Abstract. It is well known that the response of cancer cells to chemotherapeutic drugs involves the activation of apoptotic pathways. Benzyl isothiocyanate (BITC) is an important compound found in plant food and has been shown to have anti-cancer effects on human cancer cells, but its effect on prostate cancer cells in vitro remains unknown. The aim of the present study was to investigate the effects of BITC on DU 145 human prostate cancer cells in order to clarify whether a time/concentration range for optimal BITC-induced apop- tosis exists and to find the associated signaling pathway. Cell morphological changes, percentage of cell viability, DNA damage and apoptosis in DU 145 cells were examined by phase-contrast microscopy, flow cytometric assay, 4',6- diamidine-20-phenylindole dihydrochloride staining, comet assay and Western blotting analysis. The results indicate that BITC induces cell morphological changes, decreases the percentage of viable cells (induction of cell cytotoxicity), and induces DNA damage and apoptosis in DU 145 cells in a time- and dose-dependent manner. Flow cytometric assays indicated that BITC promoted reactive oxygen species and Ca
2+productions and decreased the levels of mitochondrial
membrane potential (Δ
Ψm), while the pre-treatment with N-acetylcysteine caused an increase in the percentage of viable cells. BITC also promoted caspase-3, -8 and -9 activities. Furthermore, when cells were pre-treated with the caspase-3 inhibitor and then treated with BITC, this led to an increase in the percentage of viable cells. Confocal laser microscopy examination indicated that BITC promoted the expression of AIF and Endo G, which were released from the mitochondria in DU 145 cells. In conclusion, BITC induces apoptosis in DU 145 cells through the release of AIF and Endo G from the mitochondria and also promotes caspase-3 activation.
Introduction
Apoptosis (programmed cell death), is a natural process for the elimination of unwanted cells, and plays an essential role in the normal development and the maintenance of homeo- stasis (1,2). It plays a critical role in protecting mechanisms against damaged, improperly produced or tumorigenic cells (3) and also in eliminating damaged cells (4). Many studies have shown that dysregulation of apoptosis can lead to cancer development and this provides a strategy for developing therapeutic agents through the induction of apoptosis and results in the elimination of premalignant or malignant cells (5,6). Therefore, the best strategy is focused on selectively destroying tumor cells through its induction (7) for the development of new anti-cancer agents (8-10).
Prostate cancer is one of the major causes of cancer death in male populations in developed countries. Even though various novel therapeutic strategies are now being considered, the cytotoxic drugs being used clinically are still limited based on the intrinsic or acquired resistance and toxicity.
Evidence from epidemiological studies has demonstrated that the dietary intake of cruciferous vegetables can have a protective effect against the risk of various types of malignancies (11,12). Isothiocyanates (ITCs) are the major
The roles of AIF and Endo G in the apoptotic effects of benzyl isothiocyanate on DU 145 human prostate cancer cells
via the mitochondrial signaling pathway
KUO-CHING LIU
1*, YA-TING HUANG
1*, PING-PING WU
4, BIN-CHUAN JI
6, JAI-SING YANG
2, JIUN-LONG YANG
5, TSAN-HUNG CHIU
7, FU-SHIN CHUEH
8and JING-GUNG CHUNG
3,9*1
Department of Medical Laboratory Science and Biotechnology; Departments of
2Pharmacology and
3Biological Science and Technology;
4School of Pharmacy;
5Graduate Institute of Chinese Pharmaceutical Science, China Medical University,
Taichung 404;
6Division of Respiratory Care Center, Department of Medicine, Changhua Christian Hospital, Changhua 500;
7Department of OBS/GYN, China Medical University Hospital, Taichung 404;
Departments of
8Health and Nutrition Biotechnology and
9Biotechnology, Asia University, Wufeng, Taichung 413, Taiwan, R.O.C.
Received October 8, 2010; Accepted December 2, 2010 DOI: 10.3892/ijo.2010.894
_________________________________________
Correspondence to:
Dr Jing-Gung Chung, Department of Biological Science and Technology, China Medical University, No.91 Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C.
E-mail: [email protected]
*Contributed equally
Key words:
benzyl isothiocyanate, cell cycle arrest, apoptosis, mitochondria, prostate cancer cellscomponents in cruciferous vegetables including broccoli, mustard, horseradish, watercress and cabbage, and thus play a role in their anticarcinogenic effect (13). Benzyl ITC (BITC) is one of the ITCs, and has been shown to prevent mammary cancer in MMTV-neu mice via the suppression of cellular proliferation, increased apoptosis and the enhanced infiltration of T cells in carcinoma (14). Many studies have shown that BITC induces apoptotic cell death of cultured human breast cancer cells in vitro (15-18). Furthermore, antioxidant mechanisms also contribute to the BITC-mediated chemo- prevention against inflammation-related carcinogenesis (19).
However, the exact mechanism underlying the role of BITC- mediated apoptosis in human prostate cancer has not been fully elucidated.
It has been reported that BITC can act as an anti-cancer agent through the inhibition of the growth of human cancer cells. However, little is known about the anti-cancer effect of BITC on human prostate cancer cells. In the present study, we evaluated the effects of different concentrations of BITC on DU 145 prostate cancer cells in relation to cell growth and death. We also observed that the BITC-induced reactive oxygen species (ROS) production is closely correlated with the modification of the mitochondrial function.
Materials and methods
Reagents. BITC was purchased from Sigma-Aldrich Corp.
(St. Louis, MO, USA). Stock solution of BITC was prepared in DMSO, and an equal volume of DMSO (0.1%) was added to the controls. Propidium iodide (PI), RNase A and 4,6- diamidino-2-phenylindole (DAPI) were from Sigma-Aldrich.
RPMI-1640, fetal calf serum, penicillin and streptomycin antibiotic mixture were purchased from Life Technologies/
Invitrogen (Grand Island, NY). Anti-actin antibody was from Sigma. The antibodies against Bak, Bax, Bcl-x
L, cyclin- dependent kinase (Cdk) 1, cell division cycle 25C (Cdc25C) and Smac/DIABLO were from Santa Cruz Biotechnology (Santa Cruz, CA), and the antibodies against cytochrome c, and caspase-8 and -9 were from PharMingen (Palo Alto, CA). DiOC
6, 6-carboxy-2,7-dichlorodihydrofluorescein diacetate (H
2DCFDA) and Fluo-3/AM were purchased from Invitrogen (Carlsbad, CA, USA).
Cell culture. The human prostate carcinoma cell line (DU 145) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). DU 145 cells were maintained in RPMI-1640 with 2 mM L-glutamine supplemented with 10% heat-inactivated fetal bovine serum and 1% antibiotic/antimycotic. Cells (1x10
5cells/ml) were maintained at 5% CO
2and 37˚C in humidified 5% CO
2and 95% air.
Examinations of cell morphological changes and viability. A phase contrast microscope was used for examining the morphological changes of cells. The PI exclusion assay and flow cytometry were performed for the quantitative analysis of cell viability. DU 145 cells (1x10
5cells/ml) were placed in 12-well plates and were then treated with 0, 2.5, 5, 7.5, 10 and 15 μM of BITC and were incubated for different time periods. For the morphological changes examination, cells
were examined and photographed under a phase contrast microscope at x200 magnification. After culturing with BITC at 37˚C for different time periods, 0.5 ml of PBS containing 5 μg/ml PI were added to the cells and viable cells were determined using a FACScalibur with CellQuest software (Becton-Dickinson, San Jose, CA, USA) as previously described (20,21). For the trypan blue dye exclusion assay, the cells were collected from each well and re-suspended in PBS. The cell suspensions were mixed with 0.4% trypan blue stain. The viable cells (cells that were not stained blue) and dead cells (cells that were stained blue) were counted using a hemocytometer under a light microscope (22,23).
DNA content and sub-G1 phase analyses. The effects of BITC on cell cycle distribution including sub-G1 (apoptosis) were assessed by flow cytometry as described previously (22,23). Briefly, 1x10
5cells were plated in 12-well plates in 2 ml RPMI-1640 medium and allowed to attach overnight.
The medium was replaced with fresh medium and then the cells were treated with or without 0, 2.5, 5, 7.5, 10 and 15 μM BITC for 48 h. Control cells were treated with 0.1% DMSO only. After incubation at 37˚C for a specified time, cells were harvested by using 0.05% trypsin and were then washed twice with cold phosphate-buffered saline (PBS) and fixed with ice-cold 70% ethanol overnight at -20˚C. The cells were then washed twice again with cold PBS and then treated with 80 μg/ml RNase A and 50 μg/ml PI for 30 min. The stained cells were analyzed by flow cytometry (22,23). Approximately 10,000 cells were evaluated from each treatment. The cell cycle data were re-analyzed using ModFit software.
DAPI nuclear staining. The morphological changes of apop- tosis (chromatin condensation) were assessed by staining with DAPI (Molecular Probes, Eugene, OR, USA). The DU 145 cells at a density of 1x10
5cells/well were exposed to various doses of BITC before the cells were fixed in 4% para- formaldehyde-PBS solution for 15 min and stained with DAPI (300 nmol/l) for 30 min at room temperature. Then nuclear morphology was visualized and photographed by fluorescence microscopy. Apoptotic cells were recognized by the condensed, fragmented and degraded nuclei (22,24).
Comet assay. The alkaline comet assay was performed as previously described by Del Bo et al and Riso et al (25,26).
The DU 145 cells at a density of 2x10
5cells/well were exposed to various doses of BITC before the cells were agarose-embedded and spread out on fully frosted slides. One slide was subjected to a treatment with H
2O
2(positive control) for 5 min, where the cells were then kept for 40 min in electrophoresis buffer before the electrophoresis was performed at 25 V, 300 mA (1.1 V/cm) for 20 min. All slides were stained with PI (5 μg/ml), washed in PBS, drained and covered with coverslips and were then examined and photo- graphed under a fluorescence microscope as described previously (23,25,27). The dispersion of DNA from the nucleus of each cell was measured by the lengths or para- meters of tail moments.
Determinations of ROS, mitochondrial membrane potential (Δ
Ψm) and intracellular Ca
2+levels. The levels of ROS, Δ
Ψm
LIU et al: BITC INDUCES APOPTOSIS IN HUMAN PROSTATE CANCER CELLS
788
787-796.qxd 26/1/2011 09:54 Ì ™ÂÏ›‰·788
and Ca
2+were measured by flow cytometric assay as described previously (23,28). DU 145 cells (2x10
5cells/well) were placed on 12-well plates for 24 h and then 0 and 10 μM of BITC were individually added to the wells prior to incubation for various time periods. At the end of incubation, the cells from each treatment group were trypsinized and centrifuged and then collected and washed twice with PBS, re-suspended in 500 μl of H
2DCF-DA (10 μM) for ROS, 500 μl of DiOC
6(1 μmol/l) for Δ
Ψm and 500 μl of Fluo-3/AM (2.5 μg/ml) for intracellular Ca
2+. All samples were incubated with these specific fluorochromes at 37˚C for 30 min and the differ- ential expressions from each treatment were analyzed by flow cytometry (23).
Assays for caspase-3, -8 and -9 activity. Caspase-3, -8 and -9 activities were determined by flow cytometric assay as described by Wu et al (22). In brief, DU 145 cells (2x10
5cells/well) were plated on 12-well plates for 24 h and then 0 and 10 μM of BITC were individually added to the wells for incubation for 0, 12, 24 and 48 h. At the end of incubation, all the cells were trypsinized and centrifuged, collected and washed twice with PBS. All samples were re-suspended in 50 μl of 10 μM substrate solution (PhiPhiLux-G
1D
1for caspase-3, CaspaLux-L
1D
2for caspase-8 and CaspaLux- M
1D
2for caspase-9) (OncoImmunin, Inc. Gaithersburg, MD, USA) before being incubated at 37˚C for 60 min. All samples
were washed again with PBS and were analyzed by flow cytometry as described previously (23).
Caspase-3 inhibitors and N-acetylcysteine (NAC) affect the BITC-decreased viability of DU 145 cells. Cultured DU 145 cells were pre-treated with Z-DEVE-FMK (a caspase-3 inhibitor) or NAC (an antioxidant) for 3 h and were then treated with or without 10 μM of BITC for 24 h. The cells from each treatment group were harvested and were analyzed for cell viability as described previously (23).
Western blotting analysis. DU 145 cells were seeded at a density of 1x10
6cells/ml in RPMI-1640 medium with 10%
fetal bovine serum for 24 h. Cells were treated with 0 and 10 μM of PEITC for 6, 12, 24 and 48 h. Cells from each treatment group were collected and protein was extracted into a high-salt buffer containing 1% protease inhibitor cocktail from Sigma-Aldrich, sonicated and centrifuged at 12,000 rpm for 10 min at 4˚C. Protein concentrations were determined by the BCA Protein Assay kit and Bradford assay (23,24). Cell lysates were treated with reducing sample buffer and boiled for 5 min at 100˚C. Proteins were resolved on an SDS polyacrylamide gel via electrophoresis. After the gel was transferred into a PVDF membrane in Western transfer buffer, the membrane was stained with primary antibody (anti-CDC25c, Wee1, p53, Fas, Fad ligand, Bcl-2,
Figure 1. BITC induces cell morphological changes and decreases the viability in DU 145 cells. Cells were cultured in RPMI-1640 + 10% FBS with various doses of BITC for 24 and 48 h. (A) Then the cells were examined for morphological changes and were photographed by phase-contrast microscopy (x200). (B) The percentage of cell viability was measured by flow cytometric assays, as described in Materials and methods. ***p<0.001, indicates significant difference between the control and PEITC-treated groups.
Bak, Bid, PARP, cytochrome c, AIF, GRP78 and GADD153) and then washed, followed by appropriate horseradish peroxi- dase-conjugated secondary antibodies (GE Healthcare). The intensity of the immunoreactive bands was determined using a densitometer (Molecular Dynamics, Sunnyvale, CA) equipped with ImageJ NIH software.
Immunofluorescence staining and confocal laser scanning microscopy. DU 145 cells (5x10
4cells/well) were maintained on 4-well chamber slides before being treated with 10 μM of BITC for 24 h. All samples were fixed in 3% formaldehyde in PBS for 15 min and permeabilized with 0.1% Triton X-100 in PBS for 1 h with the blocking of non-specific binding sites using 2% BSA as described previously (29). All fixed cells on the slides were stained with anti-AIF and anti-Endo G (1:200 dilution) as the primary antibodies overnight and were then stained with FITC-conjugated goat anti-mouse IgG at 1:100 dilution) (green fluorescence) as the secondary antibody, followed by nuclei counterstaining with PI (red fluorescence). All samples were photomicrographed using a Leica TCS SP2 confocal spectral microscope (23,30).
Statistical analysis. Results are expressed as the means ± standard error (SD) of the mean of three independent experi- ments, each conducted in triplicate. Data were analyzed by one-way Student's t-test. Differences were considered to be statistically significant at values of p<0.05, p<0.01 and p<0.001 when comparing the control to the BITC treatment.
Results
BITC induces morphological changes and decreases the percentage of DU 145 viable cells. The morphological changes of DU 145 cells in response to BITC were investi- gated. Incubation for 24 h with 2.5-15 μM of BITC revealed condensation both in the cytoplasm and the nucleus, thereby resulting in granulation in the majority of cells (Fig. 1A). The treatment with BITC further morphological changes and also to more cells floating in the well compared to the control.
After the cells were harvested and measured, the percentage of viable cells revealed that BITC decreased the percentage of viability. After 24- and 48-h incubation, the cell viability at concentrations of 2.5-15 μM BITC was 11-38% and 42- 80%, respectively (Fig. 1B), showing a dose-and time- dependent cytotoxic effect (Fig. 1B).
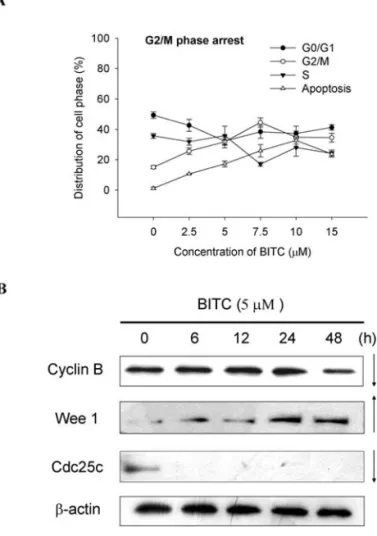
BITC induces cell cycle arrest in DU 145 cells. To further investigate the inhibition of proliferation of DU 145 cells in the presence of BITC, the cell cycle progression was examined by flow cytometry after the cells were treated with various doses of BITC and the results are shown in Fig. 2A.
The cells exposed to 2.5-7.5 μM BITC significantly increased the accumulation of the DNA contents up to 22- 45% in the G2/M phase in contrast to the control (Fig. 2A).
However, when the cells were exposed to 15 and 20 μM BITC, the accumulation of the DNA contents also increased significantly up to 7.6 and 27.7%, respectively, in the G2/M phase. However, the DNA accumulation in the sub-G1 phase was increased when the doses of BITC increased in contrast to the control (Fig. 2A), thus showing dose-dependent
effects. Based on the results (Fig. 2B) from Western blotting, BITC promoted the Wee1 levels and inhibited the levels of cyclin B and CDC25c, which led to the inhibition of the G2/M phase in cell cycle progression.
BITC induces apoptosis and DNA damage in DU 145 cells.
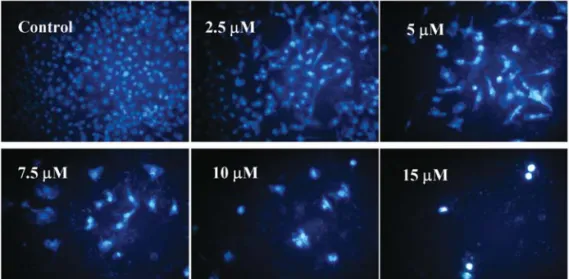
The induction of apoptosis by BITC in DU 145 cells was further confirmed in the fluorescence photomicrographs of the DU 145 cells stained with DAPI after the treatment with 2.5-15 μM BITC for 24 h. The condensation of chromatin is one of the morphological features of apoptosis examined and the results shown in Fig. 3A indicate that the control cells are round and homogeneous nuclei, whereas the BITC- treated cells show condensed and fragmented nuclei (Fig.
3A). As shown in Fig. 3A, only 2.5-10 μM BITC can induce a significant increase in apoptosis in DU 145 cells and these effects are dose-dependent. The induction of DNA damage by BITC in DU 145 cells was confirmed with a fluorescence photomicrograph of DU 145 cells stained with PI after treatment with 2.5-15 μM BITC for 24 h. The DNA damage was examined by comet assay and the results are shown in Fig. 3B, indicating that BITC induces DNA damage (comet tail occur) and that these effects are dose-dependent (Fig. 3B).
LIU et al: BITC INDUCES APOPTOSIS IN HUMAN PROSTATE CANCER CELLS
790
Figure 2. BITC induces cell cycle arrest and affects associated protein in DU 145 cells. Cells were placed in RPMI-1640 + 10% FBS with various doses of BITC for 48 h and were then harvested for analysis of cell cycle distribution (A) by flow cytometric assay or were examined for the protein associated with G2/M arrest by Western blotting (B) as described in Materials and methods.
787-796.qxd 26/1/2011 09:54 Ì ™ÂÏ›‰·790
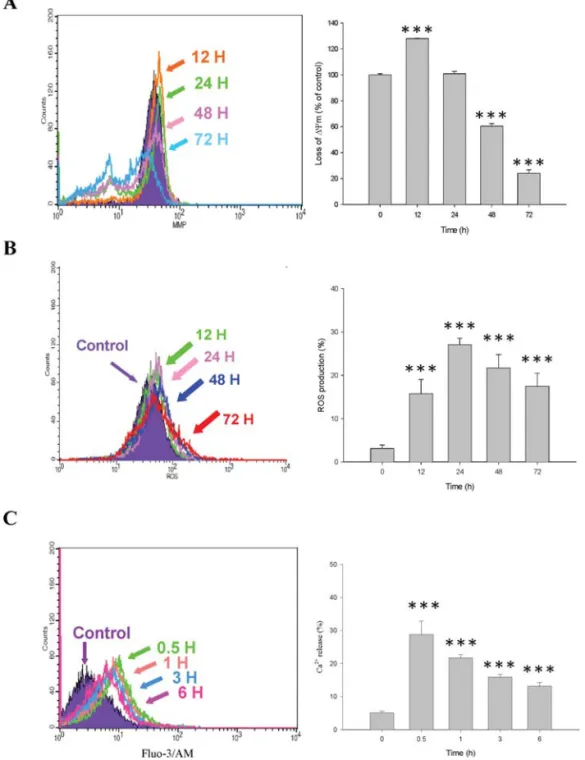
BITC decreases the levels of mitochondrial membrane potential Δ
Ψm) and increases the productions of ROS and Ca
2+in DU 145 cells. The levels of Δ
Ψm, ROS and Ca
2+in DU 145 cells after exposure to BITC were measured by flow cytometry. DU 145 cells were treated with BITC for various time periods and were then harvested in order to measure the levels of Δ
Ψm, ROS and Ca
2+, and the results are shown in Fig. 4A-C. Fig. 4A shows that BITC significantly decreases the levels at 48 and 72 h of treatment (Fig. 4A) and promotes the ROS production at 12 and 72 h of treatment (Fig. 4B).
However, BITC promotes the Ca
2+production during the early stages of treatment (0.5 h) and for up to 6 h and this effect is time-dependent (Fig. 4C).
BITC affects the activities of caspase-3, -8 and -9 in DU 145 cells. The activities of caspase-3, -8 and -9 in DU 145 cells were determined by flow cytometry. DU 145 cells were treated with BITC for various time periods and were then harvested in order to measure the activities of caspase-3, -8
and -9 and the results are shown in Fig. 5A-C. The results indicate that BITC increases the activities of caspase-3 at 12-72 h in a time-dependent manner (Fig. 5A), and promotes the caspase-8 and -9 activities at 48 h of treatment (Fig. 5B and C). Based on these observations, we suggest that caspases are involved in the induction of apoptosis by BITC in DU 145 cells.
The NAC antioxidant and caspase-3 inhibitors reduce the ability of BITC to decrease the percentage of viable DU 145 cells. DU 145 cells were pre-treated with NAC (an anti- oxidant agent) and the caspase-3 inhibitor (Z-DEVE-FMK) before the cells were harvested for the measurement of the percentage of viability and the results are shown in Fig. 6A and B. DU 145 cells were pre-treated with NAC to decrease the cytotoxic effects of BITC and this led to an increase in the percentage of viable cells (Fig. 6A). Z-DEVE-FMK significantly promotes the percentage of viable cells (Fig.
6B). Based on both observations, it is evident that BITC
Figure 3. BITC induces apoptosis and DNA damage in DU 145 cells. Cells were treated with BITC at various doses and were then incubated for 24 h. After incubation, cells in each treatment group were stained by DAPI and were then examined and photographed under a fluorescence microscope to examine the apoptosis (A) or were stained with PI to examine the DNA damage by comet assay (B) as described in Materials and methods.
decreases the percentage of viable DU 145 cells through ROS production and the caspase-3 dependent pathway.
BITC affects the apoptotic-associated protein levels in DU 145 cells. We examined the apoptotic associated protein levels by Western blotting. DU 145 cells were treated with 10 μM BITC and were then incubated for various time periods and the cells were harvested for Western blotting. The results are shown in Fig. 7A and B. The results indicate that BITC promotes the protein levels of Bid and AIF (Fig. 7A), GRP78, IRE1a and caspase-12 (Fig. 7B) but decreases the levels of PARP and XIAP (Fig. 7A).
BITC affects the distribution in DU 145 cells. We investi- gated the effects of BITC on the levels and distribution of
apoptotic-associated protein (AIF and Endo G) in DU 145 cells which were examined with a confocal laser microscope.
The results are shown in Fig. 8A and B. Fig. 8 indicates that BITC promotes the protein levels of AIF (Fig. 8A) and Endo G (Fig. 8B) and also promotes them to translocate to the nuclei.
AIF and Endo G were released from mitochondria and then trafficked to the nuclei, causing apoptosis. These observa- tions demonstrate that BITC induces apoptosis through the mitochondrial dependent pathway.
Discussion
Prostate cancer is one of the leading causes of death in the male population. Many dietary natural substances have been found to induce apoptosis in various tumor cell lines, and
LIU et al: BITC INDUCES APOPTOSIS IN HUMAN PROSTATE CANCER CELLS
792
Figure 4. BITC affects the levels of mitochondria membrane potential ΔΨm), ROS production and Ca2+and in DU 145 cells. Cells were treated with 10 μM of BITC for various time periods and were then collected and stained by DiOC6to determine the ΔΨm levels (A), or with 2,7-dichlorodihydrofluorescein diacetate to determine ROS production (B), or with Indo 1/AM to determine Ca2+levels (C), as described in Materials and methods. Staining with DiOC6for the determination of ΔΨm levels revealed a significant difference between the control and BITC-treated groups (***p<0.001).
787-796.qxd 26/1/2011 09:54 Ì ™ÂÏ›‰·792
thus, could be useful for the treatment of cancer complica- tions (31). Many studies have focused on the manipulation of the apoptotic process to treat and prevent cancer. Furthermore, more compounds were investigated for their influence on apoptosis in order to understand their mechanisms of action (32,33). BITC is one of the compounds found in cruciferous vegetables and has been shown to induce apoptosis in many human cancer cell lines (15-18). However, the characteristic chemical compounds of BITC against prostate cancer have not been well defined.
Herein, we provide evidence showing that BITC inhibits the tumor cell growth of the prostate cancer cell line, DU 145, at lower IC
50values (Fig. 1). It is well known that apoptosis
is modulated by complex pathways that involve a series of biochemical regulators including pro-apoptotic and anti- apoptotic proteins and molecular interactions (33). Flow cytometry analysis of DU 145 cells after exposure to BITC showed a distinct sub-G1 peak cell cycle distribution (Fig. 2A) which indicates that BITC induces apoptotic cell death.
Apoptotic cell death was also confirmed by DAPI staining in the DU 145 cells after exposure to various doses of BITC (Fig. 3A). The BITC-induced DNA damage in DU 145 cells was also confirmed by comet assay (Fig. 3B).
Caspases play an important role in the process of apop- tosis and in certain cases, caspases are essential for the induction of apoptosis (34). Therefore, we examined the
Figure 5. BITC induce caspase-3, -8 and -9 activity in DU 145 cells. DU 145 cells were treated with 10 μM of BITC for various time periods, and cells were collected for the determination of the activities of caspase-3 (A), caspase-8 (B) and caspase-9 (C) as described in Materials and methods. Data represent the means ± SD of three experiments. *p<0.05 and ***p<0.001, indicate significant difference between the control and BITC-treated groups.
activities of caspase-3, -8 and -9 in DU 145 cells. The results indicate that BITC promotes the activities of caspase-3, -8 and -9 (Fig. 5A-C). In particular, BITC induced the activation of caspase-3 in a time-dependent manner. We also utilized a caspase-3 inhibitor (Z-DEVE-FMK) system, which resulted in the inhibition of the apoptotic cell death induced by BITC.
This means that the pre-treatment of cells with Z-DEVE- FMK prior to treatment with BITC leads to an increase in the percentage of viable cells (Fig. 6A). Based on these observa- tions, it can be concluded that the compound BITC induces apoptosis in the prostate cancer cell line DU 145.
Other studies have demonstrated that mitochondria play an important role in the commitment of cells to apoptosis (35), and in order to elucidate the molecular mechanism responsible for the BITC-induced apoptosis in DU 145 cells, the role of mitochondria in BITC-induced apoptosis was investigated. In this study, BITC exhibited significant mito- chondrial membrane potential Δ
Ψm) disruption (Fig. 5A). It is known that AIF and Endo G were released from the mitochondria and this led to apoptosis which means that the agent induced apoptosis through the mitochondria (36). The present data show that BITC induces AIF and Endo G release from the mitochondria to the cytosol (Fig. 7A and B). This
finding suggests that the effect of the AIF and Endo G products via the mitochondria could be responsible for the modulation of BITC-induced apoptosis in the DU 145 cells.
It has been reported that aberrant mitochondria can generate intracellular ROS and that excessive ROS can induce oxidative stress which can cause DNA injury, resulting in growth arrest and apoptosis (36). Furthermore, it has been reported that the enhancement of ROS production involved with the apoptotic response is induced by anti-cancer agents (37). Exploring the possible oxidant-triggered molecular mechanisms underlying the DU 145 cell apoptosis by BITC, we found that the pro- apoptotic activity of BITC was accompanied by the accumu- lation of ROS, such as hydrogen peroxide (H
2O
2) (Fig. 5A), suggesting that the DU 145 cell death by BITC is a ROS- dependent process.
Our results from confocal laser microscopy indicate that BITC induces the release of AIF and Endo G from the mitochondria and movement into the nuclei (Fig. 8A and B).
These results suggest that the mitochondrial translocation of AIF and Endo G contributes to the BITC-induced mito- chondrial dysfunction and apoptosis. Mitochondria appearance is the intracellular source of ROS and it could also be a primary target for the damaging effects of ROS. In this study, time-course experiments showed that the elevation of ROS generation in DU 145 cells occurred as early as 12 h post- BITC exposure, indicating that this event occured earlier than mitochondrial membrane potential disruption and apoptotic execution (Fig. 4A-C). The BITC-triggered mitochondrial disruption, cytochrome c release, caspase activation and mitochondrial translocation of AIF and Endo G in DU 145
LIU et al: BITC INDUCES APOPTOSIS IN HUMAN PROSTATE CANCER CELLS
794
Figure 6. The NAC antioxidant and caspase-3 inhibitors affect the BITC- induced cytotoxicy in DU 145 cells. Cells were pre-treated with NAC (antioxidant agent) (A) or caspase-3 inhibitor ( Z-DEVE-FMK) (B) and were then treated with 10 μM BITC for 24 h. The cells were then collected for the determination of the percentage of viable cells as described in Materials and methods. Data represent the means ± SD of three experiments.
*p<0.05 and ***p<0.001, indicate significant difference between the control and BITC-treated groups.
Figure 7. BITC affects the levels of apoptotic-associated proteins in DU 145 cells. DU 145 cells were treated with 10 μM of BITC for 0, 6, 12, 24 and 48 h before the cytosolic fraction and total proteins were prepared and determined, as described in Materials and methods. The levels of associated protein expressions (A) Bid, PARP, AIF and XIAP and (B) GRP78, IRE1 and caspase-12 were estimated by Western blotting, as described in Materials and methods.
787-796.qxd 26/1/2011 09:54 Ì ™ÂÏ›‰·794
Figure 8. BITC affects AIF and Endo-G distributions in DU 145 cells. Cells were incubated with 10 μM of BITC for 24 h and were then fixed and stained with primary antibodies against AIF (A) and Endo-G (B), and were then stained with FITC-labeled secondary antibodies (green fluorescence) and the proteins were detected by a confocal laser microscopic system. The nuclei were stained with PI (red fluorescence). Areas of co-localization between the AIF and Endo G expressions and the cytoplasm and nuclei in the merged panels are shown in yellow. Scale bar, 10 μm.
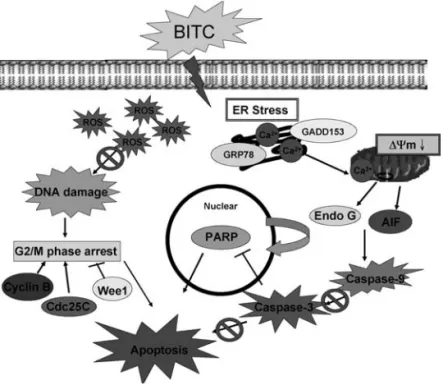
Figure 9. The proposed signaling pathways of the BITC-induced G2/M arrest and apoptosis in DU 145 cells.
cells is dependent on ROS generation. When the cells were pre-treated with NAC (antioxidant agent) prior to treatment with BITC, this led to an increase in the percentage of viable cells.
In conclusion, this study shows for the first time that BITC has anti-cancer activity in a human prostate cancer cell line (DU 145) through a ROS-provoked mitochondrial apoptotic pathway and the possible signaling pathways are shown in Fig. 9. These data provide a basic mechanism for the chemotherapeutic properties of BITC in prostate cancer cells.
Acknowledgements
This study was supported by the grant CMU98-Asia-10 from China Medical University, Taichung, Taiwan.
References
1. Zobel K, Wang L, Varfolomeev E, et al: Design, synthesis, and biological activity of a potent Smac mimetic that sensitizes cancer cells to apoptosis by antagonizing IAPs. ACS Chem Biol 1: 525-533, 2006.
2. Fesik SW: Promoting apoptosis as a strategy for cancer drug discovery. Nat Rev Cancer 5: 876-885, 2005.
3. Singh RP, Agrawal P, Yim D, Agarwal C and Agarwal R:
Acacetin inhibits cell growth and cell cycle progression, and induces apoptosis in human prostate cancer cells: structure-activity relationship with linarin and linarin acetate. Carcinogenesis 26:
845-854, 2005.
4. Yu SW, Wang H, Poitras MF, et al: Mediation of poly(ADP- ribose) polymerase-1-dependent cell death by apoptosis-inducing factor. Science 297: 259-263, 2002.
5. Hail N Jr, Kim HJ and Lotan R: Mechanisms of fenretinide- induced apoptosis. Apoptosis 11: 1677-1694, 2006.
6. Sun SY, Hail N Jr and Lotan R: Apoptosis as a novel target for cancer chemoprevention. J Natl Cancer Inst 96: 662-672, 2004.
7. Ferreira CG, Epping M, Kruyt FA and Giaccone G: Apoptosis:
target of cancer therapy. Clin Cancer Res 8: 2024-2034, 2002.
8. Hengartner MO: The biochemistry of apoptosis. Nature 407:
770-776, 2000.
9. Liu H, Dong A, Gao C, Tan C, Zu X and Jiang Y: The design, synthesis, and anti-tumor mechanism study of N-phosphoryl amino acid modified resveratrol analogues. Bioorg Med Chem 16: 10013-10021, 2008.
10. Gao C, Jiang Y, Tan C, Zu X, Liu H and Cao D: Synthesis and potent antileukemic activities of 10-benzyl-9(10H)-acridinones.
Bioorg Med Chem 16: 8670-8675, 2008.
11. Verhoeven DT, Goldbohm RA, van Poppel G, Verhagen H and van den Brandt PA: Epidemiological studies on brassica vegetables and cancer risk. Cancer Epidemiol Biomarkers Prev 5: 733-748, 1996.
12. Cohen JH, Kristal AR and Stanford JL: Fruit and vegetable intakes and prostate cancer risk. J Natl Cancer Inst 92: 61-68, 2000.
13. Conaway CC, Yang YM and Chung FL: Isothiocyanates as cancer chemopreventive agents: their biological activities and metabolism in rodents and humans. Curr Drug Metab 3: 233-255, 2002.
14. Warin R, Chambers WH, Potter DM and Singh SV: Prevention of mammary carcinogenesis in MMTV-neu mice by cruciferous vegetable constituent benzyl isothiocyanate. Cancer Res 69:
9473-9480, 2009.
15. Xiao D, Vogel V and Singh SV: Benzyl isothiocyanate-induced apoptosis in human breast cancer cells is initiated by reactive oxygen species and regulated by Bax and Bak. Mol Cancer Ther 5: 2931-2945, 2006.
16. Zhang Y, Tang L and Gonzalez V: Selected isothiocyanates rapidly induce growth inhibition of cancer cells. Mol Cancer Ther 2: 1045-1052, 2003.
17. Tseng E, Scott-Ramsay EA and Morris ME: Dietary organic isothiocyanates are cytotoxic in human breast cancer MCF-7 and mammary epithelial MCF-12A cell lines. Exp Biol Med (Maywood) 229: 835-842, 2004.
18. Xiao D, Powolny AA and Singh SV: Benzyl isothiocyanate targets mitochondrial respiratory chain to trigger reactive oxygen species-dependent apoptosis in human breast cancer cells. J Biol Chem 283: 30151-30163, 2008.
19. Miyoshi N, Takabayashi S, Osawa T and Nakamura Y: Benzyl isothiocyanate inhibits excessive superoxide generation in inflammatory leukocytes: implication for prevention against inflammation-related carcinogenesis. Carcinogenesis 25: 567-575, 2004.
20. Ban JY, Nguyen HT, Lee HJ, et al: Neuroprotective properties of gallic acid from Sanguisorbae radix on amyloid beta protein (25-35)-induced toxicity in cultured rat cortical neurons. Biol Pharm Bull 31: 149-153, 2008.
21. Lin SS, Huang HP, Yang JS, et al: DNA damage and endo- plasmic reticulum stress mediated curcumin-induced cell cycle arrest and apoptosis in human lung carcinoma A-549 cells through the activation caspases cascade- and mitochondrial- dependent pathway. Cancer Lett 272: 77-90, 2008.
22. Wu SH, Hang LW, Yang JS, et al: Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways. Anticancer Res 30: 2125-2133, 2010.
23. Yang JS, Hour MJ, Huang WW, Lin KL, Kuo SC and Chung JG:
MJ-29 inhibits tubulin polymerization, induces mitotic arrest, and triggers apoptosis via cyclin-dependent kinase 1-mediated Bcl-2 phosphorylation in human leukemia U937 cells. J Pharmacol Exp Ther 334: 477-488, 2010.
24. Ji BC, Hsu WH, Yang JS, et al: Gallic acid induces apoptosis via caspase-3 and mitochondrion-dependent pathways in vitro and suppresses lung xenograft tumor growth in vivo. J Agric Food Chem 57: 7596-7604, 2009.
25. Del Bo C, Martini D, Vendrame S, et al: Improvement of lymphocyte resistance against H(2)O(2)-induced DNA damage in Sprague-Dawley rats after eight weeks of a wild blueberry (Vaccinium angustifolium)-enriched diet. Mutat Res: Aug 26, 2010 (Epub ahead of print).
26. Riso P, Martini D, Visioli F, Martinetti A and Porrini M: Effect of broccoli intake on markers related to oxidative stress and cancer risk in healthy smokers and nonsmokers. Nutr Cancer 61: 232-237, 2009.
27. Riso P, Pinder A, Santangelo A and Porrini M: Does tomato consumption effectively increase the resistance of lymphocyte DNA to oxidative damage? Am J Clin Nutr 69: 712-718, 1999.
28. Lu CC, Yang JS, Huang AC, et al: Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Mol Nutr Food Res 54:
967-976, 2010.
29. Chen JC, Lu KW, Tsai ML, et al: Gypenosides induced G0/G1 arrest via CHk2 and apoptosis through endoplasmic reticulum stress and mitochondria-dependent pathways in human tongue cancer SCC-4 cells. Oral Oncol 45: 273-283, 2009.
30. Lo C, Lai TY, Yang JH, et al: Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int J Oncol 37: 377-385, 2010.
31. Schwartz M, Roayaie S and Konstadoulakis M: Strategies for the management of hepatocellular carcinoma. Nat Clin Pract Oncol 4: 424-432, 2007.
32. Banerjee S, Wang Z, Mohammad M, Sarkar FH and Mohammad RM: Efficacy of selected natural products as therapeutic agents against cancer. J Nat Prod 71: 492-496, 2008.
33. Kurokawa M and Kornbluth S: Caspases and kinases in a death grip. Cell 138: 838-854, 2009.
34. Ghavami S, Hashemi M, Ande SR, et al: Apoptosis and cancer:
mutations within caspase genes. J Med Genet 46: 497-510, 2009.
35. Balaban RS: Modeling mitochondrial function. Am J Physiol Cell Physiol 291: C1107-C1113, 2006.
36. Zaja-Milatovic S, Gupta RC, Aschner M and Milatovic D:
Protection of DFP-induced oxidative damage and neuro- degeneration by antioxidants and NMDA receptor antagonist.
Toxicol Appl Pharmacol 240: 124-131, 2009.
37. Visconti R and Grieco D: New insights on oxidative stress in cancer. Curr Opin Drug Discov Devel 12: 240-245, 2009.
LIU et al: BITC INDUCES APOPTOSIS IN HUMAN PROSTATE CANCER CELLS
796
787-796.qxd 26/1/2011 09:54 Ì ™ÂÏ›‰·796