Contents lists available at
ScienceDirect
Journal of Ethnopharmacology
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / j e t h p h a r m
Toona sinensis (leaf extracts) inhibit vascular endothelial growth factor (VEGF)-induced angiogenesis in vascular endothelial cells
You-Cheng Hseu a , Ssu-Ching Chen b , Wen-Hsin Lin c , Dong-Zong Hung d , Ming-Kuem Lin e , Yueh-Hsiung Kuo e , f , g , Mei-Tsun Wang h , Hsin-Ju Cho h , Lai Wang h , Hsin-Ling Yang h ,∗
aDepartment of Cosmeceutics, College of Pharmacy, China Medical University, Taichung, Taiwan
bDepartment of Life Sciences, National Central University, Chung-Li, Taiwan
cSchool of Pharmacy, China Medical University, Taichung, Taiwan
dDivision of Toxicology, China Medical University Hospital, Taichung, Taiwan
eGraduate Institute of Chinese Pharmaceutical Sciences College of Pharmacy, China Medical University, Taichung, Taiwan
fAgricultural Biotechnology Research, Academia Sinica, Taipei, Taiwan
gDepartment of Chemistry, National Taiwan University, Taipei, Taiwan
hInstitute of Nutrition, China Medical University, Taichung, Taiwan
a r t i c l e i n f o
Article history:
Received 23 July 2010 Received in revised form 22 November 2010 Accepted 25 November 2010 Available online xxx
Keywords:
Traditional Chinese medicine Toona sinensis
Gallic acid Angiogenesis EA.hy 926 cells HUVECs
a b s t r a c t
Aim of the study: Toona sinensis is well known as a traditional Chinese medicine; also, it has been shown to exhibit anticancer and anti-inflammatory effects. This study was aimed at evaluating the anti-angiogenesis effect of the aqueous extracts of Toona sinensis (TS extracts) or gallic acid, a major component of TS extracts, against both VEGF-induced EA.hy 926 and human umbilical vein endothelial cells (HUVECs).
Materials and methods: Anti-proliferative activity of TS extracts or gallic acid, was determined against EA.hy 926 and HUVECs by trypan blue exclusion method. Invasion, tube formation and chick chorioal- lantoic membrane assay were carried out to determine the in vitro and in vivo anti-angiogenic effects.
Results: Non-cytotoxic concentration of TS extracts (50–100g/mL) and gallic acid (5 g/mL) inhibited the proliferation of VEGF-stimulated EA.hy 926 and HUVECs. Inhibitory effects of TS extracts and gallic acid on angiogenesis were assessed by VEGF-induced migration/invasion and capillary-like tube forma- tion by EA.hy 926 and HUVECs. Additionally, gelatin zymography assays showed that TS extracts and gallic acid suppressed the activity of metalloproteinase (MMP)-9 and MMP-2 activated by VEGF. In vivo, TS extracts and gallic acid strongly suppressed neovessel formation in the chorioallantoic membrane of chick embryos. Flow cytometry analyses and Western blot demonstrated that treatment with TS extracts and gallic acid induced G0/G1arrest in VEGF-stimulated EA.hy 926 cells via a reduction in the amounts of cyclin D1, cyclin E, CDK4, hyperphosphorylated retinoblastoma protein (pRb), VEGFR-2, and eNOS.
Conclusions: These results support an anti-angiogenic activity of Toona sinensis that may contribute critically to its cancer and inflammation chemopreventive potentials.
© 2010 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Toona sinensis Roem. (Meliaceae; Toona sinensis) is a type of arbor that is widely distributed in Asia. It has long been used as a traditional Chinese medicine for a wide variety of conditions in Chinese society and is very popular in vegetarian cuisine in Taiwan.
The edible leaves have been used as an oriental medicine for treating rheumatoid arthritis, cervicitis, urethritis, tympanitis, gas- tric ulcers, enteritis, dysentery, itchiness, and cancer (Xien, 1996;
Edmonds and Staniforth, 1998). While the underlying pharmaco-
∗ Corresponding author. Tel.: +886 4 22053366x7503; fax: +886 4 22062891.
E-mail address:[email protected](H.-L. Yang).
logical mechanisms of this new drug are still a matter of debate, various biological activities of Toona sinensis leaf extracts have been reported, including anti-cancer (Chang et al., 1998; Chang et al., 2002; Chang et al., 2006), anti-inflammatory (Yang et al., 2006), anti-diabetes (Yu, 2002; Hsu et al., 2003), and antioxidant (Cho et al., 2003; Hseu et al., 2008) effects, as well as inhibiting Leydig cell steroidogenesis (Poon et al., 2005) and improving the dynamic activity of human sperm (Yang, 2003). Moreover, the safety levels and nontoxic characteristics of aqueous extracts of Toona sinensis were evaluated using acute and sub-acute toxicity studies in mice (Liao et al., 2006).
Angiogenesis, the formation of new vessels from preexisting vasculature, is an essential process in a variety of physiologi- cal and pathological conditions, including inflammatory diseases,
0378-8741/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.jep.2010.11.058
and metastasis (Risau, 1997; Folkman, 2002). Complex sequential steps are involved in angiogenesis, such as basement membrane degradation by proteases, endothelial cell proliferation and migra- tion/invasion, formation of capillary tubes and survival of newly formed blood vessels (Bussolino et al., 1997). Angiogenesis critically depends on several conditions such as endothelial cell proliferation, endothelial cell secretion of matrix metalloproteinases (MMPs) required to break down surrounding tissue matrix and the endothe- lial cell movement/migration. Angiogenesis is tightly regulated by an intricate balance between stimulators and inhibitors (Bussolino et al., 1997). Among these, vascular endothelial growth factor (VEGF), a soluble angiogenic factor produced by many tumors as well as normal cell lines, plays a key role in regulating normal and pathologic angiogenesis (Ferrara and Davis-Smyth, 1997). VEGF contributes to the angiogenic process by stimulating proliferation, migration/invasion, and the formation of new blood vessels by endothelial cells (Ferrara and Davis-Smyth, 1997). Because of the critical dependence of human cancer and inflammatory diseases on angiogenesis, therapeutic strategies have been developed targeting various aspects of the angiogenic processes, many with promising results (Jackson et al., 1997; Folkman, 2002; Felmeden et al., 2003).
Many medicinal herbs have been shown to be rich sources of phytochemicals with chemoprevention potential for various types of human cancer and inflammatory diseases. Because tumor epithelial cells in vivo depend on angiogenesis to provide nutrients for their growth and survival, it is plausible that this anti- angiogenic effect may play a primary role in mediating the cancer chemopreventive activity (Folkman, 2002; Felmeden et al., 2003).
Moreover, the pharmacology of many anti-inflammatory drugs has revealed that at least part of their efficacy is attributable to their anti-angiogenic effects (Jackson et al., 1997). Hence, we hypothe- sized that the anticancer and anti-inflammatory activities of Toona sinensis could be partly attributable to its anti-angiogenic activity, and that Toona sinensis could be a promising candidate drug for the treatment of diseases with impaired angiogenesis. In this study, the ability of the aqueous leaf extracts of Toona sinensis (TS extracts) (50–100 g/mL) and gallic acid (3,4,5-trihydroxybenoic acid) (5 g/mL), a major component of TS extracts, to inhibit VEGF-induced angiogenesis in culture using EA.hy 926 cells and human umbilical vein endothelial cells (HUVECs) was investi- gated. The levels of angiogenic control and related molecules were assayed to determine the Toona sinensis-mediated anti-angiogenic mechanism.
2. Materials and methods
2.1. Chemicals
Dulbecco’s Modified Eagle’s medium (DMEM), fetal bovine serum (FBS), M-199 medium, glutamine, and penicillin–streptomycin–neomycin (GIBCO BRL, Grand Island, NY), anti-cyclin E antibody (Santa Cruz Biotechnology Inc., Heidel- berg, Germany), anti- -actin (Sigma Chemical Co., St. Louis, MO), anti-cyclin D1, anti-CDK4, anti-pRb, anti-VEGFR-2, and anti-eNOS antibodies (Cell Signaling Technology Inc., Danvers, MA) were obtained from their respective suppliers. All other chemicals were of the highest grade commercially available and supplied either by Merck (Darmstadt, Germany) or Sigma.
2.2. Toona sinensis preparation and extraction
The leaves of Toona sinensis were sourced from Fooyin Univer- sity, Kaohsiung Hsien, Taiwan. A voucher specimen (FY-001) was characterized by Dr. Horng-Liang Lay, Graduate Institute of Biotech- nology, National Pingtung University of Science and Technology,
Kaohsiung Hsien, Taiwan (Hsieh et al., 2004). The aqueous extracts of Toona sinensis were prepared by adding 1000 mL water to 1000 g fresh Toona sinensis leaves and boiling until it was reduced to 100 mL, as previously described (Hsu et al., 2003). The crude extracts were centrifuged at 3000 rpm for 12 min and the super- natant was used for this study. The crude extracts (50 g) were concentrated in a vacuum and freeze dried to form powder; the stock (2 mg/mL in PBS buffer [pH 7.4]) was subsequently stored at
−20
◦C until analyzed for anti-angiogenic properties. The crude TS extracts separated from fresh Toona sinensis leaves had a yield of 5%, which was based on the initial weight of the crude extracts (Yang et al., 2006).
2.3. Isolation of gallic acid from TS extracts
The TS extracts were dissolved in a mobile phase consist- ing of methanol–water (50:50, v/v) before high performance liquid chromatography (HPLC) analysis and separation. Eight com- pounds (gallic acid, methyl gallate, ethyl gallate, kaempferol, kaempferol-3-O- -d-glucoside, quercetin, quercitrin, quercetin-3- O- -d-glucoside and rutin) were isolated from the TS extracts, as previously described (Yang et al., 2006). The identity of the com- pounds was fully characterized by comparison of their spectral data (IR, NMR and mass) with the analogous information reported in the literature (Hsu et al., 2003; Yang et al., 2006). Gallic acid, the natu- ral phenolic component purified from the TS extracts, was collected for use in this study at a yield of 6% (Yang et al., 2006). The stock of gallic acid (2 mg/mL in PBS buffer [pH 7.4]) was subsequently stored at −20
◦C until analyzed for anti-angiogenic properties.
2.4. Endothelial cell culture
EA.hy 926 cells, a human vascular endothelial cell line (Edgell et al., 1983), was grown in DMEM supplemented with 15% FBS, HAT (100 mM sodium hypoxanthine, 0.4 mM aminopterin, and 16 mM thymidine), 1% glutamine, and 1%
penicillin–streptomycin–neomycin at 37
◦C in a 5% CO
2humidified incubator. This EA.hy 926 cell line was used because it possessed endothelial characteristics including the formation of tube-like structures (Bauer et al., 1992). The use of a cell line also allowed us to overcome the difficulty of obtaining larger numbers of uncon- taminated primary cells as well as the requirement of expensive growth factors associated with the use of primary endothelial cells.
In an effort to avoid interspecies variation, a number of reports
have used human umbilical vein endothelial cells (HUVECs) in
proliferation assays, migration/invasion assays, and tube forma-
tion assays, or as sources of metalloproteinases (MMPs) in gelatin
zymography assays. HUVECs were prepared from human umbili-
cal veins as described previously (Jaffe et al., 1973). In brief, the
umbilical cord was infused with 0.05% collagenase solution con-
taining the ECs and flushed from the cord by perfusion with cord
buffer and centrifuged. The resulting cell suspension was divided
equally between several 10-cm petri dishes, and grown to con-
fluence in M-199 medium and supplemented with 20% FBS at
37
◦C in 5% CO
2. Upon confluence, the cells of primary cultures
were detached using trypsin–EDTA, and sub-cultured to conflu-
ence in tissue culture wells at 37
◦C. EA.hy 926 (1 × 10
5cells/well)
or HUVECs (4 × 10
5cells/well) were grown to confluence on tis-
sue culture plates, pre-incubated with TS extracts (0, 50, 75, or
100 g/mL) or gallic acid (5 g/mL) for 1 h followed by incuba-
tion with or without VEGF (20 ng/mL) and allowed to proliferate for
24 h. Cultures were harvested and the cell number was determined
using a hemocytometer. Cell viability was checked before and after
treatment using trypan blue exclusion and examined using phase
contrast microscopy. For all experiments, the supernatant was
1 h, the cells were washed with PBS, and the culture media were replaced with new medium containing 20 ng/mL VEGF for the indi- cated time.
2.5. In vitro cell wound healing repair assay
To determine the effects of TS extracts or gallic acid on cell migration, an in vitro wound healing repair assay was performed.
EA.hy 926 (1 × 10
5cells/well) or HUVECs (4 × 10
5cells/well) were cultured on a 1% gelatin-coated 12-well plate and incubated with the indicated concentration of TS extracts or gallic acid for 1 h in 1% FBS-medium. At confluence, monolayers were wounded using a 200 L micropipette tip, washed twice with PBS and incubated for 12 or 24 h in 1% FBS medium with VEGF (20 ng/mL). Next, cells were washed twice with PBS, fixed with 100% methanol, and stained with Giemsa stain solution. The cultures were photographed (200 × mag- nification) to monitor the migration of cells into the wounded area, and the closure of wounded area was calculated.
2.6. Endothelial cell invasion assay
Invasion assays were performed using BD Matrigel invasion chambers (Bedford, MA, USA). For the invasion assay, 10 L Matrigel (25 mg/50 mL) was applied to 8- m polycarbonate mem- brane filters, and the bottom chamber of the apparatus contained standard medium. Matrigel is a solubilized basement membrane preparation extracted from the Engelbreth–Holm–Swarm mouse sarcoma, a tumor rich in extracellular matrix proteins. Briefly, the top chambers were seeded with EA.hy 926 cells (1 × 10
5cells/well) or HUVECs (4 × 10
5cells/well) in 500 L serum-free medium, and the cells were incubated with TS extracts (0, 50, 75, and 100 g/mL) or gallic acid (5 g/mL) for 1 h prior to the addition of 20 ng/mL VEGF. Cells were placed in the bottom chambers (750 L), which were filled with serum-free medium. Cells were allowed to migrate for 12 h or 24 h at 37
◦C. After the incubation period, non-migrated cells on the top surface of the membrane were removed with a cotton swab. The migrated cells on the bottom side of the membrane were fixed in cold 100% methanol for 8 min and washed twice with PBS. The cells were stained with Giemsa stain solution and then de-stained with PBS. Images were obtained using an optical microscope (200 × magnification); invad- ing cells were quantified by manual counting. Percent inhibition of invading cells was quantified and expressed with untreated cells (control) representing 100%.
2.7. Endothelial cell tube formation assay
To determine whether TS extracts or gallic acid affected the angiogenic process, tube formation was evaluated using the BD Bio- Coat Angiogenesis System: Endothelial Cell Tube Formation Assay kit (BD Biosciences, Bedford, MA). In brief, after a treatment with TS extracts (0, 50, 75, and 100 g/mL) or gallic acid (5 g/mL), cells were harvested and seeded in a BD Matrigel Matrix coated 96-well plates with EA.hy 926 cells (1 × 10
5cells/well) or HUVECs (4 × 10
5cells/well) in serum-free medium, for 30 min followed by incubating with or without VEGF (20 ng/mL) at 37
◦C. After 18 h, the capillary networks were photographed using a phase-contrast microscope, and a 200 × magnification; the number of tubes was quantified from three random fields (Ashton et al., 1999). The percent inhibition was expressed with untreated cells (control) representing 100%.
2.8. In vivo chick chorioallantoic membrane (CAM) assay
For the CAM assay (Kim et al., 2002), fertilized chick embryos were pre-incubated for 8 days at 37.5
◦C in 85% humidity. A hole
zone was identified in the CAM. A 1 cm × 1 cm window in the shell was sectioned to expose the CAM. Filter-paper disks were sterilized and loaded with TS extracts (0, 50, 75 and 100 g/mL) or gallic acid (5 g/mL) and applied to the CAM surface. Windows were sealed with clear tape and eggs were incubated for 48 h. Blood vessels were viewed and photographed. The anti-angiogenic effects of TS extracts or gallic acid on CAMs were quantified by counting the number of blood vessel branch points.
2.9. Determination of MMP-9 and -2 by zymography
The activities of MMP-9 and -2 released from cells were mea- sured by gelatin zymography protease assays. EA.hy 926 cells (1 × 10
5/well) or HUVECs (4 × 10
5/well) were seeded into 12-well culture dishes and grown in medium with 15% or 20% FBS to a nearly confluent monolayer. The cells were resuspended in medium, and then incubated with up to 100 g/mL TS extracts or 5 g/mL gal- lic acid 1 h prior to the addition of VEGF (20 ng/mL). After 24 h, collected media of an appropriate volume (adjusted by vital cell number) were prepared using SDS sample buffer, without boiling or reduction, and were subjected to 1 mg/mL gelatin–8% SDS-PAGE electrophoresis. After electrophoresis, gels were washed with 2.5%
Triton X-100 and then incubated in a reaction buffer (50 mM Tris- base [pH 7.5], 200 mM NaCl, 5 mM CaCl
2and 0.02% Brij 35) at 37
◦C for 24 h. Then, the gels were stained with Coomassie brilliant blue R-250.
2.10. Cell cycle distribution analysis (flow cytometric analysis)
Cellular DNA content was determined by flow cytometric analysis of propidium iodide (PI)-labeled cells. EA.hy 926 cells (5 × 10
5cells/60-mm dish) were incubated with TS extracts (0, 50, 75 and 100 g/mL) or gallic acid (5 g/mL) for 1 h, and then stimulated with VEGF (20 ng/mL) for 24 h. Cells were then col- lected by trypsinization, and fixed in ice-cold 70% ethanol at −20
◦C overnight. The cells were resuspended in PBS containing 1% Triton X-100, 0.5 mg/mL of RNase and 4 g/mL of PI at 37
◦C for 30 min.
A FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA) equipped with a single argon-ion laser (488 nm) was used for flow cytometric analysis. Forward and right angle light scatter, which correlated with the size of the cell and the cytoplasmic complexity, respectively, were used to establish size gates and exclude cellular debris from the analysis. The DNA content of 10,000 cells per analy- sis was monitored using the FACSCalibur system. The cell cycle was determined and analyzed using ModFit software (Verity Software House, Topsham, ME).
2.11. Preparation of cell extracts and immunoblot analysis
EA.hy 926 cells (5 × 10
5cells/60-mm dish) were incubated with various concentrations of TS extracts (0, 50, 75, and 100 g/mL) or gallic acid (5 g/mL) for 1 h prior to the addition of VEGF (20 ng/mL).
After 24 h, the cells were detached and washed once in cold PBS and suspended in 100 L lysis buffer (10 mM Tris–HCl [pH 8], 0.32 M sucrose, 1% Triton X-100, 5 mM EDTA, 2 mM DTT, and 1 mM phenylmethyl sulfonyl flouride). The suspension was put on ice for 20 min and then centrifuged at 16,000 × g for 20 min at 4
◦C.
Total protein content was determined using a Bio-Rad protein assay reagent, with bovine serum albumin as the standard, pro- tein extracts were reconstituted in sample buffer (0.062 M Tris–HCl [pH 6.8], 2% SDS, 10% glycerol and 5% -mercaptoethanol), and the mixture was boiled for 5 min. Equal amounts (50 g) of the dena- tured proteins were loaded into each lane, separated on 8–15%
SDS polyacrylamide gel, followed by transfer of the proteins to
PVDF membranes overnight. Membranes were blocked with 0.1%
for 20 min at room temperature, and the membranes were reacted with primary antibodies for 2 h. They were then incubated with a horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse antibody for 2 h before being developed using the SuperSignal ULTRA chemiluminescence substrate (Pierce, Rockford, IL). Band intensities were quantified by densitometry with absorbance of the mixture at 540 nm determined using an ELISA plate reader. West- ern blot analysis, with antibodies against cyclin D1, cyclin E, CDK4, pRb, VEGFR-2, and eNOS was performed as described previously (Hseu et al., 2008).
2.12. Statistical analysis
Experiments are presented as mean ± standard deviation (mean ± SD). All study data were analyzed using an analysis of variance (ANOVA), followed by Dunnett’s test for pair-wise com- parison. Statistical significance was defined as p < .05 for all tests.
3. Results
In this study, the human endothelial cell lines, EA.hy 926 cells and HUVECs, were used to investigate the ability of the aqueous leaf extracts of Toona sinensis (TS extracts) and the natural phenoic components purified from TS extracts, gallic acid, to inhibit angio- genesis, and to elaborate the molecular mechanisms involved in the inhibition. EA.hy 926 cells and HUVECs were incubated with the indicated concentrations of TS extracts or gallic acid for 1 h prior to the addition of 20 ng/mL VEGF.
3.1. Effects of TS extracts on the viability of EA.hy 926 cells or HUVECs
Angiogenesis involves the local proliferation of endothelial cells.
To determine if TS extracts or gallic acid altered cultured endothe- lial cells, the effect of either TS extracts or gallic acid on the viability of EA.hy 926 cells and HUVECs stimulated with or without VEGF was examined. After 24 h, both cell lines displayed a very high rate of proliferation in response to VEGF, as demonstrated by trypan blue exclusion (Fig. 1A). Exposure to 50–100 g/mL TS extracts or 5 g/mL gallic acid significantly (p < .05) inhibited VEGF-induced proliferation of EA.hy 926 cells or HUVECs (Fig. 1A). The concen- trations of TS extracts and gallic acid that were used in these studies were not cytotoxic to either EA.hy 926 cells or HUVECs without VEGF stimulation (Fig. 1B). No distinct cellular or mor- phological changes that are typically associated with apoptosis, such as cell detachment, rounding or chromosomal fragmentation, were detected after a 24 h incubation with TS extracts or gallic acid at a concentration below 100 g/mL or of 5 g/mL, respectively.
Therefore, for all subsequent experiments, we employed the non- cytotoxic concentration of TS extracts (i.e., ≤100 g/mL) and gallic acid (i.e., 5 g/mL), and focused on the effect of TS extracts or gallic acid on VEGF-induced angiogenesis.
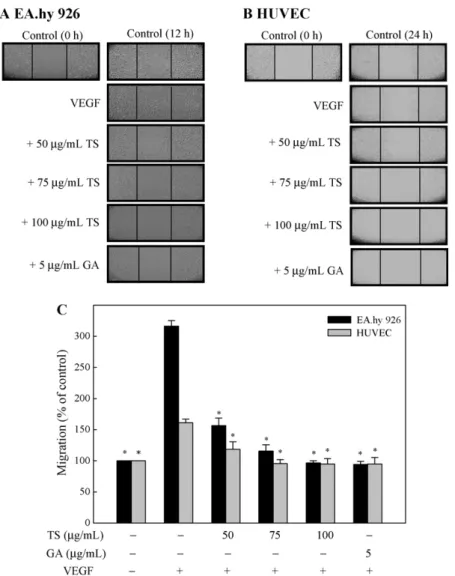
3.2. Effect of TS extracts on in vitro migration of EA.hy 926 cells or HUVECs
The migration of endothelial cells through the basement mem- brane is a crucial step in the establishment of new blood vessels (Bussolino et al., 1997). To determine the effects of TS extracts on endothelial cell migration in vitro, confluent monolayers of EA.hy 926 cells or HUVECs were wounded and cultured with control buffer plus 50–100 g/mL TS extracts or 5 g/mL gallic acid. As shown in Fig. 2A–C, treatment with VEGF significantly (p < .05) induced the migration of EA.hy 926 cells after 12 h and,
200
A
EA.hy 926HUVEC100 150
* *
* *
* * * *
50
Cell viability (% of control)
* *
0 TS (µg/mL)
VEGF GA (µg/mL)
−
−
− +
50 75 100
5
−
− +
− +
− +
− +
−
120
140 EA.hy 926
HUVEC
B
60 80 100
0 20 40
Cell viability (% of control)
− −
50 75 100−
TS (µg/mL)
GA (µg/mL)
− − − − −
5Fig. 1. Anti-proliferative activity of TS extracts and gallic acid purified from TS extracts. (A) EA.hy 926 cells and HUVECs were pre-treated with 0, 50, 75, or 100g/mL TS extracts or 5 g/mL gallic acid for 1 h followed by incubation with (A) or without (B) VEGF (20 ng/mL) and allowed to proliferate for 24 h. Cell num- bers were obtained by counting cell suspensions with a hemocytometer. Results are presented as mean± SD of three assays; *significant difference in comparison to the VEGF alone group, p < .05. GA, gallic acid.
in 24 h, HUVECs; the addition of TS extracts or gallic acid signifi- cantly (p < .05) decreased VEGF-induced migration of both EC cells (Fig. 2C).
3.3. Effects of TS extracts on the invasive potential of EA.hy 926 cells
Next, the effect of either TS extracts or gallic acid on the inva- siveness of EA.hy 926 cells or HUVECs was evaluated using the Boyden chamber assay, which allowed us to determine the ability of cells to pass through a layer of extracellular matrix on a Matrigel- coated filter. As shown in Fig. 3A–C, VEGF significantly induced the invasiveness of EA.hy 926 cells after 12 h and, after 24 h, the inva- sive potential of HUVECs; the addition of TS extracts or gallic acid (5 g/mL) significantly (p < .05) decreased VEGF-induced invasion of EA.hy 926 cells or HUVECs (Fig. 3C).
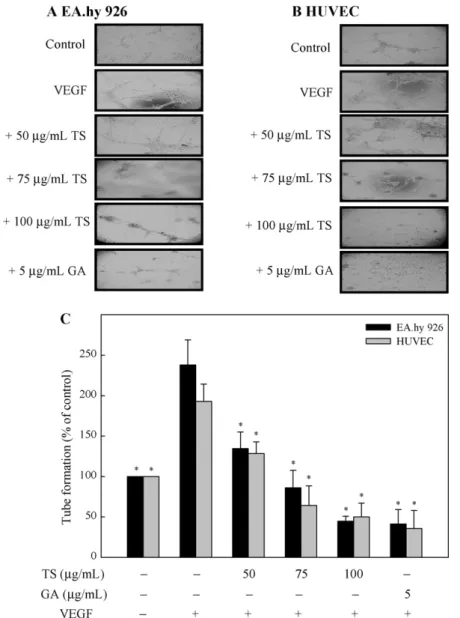
3.4. Effects of TS extracts on tube formation by EA.hy 926 cells or HUVECs
Since vascular maturation during angiogenesis is characterized
by the formation of tubular structures by capillary endothelial
cells, we performed a tube formation assay to investigate the
effect of exposure to TS extracts or gallic acid for 18 h on VEGF-
Fig. 2. TS extracts and gallic acid inhibit VEGF-induced migration of EA.hy 926 cells and HUVECs in an in vitro wound healing repair assay. Cells were pretreated with 0, 50, 75, or 100g/mL TS extracts or 5 g/mL gallic acid for 1 h. Subsequently, EA.hy 926 cells (A) or HUVECs (B) were scratched and then stimulated with VEGF (20 ng/mL) for 12 or 24 h. (C) Migration was observed using a phase-contrast microscope, at a 40× magnification, and the closure of area was calculated; *significant difference in comparison to VEGF alone group, p < .05. GA, gallic acid.
induced capillary-like structure formation by EA.hy 926 cells or HUVECs. Following stimulation by VEGF, EA.hy 926 cells or HUVECs became aligned into cords on the Matrigel, and a tube-like struc- ture was formed (Fig. 4A and B). Treatment of cells with either TS extracts or gallic acid resulted in a significant (p < .05) inhibition of VEGF-stimulated tube formation (Fig. 4A–C). The results clearly demonstrate that exposure to TS extracts or gallic acid is effective at controlling the VEGF-stimulated tube formation by endothelial cells.
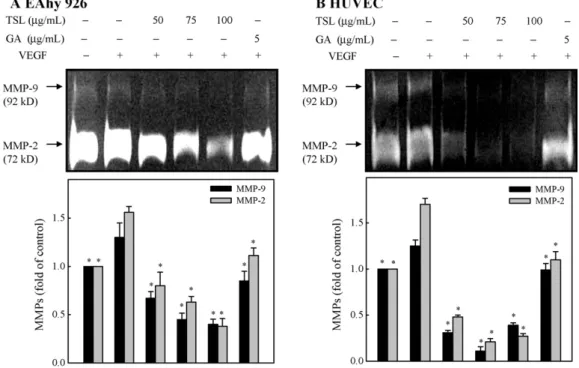
3.5. Effect of TS extracts on MMP-9 and MMP-2 activities in EA.hy 926 and HUVECs
Since gelatinase and collagenase MMPs are also involved in the angiogenic process (Egeblad and Werb, 2002), we used a gelatin zymography assay to determine the activity of secreted MMP-2 and MMP-9. To examine the effect of exposure to TS extracts or gallic acid on MMPs, EA.hy 926 cells or HUVECs were treated with TS extracts (50–100 g/mL) or gallic acid (5 g/mL) for 24 h in serum-free medium. The conditioned medium was collected and examined for MMP-9 and MMP-2 activity using gelatin zymogra- phy assays. Results from the zymography assay revealed that EA.hy 926 cells or HUVECs constitutively secreted MMP-9 and MMP-2 (Fig. 5A and B). VEGF treatment increased the level of protease
secretion, and treatment with TS extracts or gallic acid inhibited MMP-9 and MMP-2 secretion by these cells (Fig. 5A and B) (p < .05).
3.6. TS extracts inhibit angiogenesis in vivo
To determine if treatment with TS extracts or gallic acid could suppress blood vessel formation in vivo, we employed an in vivo angiogenesis model, the chick chorioallantoic membrane (CAM) assay. As shown in Fig. 6A and B, a filter paper coverslip containing TS extracts (50–100 g/mL) or gallic acid (5 g/mL) for 48 h effec- tively (p < .05) inhibited the formation of capillary vessels in the CAM compared with the coverslip containing PBS alone, which had no visible effect on the preexisting blood vessels. The results indi- cate that TS extracts or gallic acid is capable of inhibiting neovessel formation in vivo under natural conditions.
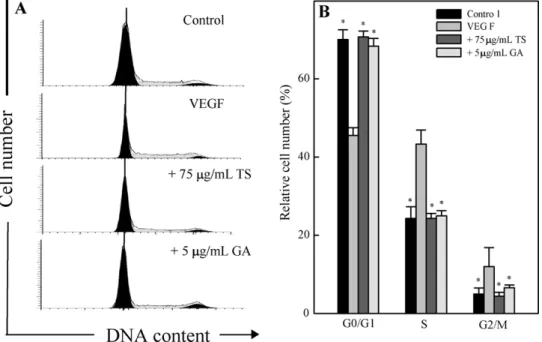
3.7. Effect of TS extracts on cell cycle in EA.hy 926 cells
To determine whether the anti-proliferative activity of TS
extracts or gallic acid involves cell cycle regulation, the profile of
the DNA content in TS extracts- or gallic acid-treated EA.hy 926
cells was evaluated using flow cytometry to measure the fluores-
cence of PI-DNA binding. The point during cell cycle progression at
which 75 g/mL TS extracts or 5 g/mL gallic acid induced growth
Fig. 3. TS extracts and gallic acid inhibit VEGF-induced invasion by EA.hy926 cells and HUVECs in a transwell assay. Cells were pretreated with 0, 50, 75, or 100g/mL TS extracts or 5g/mL gallic acid for 1 h and then stimulated with VEGF (20 ng/mL). Photomicrographs of EA.hy 926 cells (A) or HUVECs (B) invading under the membrane for 12 or 24 h. (C) The percentage inhibition of invading cells was quantified and expressed with untreated cells (control) representing 100%. Invasiveness was determined by counting cells in three microscopic fields per sample. Results are presented as mean± SD of three assays; *significant difference in comparison to VEGF alone group, p < .05.
GA, gallic acid.
inhibition in VEGF-stimulated EA.hy 926 was determined, with cellular distribution in the different phases of the treatment. Treat- ment of EA.hy 926 with VEGF for 24 h triggered the transition of cells from the G
0/G
1-phase to S-phase and pretreatment of cells with TS extracts or gallic acid significantly arrested cells in the G
0/G
1-phase (Fig. 7A and B) (p < .05). Additionally, the percent- age of cells in the G
0/G
1-phase increased, while those in the S- and G
2/M phases decreased, after treatment with TS extracts or gallic acid (Fig. 7A and B) (p < .05). Our findings suggested that both TS extracts and gallic acid specifically induced G
0/G
1arrest of VEGF-stimulated EA.hy 926 cells at a nontoxic dose; this may be responsible for the inhibition of VEGF-stimulated EA.hy 926 cell proliferation.
3.8. Effect of TS extracts on the G
0/G
1-related protein expression
In order to examine the molecular mechanism(s) and underly- ing changes in cell cycle patterns, we investigated the effects of G
0/G
1(cell cycle)-related proteins of EA.hy 926 cells activated by VEGF. We treated EA.hy 926 cells with TS extracts (75 g/mL) or gallic acid (5 g/mL) for 24 h. Western blot analysis revealed that, in VEGF-stimulated EA.hy 926 cells, reductions in cyclin D1, cyclin
E, CDK4, and hyperphosphorylated retinoblastoma protein (pRb) occurred after treatment with TS extracts, and cyclin E expression decreased in response to gallic acid (Fig. 8A). These results implied that the G
0/G
1arrest of VEGF-stimulated EA.hy 926 cells exposed to TS extracts was associated with reduced amounts of cyclin D1, cyclin E, CDK4, and pRb, and with reduced amounts of cyclin E in gallic acid-treated cells.
Additionally, because VEGF-induced activation of EA.hy 926 cells is mediated by the binding of VEGF to VEGFRs (Rahimi et al., 2000), the expression of membrane VEGFR-2 was examined by Western blot analysis. As shown in Fig. 8B, both TS extracts (75 g/mL) and gallic acid (5 g/mL) decreased the expression of membrane VEGFR-2 protein in VEGF-stimulated EA.hy 926 cells.
Furthermore, VEGF is an important mediator of NO and eNOS cat-
alyzed NO production, and this is believed to occur through the
VEGFR-2 receptor (Shizukuda et al., 1999). Thus, we investigated
the TS extracts or gallic acid-mediated eNOS protein levels in cul-
tured EA.hy 926 cells. Western blot analysis of eNOS exposed to TS
extracts (75 g/mL) or gallic acid (5 g/mL) for 24 h was resolved
on SDS-PAGE. Incubation of cells with VEGF increased eNOS pro-
tein levels and addition of TS extracts or gallic acid inhibits the
expression of eNOS induced by VEGF (Fig. 8B).
Fig. 4. TS extracts and gallic acid inhibit VEGF-induced tube formation by EA.hy 926 cells and HUVECs. Cells were pre-treated with 0, 50, 75, or 100g/mL TS extracts or 5g/mL gallic acid for 1 h. EA.hy 926 cells (A) and HUVECs (B) were then collected and replated on Matrigel-coated plates at a density of 1 × 105or 4× 105cells/well and incubated in the absence (control) or presence of VEGF (20 ng/mL). After 18 h, the presence or absence of tube formation was determined using a phase-contrast microscope (40×). (C) The capillary networks were photographed using a phase-contrast microscope, and the number of tubes was quantified from three random fields. Results are presented as mean± SD of three assays; *significant difference in comparison to VEGF alone group, p < .05. GA, gallic acid.
4. Discussion
A lot of herbs used in traditional Chinese medicine may be of great treasure. Recent studies have identified a number of phar- macological and toxicological properties of Toona sinensis. In the present study, we demonstrate that TS extracts or gallic acid, a major component of TS extracts at a yield of 6% from the total extracts, potentially inhibits the proliferation, migration/invasion and tube formation, an in vitro marker of angiogenesis, by VEGF- activated vascular endothelial cells, both EA.hy 926 cells and HUVECs. Moreover, we show that TS extracts or gallic acid exerts a potent anti-angiogenic effect in vivo, as verified from its inhi- bition of naturally induced neovascularization by capillary vessel formation in the CAM. To our knowledge, this is the first scientific demonstrating the inhibitory effect of Toona sinensis leaf extracts on vascular endothelial angiogenesis activated by growth factor VEGF.
Matrix MMPs play a major regulatory role in matrix reorgani- zation and are clearly implicated in the initiation of angiogenesis and endothelial cell differentiation and spreading during angiogen- esis (Kahari and Saarialho-Kere, 1999; Egeblad and Werb, 2002). In
particular, the release of MMP-2 and MMP-9 from endothelial cells
represents an important step in neovascularization, because this
major extracellular matrix proteolytic enzyme is secreted when
endothelial sprouting takes place, thus enhancing cell migration
across the extracellular matrix and tube-like structure formation
(Mignatti and Rifkin, 1993). Further, MMP-2 and MMP-9 are both
type IV collagenases that have been shown to be important in tumor
migration/invasion in vitro because of their ability to break down
basement membrane components, in particular collagen IV (Sato
et al., 1994; Waas et al., 2002). It has been shown that a reduc-
tion in tube formation by endothelial cells was associated with
a decrease in gelatinolytic activities of both MMP-2 and MMP-
9, whereas an enhancement of activity increased tube formation
(Egeblad and Werb, 2002). The importance of MMPs in cancer, in
particular the contribution of MMP-2 and MMP-9 to cancer metas-
tasis and angiogenesis, promoted the development of inhibitors
capable of targeting gelatinase activity in tumors (Foda and Zucker,
2001). Our results demonstrate that TS extracts or gallic acid inhib-
ited VEGF-stimulated activities of MMP-2 and MMP-9. Because
increased MMP activity is closely associated with the angiogenic
Fig. 5. TS extracts and gallic acid inhibit the activity of MMP-9 and MMP-2, respectively, in VEGF-stimulated EA.hy 926 cells (A) and HUVECs (B). Cells were pretreated with 0, 50, 75, or 100g/mL TS extracts or 5 g/mL gallic acid for 1 h and then stimulated with VEGF (20 ng/mL) for 24 h. The conditioned media and cells were subjected to gelatin zymography to analyze the activities of MMP-9 and MMP-2. Determined activities of these proteins were subsequently quantified by densitometric analysis. Representative results from three independent experiments are shown. Results are presented as mean± SD of three assays; *significant difference in comparison to VEGF alone group, p < .05.
GA, gallic acid.
pathway, the inhibition of migration/invasion and tube formation by TS extracts or gallic acid may also be due to decreased MMP activity.
A critical step in angiogenesis involves the local proliferation of endothelial cells (Risau, 1997). VEGF mediates the mitogenic activities of EA.hy 926 cells and HUVECs above the basal level. TS extracts or gallic acid effectively inhibits proliferation of EA.hy 926 cells and HUVECs. Since TS extracts or gallic acid do not have a cytotoxic effect at up to a concentration of 100 g/mL or 5 g/mL, respectively, the anti-proliferative effect may not be due to apop- tosis or necrosis of endothelial cells, but rather to the inhibition of cell cycle progression induced by VEGF. To confirm this the- ory, we examined the effect of TS extracts or gallic acid on the cell cycle progression of EA.hy 926 cells stimulated by VEGF. Accu- mulating evidence indicates that cyclin is an essential component of the cyclin-dependent kinase (CDK) complex and plays a cru- cial role in regulating the activity of CDK (Sherr, 1996). Cyclin D1 and E are critical mediators of the progression of endothelial cells through the G
1/S phase of the cell cycle, forming cyclin D1-CDK4 and cyclin E-CDK2 complex that phosphorylates and inactivates the retinoblastoma (Rb) protein, thereby releasing E2F to mediate the G
1to S transition (Sherr, 1995). D-type cyclins, like cyclin D1, are active in mid-G
1phase, whereas E-type cyclins, like cyclin E, are active in late-G
1phase prior to the transition of cell cycle from G
1to S phase (Sherr, 1996). Thus, TS extracts specifically inhibit the G
0/G
1arrest of VEGF-stimulated EA.hy 926 cells through downreg- ulation of cyclin D1, cyclin E, and CDK4 expression and blocking Rb phosphorylation, which may be responsible for the inhibition of EA.hy 926 cell proliferation. On the contrary, our results indi- cate that gallic acid treatment of VEGF-stimulated EA.hy 926 cells reduced the amounts of cyclin E. Therefore, it is possible that the antiproliferative effect of gallic acid on VEGF-stimulated endothe- lial cells is accomplished by arresting cells at the G
0/G
1phase of the cell cycle by suppressing cyclin E expression.
VEGF is one of the most potent and extensively studied angio- genic stimulators (Ferrara and Davis-Smyth, 1997). After being
released, VEGF binds to the VEGFR on the endothelial cell mem- brane and initiates a cascade of signals for vessel formation (Hicklin and Ellis, 2005). Binding of VEGF to VEGFR-2, one of the VEGFRs involved in angiogenic responses, leads to receptor autophospho- rylation and then to a cascade of angiogenic signals (Hicklin and Ellis, 2005). VEGFR-2 is a receptor tyrosine kinase composed of seven extracellular immunoglobulin domains, a transmembrane domain and a tyrosine kinase active cytoplasmic domain (de Vries et al., 1992; Terman et al., 1992). It is generally accepted that activation of the VEGFR-2, which is relatively specific for vascu- lar endothelial cells, promotes migration and, during angiogenesis (Rahimi et al., 2000). Thus, in culture studies, VEGFR-2 knockout cells block VEGF stimulation of cell migration and proliferation (Koolwijk et al., 2001). The results are consistent with the finding that inhibition of VEGFR-2 expression decreases proliferation and migration/invasion by EA.hy 926 cells, suggesting that TS extracts or gallic acid induced a decrease in VEGFR-2, leading to the inhibi- tion of endothelial cell angiogenesis.
The earliest stages of angiogenesis are defined by vasodilation mediated by NO and increased vascular permeability of pre- existing capillaries or post-capillary venules in response to VEGF (Fukumura et al., 2001). Furthermore, VEGF is an important media- tor of NO production and eNOS-catalyzed NO production, and this is believed to occur through the VEGFR-2 receptor (Shizukuda et al., 1999 ). NOS converts l-arginine to citrulline and NO in the presence of oxygen and NADPH. NOS can occur in one of three isoforms:
neuronal NOS (nNOS), inducible NOS (iNOS) and endothelial NOS (eNOS). Vasodilation by smooth muscle relaxation, mediated by NO, is a prerequisite for the endothelial cell to enter the angio- genic cascade and increased NO production correlates positively with increase of vascular density and tumor growth (Fukumura et al., 2001). Because of its prime role in angiogenesis, the inhibi- tion of the expression of eNOS by TS extracts or gallic acid resulted in significant reduction of angiogenesis.
In our previous study, a number of compounds, including gallic
acid, methyl gallate, ethyl gallate, kaempferol, kaempferol-3-O-
Fig. 6. TS extracts and gallic acid inhibit angiogenesis in vivo. A filter-paper disk with 0, 50, 75, or 100g/mL TS extracts or 5 g/mL gallic acid was placed in to chick CAMs.
After 48 h, CAMs were peeled off and photographed; blood vessels were viewed and photographed. (A) Representative photographs of chick CAM assays. (B) Quantitative analysis of neovascularization from the photographs. Results are presented as mean± SD of three assays; *significant difference in comparison to control group, p < .05. GA, gallic acid.
-d-glucoside, quercetin, quercitrin, quercetin-3-O--d-glucoside, and rutin, were isolated from the leaves of Toona sinensis; iden- tity of the compounds was determined by HPLC and based on the analogous information reported in the literature (Hsu et al., 2003;
Yang et al., 2006). The total phenolic content of the TS extract was estimated to be 130 ± 26 mg of gallic acid equivalent/g of plant extracts (Yang et al., 2006). The yield of gallic acid, the natu- ral phenolic component purified from TS extracts, was about 6%.
Although it remains unclear which of the components of Toona sinensis are active compounds, gallic acid has received increased attention recently because of some interesting new findings regard- ing its biological activities (Chen et al., 2009). Gallic acid is widely distributed in various plants and fruits, such as gallnuts, sumac, oak bark, green tea, apple peels, grapes, strawberries, pineapples, bananas, lemons and in red and white wine (Sun et al., 2002; Beer et al., 2003; Wolfe et al., 2003). It is a polyhydroxyphenolic com- pound and is one of the major bioactive compounds isolated and purified from Toona sinensis. Further, gallic acid was elucidated as one of the active angiogenesis inhibitors in a human (tissue- based fibrin–thrombin clot) angiogenesis assay (Liu et al., 2006).
Even though the therapeutic utility of gallic acid in this regard is unknown, its common occurrence in fruits and food as well as its
small molecular weight (170 Da) might be an advantage in terms of safety and dosing design. Studies have demonstrated that gallic acid selectively induces cancer cell death by apoptosis; however, gallic acid shows no cytotoxicity against normal cells (Inoue et al., 1994; Inoue et al., 1995; Isuzugawa et al., 2001). Therefore, gallic acid may be a useful phytochemical for cancer chemoprevention (Surh, 2003). These results imply that gallic acid is one of the active compounds responsible for the antiangiogenic activities of Toona sinensis leaf extracts. Moreover, in future we have planned to inves- tigate antiangiogenic effect of other bioactive compounds isolated from the aqueous leaf extracts of Toona sinensis.
In conclusion, our observations indicate that aqueous leaf
extracts of Toona sinensis exert an inhibitory effect on several
essential steps of angiogenesis, including proliferation, migra-
tion/invasion, and tube formation of vascular endothelial cells
in vitro, and on neo-angiogenesis of the chick CAM in vivo. In addi-
tion, Toona sinensis could regulate the activities of migration and
invasion-associated proteinases (MMP-2 and MMP-9); it could also
be effective at suppressing the proliferation of endothelial cells
as shown by cell cycle arrest (G
0/G
1) and expression cell cycle-
related proteins. The anti-angiogenic activity reported in this paper
supports the merit of further investigations to assess and define
Fig. 7. TS extracts and gallic acid induced VEGF-stimulated cell cycle (G0/G1) arrest in EA.hy 926 cells. Cells were pretreated without or with 75g/mL TS extracts or 5 g/mL gallic acid for 1 h and then stimulated with VEGF (20 ng/mL) for 24 h, stained with PI, and analyzed for cell cycle phase using flow cytometry. (A) Representative flow cytometry patterns are shown. (B) Cellular distribution (percentage) in different phases of the cell cycle (G0/G1, G1, S and G2/M) after TS extract and gallic acid treatment is shown.
Results are presented as mean± SD of three assays; *significant difference in comparison to VEGF alone group, p < .05. GA, gallic acid.
Fig. 8. Effects of TS extracts and gallic acid on VEGF-induced expression of cyclin D1, cyclin E, CDK4, pRb (A), VEGFR2 and eNOS (B) in EA.hy 926 cells. Cells were pretreated without or with 75g/mL TS extracts or 5 g/mL gallic acid for 1 h and then stimulated with VEGF (20 ng/mL) for 24 h. Protein (50 g) from each sample was resolved on a 8-15% SDS-PAGE and Western blotting was performed.-Actin was used as a control. Relative changes in protein bands were measured using densitometric analysis.
Representative results from three independent experiments are shown. GA, gallic acid.
its cancer and inflammation chemopreventive and/or therapeutic potential for humans.
Acknowledgements
This work was supported by grants NSC-95-2320-B-039-007, NSC-96-2320-B-039-002, NSC-97-2320-B-039-042 and CMU-99- C-02 from the National Science and China Medical University, Taiwan, ROC. This study was also supported in part by Taiwan Department of Health Clinical Trial and Research Center of Excel- lence (DOH99-TD-B-111-004) and Cancer Research Center of Excellence (DOH99-TD-C-111-005), Taiwan, ROC.
References
Ashton, A.W., Yokota, R., John, G., Zhao, S., Suadicani, S.O., Spray, D.C., Ware, J.A., 1999. Inhibition of endothelial cell migration, intercellular communication, and vascular tube formation by thromboxane A2. The Journal of Biological Chemistry 274, 35562–35570.
Bauer, J., Margolis, M., Schreiner, C., Edgell, C.J., Azizkhan, J., Lazarowski, E., Juliano, R.L., 1992. In vitro model of angiogenesis using a human endothelium-derived permanent cell line: contributions of induced gene expression, G-proteins, and integrins. Journal of Cellular Physiology 153, 437–449.
Beer, D.D., Joubert, E., Gelderblom, W.C., Manley, M., 2003. Antioxidant activity of South African red and white cultivar wines: free radical scavenging. Journal of Agricultural and Food Chemistry 51, 902–909.
Bussolino, F., Mantovani, A., Persico, G., 1997. Molecular mechanisms of blood vessel formation. Trends in Biochemical Sciences 22, 251–256.
nese herbal prescriptions on copulatory activity in aged male rats: a preliminary study. The American Journal of Chinese Medicine 26, 83–90.
Chang, H.C., Hung, W.C., Huang, M.S., Hsu, H.K., 2002. Extract from the leaves of Toona sinensis Roemor exerts potent antiproliferative effect on human lung cancer cells. The American Journal of Chinese Medicine 30, 307–314.
Chang, H.L., Hsu, H.K., Su, J.H., Wang, P.H., Chun, Y.F., Chia, Y.C., Tsai, L.Y., Wu, Y.C., Yuan, S.S., 2006. The fractionated Toona sinensis leaf extract induces apoptosis of human ovarian cancer cells and inhibits tumor growth in a murine xenograft model. Gynecologic Oncology 102, 309–314.
Chen, H.M., Wua, Y.C., Chia, Y.C., Chang, F.R., Hsu, H.K., Hsieh, Y.C., Chen, C.H., Yuan, S.S., 2009. Gallic acid, a major component of Toona sinensis leaf extracts, con- tains a ROS-mediated anti-cancer activity in human prostate cancer cells. Cancer Letters 286, 161–171.
Cho, E.J., Yokozawa, T., Rhyu, D.Y., Kim, H.Y., Shibahara, N., Park, J.C., 2003. The inhibitory effects of 12 medicinal plants and their component compounds on lipid peroxidation. The American Journal of Chinese Medicine 31, 907–917.
de Vries, C., Escobedo, J.A., Ueno, H., Houck, K., Ferrara, N., Williams, L.T., 1992.
The fms-like tyrosine kinase, a receptor for vascular endothelial growth factor.
Science 255, 989–991.
Edmonds, J.M., Staniforth, M., 1998. Toona sinensis: Meliaceae. Curtis’s Botanical Magazine 15, 186–193.
Egeblad, M., Werb, Z., 2002. New functions for the matrix metalloproteinases in cancer progression. Nature 2, 161–174.
Edgell, C.J., McDonald, C.C., Graham, J.B., 1983. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proceedings of the National Academy of Sciences of the United States of America 80, 3734–3737.
Felmeden, D.C., Blann, A.D., Lip, G.Y.H., 2003. Angiogenesis: basic pathophysiology and implications for disease. European Heart Journal 24, 586–603.
Ferrara, N., Davis-Smyth, T., 1997. The biology of vascular endothelial growth factor.
Endocrine Reviews 18, 4–25.
Foda, H.D., Zucker, S., 2001. Matrix metalloproteinases in cancer invasion, metastasis and angiogenesis. Drug Discovery Today 6, 478–482.
Folkman, J., 2002. Role of angiogenesis in tumor growth and metastasis. Seminars in Oncology 29, 15–18.
Fukumura, D., Gohongi, T., Kadambi, A., Izumi, Y., Ang, J., Yun, C.O., Buerk, D.G., Huang, P.L., Jain, R.K., 2001. Predominant role of endothelial nitric oxide synthase in vascular endothelial growth factor-induced angiogenesis and vascular perme- ability. Proceedings of the National Academy of Sciences of the United States of America 98, 2604–2609.
Hicklin, D.J., Ellis, L.M., 2005. Role of the vascular endothelial growth factor pathway in tumor growth and angiogenesis. Journal of Clinical Oncology 23, 1011–1027.
Hseu, Y.C., Chang, W.H., Chen, C.S., Liao, J.W., Huang, C.J., Lu, F.J., Chia, Y.C., Hsu, H.K., Wu, J.J., Yang, H.L., 2008. Antioxidant activities of Toona sinensis leaves extracts using different antioxidant models. Food and Chemical Toxicology 46, 105–114.
Hsieh, T.J., Liu, T.Z., Chia, Y.C., Chern, C.L., Lu, F.J., Chuang, M.C., Mau, S.Y., Chen, S.H., Syu, Y.H., Chen, C.H., 2004. Protective effect of methyl gallate from Toona sinensis (Meliaceae) against hydrogen peroxide-induced oxidative stress and DNA damage in MDCK cells. Food and Chemical Toxicology 42, 843–850.
Hsu, H.K., Yang, Y.C., Hwang, J.H., Hong, S.J., 2003. Effects of Toona sinensis leaf extract on lipolysis in differentiated 3T3-L1 adipocytes. The Kaohsiung Journal of Medical Sciences 19, 3855–3890.
Isuzugawa, K., Inoue, M., Ogihara, Y., 2001. Catalase contents in cells determine sen- sitivity to the apoptosis inducer gallic acid. Biological & Pharmaceutical Bulletin 24, 1022–1026.
Inoue, M., Suzuki, R., Koide, T., Sakaguchi, N., Ogihara, Y., Yabu, Y., 1994. Antioxidant, gallic acid, induces apoptosis in HL-60RG cells. Biochemical and Biophysical Research Communications 204, 898–904.
Inoue, M., Suzuki, R., Sakaguchi, N., Li, Z., Takeda, T., Ogihara, Y., Jiang, B.Y., Chen, Y., 1995. Selective induction of cell death in cancer cells by gallic acid. Biological &
Pharmaceutical Bulletin 18, 1526–1530.
Jackson, J.R., Seed, M.P., Kircher, C.H., Willoughby, D.A., Winkler, J.D., 1997. The codependence of angiogenesis and chronic inflammation. The FASEB Journal 11, 457–465.
lial cells derived from umbilical vein. The Journal of Clinical Investigation 52, 2745–2756.
Kahari, V.M., Saarialho-Kere, U., 1999. Matrix metalloproteinases and their inhibitors in tumour growth and invasion. Annals of Medicine 31, 34–45.
Kim, C.W., Lee, H.M., Lee, T.H., Kang, C., Kleinman, H.K., Gho, Y.S., 2002. Extracellular membrane vesicles from tumor cells promote angiogenesis via sphingomyelin.
Cancer Research 62, 6312–6317.
Koolwijk, P., Peters, E., van der Vecht, B., Hornig, C., Weich, H.A., Alitalo, K., Hick- lin, D.J., Wu, Y., Witte, L., van Hinsbergh, V.W., 2001. Involvement of VEGFR-2 (kdr/flk-1) but not VEGFR-1 (flt-1) in VEGF-A and VEGF-C induced tube for- mation by human microvascular endothelial cells in fibrin matrices in vitro.
Angiogenesis 4, 53–60.
Liao, J.W., Hsu, C.K., Wang, M.F., Hsu, W.M., Chan, Y.C., 2006. Beneficial effect of Toona sinensis Roemor on improving cognitive performance and brain degen- eration in senescence-accelerated mice. The British Journal of Nutrition 96, 400–407.
Liu, Z., Schwimer, J., Liu, D., Lewis, J., Greenway, F.L., York, D.A., Woltering, E.A., 2006.
Gallic acid is partially responsible for the antiangiogenic activities of Rubus leaf extract. Phytotherapy Research 20, 806–813.
Mignatti, P., Rifkin, D.B., 1993. Biology and biochemistry of proteinases in tumor invasion. Physiological Reviews 73, 161–195.
Poon, S.L., Leu, S.F., Hsu, H.K., Liu, M.Y., Huang, B.M., 2005. Regulatory mecha- nism of Toona sinensis on mouse Leydig cell steroidogenesis. Life Sciences 76, 1473–1487.
Rahimi, N., Dayanir, V., Lashkari, K., 2000. Receptor chimeras indicate that the vascular endothelial growth factor receptor-1 (VEGFR-1) modulates mitogenic activity of VEGFR-2 in endothelial cells. The Journal of Biological Chemistry 275, 16986–16992.
Risau, W., 1997. Mechanisms of angiogenesis. Nature 386, 671–674.
Sato, H., Takino, T., Okada, Y., Cao, J., Shinagawa, A., Yamamoto, E., Seiki, M., 1994.
A matrix metalloproteinase expressed on the surface of invasive tumour cells.
Nature 370, 61–65.
Sherr, C.J., 1995. D-type cyclins. Trends in Biochemical Sciences 20, 187–190.
Sherr, C.J., 1996. Cancer cell cycles. Science 274, 1672–1677.
Shizukuda, Y., Tang, S., Yokota, R., Ware, J.A., 1999. Vascular endothelial growth factor-induced endothelial cell migration and proliferation depend on a nitric oxide-mediated decrease in protein kinase C delta activity. Circulation Research 85, 247–256.
Sun, J., Chu, Y.F., Wu, X., Liu, R.H., 2002. Antioxidant and antiproliferative activities of common fruits. Journal of Agricultural and Food Chemistry 50, 7449–7454.
Surh, Y.J., 2003. Cancer chemoprevention with dietary phytochemicals. Nature Reviews. Cancer 3, 768–780.
Terman, B.I., Dougher-Vermazen, M., Carrion, M.E., Dimitrov, D., Armellino, D.C., Gospodarowicz, D., Bohlen, P., 1992. Identification of the KDR tyrosine kinase as a receptor for vascular endothelial cell growth factor. Biochemical and Biophysical Research Communications 187, 1579–1586.
Waas, E.T., Lomme, R.M.L.M., DeGroot, J., Wobbes, T.H., Hendriks, T., 2002. Tissue levels of active matrix metalloproteinase-2 and -9 in colorectal cancer. British Journal of Cancer 86, 1876–1883.
Wolfe, K., Wu, X., Liu, R.H., 2003. Antioxidant activity of apple peels. Journal of Agricultural and Food Chemistry 51, 609–614.
Xien, W.C., 1996. Assembly of Chinese Herbs in Mainland, vol. 2. People’s Medical Publishing House, Beijing, pp. 459–460.
Yang, H.L., Chang, W.H., Chia, Y.C., Huang, C.J., Lu, F.J., Hsu, H.K., Hseu, Y.C., 2006. Toona sinensis extracts induces apoptosis via reactive oxygen species in human premyelocytic leukemia cells. Food and Chemical Toxicology 44, 1978–1988.
Yang, F.Y., 2003. Study on the physiological function of Toona sinensis Roem in human spermatozoa. Master Thesis in National Cheng Kung University, Tainan, Taiwan.
Yu, J.Y.L., 2002. Toona sinensis extract affects gene expression of GLUT4 GLU00 se transporter in adipose tissue of alloxan-induced diabetic rats. In: Pro- ceedings of 5th Congress of the International Diabetic Society, China, May, pp. 4–7.