科技部補助專題研究計畫報告
介白素二十九在肥胖誘導之發炎及胰島素抗性中的調控角色
報 告 類 別 : 成果報告 計 畫 類 別 : 個別型計畫 計 畫 編 號 : MOST 108-2320-B-006-052-執 行 期 間 : 108年08月01日至109年08月10日 執 行 單 位 : 國立成功大學臨床醫學研究所 計 畫 主 持 人 : 許育祥 計畫參與人員: 碩士級-專任助理:林恬毓 碩士班研究生-兼任助理:張育銜 碩士班研究生-兼任助理:吳乙柔本研究具有政策應用參考價值:■否 □是,建議提供機關
(勾選「是」者,請列舉建議可提供施政參考之業務主管機關)
本研究具影響公共利益之重大發現:□否 □是
中 華 民 國 109 年 10 月 27 日
中 文 摘 要 : 肥胖是一種慢性發炎疾病,與胰島素抗性、心血管疾病以及非酒精 性脂肪肝疾病的發生有密切關係。生活習慣、基因遺傳、飲食與代 謝失調都可能造成肥胖。在肥胖發展過程中,免疫細胞會浸潤到脂 肪組織中,其中以單核細胞以及巨噬細胞為主要的族群,這些細胞 會分泌許多促發炎細胞激素,包括TNF-α、 IL-1β、 IL-6和IL-8,進而引起脂肪組織的慢性發炎反應。因此在肥胖的發炎微環境中 ,脂肪細胞與巨噬細胞間之交互作用是十分關鍵。介白素二十九 (Interleukin-29, IL-29)是第三型干擾素(interferon)家族的一員 ,透過IL-28R1和IL-10R2這對接受器來傳遞訊號啟動生物功能。IL-29在宿主對抗微生物入侵中具有重要作用,在感染病毒的細胞中其 表現量顯著上升。過去文獻得知巨噬細胞、樹突細胞、週邊血液單 核細胞都會分泌IL-29。然而,IL-29是否參與在肥胖的慢性發炎中 是未知的。在這項研究中,我們收集了肥胖患者的脂肪組織和血液 ,並根據其BMI數值對其進行了分類。我們也使用SGBS細胞來釐清分 化過後的細胞是否具有IL-28RA和IL-10R2。首先,我們發現肥胖患 者的血清中可偵測高量的IL-29,且在肥胖患者的脂肪組織會高度表 現IL-29。在人類脂肪細胞株(SGBS cells)的成熟過程中會大量的表 達IL-28R1和IL-10R2這對接受器,因此可作為IL-29的目標細胞。為 了進一步研究IL-29的生物功能,我們成功表達並純化人類IL-29蛋 白質。同時將IL-29當成抗原來建立小鼠單株抗體,我們也成功建立 並篩選專一性可拮抗IL-29的單株抗體,以利於後續研究計畫的執行 。 中 文 關 鍵 詞 : 介白素二十九,脂肪細胞,發炎
英 文 摘 要 : Obesity is a chronic inflammatory disease that is closely related to the development of insulin resistance,
cardiovascular disease and nonalcoholic fatty liver disease. In the progression of obesity, immune cells are infiltrated into adipose tissue. Most are monocytes and macrophages. These cells secrete various inflammatory factors, including TNF-α, IL-1β, IL-6, and IL-8 and then cause chronic inflammation of adipose tissue. Therefore, adipocytes and macrophages interaction play a critical role in the inflammatory environment of obesity. Interleukin-29 (IL-29), a member of type 3 interferon family, signals through a heterodimer receptor complex of 28R1 and IL-10R2 to initiate biological functions. IL-29 plays an
important role in the host against microbes and its gene is highly upregulated in cells infected with viruses. Several studies reported that macrophages, dendritic cells, and peripheral blood mononuclear cells (PBMC) expressed IL-29. However, little is known whether IL-29 is involved in
obesity. In this study, we harvested the adipose tissue and blood from obese patients that were classified based on their BMI. We also used SGBS cells to clarify whether the differentiated cells have IL-28RA and IL-10R2. We found that serum IL-29 level was significantly higher in obese
patients compared with healthy controls, and IL-29 was also detected in obese adipose tissue. IL-28R1 and IL-10R2 were expressed in mature human SGBS adipocytes which could serve as target cells of IL-29. To further investigated the
biological function of IL-29, We successfully expressed and purified human IL-29 protein. At the same time, IL-29 was used as an antigen to establish mouse monoclonal
antibodies. We also successfully established and screened specific monoclonal antibodies that could antagonize human IL-29 protein. It facilitates the progression of subsequent research plan.
I
Content
Content ... I I. 中文摘要 ... II
II. Abstract ... III
III. Background ... 1
1. Obesity ... 1
2. Interleukin-29 (IL-29) ... 1
IV. Reference literature ... 2
1. Obese-related disease ... 2
2. Obesity and chronic inflammation ... 2
3. Biological function of IL-29 ... 2
V. Research motivation ... 3
VI. Material and methods ... 4
1. Patients and Samples... 4
2. SGBS cell differentiation ... 4
3. Reverse-transcription-PCR (RT-PCR) and Quantification Real Time PCR (RT-qPCR) ... 4
4. Western blotting ... 4
5. Oil Red O staining and triglyceride quantification ... 5
6. Generation of IL-29 monoclonal antibody ... 6
7. Titer of anti-IL-29 monoclonal antibody ... 6
8. Purification of anti-IL-29 mAb ... 6
9. Specificity of monoclonal antibody against IL-29... 7
10. Statistical analysis ... 7
VII. Results ... 8
1. Higher serum IL-29 levels in obese patients ... 8
2. Expression of IL-29 receptor IL-28R1 and IL-10R2 in human SGBS adipocytes in adipogenic differentiation ... 8
3. IL-29 did not regulate lipid content of SGBS adipocytes ... 8
4. Generation of IL-29 monoclonal antibody ... 9
VIII. Discussion ... 9
IX. Reference ... 10
X. Figure and figure legend ... 13
Figure 1. Higher serum IL-29 levels in obese patients. ... 13
Figure 2. Expression of IL-29 receptors IL-28R1 and IL-10R2 in human SGBS adipocytes during adipogenic differentiation. ... 15
Figure 3. IL-29 did not regulate lipid content of SGBS adipocytes. ... 17
II 中文摘要 肥胖是一種慢性發炎疾病,與胰島素抗性、心血管疾病以及非酒精性脂肪肝 疾病的發生有密切關係。生活習慣、基因遺傳、飲食與代謝失調都可能造成肥胖。 在肥胖發展過程中,免疫細胞會浸潤到脂肪組織中,其中以單核細胞以及巨噬細 胞為主要的族群,這些細胞會分泌許多促發炎細胞激素,包括 TNF-α、 IL-1β、 IL-6 和 IL-8,進而引起脂肪組織的慢性發炎反應。因此在肥胖的發炎微環境中, 脂肪細胞與巨噬細胞間之交互作用是十分關鍵。介白素二十九 (Interleukin-29, IL-29)是第三型干擾素(interferon)家族的一員,透過 IL-28R1 和 IL-10R2 這對接 受器來傳遞訊號啟動生物功能。IL-29 在宿主對抗微生物入侵中具有重要作用, 在感染病毒的細胞中其表現量顯著上升。過去文獻得知巨噬細胞、樹突細胞、週
邊血液單核細胞都會分泌IL-29。然而,IL-29 是否參與在肥胖的慢性發炎中是未
知的。在這項研究中,我們收集了肥胖患者的脂肪組織和血液,並根據其BMI 數
值對其進行了分類。我們也使用SGBS 細胞來釐清分化過後的細胞是否具有
IL-28RA 和 IL-10R2。首先,我們發現肥胖患者的血清中可偵測高量的 IL-29,且在
肥胖患者的脂肪組織會高度表現IL-29。在人類脂肪細胞株(SGBS cells)的成熟過
程中會大量的表達IL-28R1 和 IL-10R2 這對接受器,因此可作為 IL-29 的目標細
胞。為了進一步研究IL-29 的生物功能,我們成功表達並純化人類 IL-29 蛋白質。
同時將IL-29 當成抗原來建立小鼠單株抗體,我們也成功建立並篩選專一性可拮
抗IL-29 的單株抗體,以利於後續研究計畫的執行。
III
I. Abstract
Obesity is a chronic inflammatory disease that is closely related to the development of insulin resistance, cardiovascular disease and nonalcoholic fatty liver disease. In the progression of obesity, immune cells are infiltrated into adipose tissue. Most are monocytes and macrophages. These cells secrete various inflammatory factors, including TNF-α, IL-1β, IL-6, and IL-8 and then cause chronic inflammation of adipose tissue. Therefore, adipocytes and macrophages interaction play a critical role in the inflammatory environment of obesity. Interleukin-29 (IL-29), a member of type 3 interferon family, signals through a heterodimer receptor complex of 28R1 and IL-10R2 to initiate biological functions. IL-29 plays an important role in the host against microbes and its gene is highly upregulated in cells infected with viruses. Several studies reported that macrophages, dendritic cells, and peripheral blood mononuclear cells (PBMC) expressed IL-29. However, little is known whether IL-29 is involved in obesity. In this study, we harvested the adipose tissue and blood from obese patients that were classified based on their BMI. We also used SGBS cells to clarify whether the differentiated cells have IL-28RA and IL-10R2. We found that serum IL-29 level was significantly higher in obese patients compared with healthy controls, and IL-29 was also detected in obese adipose tissue. IL-28R1 and IL-10R2 were expressed in mature human SGBS adipocytes which could serve as target cells of IL-29. To further investigated the biological function of IL-29, We successfully expressed and purified human IL-29 protein. At the same time, IL-29 was used as an antigen to establish mouse monoclonal antibodies. We also successfully established and screened specific monoclonal antibodies that could antagonize human IL-29 protein. It facilitates the progression of subsequent research plan.
1
II. Background
1. Obesity
In 1996, World Health Organization (WHO) classifies obesity as a chronic inflammatory disease. Body mass index (BMI) is used as an indicator of body fatness. It is defined as weight divided by the square of height. People are considered obese when their BMI is over 30 kg/m2. According to statistics of WHO in 2014, more than
600 million adults were obese in the world and it is further projected that 1.12 billion individuals will be classified as obese by 20301,2. The main cause is due to the
imbalance between intake and consumption of calories, so that excessive energy stored in adipocytes leads to increase of cell numbers (hyperplasic adipocytes) and expansion of cell size (hypertrophic adipocytes).
2. Interleukin-29 (IL-29)
In 2002, two research groups identified newly three cytokines, IL-28A, IL-28B, and IL-29 (One of group named IFN-λ2, IFN-λ3, and IFN-λ1)3,4. IL-28A, IL-28B, and
IL-29 are present in humans;IL-28A and IL-28B are exist in mice except IL-29. These cytokines belong to antiviral family of cytokines that are related to type I IFNs and IL-10 family members. IL-29 gene is located on a chromosomal region mapped to 19q13. IL-29 signals through a heterodimeric class II cytokine receptor that consisted of IL-28R1 and IL-10R2. IL-29 binds to its receptor and causes the activation of the two tyrosine kinases, Janus kinase 1 (Jak1) and tyrosine kinase 2 (Tyk2) which then leads to the phosphorylation and activation of STAT1 and STAT2. Phosphorylated STAT1 and STAT2 combine with interferon regulatory factor 9 (IRF9) to form a heterodimeric complex called IFN-stimulated gene factor 3 (ISGF3) which rapidly translocate to the cell nucleus to induce the expression of IFN-responsive genes (ISG). Besides the Jak/STAT pathway, IL-29 also leads to the activation of different mitogen-activated protein kinase (MAPK) pathways, including extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinase (JNK), and p38 kinase5.
2
III. Reference literature
1. Obese-related disease
Obesity is considered as a serious global epidemic that significantly affects population health6,7. Obesity elevates the morbidity of cardiovascular disease, type 2
diabetes, sleep apnea, cancers, gout and degenerative arthritis compared with healthy people8-10. In 2015, the top ten cause of death in Taiwan, including malignant tumors,
heart disease, cerebrovascular disease, diabetes, hypertensive diseases are highly associated with obesity.
2. Obesity and chronic inflammation
Previous research reported that tumor necrosis factor-α (TNF-α) levels were increased in obese adipose tissue and directly induced insulin resistance11. Several
studies have identified that the production of IL-1β, IL-6, and MCP-1 were elevated in obesity, which impaired insulin signaling indicating that inflammation is critical in the pathogenesis of obesity-induced metabolic disorders12-18. Obesity is a low-grade
chronic inflammation in adipose tissue. In the lean state, TH2 cells, Treg cells,
eosinophils, and M2-like (anti-inflammatory) macrophages predominate in adipose tissue. Treg cells secrete IL-10 to maintain M2-like macrophages and inhibit
macrophages migration. IL-4 released from TH2 cells and eosinophils induces the
expression of IL-10 in M2-like macrophages, which in turn keep anti-inflammatory and insulin-sensitive phenotype. In obese state, adipose tissue expands and the numbers of natural killer (NK) cells, neutrophils, and macrophages are rapidly increased in the adipose tissue. NK cells produce IFN-γ and drive TH1 cell expansion. Hypertrophic
adipocytes secrete MCP-1 to trigger the infiltration of monocytes and macrophages into the adipose tissue and then polarized to the M1-like (pro-inflammatory) macrophages
19. These immune cells release cytokines such as TNF-α, IL-1β, and IL-6 and then
contribute to adipose tissue inflammation, and insulin resistance. The number of Treg
cells and eosinophils declines in obese adipose tissue. Obesity also induces an increase in CD4+ T
H1 and CD8+ effector T cells, which secrete pro-inflammatory cytokines. B
cell numbers also increase and activate T cells, which potentiate M1-like macrophage polarization, inflammation and insulin resistance19-24. Adipocytes secrete
pro-inflammatory cytokines and adipokines are important factors in obesity and insulin resistance22,23,25. Cytokines and chemokines from the adipose tissue can also be released
into the circulation to promote inflammation in other tissues.
3. Biological function of IL-29
IL-29 predominantly expressed in the epithelial tissues. Besides, several immune cells like dendritic cells (DC), macrophages, and TH17 cells are cellular sources of IL-2926-28. Like type I IFN, IL-29 plays a critical role in host defense against microbes and
3
including lung cancer, esophageal carcinomas, pancreatic cancer, and colorectal cancer29. In addition to its anti-viral and anti-tumor activities, IL-29 acts as an
immune-modulator in rheumatoid arthritis (RA) and allergic asthma.
IV. Research motivation
Obesity is associated with a state of chronic low-grade inflammation and elevates the morbidity of insulin resistance, cardiovascular disease and NAFLD. During the development of obesity, macrophages accumulate in adipose tissue and secrete inflammatory factors to induce inflammation30. Therefore, adipocyte-macrophage
crosstalk is critical in obesity.
IL-29 regulates immune response in RA and allergic asthma. However, it is unclear the relation between IL-29 and obesity. Here, we aimed to investigate the role of IL-29 in obese patients and adipocyte-macrophage interaction.
4
V. Material and methods
1. Patients and Samples
Adipose tissues were harvested from obese patients. BMI can be used for patient classification. Blood samples were collected from 41 obese patients, 13 pre-obese patients and 10 healthy controls. Serum from patients was collected for ELISA analysis. RNA extract from obese adipose tissue were used for RT-PCR analysis. The study was approved by the Ethics Committee of National Cheng Kung University Hospital Institutional Review Board. Signed informed consent was obtained from all participants.
2. SGBS cells differentiation
SGBS preadipocytes were kindly provided by Professor Wabitsch (University of Ulm, Germany). Human SGBS preadipocytes were cultured in DMEM/F12 medium containing 10% fetal calf serum (FCS; Invitrogen), 3.3 μM biotin (Sigma), 1.7 μM pantothenate (Sigma), 100 units/ml of penicillin (Caisson) and 100 µg/ml streptomycin (Caisson) until reaching confluence. To induce SGBS preadipocytes differentiation, cells were washed with PBS twice and cultured in differentiation medium (DMEM/F12 supplemented with 3.3 μM biotin, 1.7 μM pantothenate, 2 μM rosiglitazone, 25 nM dexamethasone, 0.5 mM methylisobuthylxantine (IBMX), 1 µM cortisol, 0.01 mg/ml transferrin, 0.2 nM triiodotyronin, and 20 nM human insulin) for the first 4 days. After 4 days, the differentiating cells were kept in differentiation medium excluding dexamethasone, IBMX, and rosiglitazone. The medium was changed every 4 days.
3. Reverse-transcription-PCR (RT-PCR) and Quantification Real Time PCR (RT-qPCR)
The total volume of each reverse-transcription reaction was 20 μL. 10 μL RNA (2 μg) was mixed with 5 pM random primer and 0.5 mM dNTP. The mixture was heated at 65°C for 5 mins and incubated on ice. 1 μL M-MLV reverse transcriptase (Invitrogen, USA), 5 mM DTT, and 1× first strand buffer were added and then reacted for 10 mins at 25 °C, 50 mins at 37 °C and 15 mins at 70 °C. The cDNA was diluted for ten-fold with RNase free water for analysis.
RT-qPCR was performed with a Rotor-Gene Q detection system (QIAGEN). Primers used for RT-qPCR are listed in Table 1. Gene expression was normalized to human β-actin using the ΔCt method. The reaction components contained 2 μL diluted cDNA sample, 0.7 μM primers of each, and 5 μL SYBR green master mix. The final volume of each reaction was filled up to 10 μL with RNase-free water. The RT-qPCR conditions consisted of an initial activation at 95°C for 2 mins, followed by 40-45 cycles of 5 s at 95°C and 10 s at 60°C.
4. Western blotting
Proteins were separated by 12%–16% SDS-PAGE and transferred electrophoretically to PVDF membranes (Millipore, USA). Membranes were blocked
5
with 5% (w/v) non-fat milk in PBST (140 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4,
1.8 mM KH2PO4, 0.01% Tween-20, pH=7.4) for 2 hrs at room temperature, and then
incubated overnight at 4 °C with primary antibody. After binding of primary antibodies, membranes were washed with PBST three times and incubated for 1 hr at room temperature with the species-specific horseradish peroxidase-labeled (HRP) secondary antibodies. Binding of secondary antibodies was detected with SuperSignal West Pico Chemiluminescent Substrate where the chemiluminescent signals were visualized and imaged following exposure and development of Hyperfilm ECL molecular on Luminescence Imaging System.
5. Oil Red O staining and triglyceride quantification
Pre-SGBS cells were incubated for 14 days in differentiation medium. Mature SGBS adipocytes were washed twice with PBS and fixed with 3.7% formaldehyde for 30 mins. After fixation, the cells washed twice with distilled water and once with 1, 2-isopropanediol (Sigma). Then, the cells stained with Oil red O (Sigma) solution (0.3 g Oil red O in 100mL 60% 1,2-isopropanediol) for 15-30 mins at room temperature. Then, the cells were washed with distilled water three times to remove background staining. Finally, cells were visualized through 4 × or 10 × objectives mounted on for observation. For the quantification, the oil red-O dye was extracted with 100% isopropanol and measured by a micro-plate reader (Thermo Scientific™) at 492 nm.
6. Immunocytochemistry staining
Cells were rinsed with PBS twice and fixed in 3.7% formaldehyde for 30 mins. Cells were then blocked in antibody diluent (DAKO®) for 2 hrs at room temperature and incubated with primary antibody at 4 °C overnight. Next day, samples were washed with PBS for 10 mins three times and incubated for 1 hr at room temperature with appropriate secondary antibody (Invitrogen Life Technologies). The staining was visualized using 3-amino-9-ethylcarbazole (AEC) solution.
7. Construction of IL-29 in pSecTag/Hygro A vector
A cDNA fragment of human IL-29 was inserted into the expression vector of pSecTag/Hygro A. IL-29 fragment was amplified by PCR with specific primers (see Table1). After amplification, the PCR product was purified using a PCR purification kit (QIAGEN). PCR product and pSecTag/Hygro A vector were digested for 3 hrs at 37°C using restriction endonucleases BamHI and NotI (NEB) and ligated with T4 DNA ligase for 16 hrs at 4°C (NEB).
8. Transfection of IL-29_pSecTag/Hygro A into mammalian cells
293T cells were cultured in DMEM/HG medium with 10% FBS, 100 units/ml of penicillin and 100 µg/ml streptomycin. The cells were seeded onto 6-well cell culture plates (NUNC) at a density of 3 × 105 cells per well. After overnight culture, the cells
6
transfection reagent (Invitrogen). After transfection for 48 hrs, cell was treated with 400 μg/mL hygromycin B (InvivoGen) to select the positive cells.
9. Purification of human recombinant IL-29 (rhIL-29) from mammalian cells
The conditioned medium from 293T cells that expressed IL-29 was collected and purified by affinity chromatography. Ni2+ charged metal chelate column (1 mL bed
volume; GE Healthcare) equilibrated in pSec Tag2/HygroA protein binding buffer (A11; 20 mM NaCl, 0.5 mM Tris-HC1, 1mM imidazole and 20 mM β-mercaptoethanol, pH 7.6). Protein was eluted with pSec Tag2/HygroA protein elution buffer (B1; 0.7 M NaC1, 6.5 mM Tris-HC1, 500 mM imidazole and 20 mM β-mercaptoethanol, pH 7.6). Fractions were collected at a flow-rate of 1 ml/min. The purified IL-29 was dialyzed into PBS overnight at 4 °C. This purified IL-29 protein was used for the selection of IL-29 mAbs as described below.
10. Generation of IL-29 monoclonal antibody
BALB/cJ mice were immunized subcutaneously every two weeks for 10 weeks with rhIL-29 (25 μg/mouse for first immunization, 50 μg/mouse for subsequent boosting) emulsified with an equal volume of Freund's complete/incomplete adjuvant. The mice were scarificed and spleen cells were isolated. Spleen cells were fused with SP2 myeloma cells in ratio 1:5 with PEG 4000 (Merck and Co., Inc., Whitehouse Station, NJ). After fusion, the cells were resuspended into 96-well plates and cultured with HAT medium (Sigma) for 2 days then replaced with HT medium (Sigma). Using ELISA, culture supernatant was tested for antibody reacting with rhIL-29. To clone the selected hybridoma cells, the limiting dilution was carried out twice. The hybridoma cells were cultured in Dulbecco's Modified Eagle medium (Invitrogen) containing 10% FBS, 100 units/ml of penicillin, 100 µg/ml streptomycin, 2% L-glutamine adjusted NaHCO3 solution. The isotype of the selected antibody, mIgG, was determined using
RT-PCR with specific primer.
11. Titer of anti-IL-29 monoclonal antibody
Serum titer of anti-IL-29 monoclonal antibody titer is determined using ELISA. To test the titer of antibody, rhIL-29 from mammalian cells was coated on 96-well plates overnight at 4oC. Plate was washed with PBST and blocked with blocking buffer (1%
BSA in PBS) for 2 hrs at room temperature. The serum from immunized mice as primary antibody was added into the plates and incubated for 1 hr at room temperature. After washing with PBST, HRP-conjugated goat anti-mouse IgG was added and incubated 1 hr at room temperature. After washing, plate was incubated with tetramethylbenzidine (TMB) substrate solution. The reaction was stopped with 2 N H2SO4. The absorbance at 450 nm was measured with a micro-plate reader.
12. Purification of anti-IL-29 monoclonal antibody
7
purified by affinity chromatography. Protein A column (5 mL bed volume; GE Healthcare) equilibrated in protein A column binding buffer (A11; 19.5 mM NaH2PO4,
30.5 mM Na2HPO4, pH 7.4). Protein was eluted with protein A column elution buffer
(B1; 500 mM citric acid, pH 3.5). Fractions were collected at a flow-rate of 1 ml/min. The purified IL-29 mAb was rapidly adjusting to neutral pH (pH=7.4) with 1M Tris-base (pH 9.0, 100 μL/1mL elution sample) and dialyzed into PBS overnight at 4 °C.
13. Specificity of monoclonal antibody against IL-29
Cytokines including IL-17, IL-19, IL-20, IL-22, and IL-24 were coated on 96-well plates overnight at 4oC. Plate was washed with PBST three times and blocked with
blocking buffer (1% BSA in PBS) for 2 hrs at room temperature. After washing with PBST three times, IL-29 mAb was added into plates for overnight at 4oC. The plate was
washed with PBST three times and then incubated with second antibody for 1 hr. After washing, plate was incubated with TMB substrate solution. The reaction was stopped with 2 N H2SO4. The absorbance at 450 nm was measured with a micro-plate reader.
14. Statistical analysis
Prism 6.0 (GraphPad Software) was used for the statistical analysis. T-test and A one-way ANOVA nonparametric test (Kruskal-Wallis test) was used to compare the data between groups. Data are expressed as the mean of replicate measurements or mean normalized values between multiple experiments ± SEM or SD. P < 0.05 was considered statistically significantly.
8
VI. Results
1. Higher serum IL-29 levels in obese patients
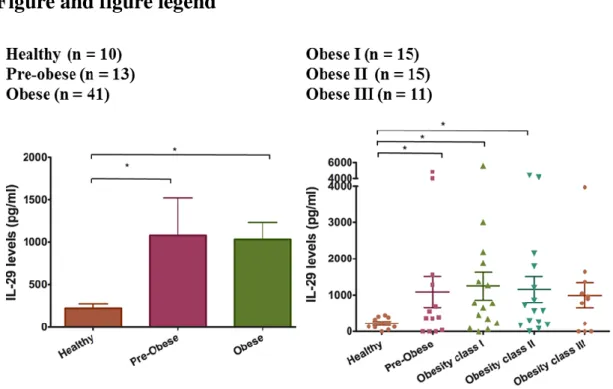
First, we investigated the clinical implication of IL-29 in obesity. We analyzed the serum level of IL-29 in patients with different weight. According to BMI, we divided them into several groups: healthy control (BMI:18.5~24.9 kg/m2 ), overweight (BMI:
25.0~29.9 kg/m2), and obesity (BMI ≥ 30.0 kg/m2). Among obese patients, they were
divided into obesity class I obesity (BMI:30.0~34.9 kg/m2), class II obesity (BMI:
35.0~39.9 kg/m2), and class III obesity (BMI ≥ 40.0 kg/m2). We found that serum level
of IL-29 was significantly higher in the patients with overweight and obesity than normal weight (p < 0.05) and serum IL-29 level was raised accompanying with increase of the obesity degree.
2. Expression of IL-29 receptor IL-28R1 and IL-10R2 in human SGBS adipocytes in adipogenic differentiation
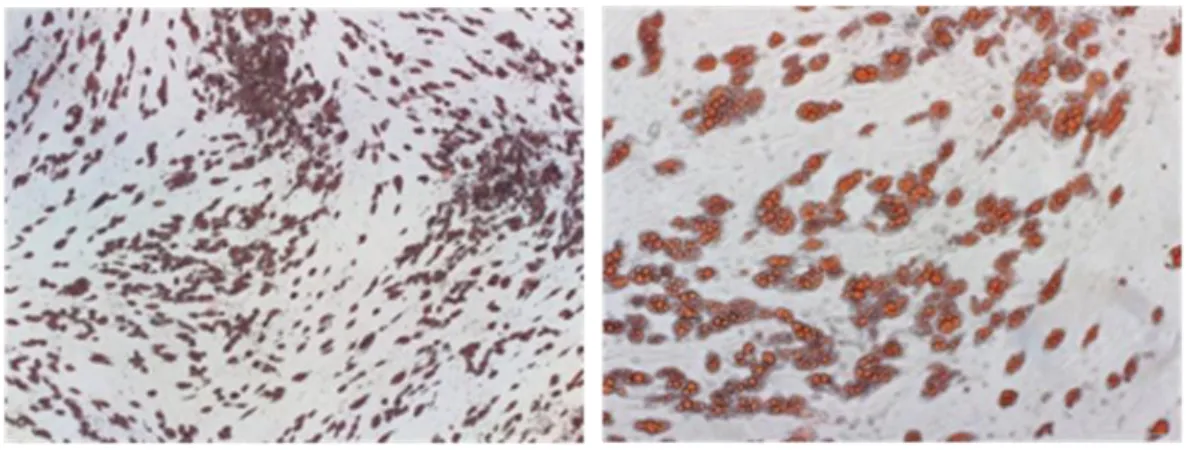
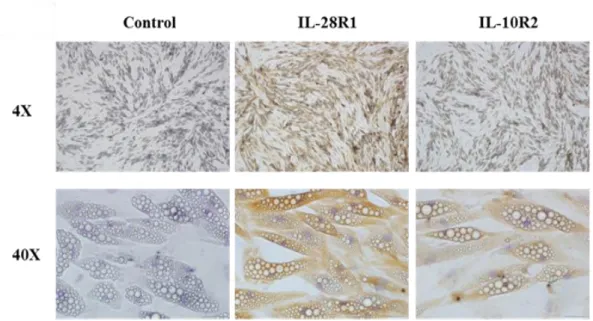
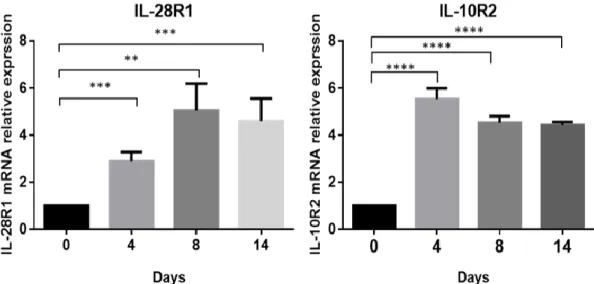
To explore the role of IL-29 in adipocytes, we used a human cell line SGBS preadipocytes, which was originally obtained from an adipose tissue specimen of a patient with Simpson-Golabi-Behmel syndrome. It took 14 days to differentiate into mature SGBS adipocytes (Figure 2). First, we examined whether SGBS adipocytes are target cells for IL-29, we used RT-PCR and immunocytochemistry staining to analyze the expression of IL-29 receptor, IL-28R1 and IL-10R2, in differentiated SGBS adipocytes. The results showed that both IL-28R1 and IL-10R2 mRNA and protein were expressed in SGBS adipocytes (Figure 3). RT-qPCR confirmed that the expression of IL-28R1 and IL-10R2 were significantly (p < 0.05) increased during adipocyte differentiation (Figure 4).
3. IL-29 did not regulate lipid content of SGBS adipocytes
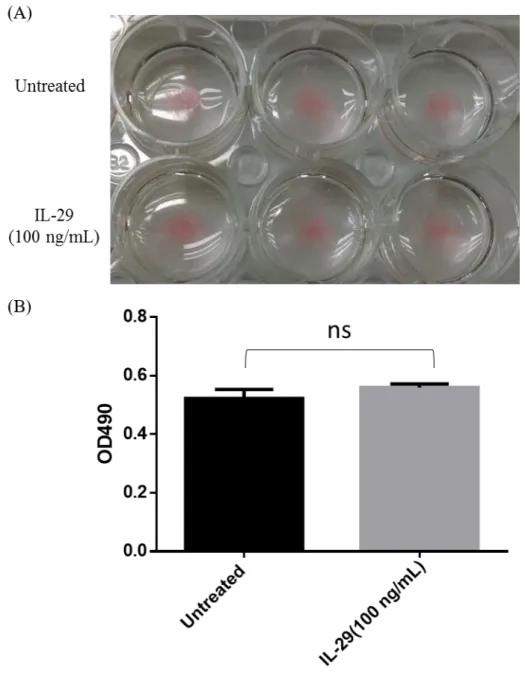
Because IL-28R1 and IL-10R2 were increased during adipocyte differentiation, we wanted to identify whether IL-29 affected adipocyte differentiation and oil droplets formation. SGBS adipocytes were treated with 100 ng/mL IL-29 during differentiation and lipid content was determined by Oil Red O staining. We found that IL-29 did not regulate oil droplets content of SGBS adipocytes (Figure 5A, B).
4. Expression and purification of IL-29 in mammalian cell system
We constructed IL-29 into pSecTag/Hygro A vector and transiently transfected into 293T cells. Western blotting was performed to detect 29 expression using IL-29 antibody (Figure 6A). After transfection for 48 hrs, cell was selected using hygromycin B (Invitrogen) to generate stable cell lines expressing IL-29. A large volume of culture medium was collected and purified by affinity chromatography. (Figure 6B) The approximate molecular weight of purified rhIL-29 from mammalian cells was 29-42kDa (Figure 6C). These proteins from different sources were used for
9
in vitro experiments and for generation of IL-29 mAb.
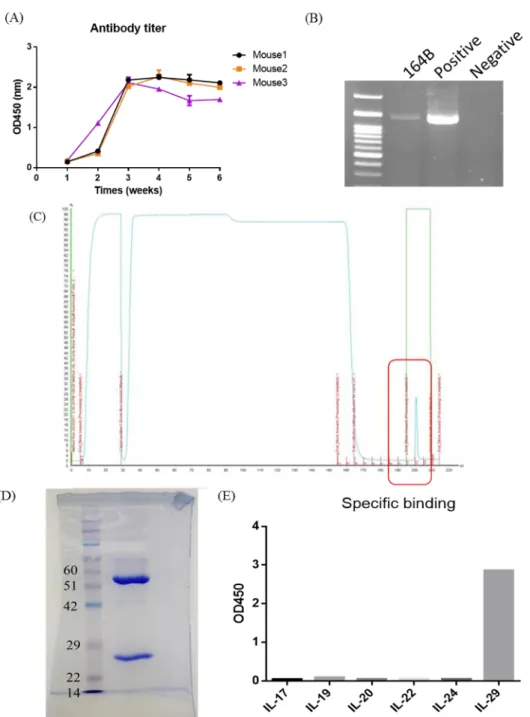
5. Generation of IL-29 monoclonal antibody
We immunized mice with rhIL-29, which was purified from E.coli system and then collected serum to test the titer of IL-29 antibodies after the third and fifth time boosting. The production of IL-29 antibodies from immunized mice was observed after the third time boosting (Figure 7A). After the fifth time boosting, mice were sacrificed and isolated splenocytes. We fused myeloma SP2 cells with splenocytes using PEG to generate hybridoma cells. We selected positive hybridoma using direct ELISA with coating rhIL-29 which was purified from IL-29 stable clone cells. We successfully generated and selected 10 antibody clones with specific recognizing IL-29. RT-PCR confirmed the isotype of IL-29 mAb (164B) was mIgG1 (Figure 7B). IL-29 mAb (164B) was purified by affinity chromatography using protein A column (Figure 7C,
D). We also analyzed 17, 19, 20, 22, 24 to confirm the specificity of
IL-29 mAb (164B) using ELISA. Only IL-IL-29 was recognized by IL-IL-29 mAb 164B (Figure
7E).
VII.
Discussion
Previously studies reported that IL-29 regulates innate and adaptive immunity and plays a role in RA and allergic asthma5,31. In this study, we found that the serum level
of IL-29 was significantly higher in obese patients and was increased accompanying with the raise of the obesity degree. RT-PCR showed that IL-29 was expressed in obese adipose tissue. These results suggested that IL-29 might be involved in obesity. To further understand the relationship between IL-29 and inflammation, we used human SGBS adipocytes. This cells have been used for a number of studies on adipose differentiation, adipocyte glucose uptake, lipolysis, apoptosis, regulation of expression of adipokines and protein translocation32. Therefore, we firstly confirmed the IL-29
receptors (IL-28R1 and IL-10R2) were expressed in SGBS cells, which indicated that SGBS adipocytes are the target cell for IL-29. Next, we performed SGBS adipocytes were treated with IL-29 during differentiation, but IL-29 did not regulate oil droplets content of SGBS adipocytes. Obesity is associated with chronic low-grade systemic inflammation. Various cytokines such as TNF-α, IL-1β, IL-6, and IL-8 are involved in the development of obesity2. Therefore, we speculated IL-29 may regulate SGBS
10
VIII.
Reference
1 Kahn, B. B. & Flier, J. S. Obesity and insulin resistance. J Clin Invest 106, 473-481, doi:10.1172/JCI10842 (2000).
2 Hotamisligil, G. S. Inflammation, metaflammation and immunometabolic disorders. Nature 542, 177-185, doi:10.1038/nature21363 (2017).
3 Sheppard, P. et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nature immunology 4, 63-68, doi:10.1038/ni873 (2003).
4 Kotenko, S. V. et al. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nature immunology 4, 69-77, doi:10.1038/ni875 (2003).
5 Xu, L. et al. Interleukin-29 induces receptor activator of NF-kappaB ligand expression in fibroblast-like synoviocytes via MAPK signaling pathways. International journal of rheumatic diseases 18, 842-849, doi:10.1111/1756-185x.12747 (2015).
6 Maffetone, P. B., Rivera-Dominguez, I. & Laursen, P. B. Overfat and Underfat: New Terms and Definitions Long Overdue. Frontiers in Public Health 4, 279, doi:10.3389/fpubh.2016.00279 (2016).
7 Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet (London, England) 387, 1377-1396, doi:10.1016/s0140-6736(16)30054-x (2016).
8 Subramanian, V. & Ferrante, A. W., Jr. Obesity, inflammation, and
macrophages. Nestle Nutrition workshop series. Paediatric programme 63, 151-159; discussion 159-162, 259-168, doi:10.1159/000209979 (2009). 9 Guilherme, A., Virbasius, J. V., Puri, V. & Czech, M. P. Adipocyte
dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nature reviews. Molecular cell biology 9, 367-377, doi:10.1038/nrm2391 (2008). 10 Kim, J. B. Dynamic cross talk between metabolic organs in obesity and
metabolic diseases. Exp Mol Med 48, e214, doi:10.1038/emm.2015.119 (2016).
11 Hotamisligil, G., Shargill, N. & Spiegelman, B. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science
259, 87-91, doi:10.1126/science.7678183 (1993).
12 Vandanmagsar, B. et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med 17, 179-188,
doi:http://www.nature.com/nm/journal/v17/n2/abs/nm.2279.html#supplementa
ry-information (2011).
11
members in morbid obesity and effects of weight loss. Molecular medicine (Cambridge, Mass.) 17, 840-845, doi:10.2119/molmed.2010.00108 (2011). 14 Roytblat, L. et al. Raised Interleukin-6 Levels in Obese Patients. Obesity
Research 8, 673-675, doi:10.1038/oby.2000.86 (2000).
15 Kern, P. A., Ranganathan, S., Li, C., Wood, L. & Ranganathan, G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. American Journal of Physiology - Endocrinology And Metabolism 280, E745-E751 (2001).
16 Eder, K., Baffy, N., Falus, A. & Fulop, A. K. The major inflammatory mediator interleukin-6 and obesity. Inflammation Research 58, 727, doi:10.1007/s00011-009-0060-4 (2009).
17 Panee, J. Monocyte Chemoattractant Protein 1 (MCP-1) in obesity and diabetes. Cytokine 60, 1-12, doi:http://dx.doi.org/10.1016/j.cyto.2012.06.018
(2012).
18 Kim, C. S. et al. Circulating levels of MCP-1 and IL-8 are elevated in human obese subjects and associated with obesity-related parameters. Int J Obes 30, 1347-1355 (2006).
19 Lackey, D. E. & Olefsky, J. M. Regulation of metabolism by the innate immune system. Nat Rev Endocrinol 12, 15-28, doi:10.1038/nrendo.2015.189 (2016).
20 Sell, H., Habich, C. & Eckel, J. Adaptive immunity in obesity and insulin resistance. Nat Rev Endocrinol 8, 709-716 (2012).
21 Lumeng, C. N. Innate Immune Activation in Obesity. Molecular aspects of medicine 34, 12-29, doi:10.1016/j.mam.2012.10.002 (2013).
22 Osborn, O. & Olefsky, J. M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat Med 18, 363-374 (2012). 23 McArdle, M., Finucane, O., Connaughton, R., McMorrow, A. & Roche, H.
Mechanisms of Obesity-Induced Inflammation and Insulin Resistance: Insights into the Emerging Role of Nutritional Strategies. Frontiers in Endocrinology 4, doi:10.3389/fendo.2013.00052 (2013).
24 Dalmas, E., Clément, K. & Guerre-Millo, M. Defining macrophage phenotype and function in adipose tissue. Trends in Immunology 32, 307-314,
doi:10.1016/j.it.2011.04.008.
25 Choe, S. S., Huh, J. Y., Hwang, I. J., Kim, J. I. & Kim, J. B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Frontiers in Endocrinology 7, doi:10.3389/fendo.2016.00030 (2016).
26 Wolk, K. et al. Maturing dendritic cells are an important source of IL-29 and IL-20 that may cooperatively increase the innate immunity of keratinocytes.
12
Journal of leukocyte biology 83, 1181-1193, doi:10.1189/jlb.0807525 (2008). 27 Wolk, K. et al. IL-29 is produced by T(H)17 cells and mediates the cutaneous antiviral competence in psoriasis. Science translational medicine 5, 204ra129, doi:10.1126/scitranslmed.3006245 (2013).
28 Siren, J., Pirhonen, J., Julkunen, I. & Matikainen, S. IFN-alpha regulates TLR-dependent gene expression of IFN-alpha, IFN-beta, IL-28, and IL-29. Journal of immunology (Baltimore, Md. : 1950) 174, 1932-1937 (2005).
29 Kelm, N. E. et al. The role of IL-29 in immunity and cancer. Critical Reviews in Oncology/Hematology 106, 91-98,
doi:http://dx.doi.org/10.1016/j.critrevonc.2016.08.002 (2016).
30 Weisberg, S. P. et al. Obesity is associated with macrophage accumulation in adipose tissue. The Journal of clinical investigation 112, 1796-1808,
doi:10.1172/jci19246 (2003).
31 Koch, S. & Finotto, S. Role of Interferon-lambda in Allergic Asthma. Journal of innate immunity 7, 224-230, doi:10.1159/000369459 (2015).
32 Fischer-Posovszky, P., Newell, F. S., Wabitsch, M. & Tornqvist, H. E. Human SGBS Cells – a Unique Tool for Studies of Human Fat Cell Biology. Obesity Facts 1, 184-189 (2008).
13
Figure and figure legend
Figure 1. Higher serum IL-29 levels in obese patients.
(A) Level of IL-29 in serum from 10 healthy controls, 13 overweight patients (pre-obese), 15 class I obese patients, 15 class II obese patients, and 11 class III obese patients were analyzed using ELISA. Data are expressed as mean ± SD of triplicate samples from a single experiment and are representative of three independent experiments. *P < 0.05 compared with healthy controls. (B) The expression of IL-29 in adipose tissues isolated from obese patients was detected and analyzed using RT-PCR with specific primers. β-actin was an internal control.
14
Figure 2. Human SGBS preadipocytes differentiated into mature SGBS adipocytes.
The SGBS preadipocytes were differentiated into mature SGBS cells in differentiation medium for 14 days. The SGBS preadipocytes and mature SGBS cells were stained with Oil Red O.
15
Figure 3. Expression of IL-29 receptors IL-28R1 and IL-10R2 in human SGBS adipocytes by immunohistochemistry staining.
The expression of IL-28R1 and IL-10R2 in human mature SGBS adipocytes was analyzed using RT-PCR with IL-28R1 and IL-10R2 antibody by immunohistochemistry staining.
16
Figure 4. Expression of IL-29 receptors IL-28R1 and IL-10R2 in human SGBS adipocytes during adipogenic differentiation by RT-qPCR.
The expression of IL-28R1 and IL-10R2 in human mature SGBS adipocytes was analyzed using RT-PCR with specific primer. *P < 0.05, **P < 0.01, ***P < 0.001 compared with day 0 controls.
17
Figure 5. IL-29 did not regulate lipid content of SGBS adipocytes.
(A) SGBS preadipocytes were incubated for 14 days in differentiation medium with IL-29 (100 ng/ml). The lipid droplets were stained with Oil Red O. (B) The OD value of Oil Red O eluted solution, which represents lipid droplet accumulation in the cytoplasm. ns: non-significant.
18
Figure 6. Expression and purification of human IL-29 recombinant protein.
(A) Generation of stable clone expressing IL-29 from 293T cells mammalian system and analyzed using western blotting with specific antibody against IL-29. (B-C) Human IL-29 recombinant protein was expressed and purified from the culture medium of IL-29 stable 293T cells using FPLC and analyzed the protein purity using Coomassie Blue staining of 12% SDS-PAGE gels.
19
Figure 7. Generation of anti-IL-29 monoclonal antibody (164B).
(A) The serum titer of anti-IL-29 mAb from immunized mice. (B) The isotype of anti-IL-29 mAb (164B). (C-D) Anti-IL-29 mAb (164B) was expressed and purified from the culture medium using FPLC with protein A column and analyzed the antibody purity using Coomassie Blue staining of 12% SDS-PAGE gels. (E)Analysis of anti-IL-29 mAb specific binding affinity. IL-17, IL-19, IL-20, IL-22, and IL-24 were coated on 96 wells plate, the specificity of IL-29 mAb (164B) were confirmed using ELISA. Only IL-29 was recognized by IL-29 mAb (164B).
108年度專題研究計畫成果彙整表
計畫主持人:許育祥 計畫編號:108-2320-B-006-052-計畫名稱:介白素二十九在肥胖誘導之發炎及胰島素抗性中的調控角色 成果項目 量化 單位 質化 (說明:各成果項目請附佐證資料或細 項說明,如期刊名稱、年份、卷期、起 訖頁數、證號...等) 國 內 學術性論文 期刊論文 0 篇 研討會論文 1 台灣臨床醫學研究所聯合教學研究研討 會 專書 0 本 專書論文 0 章 技術報告 0 篇 其他 0 篇 國 外 學術性論文 期刊論文 1 篇Cell Mol Immunol. 2020 Apr;17(4):369-379. doi: 10.1038/s41423-019-0262-9. 研討會論文 0 專書 0 本 專書論文 0 章 技術報告 0 篇 其他 0 篇 參 與 計 畫 人 力 本國籍 大專生 0 人次 碩士生 2 兩位碩士班學生參與研究 博士生 0 博士級研究人員 0 專任人員 1 一位研究助理參與研究 非本國籍 大專生 0 碩士生 0 博士生 0 博士級研究人員 0 專任人員 0 其他成果 (無法以量化表達之成果如辦理學術活動 、獲得獎項、重要國際合作、研究成果國 際影響力及其他協助產業技術發展之具體 效益事項等,請以文字敘述填列。)