27 51 145 201 315 438 486 963 969
YM020112 0.25 G/A C/T T G/T G A/C G/C G/T A/G C/T
YM020715 0.25 G/A C/T C/T G G/A A/C G G A C
YM060369 0.25 G/A C/T T G G A/C G G A C
YM060616 0.25 G/A C/T T G G A/C G G A C
YM020291 0.5 A C T G/T G C G/C G/T A/G C/T
YM020743 0.5 A C T G/T G C G/C G/T A/G C/T
YM060800 0.5 A C T G/T G C G/C G/T A/G C/T
YM020438 0.5 G/A C/T T G G A/C G G A C
YM060210 2 G/A C/T T G G A/C G G A C
YM020438-1 8 G/A C/T T G G A/C G G A C
YM060616-1 8 G/A C/T T G G A/C G G A C
YM060088-2 16 A C T T G C C T G T YM020347-1 32 A C T T G C C T G T YM060607 64 A C T G/T G C G/C G/T A/G C/T YM020112-1 64 A C T T G C C T G T YM020112-2 64 A C T T G C C T G T YM020347-2 64 A C T T G C C T G T YM060088-1 64 A C T T G C C T G T YM060800-2 64 A C T T G C C T G T YM020715-1 64 G T T G A A G G A C YM020715-2 64 G T T G A A G G A C
YM020438-2 64 G/A C/T T G G A/C G G A C
YM060369-2 64 G/A C/T T G G A/C G G A C
YM020347 ⬍0.125 G/A C/T T G/T G A/C G/C G/T A/G C/T
YM060088 ⬍0.125 G/A C/T T G/T G A/C G/C G/T A/G C/T
YM020291-1 ⬎64 A C T T G C C T G T
YM020291-2 ⬎64 A C T T G C C T G T
YM020743-1 ⬎64 A C T T G C C T G T
YM020743-2 ⬎64 A C T T G C C T G T
YM060210-1 ⬎64 G/A C/T T G G A/C G G A C
YM060800-1 ⬎64 A C T T G C C T G T
YM060369-1 ⬎64 G/A C/T T G G A/C G G A C
YM060616-2 ⬎64 G/A C/T T G G A/C G G A C
aNumbers in boldface indicate a change in amino acid due to different nucleotides. Genetic details: (i) FCY2, 145th G, Glu; T, stop; 201th G, Trp; A, stop; 486th
each clinical parental isolate was confirmed by Etest, such as those of YM020291-R1 (Fig. 1f), YM060097-R1 (Fig. 1h), and YM060800-R1 (Fig. 1j). In contrast, YM060051-R1 was still susceptible to 5FC, which is also consistent with the results of the broth microdilution method (Fig. 1d).
Sequencing the four known genes involved in flucytosine resistance.To determine the mechanisms contributing to 5FC resistance, we sequenced the FCY1, FCY2, FUR1, and URA3 ORFs of 33 isolates (Table 3), a group comprising 1 resistant isolate (YM060607), 11 susceptible parental isolates (ran-domly selected from the 30 isolates producing 5FC-resistant progeny), and 21 progeny from these isolates. Unlike FCY1 in either C. albicans or C. lusitaniae (20, 23), the sequence of
FCY1 in the present study is highly conserved. Neither
single-nucleotide polymorphism (SNP) nor any other variation was detected among all of the tested isolates. For FUR1, six SNPs were detected. Nevertheless, all were synonymous alterations since they did not change the amino acid residues in the en-coded proteins. There were three SNPs detected in URA3. The SNP at position 345 was a synonymous alteration. The one at 775th position allowed the 259th amino acid residue to be either threonine (ACC) or alanine (GCC), and that at 971st position made the 264th residue to be either threonine (ACC) or isoleucine (ATC).
When we compared the FCY2 sequences of the isolates from
the present study to that of CTRG_02059 from the C. tropicalis database of the Broad Institute (http://www.broadinstitute .org/annotation/genome/candida_group/GeneDetails.html?sp ⫽S7000000625961821), we found that CTRG_02059 contained a nonsense mutation at the 201st position, which caused the ATG at positions 214 to 216 to be denoted as the translation initiation site. Therefore, FCY2 in fact encodes a 509-amino-acid purine-cytosine permease (HQ166001), and its translational initiation site corresponds to position 2136158 at the supercontig 2 (Fig. 2). Of 10 SNPs detected in FCY2, 7 were synonymous alterations. In contrast, both the 145th nucleotide alteration, G to T, and the 201st nucleotide alteration, G to A, resulted in truncated purine-cytosine permeases. The remaining one was at the 486th position, a G-to-T alteration changing methionine to isoleucine at the 162nd amino acid residue.
Of the 11 susceptible parental isolates, 6, including YM020112, YM020291, YM020347, YM020743, YM060088, and YM060800, had a heterozygous G/T at position 145, and their progeny had a homozygous T/T in FCY2. Three isolates (YM020112, YM020347, and YM060088) had eight SNPs, and the other three (YM020291, YM020743, and YM060800) had five SNPs within the FCY2 ORF. We assessed the results by cloning PCR products of FCY2 from YM020291 into a vector and sequencing several independent clones. We found that there were two distinct FCY2 alleles in the YM020291 isolate.
TABLE 3—Continued Gene sequencea FUR1 (657 nt) URA3 (807 nt) 21 93 127 431 501 507 345 775 791 A T A/G C T T A G C/T A T A/G C T T A G C/T A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C A/T T A C C/T C/T A G C A T A/G C T T A G C/T A C/T A C/T C/T C/T A/G A/G C A T A/G C T T A G T A T A/G C T T A G T A T A/G C T T A G C/T A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A T A/G C T T A G C/T A T A/G C T T A G C/T A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C A T A/G C T T A G C/T A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A/T T A C C/T C/T A G C A C/T G C C/T C/T A G C A/T T A C C/T C/T A G C A C/T G C C/T C/T A G C A C/T G C C/T C/T A G C
One allele was noted as a G allele, containing G, G, G, A, and C at positions 145, 438, 486, 963, and 969, respectively, and the other as the T allele, containing T, C, T, G, and T at the same positions. Hence, the G/T to T/T alteration at position 145 did not occur via single site mutations. Most likely, it was achieved via replacing the G allele with the T allele.
Loss of heterozygosity in FCY2 heterozygous nonsense mu-tants leading to 5FC resistance. Strains with homozygous G alleles and T alleles of FCY2 were constructed in YM020291 and YM060800 strains. The YLO415 and YLO 447 (GGGAC/ GGGAC) isolates, harboring homozygous G alleles, were sus-ceptible to 5FC (Fig. 3c and d), whereas YLO417 and YLO440 (TCTGT/TCTGT), harboring homozygous T alleles, were re-sistant to 5FC (Fig. 3e and f). Furthermore, we used YM020291 as the parental stain to determine the effect of the nonsense mutation at position 145 of FCY2. We performed site-directed mutagenesis to construct the YLO466 (TGGAC/ TCTGT) strain, in which the G at position 145 in the G allele of FCY2 of YM020291 (GGGAC/TCTGT) was replaced by a T, and the YLO468 (GGGAC/GCTGT) strain, in which the T at position 145 in the T allele was replaced by a G. We found that the YLO468 isolate, with a homozygous G, was suscepti-ble to 5FC (Fig. 3g), whereas the YLO466 isolate, containing a homozygous T at position 145, was resistant to 5FC (Fig. 3h). These results demonstrated that a nonsense mutation in FCY2 at position 145, when in a homozygous state, sufficiently con-tributed to the 5FC-resistant phenotype.
Of the 30 clinical isolates producing 5FC-resistant progeny, 22 had the G/T SNP at position 145 of FCY2 (the first 30 isolates in Table 1). Furthermore, all but one (YM060416-2) progeny derived from these 22 clinical isolates had a homozy-gous T/T at that position. In fact, except for YM060512, all parental clinical isolates containing the G/T SNP at position
145 produced 5FC-resistant progeny. In contrast, all eight tested parental clinical isolates with clear inhibitory ellipses had a homozygous G/G at position 145. Hence, a nonsense mutation in FCY2, followed by loss of heterozygosity can be a major mechanism contributing to 5FC resistance. This is the first study to demonstrate that loss of heterozygosity and al-teration in 5FC susceptibility can be readily detected in C.
tropicalis after exposure to the drug.
Loss-of-heterozygosity can be due to one of the following: chromosome loss and duplication, break-induced replication, allelic recombination, and gene conversion (1, 11, 17, 26). Various mechanisms may reflect different expenses for fitness under diverse environmental conditions. Previous studies in C.
albicans suggest that loss of heterozygosity under in vitro
lab-oratory culture conditions mainly resulted from chromosome loss and duplication, whereas in clinical isolates it occurred via mitotic recombination (12, 17, 39). Our preliminary result sug-gests that the loss of heterozygosity of FCY2 predominantly resulted from break-induced replication/allelic recombination (unpublished data).
YM020715 had G/A at position 201. The YM020715-1 and YM020715-2 recovered from within the inhibitory ellipse had homozygous A/A at that position (Table 3). This G-to-A substitution, causing a nonsense mutation, followed by con-verting to A/A, contributed to the 5FC-resistant phenotype of YM020715-1 and YM020715-2. Of the isolates collected in Taiwan, we found that the 145th SNP occurred at a much higher frequency than the 201st one (23 versus 1, thus far). Furthermore, 42 of the 43 resistant progeny from the 22 clin-ical parental isolates with the G/T alleles contained homozy-gous T/T alleles. Hence, genomic alterations, resulting in ho-mozygosity occur more frequently than the acquisition of an independent mutation in the second allele or a mutation on a
FIG. 2. Sequence of FCY2. The FCY2 encodes a purine-cytosine permease consisting of 509 amino acids. The A of the translational initiation site of FCY2 corresponds to position 2136158 at the supercontig 2. Positions 145 and 201 are marked by arrowheads.
different gene. Similar phenomena have been reported in C.
al-bicans in the mechanistic studies of fluconazole resistance. One
involved two hyperactive TAC1 alleles from isolates overex-pressing CDR1 and CDR2 (11) and another two different
MRR1 mutants overexpressing MDR1 (17).
Existence of resistance with unknown mechanisms. The YM060607 isolate was the only 5FC-resistant clinical isolate among our collection in the TSARY studies. It had a G/T at position 145 of FCY2 and an A/G at position 775 of URA3. Hence, the development of resistance is not based on the mechanism and genes mentioned above. In addition to muta-tions within ORFs, alteramuta-tions on the level of gene expression due to mutations in the untranslated regions or their trans-regulators may also result in resistance. The mechanisms con-tributing to the increase in 5FC MICs of other progeny are under investigation. These progeny included 10 resistant iso-lates (YM020438-2, YM060210-1, YM060075-1, YM020671-1,
YM020671-2, YM060097-1, YM060097-2, YM060369-1,
YM060369-2, and YM060616-2) and 3 intermediate isolates (YM020438-1, YM060616-1, and YM060512-1).
Conclusion.In the present study, we found that FCY2’s loss of heterozygosity is the major molecular mechanism con-tributing to the 5FC-resistant phenotype of C. tropicalis. The increasing rate of reduced susceptibility to fluconazole in C.
tropicalis has considerable clinical importance. In addition,
ap-proximately half of the fluconazole-resistant C. tropicalis iso-lates collected in Taiwan belonged to diploid sequence type 98 (DST98) and DST140 (9, 35). In the present study, we found that DST98 and DST140 isolates had homozygous G/G at position 145, and none produced 5FC-resistant progeny within the inhibitory ellipses. Among all of the tested isolates, only
one, YM060607, was resistant to both 5FC and fluconazole. Hence, 5FC in combination with another antifungal drug can be considered for treating fluconazole-resistant C. tropicalis.
ACKNOWLEDGMENTS
We thank H. T. Chen and C. C. Lin for their technical assistance. This study was supported in part by grants ID-099-PP-09 (H.-J.L.), NSC 98-3112-B-009-001, NSC 99-2320-B-009-001-MY3, and ATU Program NCTU 99W962 (Y.-L.Y.).
REFERENCES
1. Andersen, M. P., Z. W. Nelson, E. D. Hetrick, and D. E. Gottschling. 2008. A genetic screen for increased loss of heterozygosity in Saccharomyces
cerevi-siae. Genetics 179:1179–1195.
2. Barchiesi, F., et al. 2000. Experimental induction of fluconazole resistance in
Candida tropicalis ATCC 750. Antimicrob. Agents Chemother. 44:1578–
1584.
3. Calvet, H. M., M. R. Yeaman, and S. G. Filler. 1997. Reversible fluconazole resistance in Candida albicans: a potential in vitro model. Antimicrob. Agents Chemother. 41:535–539.
4. Chai, Y. A., et al. 2007. Predominance of Candida tropicalis bloodstream infections in a Singapore teaching hospital. Med. Mycol. 45:435–439. 5. Chapeland-Leclerc, F., et al. 2005. Inactivation of the FCY2 gene encoding
purine-cytosine permease promotes cross-resistance to flucytosine and flu-conazole in Candida lusitaniae. Antimicrob. Agents Chemother. 49:3101– 3108.
6. Chen, K. W., Y. C. Chen, Y. H. Lin, H. H. Chou, and S. Y. Li. 2009. The molecular epidemiology of serial Candida tropicalis isolates from ICU pa-tients as revealed by multilocus sequence typing and pulsed-field gel elec-trophoresis. Infect. Genet. Evol. 9:912–920.
7. Chen, P. L., et al. 17 December 2009. Species distribution and antifungal susceptibility of blood Candida isolates at a tertiary hospital in southern Taiwan, 1999–2006. Mycoses [E-pub ahead of print.]
8. Cheng, M. F., et al. 2004. Distribution and antifungal susceptibility of
Can-dida species causing candidemia from 1996 to 1999. Diagn. Microbiol. Infect.
Dis. 48:33–37.
9. Chou, H. H., H. J. Lo, K. W. Chen, M. H. Liao, and S. Y. Li. 2007. Multilocus sequence typing of Candida tropicalis shows clonal cluster enriched in iso-lates with resistance or trailing growth of fluconazole. Diagn. Microbiol. Infect. Dis. 58:427–433.
FIG. 3. Effects on flucytosine susceptibility of different FCY2 mutants. The susceptibilities of different strains were determined using Etest. Parental isolates YM020291 (a) and YM060800 (b) with GGGAC/TCTGT at positions 145, 438, 486, 963, and 969 of FCY2 are shown. Additional isolates: YLO415 (c) and YLO447 (d) with GGGAC/GGGAC, YLO417 (e) and YLO440 (f) with TCTGT/TCTGT, YLO468 with GGGAC/ GCTGT (g), and YLO466 with TGGAC/TCTGT (h). The results were photographed after 48 h of incubation at 35°C.
10. Clinical Laboratory Standards Institute. 2008. Reference method for broth dilution antifungal susceptibility testing of yeasts; approved stan-dard, 3rd ed. CLSI document M27–A3. Clinical Laboratory Standards Institute, Wayne, PA.
11. Coste, A., et al. 2006. A mutation in Tac1p, a transcription factor regulating
CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to
mediate antifungal resistance in Candida albicans. Genetics 172:2139–2156. 12. Coste, A. T., M. Karababa, F. Ischer, J. Bille, and D. Sanglard. 2004. TAC 1, transcriptional activator of CDR genes, is a new transcription factor in-volved in the regulation of Candida albicans ABC transporters CDR1 and
CDR2. Eukaryot. Cell 3:1639–1652.
13. Cowen, L. E., et al. 2000. Evolution of drug resistance in experimental populations of Candida albicans. J. Bacteriol. 182:1515–1522.
14. Desnos-Ollivier, M., et al. 2008. Clonal population of flucytosine-resistant
candida tropicalis from blood cultures, Paris, France. Emerg. Infect. Dis.
14:557–565.
15. Diasio, R. B., J. E. Bennett, and C. E. Myers. 1978. Mode of action of 5-fluorocytosine. Biochem. Pharmacol. 27:703–707.
16. Dodgson, A. R., K. J. Dodgson, C. Pujol, M. A. Pfaller, and D. R. Soll. 2004. Clade-specific flucytosine resistance is due to a single nucleotide change in the FUR1 gene of Candida albicans. Antimicrob. Agents Chemother. 48: 2223–2227.
17. Dunkel, N., J. Blass, P. D. Rogers, and J. Morschha¨user.2008. Mutations in the multidrug resistance regulator MRR1, followed by loss of heterozygosity, are the main cause of MDR1 overexpression in fluconazole-resistant Candida
albicans strains. Mol. Microbiol. 69:827–840.
18. Edlind, T. D., and S. K. Katiyar. 2010. Mutational analysis of flucytosine resistance in Candida glabrata. Antimicrob. Agents Chemother. 54:4733– 4738.
19. Falagas, M. E., N. Roussos, and K. Z. Vardakas. 2010. Relative frequency of albicans and the various non-albicans Candida spp. among candidemia iso-lates from inpatients in various parts of the world: a systematic review. Int. J. Infect. Dis. [Epub ahead of print.]
20. Florent, M., et al. 2009. Nonsense and missense mutations in FCY2 and
FCY1 genes are responsible for flucytosine resistance and
flucytosine-flu-conazole cross-resistance in clinical isolates of Candida lusitaniae. Antimi-crob. Agents Chemother. 53:2982–2990.
21. Forche, A., G. May, and P. T. Magee. 2005. Demonstration of loss of heterozygosity by single-nucleotide polymorphism microarray analysis and alterations in strain morphology in Candida albicans strains during infection. Eukaryot. Cell 4:156–165.
22. Franz, R., et al. 1998. Multiple molecular mechanisms contribute to a step-wise development of fluconazole resistance in clinical Candida albicans strains. Antimicrob. Agents Chemother. 42:3065–3072.
23. Hope, W. W., L. Tabernero, D. W. Denning, and M. J. Anderson. 2004. Molecular mechanisms of primary resistance to flucytosine in Candida
albi-cans. Antimicrob. Agents Chemother. 48:4377–4386.
24. Jacobsen, M. D., et al. 2008. Molecular phylogenetic analysis of Candida
tropicalis isolates by multilocus sequence typing. Fungal Genet. Biol. 45:
1040–1042.
25. Jacques, N., et al. 2010. Population polymorphism of nuclear mitochondrial DNA insertions reveals widespread diploidy associated with loss of heterozy-gosity in Debaryomyces hansenii. Eukaryot. Cell 9:449–459.
26. Niimi, K., et al. 2010. Clinically significant micafungin resistance in Candida
albicans involves modification of a glucan synthase catalytic subunit GSC1
(FKS1) allele followed by loss of heterozygosity. J. Antimicrob. Chemother.
65:842–852.
27. Papon, N., et al. 2007. Molecular mechanism of flucytosine resistance in
Candida lusitaniae: contribution of the FCY2, FCY1, and FUR1 genes to
5-fluorouracil and fluconazole cross-resistance. Antimicrob. Agents Che-mother. 51:369–371.
28. Pfaller, M. A., and D. J. Diekema. 2007. Epidemiology of invasive candidi-asis: a persistent public health problem. Clin. Microbiol. Rev. 20:133–163. 29. Pfaller, M. A., et al. 2000. Bloodstream infections due to Candida species:
SENTRY antimicrobial surveillance program in North America and Latin America, 1997–1998. Antimicrob. Agents Chemother. 44:747–751. 30. Polak, A., and H. J. Scholer. 1975. Mode of action of 5-fluorocytosine and
mechanisms of resistance. Chemotherapy 21:113–130.
31. Reuss, O., A. Vik, R. Kolter, and J. Morschhauser. 2004. The SAT1 flipper, an optimized tool for gene disruption in Candida albicans. Gene 341:119– 127.
32. Sherman, F. 2002. Getting started with yeast. Methods Enzymol. 350:3–41. 33. Vermes, A., H. J. Guchelaar, and J. Dankert. 2000. Flucytosine: a review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 46:171–179.
34. Waldorf, A. R., and A. Polak. 1983. Mechanisms of action of 5-fluorocyto-sine. Antimicrob. Agents Chemother. 23:79–85.
35. Wang, J. S., S. Y. Li, Y. L. Yang, H. H. Chou, and H. J. Lo. 2007. Association between fluconazole susceptibility and genetic relatedness among Candida
tropicalis isolates in Taiwan. J. Med. Microbiol. 56:650–653.
36. Warnock, D. W. 2007. Trends in the epidemiology of invasive fungal infec-tions. Nippon Ishinkin Gakkai Zasshi 48:1–12.
37. White, T. C. 1997. The presence of an R467K amino acid substitution and loss of allelic variation correlate with an azole-resistant lanosterol 14alpha demethylase in Candida albicans. Antimicrob. Agents Chemother. 41:1488– 1494.
38. White, T. C., K. A. Marr, and R. A. Bowden. 1998. Clinical, cellular, and molecular factors that contribute to antifungal drug resistance. Clin. Micro-biol. Rev. 11:382–402.
39. Wu, J. M., et al. 2007. Solution structure of a novel D-naphthylalanine substituted peptide with potential antibacterial and antifungal activities. Biopolymers 88:738–745.
40. Xess, I., N. Jain, F. Hasan, P. Mandal, and U. Banerjee. 2007. Epidemiology of candidemia in a tertiary care centre of north India: 5-year study. Infection
35:256–259.
41. Yang, Y. L., et al. 2010. The distribution of species and susceptibility of amphotericin B and fluconazole of yeast pathogens isolated from sterile sites in Taiwan. Med. Mycol. 48:328–334.
42. Yang, Y. L., Y. A. Ho, H. H. Cheng, M. Ho, and H. J. Lo. 2004. Susceptibilities of Candida species to amphotericin B and fluconazole: the emergence of fluconazole resistance in Candida tropicalis. Infect. Control Hosp. Epide-miol. 25:60–64.
43. Yang, Y. L., S. Y. Li, H. H. Cheng, and H. J. Lo. 2005. Susceptibilities to amphotericin B and fluconazole of Candida species in TSARY 2002. Diagn. Microbiol. Infect. Dis. 51:179–183.
44. Yang, Y. L., and H. J. Lo. 2001. Mechanisms of antifungal agent resistance. J. Microbiol. Immunol. Infect. 34:79–86.
45. Yang, Y. L., et al. 2008. Susceptibilities to amphotericin B and fluconazole of