科技部補助專題研究計畫成果報告 期末報告
情感型疾病與代謝症候群交互作用之整合性研究-長期壓力對情 緒和代謝中樞神經可塑性的影響(2/2)
計 畫 類 別 : 整合型計畫
計 畫 編 號 : MOST 105-2321-B-006-019- 執 行 期 間 : 105年08月01日至106年07月31日
執 行 單 位 : 國立成功大學解剖學科暨細胞生物與解剖學研究所
計 畫 主 持 人 : 郭余民
計畫參與人員: 碩士級-專任助理:陳韻文
碩士班研究生-兼任助理:黃婉寧
報 告 附 件 : 出席國際學術會議心得報告
中 華 民 國 106 年 10 月 30 日
中 文 摘 要 : 流行病學研究指出,代謝型疾病以及情感型疾病間存在著相互關聯 性。而此二疾病間高度的併發性,暗示著此兩疾病間可能存有相似 的致病機轉。而在諸多代謝型疾病中,糖尿病是一表現長期高血糖 以及胰島素失能兩症狀之代謝型疾病,同時也是當今一個棘手的全 球性健康問題。而長期壓力除了是一個已知的情感型疾病成因之外
,近期也被認為是罹患糖尿病的高風險因子。然而,有關壓力與糖 尿病間交互作用的詳細機轉,至今仍少被著墨。此外,大腦中,杏 仁核為情緒處理中樞,專司處理壓力相關事件。但杏仁核是否參與 在調節壓力引發之代謝反應中,至今亦不清楚。為探究壓力對於糖 尿病發生過程的影響,於本研究中我們使用為期11週的高脂飲食餵 養,來於小鼠中誘發高血糖與胰島素阻抗,並採用為期10日的社交 挫敗來作為給予小鼠的壓力模式。結果顯示,在高脂飲食餵養前期 給予的社交挫敗壓力,可加劇高脂飲食所導致的胰島素阻抗,包括 提升空腹後血中胰島素含量,以及提高葡萄糖耐受性與胰島素耐受 性測試中的血糖濃度。爾後,我們進一步研究杏仁核於此些社交挫 敗壓力所誘發之反應中扮演的角色。我們發現若於社交挫敗壓力給 予之前,損壞雙側杏仁核之中央核,則可抑制前述之社交挫敗引起 之加劇效應。此外,社交挫敗壓力亦會增加杏仁核中,與神經再塑 相關的TrkB蛋白表現。綜合來看,我們的結果顯示,壓力會加劇高 脂飲食所導致之胰島素阻抗,亦會引發杏仁核中的神經再塑。與之 一致,我們也證實此壓力所引起之加劇效應是與杏仁核相關的。再 者,本研究也提出一個可供日後研究,中樞系統如何調節壓力加劇 代謝失調此一過程時,可使用的新穎動物模式。
中 文 關 鍵 詞 : 壓力、糖尿病、肥胖、杏仁核
英 文 摘 要 : Epidemiological studies indicate a bidirectional
association between metabolic disorders and mood disorders.
The concurrence of these two diseases suggests that these two diseases may share common pathophysiological
mechanisms. Among the metabolic disorders, diabetes which is mainly characterized by a prolonged elevation of blood sugar and insulin dysfunctions has become a critical global health issue. Moreover, chronic stress, a well-known cause for mood disorders, has been recently considered as an independent risk factor for diabetes. However, the
mechanism underlying the interactions between stress and diabetes is less explored. The amygdala is the emotional center responsible for processing the stress-associated events. However, whether amygdala is involved in regulating of the stress-related metabolic responses remains unclear.
To examine the effect of stress on the development of diabetes, we used a 11-week high-fat diet (HFD) feeding paradigm to induce hyperglycemia and insulin resistance in mice and exposed these mice with a 10-day social defeat (SD) stressor. Our results showed that mice given the SD at the beginning of the HFD feeding aggravated the HFD-induced insulin resistance (i.e., increasing the fasting insulin
level and elevating the blood glucose during glucose tolerance and insulin tolerance tests). We further investigated the role of amygdala in these SD-induced effects, and found that the bilateral lesions of the central amygdalae before the SD inhibited the stress- induced aggravation effect. Moreover, the SD stress also upregulated the expression of neuroadaptation-related protein, TrkB, in the amygdala of HFD mice. In conclusion, stress aggravated the HFD-induced insulin resistance and induced neuroadaptation in the amygdala of HFD mice. The stress-induced aggravation effect is amygdala-dependent.
This study also provides a new animal model to investigate the neural regulation underlying the stress-enhanced
metabolic dysregulation.
英 文 關 鍵 詞 : stress, diabetes, obesity, amygdala
Introduction
Type II diabetes (T2D) is a group of metabolic disorders which are characterized by insulin resistance and high blood glucose. The etiology of T2D is related to lifestyle and genetic factors. Obesity is the major risk factor of T2D. Chronic exposure to stressors is also known to increase the risk of T2D [1]. Long-term stress has been suggested to affect metabolism via changing eating behaviors and energy homeostasis in humans and animals [2, 3]. However, the underlying mechanism for stress to induce T2D remains unclear.
In the central nervous system, the amygdala is the emotional center responsible for processing the emotional reactions, and the formation and storage of memories associated with stress-associated events [4]. Anatomically, both emotional center (i.e.
amygdala) and metabolic center (i.e. hypothalamic paraventricular nucleus and arcuate nucleus) are located in the limbic system with direct neural circuit connections.
Upon stress stimulation, neurons in the central nucleus of amygdala activate and can excite one of its targets the hypothalamic paraventricular nucleus to trigger the hypothalamic-pituitary-adrenocortical (HPA) axis. Consequently, stress hormones are released to induce whole body stress responses including fuel liberation helping mammals to cope with stressors [5]. However, excess levels of stress hormones, e.g.
glucocorticoids, drive the intake of comfort foods in humans and rodents [6-8], and has been associated with the development of obesity [9, 10].
Long-term stress induces neuroadaptation in the amygdala. Single episode of stress facilitates fear memory and enhances the induction of long-term potentiation in the amygdala of rat [11]. Repeated stress enhances dendritic arborization and increases spine density of the basolateral amygdala neurons [12, 13]. However, whether the stress-induced amygdalar neuroadaptation is involved in the interplay between stress and T2D remains unclear.
Purpose
Therefore, the objective of this study is to investigate whether and how stress affects the T2D pathology in mice and characterizing the role of the stress-induced amygdalar neuroadaptations in this process.
To reach this goal, three specific aims were generated:
Aim 1: To examine the effects of stress on the T2D pathology in mice.
Aim 2: To examine the role of amygdala neuroadaptation in the stress-induced effects.
Aim 3: To examine the role of stress hormone responses in stress-induced effects.
Experimental Approaches
In this study, we fed young male C57BL/6N mice with a high-fat diet (HFD) for
11 weeks to induce hyperglycemia and insulin resistance, the two major features of T2D. To examine the effect of stress on the development of T2D, mice were exposed to social defeat (SD) stress in the early and late stages of HFD feeding. The expression of synaptic plasticity-related proteins, including TrkB, brain-derived neurotrophic factor (BDNF), synaptotagmin-1 (SYT-1), SYT-4 and SNAP-25 in the amygdala were used as indicators of neuroadaptation. In some mice, their central amygdalae were bilaterally lesioned before stress and HFD treatments. The effects of stress and amygdalar lesions on the HFD-induced insulin resistance were examined by measuring the fasting levels of plasma glucose and insulin, and by the glucose and insulin tolerance tests in these mice.
Materials and Methods
Animals
All experiments were performed in accordance with the National Institutes of Health Guideline for Animal research (Guide for the Care and Use of Laboratory Animals) and approved by the National Cheng Kung University Institutional Animal Care and Use Committee (IACUC approval number: 104211). All the mice used in this study were a C57BL/6N strain, obtained from National Cheng Kung University Laboratory Animal Center (http://www.ncku.edu.tw/animal/eng/nckulac.html). Mice were housed in pairs with a stable temperature (24 ± 1°C), a 12-h light/dark cycle, and unrestricted access to food and water. The housing environment and animal health were monitored by the Laboratory Animal Center. One day after the end of experiments, the mice were subjected to further tests or specimen collections.
High-fat diet feeding
All mice were initially fed a standard chow (laboratory autoclavable rodent diet 5010, LabDiet, St. Louis, MO, USA) until the start of the HFD feeding. Began at 8-week-old, half of the mice were fed a high-fat diet, which contains 60 kcal % fat (58Y1, TestDiet, St. Louis, MO, USA), while the other half of the mice were allowed continuous free-access to the standard chow diet. The HFD feeding lasted for 11 weeks (from 8 to 19 weeks old).
Social defeat stress
A 10-day SD stress was given to mice as previously described [14]. Briefly, male CD-1 retired breeder mice (4-6 months of age) with reliable attack latencies were individually housed in a compartment of a defeat cage which was divided into two separated compartments with a perforated Plexiglas separator. The defeated C57BL/6 mouse was first placed in the same compartment as the CD1 aggressor for 10 mins
and then transferred to the other compartment adjacent to its CD1 aggressor for the rest of the day. This procedure was repeated by exposing the defeated mouse to a novel CD1 aggressor each day, for 10 consecutive days. The non-defeat control mice were housed in pairs in a defeat cage with one mouse per component. All control mice were daily rotated in a manner similar to that of mice undergoing defeat. During the 10-day rotation, the non-defeat control mice were never allowed physical contact with their cage mate.
Measuring fasting plasma levels of glucose and insulin
After a 12-h fasting, mice were anesthetized by pentobarbital (1g/kg, i.p.) and their blood was collected from the retro-orbital sinus using heparinized capillary tubes.
Plasma was obtained after centrifuging the blood at 3,000×g for 10 min. The glucose levels were measured using a commercial glucose-oxidase kit (BioSystems). The insulin levels were measured using a commercial mouse insulin ELISA kit (code#10-1247-01, Mercodia, Uppsala, Sweden). The homeostasis model assessment-insulin resistance (HOMA-IR) index was calculated as follows: fasting glucose (mM) × fasting insulin (mU/l)/22.5.
Glucose and insulin tolerance tests
Intra-peritoneal glucose tolerance test (IPGTT) and intra-peritoneal insulin tolerance test (IPITT) were performed as previously described [15]. After a 12-hr fasting, the mice were intraperitoneally injected either with 1.0 g/kg glucose or 1.0 U/kg insulin. Blood samples were collected from the orbital sinus of the mice at 0, 30, 60, 90 and 120 min after the injection. Blood glucose levels were determined using a commercial glucose assay kit (code#11538, Biosystems, Barcelona, Spain).
Central amygdala lesion
Intra-amygdalar microinjection of ibotenic acid (IBO) was used to chemically lesion the central amygdala of mice [16]. One week before the start of diet feeding (at 7 weeks of age), anesthetized mice (pentobarbital, 1g/kg, i.p.) were mounted on a stereotaxic apparatus. IBO (20 μg each side, dissolved in 0.5 μl artificial cerebrospinal fluid) was bilaterally injected into the central amygdalae (AP: -0.16 mm; ML: ± 2.5 mm; DV: -4.75 mm from the bregma reference point) via a 30-gauge stainless steel needle. The mice were handled daily and given at least 7 days to recover. The mice with central amygdalar lesions were termed as IBO mice, and the mice injected with equal volume of artificial cerebrospinal fluid (aCSF) were served as control and termed as aCSF mice.
Tissue preparation for immunoblotting and Nissl staining
The anesthetized mice (pentobarbital, 1g/kg, i.p.) were perfused from the left ventricle with chilled phosphate-buffered saline, and their brains were removed. For immunoblotting, the amygdalae were quickly dissected out, freezing in liquid nitrogen, and stored at -80 °C until use. For Nissl staining, whole brains were post-fixed with 4% buffered paraformaldehyde at room temperature for 2 days, and then coronally sliced into 25 µm sections using a cryomicrotome.
Western blot
Immunoblotting for detecting the expression of TrkB, BDNF, SYT-1, SYT-4, and SNAP-25 in the central amygdala was performed as described previously [17].
Relative protein levels (versus β-actin) were estimated by immunoblotting using the following primary antibodies: TrkB (610102, Becton Dickinson, Franklin Lakes, NJ, USA), BDNF (sc-546, Santa Cruz Biotechnology, Santa Cruz, CA, USA), SYT-1 (SYA-130, Stressgen, Victoria, BC, Canada), SYT-4 (SC-30095, Santa Cruz Biotechnology) and SNAP-25 (VAM-SV012, Stressgen). The bound antibodies were detected using an enhanced chemiluminescence detection kit (PerkinElmer, Boston, MA, USA). The band densities were measured using an imaging system (BioChemi;
UVP, Upland, CA, USA) and analyzed using ImageJ Fiji software (version 1.51K) [18].
Nissl staining
Nissl staining was used to verify the successful lesion of the central amygdala.
The brain sections containing the central amygdalae were collected in the range, stereotaxic reference point, from 0.70 mm to 1.94 mm posterior to the Bregma. The brain sections mounted on the gelatin-coated slides were soaked in 1:1 alcohol/chloroform solution overnight to reduce staining background. Then, the brain sections were rehydrated through 100% and 95% alcohol to distilled water and stained with 0.1% cresyl violet solution for 10 minutes in 50 ºC oven. The stained sections were washed by quickly rinsing in distilled water, dehydrated through 100% alcohol to xylene, and mounted with permanent mounting medium.
Plasma corticosterone measurement
Blood specimens were collected using heparin as an anticoagulant. After centrifuging, the plasma was collected and the plasma level of corticosterone was quantified using a commercial ELISA kit (501320, Cayman, Ann Arbor, Michigan, USA).
Statistical analysis
Data are expressed as mean ± S.E.M. Significance was set at p < 0.05. The body weight gains and the plasma glucose levels in the glucose and insulin tolerance tests of mice were analyzed by repeated measured two-way ANOVAs with time (ages or experimental time points) and treatment as main factors. Sidak’s multiple comparisons were used to perform post hoc analysis after the two-way ANOVAs.
Other data sets were analyzed using one-way ANOVAs followed by Tukey’s multiple comparisons.
Results
Effect of high-fat diet on body weight and insulin sensitivity
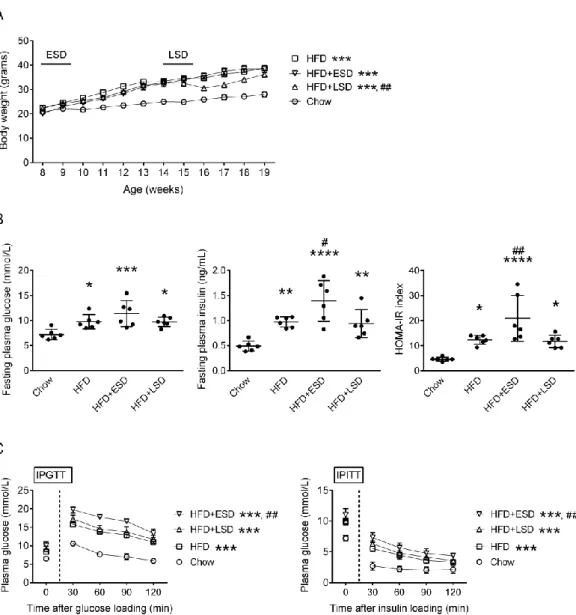
To determine the effect of HFD on systemic metabolism, we measured the body weight gain, fasting levels of plasma glucose and insulin, and glucose and insulin tolerance. HFD increased the body weight gain (F = 99.3, d.f. 1/132, p < 0.0001), and induced insulin resistance by elevating the fasting levels of plasma glucose (p < 0.05) and insulin (p < 0.01) as well as the HOMA-IR index (p < 0.05) in mice (HFD vs.
Chow, Figure 1A and 1B). HFD mice also showed impaired glucose utility and insulin resistance with higher plasma glucose levels during the IPGTT (F = 84.5, d.f.
1/24, p < 0.0001) and IPITT (F = 14.5, d.f. 1/36, p < 0.01), respectively (HFD vs.
Chow, Figure 1C).
Effect of stress on high-fat diet-induced metabolic dysregulations
To examine if stress affects the HFD-induced metabolic disorders, we exposed mice to a 10-day SD stress at the early (started from the 1st week of HFD feeding;
ESD group) or late (started from the 7th week of HFD feeding; LSD group) stages during the HFD feeding. ESD did not alter the HFD-induced body weight gains (p = 0.2) (HFD+ESD vs. HFD, Figure 1A), but enhanced the HFD-induced insulin resistance by increasing the fasting insulin level (p < 0.05) and HOMA-IR index (p <
0.01) (HFD+ESD vs. HFD, Figure 1B) and by elevating the plasma glucose levels during the IPGTT (F = 26.8, d.f. 1/18, p < 0.01) and IPITT (F = 5.1, d.f. 1/39, p < 0.05) (HFD+ESD vs. HFD, Figure 1C). On the other hand, HFD+LSD mice showed lower body weight gains than the HFD group, particularly following the exposure to SD stress (F = 10.1, d.f. 1/154, p < 0.01) (HFD+LSD vs. HFD, Figure 1A). Furthermore, LSD did not alter the HFD-induced insulin resistance (HFD+LSD vs. HFD, Figure 1B and 1C).
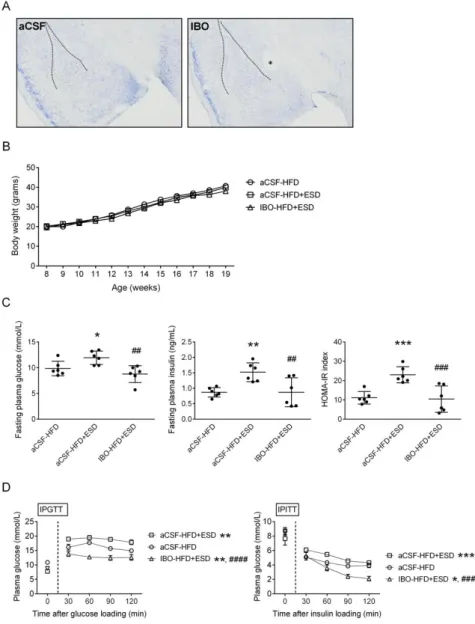
Effect of amygdala on stress-induced aggravation of high-fat diet-induced insulin resistance
To examine the involvement of neural regulation in the stress-induced aggravation of metabolic abnormalities in HFD mice, we performed bilateral intra-amygdalar injections of IBO to chemically lesions of the central amygdalae, the brain stress center (Figure 2A). The control mice received an equal volume of aCSF injections to the intra-amygdalae. Because only the ESD aggravated the HFD-induced metabolic abnormalities (Figure 1), IBO injections were given one week before the inductions of ESD. As shown in Figure 2B, neither ESD nor IBO injections altered the HFD-induced weight gain. However, ESD-induced aggravation of HFD-induced insulin resistance, including increased fasting levels of plasma glucose (p < 0.01) and insulin (p < 0.01), and HOMA-IR index (p < 0.001) (IBO-HFD+ESD vs.
aCSF-HFD+ESD, Figure 2C), as well as increased plasma glucose levels in the IPGTT (F = 46.4, d.f. 1/30, p < 0.0001) and IPITT (F = 28.1, d.f. 1/30, p < 0.001) (IBO-HFD+ESD vs. aCSF-HFD+ESD, Figure 2D) were inhibited by bilateral lesions of amygdalae. These results suggest that the central amygdala is essential for the stress-induced enhancements of the HFD-induced insulin resistance in mice.
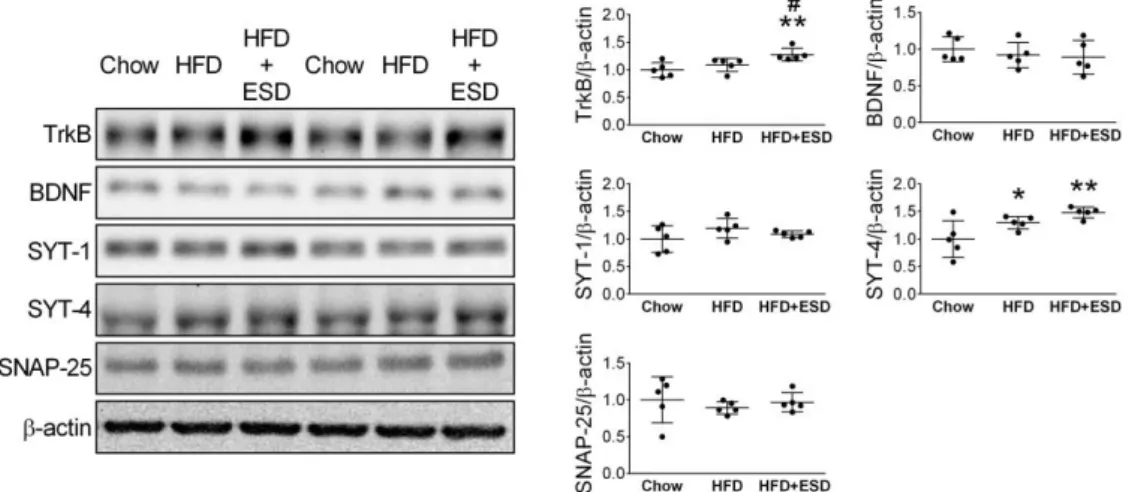
Effects of high-fat diet and social defeat on the expression of neuroplasticity-related proteins in the amygdala
Western blot was used to semi-quantify the expressions of neuroplasticity-related proteins, including the TrkB, BDNF, SYT-1, SYT-4, and SNAP-25, in the amygdala.
Results showed that HFD increased the amygdalar expression of SYT-4 (p < 0.05), but not TrkB (p = 0.3), BDNF (p = 0.5), SYT-1(p = 0.1) or SNAP-25 (p = 0.4) (HFD vs. Chow, Figure 3). Compared to the Chow mice, the HFD+ESD mice had higher expression of TrkB (p < 0.01) and SYT-4 (p < 0.01) in the amygdala (Figure 3).
Moreover, the amygdalar levels of TrkB in the HFD+ESD mice were also higher than those in the HFD mice (p < 0.05) (Figure 3).
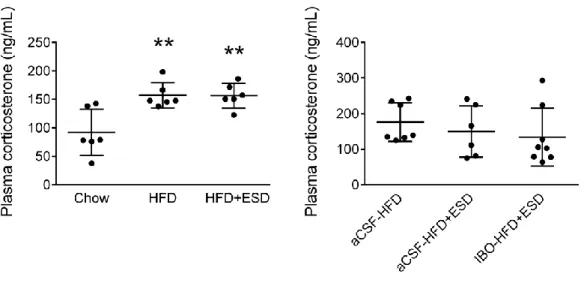
Effects of high-fat diet and social defeat on circulating level of corticosterone To examine if stress hormone is involved in the stress-induced aggravation of high-fat diet-induced insulin resistance, the levels of corticosterone, the primary rodent stress hormone, were measured in different groups of mice. Plasma levels of corticosterone in the HFD (p < 0.01) and HFD+ESD (p < 0.01, SD-HFD vs. Chow) groups were higher than those of the Chow group; there was no difference between the HFD and HFD+ESD groups (p = 1.0) (Figure 4, left panel). Furthermore, bilateral amygdalar lesions did not alter plasma levels of corticosterone in the HFD+ESD mice (p = 0.7, Figure 4).
Discussions and Conclusions
This study aimed to determine the effects of chronic stress on the HFD-induced metabolic dysregulations and whether the stress-induced amygdalar neuroadaptation is involved in this phenomenon. Our results showed that only when the SD stress was given to mice at the early stage, but not at the late stage, of the HFD feeding period aggravated the HFD-induced insulin resistance. Bilateral lesions of the central amygdalae before the ESD and the HFD feeding abolished the stress-induced aggravation of the HFD-induced insulin resistance. Both the HFD feeding and the ESD altered the expressions of some of the neuroplasticity-related proteins in the amygdala, but only the HFD feeding changed plasma levels of corticosterone. These findings suggest that chronic stress augments the diet-induced insulin resistance; this event is amygdala-dependent, but corticosterone-independent. This study also suggests a new animal model for investigating the neural regulation underlying the stress-enhanced metabolic dysregulation.
Accumulated evidence suggests that chronic stress is an independent risk factor for the developments of T2D [1]. However, mechanism underlying the interaction between psychological stress and insulin resistance is still unclear, although a few pioneering studies have shed light on this puzzle. Li et al. reported that acute stress could lead to a rapid development of insulin resistance [19]. Mice that were exposed to a 180-episode inescapable foot shock had increased blood glucose levels in both the glucose tolerance and insulin tolerance tests. This acute stress impaired the insulin signaling pathway in the liver, but not in the adipose, muscle or brain (i.e., hypothalamus, hippocampus, and amygdala) tissues [19]. They concluded that psychological stress acutely alters hepatic insulin sensitivity and affects the systemic glucose metabolism in normal mice [19]. In agreement with their findings, we found that chronic SD stress could enhance the HFD-induced insulin resistance. We also found that the central amygdala is necessary for such effect. Various forms of chronic stressors are known to induce neuroplasticity changes in the amygdala and affect downstream sympathetic outputs [17, 20]. Together, these findings suggest that stress aggravates HFD-induced insulin resistance and the stress-induced neuroplasticity in the central amygdala is involved in this aggravation effect.
Chronic activation of HPA axis contributes to the development of metabolic disorders [10, 21]. In rodents, excess corticosterone induces obesity [10] and insulin resistance [21]. We found that HFD along increased levels of plasma corticosterone, which suggests the critical role of stress hormone in the development of metabolic disorders. This proposition is supported by the findings that HFD and HFD+ESD mice had similar levels of plasma corticosterone and comparable body weight gains.
However, both the ESD and lesions of amygdalae did not alter levels of plasma
corticosterone in the HFD mice, but the ESD positively and bilateral lesions of amygdalae negatively affected the FHD-induced insulin resistance. These findings suggest that the HPA axis is not involved in the chronic stress-induced aggravation of HFD-induced insulin resistance. In addition to the HPA axis, stress could also activate the sympatho-adrenal medullary axis to modulate energy metabolism [20].
Sympathetic overactivity has been linked to the developments of insulin resistance [22, 23]. Overexciting sympathetic innervations can cause α-adrenergic vasoconstriction, which in turn impairs the muscular glucose uptake by blunting the postprandial increases of blood flows in the skeletal muscles, and stimulate additional insulin production by the pancreas leading to insulin resistance [24]. Moreover, excess epinephrine increases hepatic glucose production and inhibits the insulin-induced glucose uptake by tissues [25, 26]. These sympathetic overactivity-induced effects on the glucose metabolism are similar to the acute stress-caused outcomes reported by Li et al. [19]. Thus, SD may enhance the HFD-induced insulin resistance by strengthening the sympathetic outputs. it will be interesting to investigate this hypothesis in the future.
The differential effects of ESD and LSD on the aggravation of the HFD-induced insulin resistance deserve further discussion. It is possible that the HFD feeding period after the LSD is too short to display the stress effects. The HFD feeding period after the ESD was approximately 9.5 weeks, while the feeding period after the LSD was only about 3.5 weeks. Another possibility is that the stress-induced neuroadaptation is age dependent. Although C57BL/6J mice are sexually mature by 35 days, it has been recommended that mice should be at least 3 months old to be considered as mature adults (https://www.jax.org/research-and-faculty/research-labs/
the-harrison-lab/gerontology/life-span-as-a-biomarker). The 10-day ESD was given to mice at 8-week-old, which is about the late adolescent stage of mice life span.
Adolescence is a particular period of life when significant physiological and psychological changes occur. Intuitively such changes appear to arise from structural and functional changes in the brain. In adolescence, the brain reorganizes the synaptic connections and many neurotransmitter systems, and is more readily influenced by environmental factors, such as stress [27]. It has been demonstrated that social stress exerts opposite effects on the amygdalar structures and functions in the adolescent and adult rats [17]. It is possible that SD stress may differentially affect the amygdalar neurotransmissions, which are involved in the modulation of the systemic glucose metabolism in the adolescent and adult HFD mice. As a result of that, the stress-enhanced metabolism dysregulation only becomes manifestation in those mice with stress exposure at the adolescent or younger ages. This scenario is supported by recent reports addressing the links between psychological stress and early-onset T2D
[28-30]. Our mouse model is an appropriate tool to investigate the role of age-dependent stress experience in the pathogenesis of metabolic disorders.
Conclusion
Our findings indicated that chronic HFD feeding and SD stress alter the expressions of the neuroplasticity-related proteins in the amygdala. SD stress aggravates the HFD-induced insulin resistance, which is amygdala dependent, but corticosterone independent. The SD stress-induced aggravation seems to depend on the exposing age.
This study may also open a new corridor to investigate the neural regulation underlying the age-dependent stress-enhanced metabolic dysregulation.
Reference
[1] Annu Rev Public Health. 2015;36:441-62.
[2] Trends Endocrinol Metab. 2010;21:159-65.
[3] Neurosci Biobehav Rev. 2009;33:537-50.
[4] Nat Rev Neurosci. 2009;10:423-33.
[5] Annu Rev Neurosci. 2009;32:289-313.
[6] Am J Physiol. 1996;271:E317-25.
[7] Brain Behav Immun. 2005;19:275-80.
[8] Prog Brain Res. 2006;153:75-105.
[9] Stress. 2011;14:233-46.
[10] Endocrinology. 2000;141:560-3.
[11] J Neurosci. 2005;25:8725-34.
[12] Proc Natl Acad Sci U S A. 2005;102:9371-6.
[13] Neuroscience. 2006;143:387-93.
[14] Nat Protoc. 2011;6:1183-91.
[15] Diabetologia. 2016;59:1732-42.
[16] Eur J Neurosci. 2004;20:240-8.
[17] Front Behav Neurosci. 2014;8:27.
[18] Nat Methods. 2012;9:676-82.
[19] J Endocrinol. 2013;217:175-84.
[20] Nat Rev Neurosci. 2009;10:397-409.
[21] Endocrinol Metab Clin North Am. 2014;43:75-102.
[22] Am J Hypertens. 2001;14:304S-9S.
[23] J Diabetes Res. 2015;2015:341583.
[24] Blood Press Suppl. 1998;3:5-13.
[25] Diabetologia. 2000;43:533-49.
[26] Anat Rec A Discov Mol Cell Evol Biol. 2004;280:854-67.
[27] Neurosci Biobehav Rev. 2000;24:417-63.
[28] Diabetes Care. 2016;39:1635-42.
[29] J Clin Endocrinol Metab. 2011;96:159-67.
[30] Pediatr Diabetes. 2016;17:266-73.
Figures
Figure 1. Effects of SD on the HFD-induced obesity and insulin resistance. (A) The body weight gains of mice. The two short horizontal lines indicate the periods of SD treatment. (B) The fasting levels of glucose and insulin and the HOMA-IR index of mice. (C) The plasma glucose levels during the intra-peritoneal glucose tolerance test (IPGTT) and intra-peritoneal insulin tolerance test (IPITT). The symbols labeled for the statistical significances show the results of two-way ANOVAs followed by Sidak’s multiple comparisons in panels (A) and (C), and the results of one-way ANOVAs followed by Tukey’s post-hoc analysis in panel (B). *p < 0.05, **p < 0.01,
***p < 0.001, ****p < 0.0001 versus Chow group. #p < 0.05, ##p < 0.01 versus HFD group. Body weight gains, n = 6-10; Fasting levels of glucose/insulin and HOMA-IR index, n = 6; IPGTT, n = 3-7; IPITT, n = 6-8.
Figure 2. Effects of bilateral lesions of central amygdalae on the SD-induced aggravation of the HFD-induced insulin resistance. (A) Representative Nissl staining micrographs show the lesion of central amygdala. The dash lines show the margins of the basolateral amygdala. The asterisk indicates the lesion site located in the central amygdala. (B) The body weight gains of mice. (C) The fasting levels of glucose and insulin and the HOMA-IR index of mice. (D) The plasma glucose levels during the intra-peritoneal glucose tolerance test (IPGTT) and intra-peritoneal insulin tolerance test (IPITT). The symbols labeled for the statistical significances show results of one-way ANOVAs followed by Tukey’s post-hoc analysis in panel (C) and results of two-way ANOVAs followed by Sidak’s multiple comparisons in panel (D).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 versus aCSF-HFD group. ##p <
0.01, ###p < 0.001, #### p < 0.0001 versus aCSF-HFD+ESD group. Body weight gains, n = 15; Fasting levels of glucose/insulin and HOMA-IR index, n = 6; IPGTT, n
= 6; IPITT, n = 6.
Figure 3. Effects of HFD and ESD on the expression of neuroplasticity-related proteins in the amygdala of mice. *p < 0.05, **p < 0.01 versus Chow group; # p <
0.05 versus HFD group, one-way ANOVAs followed by Tukey’s post-hoc analysis. n
= 5.
Figure 4. Effects of HFD, SD and amygdala lesions on the circulating levels of corticosterone in mice. **p < 0.01, ***p < 0.001 versus Chow group, one-way ANOVAs followed by Tukey’s post-hoc analysis. n = 6-8.
1
科技部補助專題研究計畫出席國際學術會議心得報告
日期: 106 年 8 月 2 日
一、參加會議經過
今年阿茲海默症 2017 年度國際研討會議選在英國倫敦盛大舉辦。此一研討會是全世界有關阿茲 海默症相關研究最大的會議。過去每兩年舉辦一次,但是由於阿茲海默症的發病率隨人口老化而不斷 攀升,對家庭、社會的衝擊愈趨嚴峻,相關的研究在過去幾年蓬勃發展,於是於 2010 年改為每年一 舉辦一次。
我於 7 月 15 日下午,由高雄機場出發,搭乘港龍航空班機,在香港轉機,於該日晚間抵達倫敦 希斯洛(Heathrow)機場,隨後搭火車、轉地鐵下塌旅館。倫敦是英國的首都所在地,7 月中的氣溫約 在 18-23 度間,氣候涼爽,是一年中最好的季節。倫敦市民相當友善、多元,街景乾淨,是一個通滿 活力的城市。
這次的會議的會場設在倫敦市區的 ExCel Convention Center 內舉辦 (圖 1)。與會學者的研究領域涵蓋範圍相當廣 泛,今年約有 5 千位來自世界各地基礎、醫藥研究人員參 加此會議,同時有廠商在會場展示各類新藥、新式儀器、
或是協會的推廣。
圖 1、ExCel Convention Center 一角。
計畫編號
MOST 105-2321-B-006-019計畫名稱
情感型疾病與代謝症候群交互作用之整合性研究-長期壓力對情緒和代謝中樞神經 可塑性的影響(2/2)出國人員
姓名 郭余民 服務機構
及職稱 國立成功大學 解剖學科(所)教授 會議時間
106 年 7 月 16 日至106 年 7 月 20 日
會議地點 倫敦,英國
會議名稱
(中文)阿茲海默症 2017 年度會議
(英文) Alzheimer’s Association International Conference 2017
發表題目
(中文) 高血壓在小鼠和豬誘發阿茲海默病理
(英文)
Hypertension induces Alzheimer’s disease-related pathologies in
mice and pigs
2
大會分 5 天進行,內容非常豐富,每天上午均安排有 symposia、featured research sessions (圖 2)、plenary session (圖 3)、oral sessions,下午則有 emerging concepts 和兩時段 oral sessions。此外每日有 poster presentations。
圖 2 (右)、Featured research sessions。
圖 3 (下)、Plenary sessions。
我的海報報告在 7 月 27 日,主題是研究高血壓和阿茲海默氏症病理的關聯。我的海報吸引一些臨 床醫師與基礎科學家的興趣,尤其是高血壓豬的部分,討論熱烈(圖 4)。
圖 4、我與海報和有興趣的學者。
大會於 7 月 20 日閉幕。會後我在英國多逗留幾天,進行數日的私人旅遊與參訪,於 7 月 28 日搭 乘國泰航空班機離開倫敦。原定經香港於 29 日返抵高雄機場,但是不巧遇到泥莎颱風,於是改往桃 園機場,再搭高鐵回到台南。
3
二、與會心得
今年大會將主題定調在生活型態、降低風險、提升診斷和早期診斷(Focus on Lifestyle, Risk Reduction, Improved Diagnosis and Early Detection)。個人認為這是非常正確的方向。此外,這次 明顯發現神經科學,尤其是阿茲海默氏症的研究已經進入另一個階段,需要很大的研究團隊,在巨額 經費支持下,跨領域、長期合作追蹤病人,才能獲得較全面的結果。台灣目前相關的研究,明顯落後 國際團隊,而且越來越遠。
三、建議
1. 由於阿茲海默氏症腦部病理造影需要正子掃描,價錢昂貴,因此科技部可考慮以專案方式集中補
助 1~2 個團隊,長期耕耘進行台灣阿茲海默氏症的資料收集和相關研究,不要急著短期驗收成 果。
2. 參與國際會議是與國際學者面對面交流的絕佳地點,能快速有效的獲得當前國際的研究進展與方
向,希望科技部能持續補助國內學者出席國際會議。
四、攜回資料名稱及內容 1. 會議摘要
2. 國外學者名片 五、其他
發表論文摘要
Hypertension induces Alzheimer’s disease-related pathologies in mice and pigs Yao-Hsiang Shih1, Yu-Min Kuo1,2*
1Institute of Basic Medical Sciences, National Cheng Kung University, Tainan, Taiwan 2Department of Cell Biology and Anatomy, National Cheng Kung University, Tainan, Taiwan
Background: Middle-life hypertension (HTN) increases the risk of late-life Alzheimer’s disease (AD).
However, whether HTN causes AD or is a distinct disease that increases the prevalence with age
(co-morbidity) remains unclear. This study aims to test if there is a causal relationship between HTN and AD.
Methods: Two animal models were used to test the role of HTN in AD pathogenesis. First, 3xTg AD model mice at the ages of 2, 5 and 7 months were subjected to “two-kidney-one-clip” (2K1C) surgery to induce HTN. One and three months after the surgery, the blood pressures, hippocampus-dependent learning and memory, and AD-related pathologies, including Aβ production and deposition and Tau phosphorylation, were evaluated. Second, Lanyu-miniature-pigs, at the age of 7 months, were subjected to abdominal aortic constriction (AAC) to induce HTN. The blood pressures and AD-related pathologies were determined 1, 2 and 3 months after the surgery.
Results: The 3xTg mouse study: In all three ages of mice, the blood pressures were increased 7 days after the 2K1C surgery and last at least 1 month. One month after the surgery, the hippocampus-dependent memories were impaired. The levels of Aβ, amyloid precursor protein, pS412-Tau in the ventral
4
hippocampi were elevated, while the levels of Tau protein kinase, GSK3β, were not affected. Furthermore, 7-month-old mice subjected to 2K1C for three months showed higher amyloid plaque loads in the
hippocampi than the Sham controls. The pig study: Three months after the AAC surgery, the blood pressures of pigs were increased. The levels of Aβ, amyloid precursor protein, pT212-Tau, pS412-Tau, activated GSK3β and RAGE were increased, while the levels of LRP1 and pAKT were unchanged in the ventral hippocampi. The activities of another GSK3β upstream inhibition kinase, PKC, were decreased by AAC surgery. Immunohistochemistry revealed that the expression levels of pS412-Tau in the mossy fiber of hippocampi were increased.
Conclusion: HTN aggravates the AD-related pathologies, including impairment of
hippocampus-dependent learning and memory, accumulation and deposition of Aβ and increases of Tau phosphorylation. Control of blood pressure during middle-life may delay the onset of AD.
This study was supported by the Taiwan Ministry of Science and Technology. (MOST 104-2321-B-006-030 and 104-2320-B-006-039)
105年度專題研究計畫成果彙整表
計畫主持人:郭余民 計畫編號:105-2321-B-006-019- 計畫名稱:長期壓力對情緒和代謝中樞神經可塑性的影響(2/2)
成果項目 量化 單位
質化
(說明:各成果項目請附佐證資料或細 項說明,如期刊名稱、年份、卷期、起 訖頁數、證號...等)
國 內
學術性論文
期刊論文 0
研討會論文 1 篇
Molecular mechanisms underlying the interaction between metabolic and mood disorders, 2017/09/23, 中華民 國基礎神經科學學會2017年學術年會暨 會員大會, 台北, 台灣
專書 0 本
專書論文 0 章
技術報告 0 篇
其他 0 篇
智慧財產權 及成果
專利權 發明專利 申請中 0
件
已獲得 0
新型/設計專利 0
商標權 0
營業秘密 0
積體電路電路布局權 0
著作權 0
品種權 0
其他 0
技術移轉 件數 0 件
收入 0 千元
國
外 學術性論文
期刊論文 1
篇
High-fat diet reduces the hippocampal content level of lactate which is correlated with the expression of glial glutamate transporters, Neuroscience Letters.
2017 Oct 16;662:142-146.
研討會論文 2
1. Hippocampal glial glutamate transporters link metabolic disorders to depression: using riluzole as a therapeutic reagent.
2017/09/08-11, 47th European Brain
& Behaviour Society Meeting, Bilbao, Spain
2. Riluzole rescues the high-fat diet-induced depression in mice:
glutamate transporters. 2017/09/11- 15, 53rd The European Association for the Study of Diabetes Annual Meeting 2017, Lisbon, Portugal.
專書 0 本
專書論文 0 章
技術報告 0 篇
其他 0 篇
智慧財產權 及成果
專利權 發明專利 申請中 0
件
已獲得 0
新型/設計專利 0
商標權 0
營業秘密 0
積體電路電路布局權 0
著作權 0
品種權 0
其他 0
技術移轉 件數 0 件
收入 0 千元
參 與 計 畫 人 力
本國籍
大專生 0
人次
碩士生 1 黃婉寧
博士生 0
博士後研究員 0
專任助理 1 陳韻文
非本國籍
大專生 0
碩士生 0
博士生 0
博士後研究員 0
專任助理 0
其他成果
(無法以量化表達之成果如辦理學術活動
、獲得獎項、重要國際合作、研究成果國 際影響力及其他協助產業技術發展之具體 效益事項等,請以文字敘述填列。)