國立臺灣大學生命科學院生態學與演化生物學研究所 碩士論文
Institute of Ecology and Evolutionary Biology College of Life Science
National Taiwan University Master Thesis
Odysseus基因對果蠅精子生成的影響
Characterization of Odysseus in Drosophila Spermatogenesis
吳家翔
Chia-Hsiang Wu
指導教授:丁照棣aa
中華民國102年1月 January, 2013
國 立 臺 灣 大 學 生 態 學 與 演 化 生 物 學 研 究 所
碩 士 論 文
O d ys seu s
基 因 對 果 蠅 精 子 生 成 的 影 響
吳 家 翔 撰
102 1
Advisor: Chau-Ti Ting
i
ii
Acknowledgements
I would like to thank my advisor, Chau-Ti Ting, for all her guidance and immense patience. I am grateful for every opportunity she gave me to learn in her lab. Her
infectious enthusiasm for science and truth always makes me wonder what it is like to be like that. I want to thank all the other members in Ting lab, especially Hsin-Yen Liu and Wen-Chiao Chan. They are always ready to help in every way and I do not think I could ever make it without them. My thanks also goes to the former members of Ting lab, Ya-Jen Cheng, Shi-Yow Yang, Chi-Chun Chen, Cheng-Lin Li, and Kevin Wei. I enjoyed and appreciated their company when I was too clumsy to know what to do.
I would like to thank Shu Fang and all the other members of Fang Lab at Academia Sinica. Without their help I would not be able to access facilities there (the Darkroom, especially). I also thank my committee members, Yu-Chen Tsai at Tunghai University and Haiwei Pi at Chang Gung University, for input and guidance to my thesis work. I thank the Core Facilities at ICOB, Academia Sinica, the DNA Sequencing Service at IBMS, Academia Sinica and the Technology Commons at National Taiwan University for technical support. I am grateful to Chuang-Yu Lin at ICOB for his help and advice about imaging, which is an important part of my work and I cannot thank him enough.
I thank Chung-I Wu for allowing me to visit his lab in spring and summer of 2011.
I thank Mao-Lien Wu for her help at the bench and in daily life. My special thanks goes to Huyen Vu Thanh and her lovely family, Kimmy and Eric, for their hospitality and kindness. My roommates at 6128 S Dorchester Ave. were wonderful people and I owe them so much. My friends Yian and Artur should be awarded the Key to the City for humanitarian work in Chicago. They are heroes who save the Windy City.
iii
I thank my college advisor, Shaw-Yhi Hwang, for all the cozy talks and nice coffee. I thank my friends for watching me and backing me up all the way: Jeff Lin, Tsung-Hsi Lin, Tzu-Wei Chan, Griffin Chang, Shi-Jun Chien, Wen-Che Tsai, Christy Lee, Eileen Lin, Flore Sun, Gennie Lin, and Tricia Lee. I am lucky and honored to meet long-time friends, Tien-Yin Huang, Cathy Hsu, and Wei-Che Hsu, in my early days when the friendship got started without self-interest. They are my most valuable assets.
Last but not least, I want to thank my family for their endless love and tolerance.
Difficult kids who want to be a rebel at age 30 can be reeeeeally troublesome. For this I owe my life to them.
iv
摘要
在物種形成的過程中,雜交不合(包括雜交不活與雜交不孕)能夠形成生殖 屏障。因此,了解造成雜交不合的基因,將有助於認識物種的起源。OdsH 是由 unc-4 基因複製而產生的一個基因。在秀麗隱桿線蟲中,unc-4 與神經發育有關。OdsH 是造成兩近緣種果蠅——擬黃果蠅與模里西斯果蠅——雜交子代雄性不孕的主要
因子,據信可能是因 OdsHmau在精巢中異常表現、或因 OdsHmau對擬黃果蠅的 Y
染色體異常結合所導致。在黃果蠅中,剔除 OdsH 使雄蟲生殖力下降;免疫染色實 驗顯示,缺少 OdsH 表現的精巢,其生殖細胞數目較少。在本研究中,我測試了六 株 OdsH RNAi 果蠅,其中四株來自果蠅品系中心,兩株為自行建立。我並使用了 在特定位置表現綠色螢光蛋白的果蠅,標記精巢內部不同細胞,觀察 OdsH 對黃果 蠅精子生成的影響。本研究展示六株 OdsH RNAi 果蠅的效果,有助於未來研究 OdsH 與 unc-4 的功能重疊。對黃果蠅精巢的觀察結果,則暗示 OdsH 可能參與精 子生成當中的減數分裂與後期分化。
關鍵字:果蠅、基因重複、雜交不合、OdsH 基因、RNA 干擾、精子生成、unc-4 基因
v
Abstract
Hybrid incompatibility, including hybrid inviability and hybrid sterility, can act as a reproductive barrier in the process of speciation. Therefore, genes involved in hybrid incompatibility may provide a window on how new biological species form. The gene Odysseus-site homeobox (OdsH) is duplicated from unc-4, a homolog of a neuron developmental gene in Caenorhabditis elegans. OdsH has been identified as a crucial element that triggers hybrid male sterility in reciprocal crosses between two sibling species, Drosophila simulans and D. mauritiana. Previous studies have suggested that misexpression of OdsHmau in testes, or additional binding of OdsHmau to the Y
chromosome of D. simulans, results in the hybrid sterility. In D. melanogaster, targeted gene knockout revealed that lack of OdsH expression reduces male fertility.
Immunostaining in OdsH null mutant (OdsH0) testes has shown a decrease in the number of germ cells. In this thesis, I generated two OdsH RNAi constructs and examined their efficiency with the other four available at stock centers. In addition, I used reporter lines that express enhanced green fluorescent protein to further
characterize this testis-specific gene at the cellular level. My results demonstrated the efficiency of these RNAi strains, which should facilitate future studies on the functional redundancy between OdsH and unc-4. Cytological observation implies that OdsH may play a role in meiosis and terminal differentiation during spermatogenesis.
Keywords: Drosophila, gene duplication, hybrid sterility, OdsH, RNA interference, spermatogenesis, unc-4
vi
Contents
口試委員會審定書 ... i
Acknowledgements ... ii
摘要 ... iv
Abstract ... v
Contents ... vi
List of Figures ... viii
List of Tables ... x
Introduction ... 1
Materials and Methods ... 7
Fly strains and husbandry ... 7
Synthesis of mir-6-1-based microRNA for RNAi constructs ... 7
Generation of RNAi strains against OdsHsim and OdsHmau in D. melanogaster ... 12
Immunostaining ... 12
Imaging and quantification ... 13
Statistical methods ... 13
RNA extraction and cDNA reverse transcription ... 13
Results ... 18
Generation of RNAi strains against OdsHsim and OdsHmau in Drosophila melanogaster ... 18
vii
RNAi knockdown rescues the OdsHmel ectopic expression phenotype in Drosophila
eye ... 18
His-GFP expression in testes of OdsH+ and OdsH0 flies ... 25
Sa-GFP expression in testes of OdsH+ and OdsH0 flies ... 26
BamP-GFP expression in testes of OdsH+ and OdsH0 flies ... 26
Discussion ... 32
OdsH RNAi strains should facilitate future studies on the functional divergence of duplicated genes and the normal function of OdsHsim ... 32
His-GFP expression implies GSC loss or disruption of subsequent spermatogenic development in young OdsH0 flies ... 35
Sa-GFP expression hints a possible role of OdsH in meiosis or spermatid differentiation ... 38
BamP-GFP expression yields no information on OdsH in early spermatogenesis .... 40
References ... 42
Appendix I: RNAi constructs for transgenesis in Drosophila simulans ... 45
Generation of pCaSpeR4-based RNAi vectors for P-element transformation ... 45
Generation of attB-P[acman]-based RNAi vectors for P-element transformation by recombineering... 45
Appendix II: mir-6-1-based RNAi design ... 51
viii
List of Figures
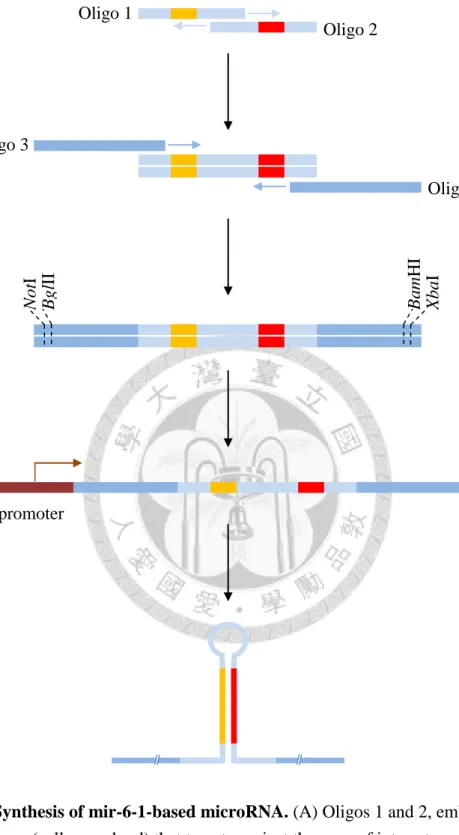
Figure 1. Synthesis of mir-6-1-based microRNA ... 10
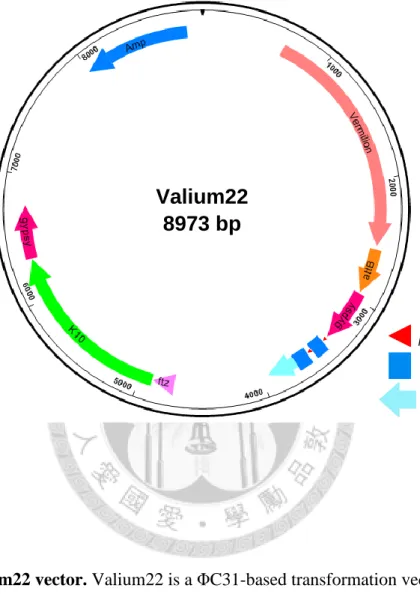
Figure 2. Valium22 vector ... 15
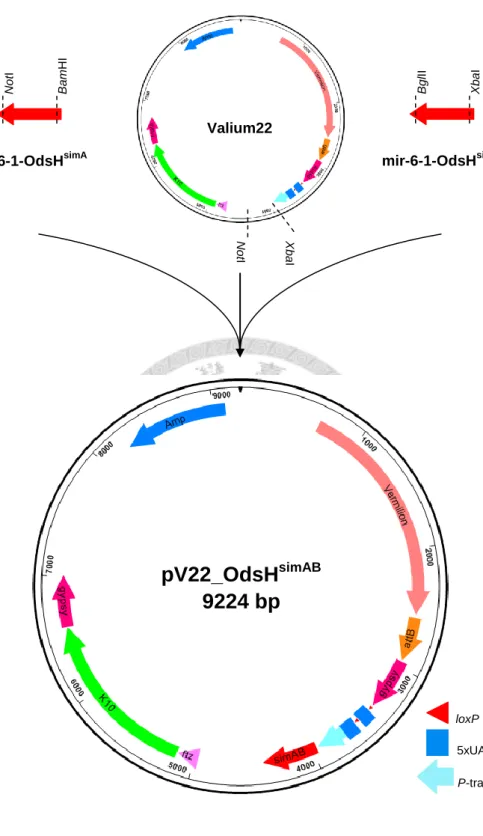
Figure 3. Generation of pV22_OdsHsimAB ... 16
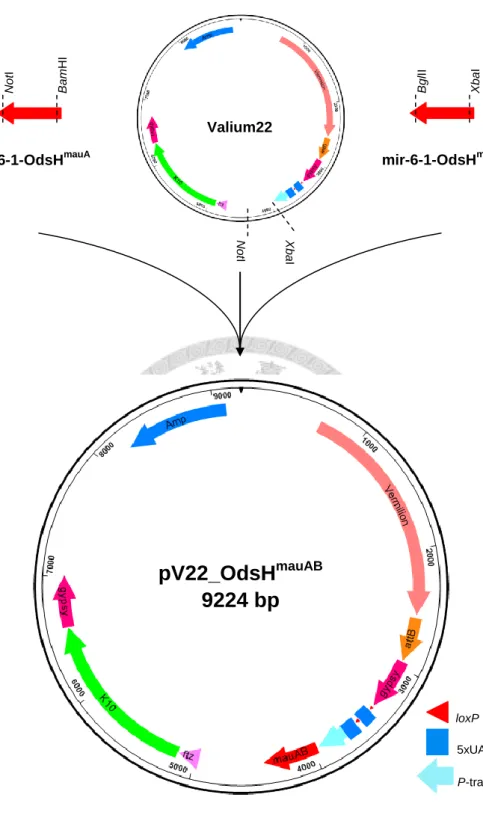
Figure 4. Generation of pV22_OdsHmauAB ... 17
Figure 5. Drosophila eye images from wild-type and transgenic flies ... 21
Figure 6. Normalized values of eye area to w1118 control ... 22
Figure 7. Normalized values of eye circumference to w1118 control ... 23
Figure 8. RT-PCR detection of OdsHmel expression in wild-type and transgenic male flies ... 24
Figure 9. His-GFP expression in testes of 0- and 32-day-old OdsH+ and OdsH0 flies ... 27
Figure 10. The average volume of His-GFP signal per testis ... 28
Figure 11. Sa-GFP expression in testes of 1-, 15-, and 55-day-old OdsH+ and OdsH0 flies ... 29
Figure 12. The average volume of Sa-GFP signals in spermatocytes ... 30
Figure 13. BamP-GFP expression in testes of 1-, 10-, and 32-day-old OdsH+ and OdsH0 flies ... 31
Figure 14. OdsH structure and the target regions of UAS-OdsHRNAi strains used in this thesis... 33
Figure 15. Male germline differentiation in Drosophila melanogaster ... 37
ix
Figure 16. tMAC (aly-class genes) and tTAFs (can-class genes) network in Drosophila melanogaster spermatocytes ... 41 Figure 17. Design of pCaSpeR4_V22_OdsHsimAB, a pCaSpeR4-based RNAi vectors for
P-element transformation ... 48 Figure 18. Retrival of miRNA construct against OdsHsim from pV22_OdsHsimAB into
attB-P[acman]-CmR by recombineering ... 50
x
List of Tables
Table 1. Fly strains used in this thesis ... 9 Table 2. Oligo names and sequences used in this thesis ... 11 Table 3. Summary of the introduction of mir-6-1-based RNAi constructs into
Drosophila melanogaster via C31 integrase-mediated recombination ... 20
Table 4. The target sequences of UAS-OdsHRNAi strains obtained from stock centers .... 34 Table 5. Homology of the target sequences of mir-6-1-based RNAi constructs in
Drosophila melanogaster, D. simulans and D. mauritiana ... 34 Table 6. Oligo sequences for mir-6-1-based RNAi synthesis ... 52 Table 7. Synthesized miRNA sequences ... 53
1
Introduction
People have long been fascinated by the origin of species. From Aristotle (384 BC-322 BC) to Carl Linnaeus (1707-1778), philosophers strived to classify the creatures on Earth and assigned the order in which all lives are created. Conceivably, one of the earliest definitions of species pertains to the morphological similarity among individuals, because there is no more instinctive way to categorize lives than the observable and measureable traits by the naked eye. In face of vast numbers of species, people held different view on their origin. In the view of creationism, species today are just the same as they were when the Earth was formed. In contrast, some people doubted and
questioned the possibility of creating new species. In the pre-Darwinian era, challenges against the deep-rooted ecclesiastical perspective begun emerging already. Such as Linnaeus, based on his classification work, pointed that a new species could arise from hybridizing plants. Jean-Baptiste Lamarck (1744-1829) also advanced the idea of hybridization as a source of new species (Reviewed in MCCARTHY 2008). By the time Charles Darwin (1809-1882) was born, the Age of Enlightenment came to an end. It was a society where reason was upheaved to the level comparable to church traditions, a society where research fellows received supports from state-funded academies, and a society where salons, coffee houses and debate clubs permeated.
Several keys contributed to Darwin’s achievement in the theory of evolution: his immense curiosity and interest in natural history, the upper-class background and financially secure life that enabled him to be a gentleman scientist on HMS Beagle (1831-1836) (BROWNE 2009), and befriending his contemporary scientists, like Asa Gray (1810-1888), John Gould (1804-1881), John Herschel (1792-1871), Joseph Dalton Hooker (1817-1911) and Charles Lyell (1797-1875). Among them, Lyell’s
2
uniformitarian view of geology—the same laws and processes of nature have always operated in the past and at present on Earth in the same way— helped shape Darwin’s speculations on the “mystery of mysteries”(DARWIN 1859). Finally, a manuscript on natural selection from Alfred Russel Wallace (1823-1913) in 1858 gave impetus to the publication of Darwin’s painstaking work and the cornerstone of evolutionary biology as well in 1859. In On the Origin of Species, Darwin proposed natural selection as the fundamental mechanism of evolution. It occurs when descents, from a common ancestor, carry heritable traits with variation related to fitness. Individuals with certain traits may have better chance to survive and reproduce than the others without. This can
consequently lead to a specialized population, or a new species, that adapts to a particular way of life (DARWIN 1859).
Upon the arrival of Darwin’s theory, the community was not ready for this radical idea. One reason that it came under suspicion was the absence of evidence to support.
Although Gregor Mendel (1822-1884) published his seminal work on modern genetics in 1865, Darwin was not aware of it and failed to explicate his crossing experiment results with the blending inheritance and his pangenesis theory (Reviewed in
CHARLESWORTH and CHARLESWORTH 2009). It was not until the turn of the 20th century that Mendel’s laws were rediscovered and advocated by Hugo de Vries (1848-1935), Carl Correns (1864-1933), Erich von Tschermak (1871-1962) and William Bateson (1861-1926) (Reviewed in KEYNES and COX 2008). Yet the evolution and species problems were not all solved happily ever after. For example, Bateson, an English biologist who coined the term “genetics,” did not lend support to Darwin’s idea. In fact, since the renaissance of Mendelian inheritance, there were dissenting voices from Darwin’s natural selection. These voices, including de Vries and Bateson, believed that
3
a new species arises from mutations that cause large, discontinuous modification of individuals. This theory—referred to as mutationism or Mendelism—was defied by the biometricians, who followed Darwin’s view and concentrated on small, continuous variations among individuals (Reviewed in AYALA and FITCH 1997). About the same time, explosive progress in genetics enabled scientists to trace the inheritance factors in cells. Thomas Hunt Morgan (1866-1945) used Drosophila melanogaster to demonstrate the chromosome theory of inheritance proposed by Walter Sutton (1877-1916) and Theodor Boveri (1862-1915). For the first time, a gene was showed physically on a specific chromosome as a carrier of traits. The Morgan group—including his students Alfred Sturtevant (1891-1970), Calvin Bridges (1889-1938) and Hermann Joseph Muller (1890-1967)—became the trailblazer for the further understanding of modern genetics. Together, they established what is common knowledge for geneticists today, such as genetic linkage, polytene chromosome map and X-ray mutagenesis (Reviewed in DAVIS 2004).
The mutationist-biometrician contradiction was reconciled by the introduction of mathematical statistics into genetics in the 1920s and 1930s. Ronald Fisher (1890-1962), Sewall Wright (1889-1988), and John Burdon Sanderson Haldane (1892-1964) were among the most influential figures that developed the theoretical framework of
population genetics. Their work demonstrated that natural selection could act on small, continuous variations that are passed on from parents to offspring through Mendelian inheritance and contribute to morphological and functional alterations. Furthermore, it led to the demise of mutationism and stimulated the forthcoming synthetic theory of evolution, the Modern Synthesis (Reviewed in AYALA and FITCH 1997; CHARLESWORTH
and CHARLESWORTH 2009).
4
At the dawn of the Modern Synthesis era, two prominent scientists, Theodosius Dobzhansky (1900-1975) and Ernst Mayr (1904-2005), made tremendous contribution in combining the evolution theory and multiple scientific disciplines. Both of them proposed what is known as the biological species concept: species are natural populations that are able to interbreed and reproductively isolated from other such populations. Since then, the discussion about isolating mechanisms has been the core to decode speciation (DOBZHANSKY 1937; MAYR 1942). In 1936, Dobzhansky published his groundbreaking work in the genetics of speciation (DOBZHANSKY 1936). He crossed two species, D. pseudoobscura and D. persimilis, and backcrossed the fertile F1 females to males of the two species. He thus obtained hybrids with many combinations of heterospecific chromosomes. With the help of visible markers, the fertility of these hybrids, which was measured by the testis size, indicated which region on whose chromosome caused hybrid sterility—a reproductive barrier that is required to create a new species. Dobzhansky gave empirical evidence that genes, instead of cytoplasm difference or chromosome rearrangement, cause hybrid sterility. Also, through this experiment, he provided an answer to Darwin’s paradox: how can sterility or inviability survive through natural selection? Dobzhansky proposed an interaction between (at least) two loci responsible for hybrid sterility: Assume two allopatric populations with the same genotype aabb. Then a mutation occurs: now one population comprises Aabb and AAbb, while the other comprises aaBb and aaBB. The new allele A is compatible with b, and B is compatible with a. In this scenario, allele A and B are never put to the test together. When the two populations meet, the incompatibility between A and B can lead the hybrid offspring to a dead end. This idea, also discussed and contributed by William Bateson (1909) and H. J. Muller (1942), has become known as the
5
Bateson-Dobzhansky-Muller Model (Reviewed in ORR 1996).
Dobzhansky’s pioneering experiment did not add zest to the seeking of hybrid incompatibility genes. Conversely, interest in this subject was not revived until early 1980s, when Jerry Coyne launched the study to answer the genetic basis of hybrid male sterility between D. simulans and D. mauritiana (COYNE 1984; JOHNSON 2008). Coyne and Charlesworth (1986) further traced the largest sterility effect to the X chromosome and found it closely linked to the marker, forked. Using molecular markers, Perez et al.
(1993) were able to associate the sterility effect to a major locus and named it Odysseus (Ods). The name “Odysseus”—the hero who devised the wooden horse that destroyed the city of Troy in the Homeric epic—is metaphorical: the D. mauritiana allele invades the D. simulans background and finally leads to a crash of the lineage. A subsequent investigation of Ods revealed that Odsmau alone cannot cause sterility; only when Odsmau and its distal region were cointrogressed, the sterility effect can be fully exerted (PEREZ
and WU 1995). Finally, an attempt of fine-scale genetic mapping successfully delineated the Ods locus to the nucleotide level (TING et al. 1998). The location of Ods was
defined by two introgression lines from a collection of 190 newly generated
recombinants: 99F (the fertile line that has the longest introgressed fragment from D.
mauritiana) and 193S (the sterile line that has the shortest introgressed fragment from D.
mauritiana), which differ only by a 3-kb region where reside the (later known) exons 3 and 4 of a gene. Since this gene contains a homeobox in the exons 2 and 3, it was named Odysseus-site homeobox gene (OdsH).
The discovery of OdsH has posed several questions: First, sequence analysis showed that OdsH, particularly its homeodomain, has undergone rapid evolution, which is asymmetric to its highly conserved paralog, unc-4 (TING et al. 1998). Second, the fact
6
that the male reproductive organ-specific OdsH duplicated from the neuron-specific unc-4 (TABUCHI et al. 1998) supports a hypothesis that a duplicated gene, which is liable to functional divergence and has not reached its stable maintenance, is a candidate
“speciation gene” (TING et al. 2004). Third, since the maladaptive defect induced by OdsH in hybrids is an evolutionary by-product, what is the normal function of OdsH? In D. melanogaster, OdsH is suggested to enhance sperm production, probably by speeding up sperm maturation (SUN et al. 2004) and/or maintaining germline stem cells (CHENG
et al. 2012). Fourth, what is the mechanism underlying the hybrid sterility? The homeodomain motif supports an educated guess that OdsH is a DNA-binding
transcription factor. The misregulated OdsH in D. simulans-D. mauritiana hybrid sterile line disrupts the expression of genes involved in spermatogenesis, which may be the cause of spermatogenic failure (LU et al. 2010; SUN et al. 2004). However, Bayes and Malik reported an additional heterochromatic localization of OdsHmau on the Y
chromosome of D. simulans may cause chromosome decondensation in sterile hybrids, which suggests a “gain-of-function” protein-DNA interaction that echoes the
Bateson-Dobzhansky-Muller Model (BAYES and MALIK 2009).
In this thesis, I tested four OdsH RNAi strains available from stock centers and two that newly generated at my hands. I used reporter lines that express green
fluorescent protein (GFP) to characterize this testis-specific gene at the cellular level.
These efforts are intended to tackle two questions: (1) The function redundancy between OdsH and its paralog, unc-4, and (2) The specific role of OdsH in D. melanogaster spermatogenesis. Together, this thesis provides some new hints on genes involved in hybrid incompatibility and materials for future study.
7
Materials and Methods
Fly strains and husbandry
All Drosophila strains were raised at 25˚C on standard cornmeal medium.
his-GFP (CLARKSON and SAINT 1999), sa-GFP (CHEN et al. 2005) (a gift from Margaret T. Fuller at Stanford University), and bamP-GFP (CHEN and MCKEARIN 2003) (a gift from Haiwei Pi at Chang Gung University) were crossed to OdsH+ and OdsH0 flies (SUN et al. 2004) to visualize germ cells in different developmental stages of spermatogenesis. UAS-OdsHRNAi strains were obtained from the Vienna Drosophila RNAi Center at the Campus Vienna Biocenter (GD51289 and KK103949) and the Transgenic RNAi Project at the Harvard Medical School (JF02198 and HMS01554).
GMR>OdsHmel was used for testing RNAi efficiency. D. melanogaster y1 sc1 v1
P{nos-phiC31\int.NLS} X; P{CaryP}attP2 was provided by Rainbow Transgenic Flies, Inc. (Camarillo, CA) and used for transgenesis. FM6 was used to make homozygous sa-GFP, OdsH+ and sa-GFP, OdsH0 flies. See Table 1 for strain information.
Synthesis of mir-6-1-based microRNA for RNAi constructs
The design of mir-6-1-based miRNA against OdsH was based on Chen et al.
(2007). A target site on OdsHsim coding region was selected according the criteria as follows: (1) a 22-nucleotide sequence, (2) a 30-52% guanine-cytosine content, (3) at least three adenine-thymine within the 16th to 20th nucleotide, (4) an adenine at the 20th nucleotide, (5) no guanine at the 13th nucleotide, (6) an adenine at the 3rd nucleotide (optional), and (7) an thymine at the 10th nucleotide (optional). The resulting 22-nt sequence was then swapped to the Oligos 1 and 2 cassettes with the 20th nucleotide in the Oligo 1 replaced by cytosine. Then, the last four nucleobases of the Oligo 2 were
8
changed to the reverse-complement counterpart of the last four nucleobases of the modified 22-nt sequence in Oligo 1. Anneal Oligos 1 and 2 by the first-round PCR using Expand High FidelityPLUS PCR System (Roche), which generated the mir-6-1 mimic sequence that later formed the miRNA precursors (Figure 1, A). This product was amplified by the second-round PCR with Oligos 3 and 4, which added the mir-6-1 flanking sequences and restriction recognition sites NotI/BglII and BamHI/XbaI at 5-prime and 3-prime ends respectively (Figure 1, B and C). The resulting
PCR-amplified fragment was confirmed by sequencing after cloned into pCR™4-TOPO® TA vector (Invitrogen) and the construct was named mir-6-1-OdsHsimA. By the same method the constructs mir-6-1-OdsHsimB, mir-6-1-OdsHmauA and mir-6-1-OdsHmauB were generated. See Table 2 for oligo sequences.
9
Table 1. Fly strains used in this thesis.
Strain Genotype Note
w1118 w1118
FM6 FM6, y1 dm+ w1/Dp(1;Y)y+ Bloomington stock 4327
OdsH+ w1118 OdsH+ SUN et al. (2004)
OdsH0 w1118 OdsH0 SUN et al. (2004)
GMR>OdsHmel w*; P{GAL4-ninaE.GMR}12; P{w+ UAS-OdsHmel 9B} LIN (2009) his-GFP w1118; P{His2AvT:Avic\GFP-S65T
}62A CLARKSON and SAINT (1999); Bloomington stock 5941
sa-GFP y w P{w+ sa-GFP} CHEN et al. (2005)
bamP-GFP hsFLP; Sp/Cyo; P{w+ -898/+133-bam:GFP} CHEN and MCKEARIN (2003)
GD51289 w1118; P{w+ GD51289} UAS-OdsHRNAi from VDRC
KK103949 w1118; P{w+ KK103949} UAS-OdsHRNAi from VDRC
JF02198 y1 v1; P{TRiP.JF02198}attP2 UAS-OdsHRNAi from TRiP HMS01554 y1 sc* v1; P{TRiP.HMS01554}attP2 UAS-OdsHRNAi from TRiP
attP2 y1 sc1 v1 P{nos-phiC31\int.NLS}X; P{CaryP}attP2 NI et al. (2008); Bloomington stock 25710
10
NotI BglII BamHI XbaI
(A)
(B)
(C)
(D)
Figure 1. Synthesis of mir-6-1-based microRNA. (A) Oligos 1 and 2, embedded a 22-nt sequence (yellow and red) that targets against the gene of interest, are annealed by the first-round PCR. (B) Oligos 3 and 4 are used to amplify the first-round PCR product, creating (C) a ~230-bp fragment flanked by NotI/BglII and BamHI/XbaI sites at 5-prime and 3-prime end, respectively. (D) As the miRNA template placed under the control of a promoter is transcribed, the hairpin-structured primary miRNA forms. This artificial primary miRNA then enters the endogenous RNAi pathway and acts as a functional miRNA.
promoter
Oligo 1
Oligo 2
Oligo 3
Oligo 4
11
Table 2. Oligo names and sequences used in this thesis.
Name Sequence (5′ to 3′)
“Oligo 1” for mir-6-1-based RNAi synthesis
miR6_OdsHsim_A1 GGCAGCTTACTTAAACTTAATCACAGCCTTTAATGTATAGCGGTTTGGTGTCAAACTATAAGTTAATATACCATATC miR6_OdsHsim_B1 GGCAGCTTACTTAAACTTAATCACAGCCTTTAATGTAAATCGTCCTTCAGCATCACATTAAGTTAATATACCATATC miR6_OdsHmau_A1 GGCAGCTTACTTAAACTTAATCACAGCCTTTAATGTGGAGAAAGCCTTCCAGGAACATTAAGTTAATATACCATATC miR6_OdsHmau_B1 GGCAGCTTACTTAAACTTAATCACAGCCTTTAATGTTCGAGGATATGGAAGTGGACGTTAAGTTAATATACCATATC
“Oligo 2” for mir-6-1-based RNAi synthesis
miR6_OdsHsim_A2 AATAATGATGTTAGGCACTTTAGGTACATAGCGGTTTGGTGTCAAAATATAGATATGGTATATTAACTTATAGT miR6_OdsHsim_B2 AATAATGATGTTAGGCACTTTAGGTACAAATCGTCCTTCAGCATCAAATTAGATATGGTATATTAACTTAATGT miR6_OdsHmau_A2 AATAATGATGTTAGGCACTTTAGGTACGGAGAAAGCCTTCCAGGAAAATTAGATATGGTATATTAACTTAATGT miR6_OdsHmau_B2 AATAATGATGTTAGGCACTTTAGGTACTCGAGGATATGGAAGTGGAAGTTAGATATGGTATATTAACTTAACGT
“Oligo 3” and “Oligo 4” for mir-6-1-based RNAi synthesis
mi6_5′ NotI_BglII GGCGCGGCCGCCGCCAGATCTTTTAAAGTCCACAACTCATCAAGGAAAATGAAAGTCAAAGTTGGCAGCTTACTTAAACTTA mi6_3′ BamHI_XbaI GGCCTCTAGAACGGATCCAAAACGGCATGGTTATTCGTGTGCCAAAAAAAAAAAAAATTAAATAATGATGTTAGGCAC
“LA_F” and “LA_R” for recombineering
VALIUM_GYPSY_L_F AGGCGCGCCTCGGTACACTAGTTG VALIUM_GYPSY_L_R CGCGGATCCTACTAGTGTTGTTGG
“RA_F” and “RA_R” for recombineering
VALIUM_GYPSY_R_F CGCGGATCCGCGTGCAGTGGCCAC VALIUM_GYPSY_R_R ACCTTAATTAAGAGCTCCTGCAGGTTG
“5′-Check-F” and “5′-Check-R” for recombineering
pacman_L for VGL GTGAGCGCGCGTAATACGACTCAC VALIUM20_22_R TAATCGTGTGTGATGCCTACC for OdsHmel expression check by RT-PCR
OdsH c0+ GCAAAAGCTAAGACGAAAATGGAT
U8 4524- TGCTTAGCTAACCACCCGAAATCA
rp49F CAGTCGGATCGATATGCTAAGCTGT
rp49R TTACCGACCTTGGGCATCAGATACT
12
Generation of RNAi strains against OdsHsim and OdsHmau in D. melanogaster Two mir-6-1-based constructs, together with the vector Valium22 (NI et al. 2011) (Figure 2), were combined by a three-way ligation with appropriate restriction enzyme digestions, which created pV22_OdsHsimAB (Figure 3). pV22_OdsHmauAB was generated by the same method (Figure 4). These two resulting constructs were then microinjected into embryos of attP flies (Bloomington Stock 25710) via ΦC31 integrase-mediated transformation by Rainbow Transgenic Flies, Inc. Hatched virgin adults (P0) were sexed and mated with the parental attP virgin flies, and transformants were screened in F1 flies to create homozygous stocks.
Immunostaining
Immunostaining was performed as described (SINGH and HOU 2008) with minor modifications. Testes were dissected in Drosophila ringer’s solution and fixed in 4%
paraformaldehyde made in phosphate-buffered saline (PBS) for 20 minutes. Fixed samples were washed three times for 20 minutes each with 1 × PBST (0.1% Triton X-100 in 1 × PBS), and blocked in 1 min blocking buffer (Bio-Cando) at 4C overnight.
Samples were then incubated in primary antibody at 4C overnight, washed five times for 30 minutes each with 1 × PBST, and incubated in secondary antibody at room temperature for two hours in the dark. After 5 washes in 1 × PBST and another three washes in 1 × PBS, samples were mounted on slides with a drop of mounting medium (50% glycerol in 1 × PBS) and stored at 4C until imaging.
Primary and secondary antibodies were diluted in the blocking buffer. Primary antibodies used were: 1:50 mouse anti-Fasciclin III (Developmental Studies Hybridoma Bank, DSHB), and 1:10,000 rabbit anti-GFP (Torrey Pines Biolabs). Alexa fluorescence
13
conjugated secondary IgG (H+L) antibodies were used at 1:200 for goat anti-rabbit 488 and 568 (Molecular Probes).
Imaging and quantification
Fluorescence images of testes were obtained using the Leica TCS SP5 AOBS confocal microscope. Signals were analyzed using MetaMorph (Molecular Devices) software. Image stacks were converted into 3D objects by 4D viewer, and signal
intensity was estimated by summing the number of voxels within the regions of interest.
Light microscope images of compound eyes were obtained using Zeiss SteREO Lumar.V12. Compound eye size was measured using MetaMorph software. Scanning electron microscope images of compound eyes were obtained using the FEI Inspect S in low vacuum mode.
Statistical methods
Statistical significance of the differences in average volume of green fluorescent signals per testis and in average compound eye size and circumference of various genotypes was estimated using the Mann-Whitney U test.
RNA extraction and cDNA reverse transcription
Total RNA was extracted from five three-day-old adult male flies with TRIzol reagent (Invitrogen) following the manufacturer’s instructions. cDNA was synthesized using SuperScript® III First-Strand Synthesis System for RT-PCR (Invitrogen)
according to the manufacturer’s protocol with OdsHmel-specific primers. Drosophila ribosomal protein 49 gene (rp49) was used as a loading control. PCR conditions
14
included one cycle of 2 min at 95°C followed by 25 cycles of 95°C for 30 s, 55°C for 90 s and 72°C for 30 s, and a final incubation at 72°C for 10 min. PCR products were separated on 0.8% agarose gels. See Table 2 for primer sequences.
15
Figure 2. Valium22 vector. Valium22 is a ΦC31-based transformation vector and specialized for expression in the female germline. ftz, the 3-prime UTR intron of fushi tarazu, contains the transport and localization signal. K10, the 3-prime UTR of female sterile (1) K10, contains the localization element. The whole design is flanked by two gypsy insulators to stabilize transgenic expression.
loxP 5xUAS
P-transposase promoter
Valium22
8973 bp
16
Figure 3. Generation of pV22_OdsHsimAB. Two synthetic miRNA templates and the vector Valium22 were digested with different restriction enzyme pairs: XbaI/BglII, BamHI/NotI, and XbaI/NotI. These three fragments were then combined in a three-way ligation to create pV22_OdsHsimAB for ΦC31 integrase-mediated transformation.
NotI
mir-6-1-OdsHsimA
BamHI BglII
mir-6-1-OdsHsimB
XbaI
NotI XbaI
Valium22
pV22_OdsH
simAB9224 bp
loxP 5xUAS
P-transposase promoter
17
Figure 4. Generation of pV22_OdsHmauAB. Two synthetic miRNA templates and the vector Valium22 were digested with different restriction enzyme pairs: XbaI/BglII, BamHI/NotI, and XbaI/NotI. These three fragments were then combined in a three-way ligation to create pV22_OdsHmauAB for ΦC31 integrase-mediated transformation.
NotI
mir-6-1-OdsHmauA
BamHI BglII
mir-6-1-OdsHmauB
XbaI
NotI XbaI
Valium22
pV22_OdsH
mauAB9224 bp
loxP 5xUAS
P-transposase promoter
18
Results
Generation of RNAi strains against OdsHsim and OdsHmau in Drosophila melanogaster
The mir-6-1-based RNAi constructs, pV22_OdsHsimAB and pV22_OdsHmauAB, were introduced into D. melanogaster y1 sc1 v1 P{nos-phiC31\int.NLS}X;
P{CaryP}attP2 embryos. For the pV22_OdsHsimAB construct, 63.1% of hatched larvae eclosed, 82.9% of the adults were fertile, and 26.5% of the fertile adults were
transformants. For the pV22_OdsHmauAB construct, 60% of hatched larvae eclosed, 70.8% of the adults were fertile, and 29.4% of the fertile adults were transformants (Table 3). Two UAS-OdsHRNAi strains were therefore generated and named
OdsHRNAi_simAB and OdsHRNAi_mauAB.
RNAi knockdown rescues the OdsHmel ectopic expression phenotype in Drosophila eye
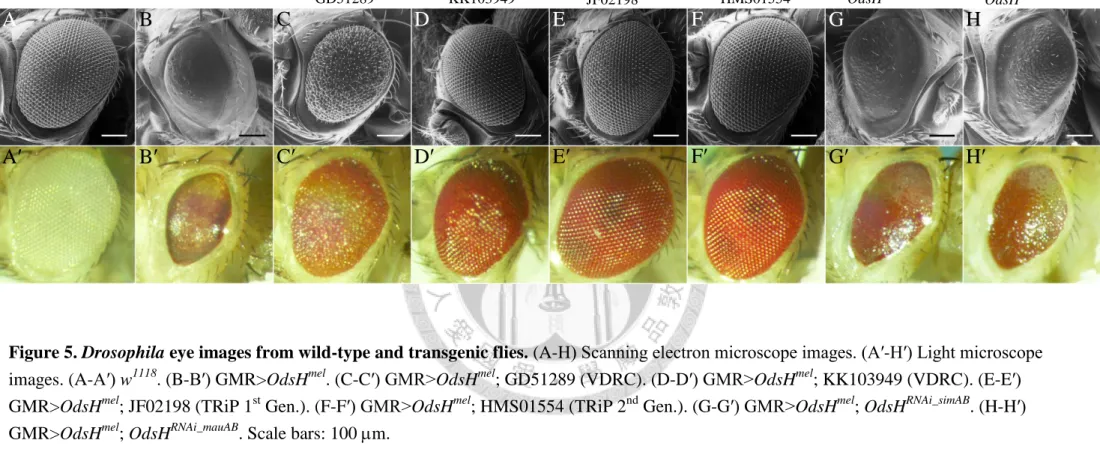
A total of six UAS-OdsHRNAi strains, including two newly generated and four obtained form stock centers, were investigated in the GMR-Gal4>UAS-OdsHmel background. Observations on Drosophila eye phenotypic changes demonstrate that all the six RNAi strains yield the knockdown effect. Compared to the wild-type eyes (Figure 5, A-A′), GMR-driven OdsHmel caused severe eye defects: reduction in size, and loss of ommatidia and bristles (Figure 5, B-B′). The two VDRC and two TRiP lines restored the eye defects evidently (Figure 5, C-F′). All of them displayed a
wild-type-like eye of normal size. However, only three of them showed regular
ommatidia and bristles (Figure 5, D-F′), except for the strain GD51289, which produced fused ommatidia and disorganized bristles (Figure 5, C-C′). The newly synthesized
19
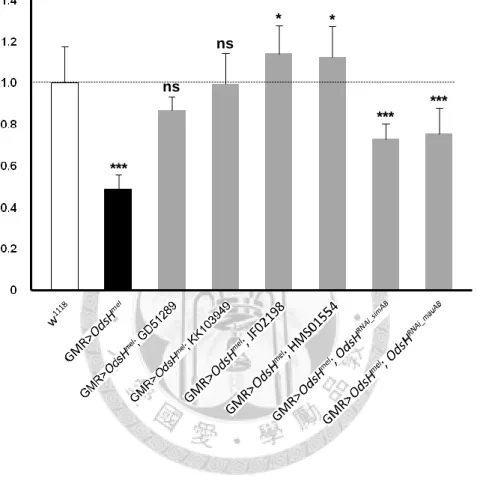
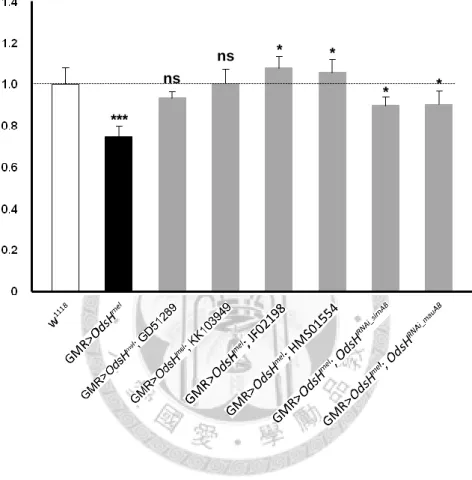
OdsHRNAi_simAB and OdsHRNAi_mauAB, whose guide and passenger strands are based on the coding sequences of OdsHsim and OdsHmau, showed minor but palpable knockdown effect. In the GMR-Gal4>UAS-OdsHmel background, OdsHRNAi_simAB and OdsHRNAi_mauAB partially restored not only the size of compound eyes but also the growth of ommatidia and bristles (Figure 5, G-G′ and H-H′). Measurements of eye area and circumference provide quantification results in strong agreement with phenotypic observations that the four strains from stock centers had higher efficiency than the two self-made (Figures 6 and 7).
To further confirm the rescue effect, I used RT-PCR to detect the OdsHmel mRNA level of the above-mentioned strains. The result showed that the VDRC and TRiP stocks obviously reduced OdsHmel expression towards the wild-type (w1118 and GMR-Gal4) level. It is expected that the decrease of OdsHmel mRNA was ambiguous in
OdsHRNAi_simAB and OdsHRNAi_mauAB, whose RNAi constructs were designed according to OdsHsim and OdsHmau coding sequences, because OdsH has experienced rapid evolution and diverged at the sequence level (Figure 8).
20
Table 3. Summary of the introduction of mir-6-1-based RNAi constructs into Drosophila melanogaster via C31 integrase-mediated recombination. *Only a part of the hatched larvae were delivered by Rainbow Transgenic Flies, Inc.
Symbol Genotype Hatching rate
(larvae/injected eggs)
Eclosing rate (P0 adults/larvae)
Fertility rate (fertile P0 adults/P0 adults)
Transformation rate (transformants/fertile P0 adults)
OdsHRNAi_simAB y1 sc1 v1; P{OdsHRNAi_simAB}attP2 * 63.1% (41/65) 82.9% (34/41) 26.5% (9/34)
OdsHRNAi_mauAB y1 sc1 v1; P{OdsHRNAi_mauAB}attP2 * 60% (24/40) 70.8% (17/24) 29.4% (5/17)
21
Figure 5. Drosophila eye images from wild-type and transgenic flies. (A-H) Scanning electron microscope images. (A′-H′) Light microscope images. (A-A′) w1118. (B-B′) GMR>OdsHmel. (C-C′) GMR>OdsHmel; GD51289 (VDRC). (D-D′) GMR>OdsHmel; KK103949 (VDRC). (E-E′) GMR>OdsHmel; JF02198 (TRiP 1st Gen.). (F-F′) GMR>OdsHmel; HMS01554 (TRiP 2nd Gen.). (G-G′) GMR>OdsHmel; OdsHRNAi_simAB. (H-H′) GMR>OdsHmel; OdsHRNAi_mauAB. Scale bars: 100 m.
A B C D E F G H
A′ B′ C′ D′ E′ F′ G′ H′
GMR>OdsHmel GMR>OdsHmel
w1118
GD51289 KK103949 JF02198 HMS01554 OdsHRNAi_simAB OdsHRNAi_mauAB
22
Figure 6. Normalized values of eye area to w1118 control. Genotypes are indicated below the graph. Error bars indicate the standard deviation of the mean. Statistical
significance was determined with the Mann-Whitney U test. *, P < 0.05; ***, P < 0.001;
ns, not significant. n = 10 testes for each data point.
***
ns
ns
* *
***
***
Normalized value of eye area
23
Figure 7. Normalized values of eye circumference to w1118 control. Genotypes are indicated below the graph. Error bars indicate the standard deviation of the mean.
Statistical significance was determined with the Mann-Whitney U test. *, P < 0.05; ***, P < 0.001; ns, not significant. n = 10 testes for each data point.
***
ns
ns * *
* *
Normalized value of eye circumference
24
Figure 8. RT-PCR detection of OdsHmel expression in wild-type and transgenic male flies. Genotypes are indicated above the graph. rp49 was used as a loading control.
25
His-GFP expression in testes of OdsH+ and OdsH0 flies
To determine which testicular cells can be observed in the his-GFP background, it is necessary to clarify the construction of this transgenic line. his-GFP, synonymous with His2AvD-GFP, encodes a variant H2A.F/Z class histone of D. melanogaster fused with the green fluorescence protein of Aequorea victoria. Its localization to
chromosomes makes it a superb reporter gene to visualize nuclear activity during interphase, mitosis and meiosis (CLARKSON and SAINT 1999; WHITE-COOPER 2004). In post-meiotic spermatids, chromatin undergoes reorganization and condensation;
histones are modified, removed and replaced by protamines (JAYARAMAIAH RAJA and RENKAWITZ-POHL 2005; RATHKE et al. 2007). Therefore, in his-GFP testes, where GFP signal appears indicates an aggregate of chromosome in germ cells, including GSCs, spermatogonia, spermatocytes and early spermatids. The only somatic cells that can be easily observed are the dividing somatic stem cells (SSCs) and the newly divided somatic cyst cells. Unlike GSCs that undergo four rounds of mitosis and two rounds of meiosis, SSCs merely divide once during spermatogenesis, which makes them relatively few in number.
I first examined the His-GFP expression in the testes of OdsH+ and OdsH0 flies to uncover how OdsH affects the male germ cells in general. Immediately, I noted that His-GFP displayed various expression patterns in OdsH+ and OdsH0 testes at day 0. The distribution of His-GFP in testes was in the apical region (Figure 9, A and E), somewhat extending (Figure 9, B and F), or further extending towards the basal end (Figure 9, C and G). On the contrary, at day 32, His-GFP distribution was quite similar in OdsH+ and OdsH0 testes (Figure 9, D and H), from the apical tip towards the distal end.
Quantification assay showed a significantly higher GFP signal in OdsH+ testes than in
26
OdsH0 testes at day 0, but not at day 32 (Figure 10).
Sa-GFP expression in testes of OdsH+ and OdsH0 flies
Since I observed a significant difference of GFP expression between OdsH+; his-GFP and OdsH0; his-GFP, I investigated the expression pattern of Sa-GFP, a primary spermatocyte marker (CHEN et al. 2005), in testes of different ages by
immunostaining (Figure 11, A-F). Quantification assay showed at days 1, 15 and 55, no significant difference of GFP signal in sa-GFP, OdsH+ and sa-GFP, OdsH0 testes (Figure 12).
BamP-GFP expression in testes of OdsH+ and OdsH0 flies
Next, I characterized the BamP-GFP expression pattern in testes of different ages.
Immunostaining shows that, in both OdsH+; bamP-GFP and OdsH0; bamP-GFP testes, bam promoter-driven GFP expression either resembled the endogenous bam expression, which starts from the 4-cell cyst stage until the entry into premeiotic G2 phase and peaks in 8-cell cysts (Figure 13, A-C and G-I), or displayed somewhat misimpression pattern that spread extensively from the apical region to the distal end (Figure 13, D-F and J-L).
No quantification was done due to inconsistency in expression pattern.
27
Figure 9. His-GFP expression in testes of 0- and 32-day-old OdsH+ and OdsH0 flies.
Testes from (A-C) 0- and (D) 32-day-old flies of OdsH+; his-GFP and from (E-G) 0- and (H) 32-day-old flies of OdsH0; his-GFP were shown. Testes were stained with anti-GFP (green). Scale bars: 100 m.
OdsH0 ; his-GFP
Day 32
OdsH+ ; his-GFP
Day 0
A B C D
E F G H
28
Figure 10. The average volume of His-GFP signal per testis. The histogram depicts the average volume of His-GFP signal per testis from OdsH+; his-GFP and OdsH0; his-GFP flies at 0 and 32 days of age. Numbers on the bars indicate the sample size for each genotype. Error bars indicate the standard deviation of the mean. Statistical significance was determined with the Mann-Whitney U test. ***, P < 0.001; ns, not significant.
ns
(14) (15) (4) (3)
***
29
Figure 11. Sa-GFP expression in testes of 1-, 15-, and 55-day-old OdsH+ and OdsH0 flies. Testes from (A) 1- (B) 15- and (C) 55-day-old flies of sa-GFP, OdsH+ and from (D) 1- (E) 15- and (F) 55-day-old flies of sa-GFP, OdsH0 were shown. Testes were stained with anti-GFP (green) and anti-Fasciclin III (*, hub cells). Scale bars: 100 m.
A B C
D E F
sa-GFP, OdsH+ sa-GFP, OdsH0
Day 15 Day 55
Day 1
*
* * *
* *
30
Figure 12. The average volume of Sa-GFP signals in spermatocytes. The histogram depicts the average volume of Sa-GFP signal in spermatocytes per testis from sa-GFP, OdsH+ and sa-GFP, OdsH0 flies at 1, 15 and 55 days of age. Numbers on the bars indicate the sample size for each genotype. Error bars indicate the standard deviation of the mean. Statistical significance was determined with the Mann-Whitney U test. ns, not significant.
ns
(17) (16) (12) (14) (14) (10)
ns
ns
31
Figure 13. BamP-GFP expression in testes of 1-, 10-, and 32-day-old OdsH+ and OdsH0 flies. Testes from (AD) 1- (BE) 10- and (CF) 32-day-old flies of OdsH+; bamP-GFP and from (GJ) 1- (HK) 10- and (IL) 32-day-old flies of OdsH0; bamP-GFP were shown. Testes were stained with anti-GFP (green) and anti-Fasciclin III (*, hub cells). Scale bars: 100 m.
A B C
D E F
G H I
J K L
Day 10 Day 32
Day 1
OdsH+ ; bamP-GFPOdsH0 ; bamP-GFP
*
*
*
*
*
*
*
* *
*
* *
32
Discussion
OdsH RNAi strains should facilitate future studies on the functional divergence of duplicate genes and the normal function of OdsHsim
Four OdsH RNAi strains from VDRC and TRiP were tested in GMR>OdsHmel background. All of them rescued the ectopic expression phenotype to different levels, which indicates their efficiency. It seems unlikely to associate their efficiency with their targeting regions; all of the targeting regions fall within the exon 4 of OdsH, with one (JF02198 from TRiP) including the 3′ UTR (Figure 14 and Table 4). The observed less efficiency of GD51289 from VDRC is probably because the RNAi construct is
generated via the P-element transformation, and its expression is subject to the position effect. Ultimately, these strains should be examined in a fertility assay to see whether their phenotype—males produce less offspring—shows consistency with OdsH null mutant. These strains should facilitate the simultaneous knockdown of OdsH and unc-4 for future studies on the functional divergence between the two paralogs.
For the newly generated OdsHRNAi_simAB and OdsHRNAi_mauAB, two target sites for each construct fall within the homeobox (mir-6-1-OdsHsimA and mir-6-1-OdsHmauA) and the exon 4 (mir-6-1-OdsHsimB and mir-6-1-OdsHmauB), respectively (Figure 14).
Considering the sequence divergence of OdsH (Table 5), it is expected to see the incomplete rescue effect. Although the phenotypic rescue effect were observed, these two strains need to be further tested in the GMR>OdsHsim and GMR>OdsHmau
background to determine the knockdown efficiency.
33
Figure 14. OdsH structure and the target regions of UAS-OdsHRNAi strains used in this thesis. Self-made RNAi constructs
(mir-6-1-OdsHsimA, mir-6-1-OdsHsimB, mir-6-1-OdsHmauA and mir-6-1-OdsHmauB) are denoted as simA, simB, mauA and mauB. Note that target regions of GD51289, HMS01554 and simB do not overlap.
OdsH
HMS01554
simA mauB simB
mauA
KK103949 GD51289
JF02198
Intron
Homeobox UTR
Coding sequence Exon 3
Exon 1 Exon 2 Exon 4
34
Table 4. The target sequences of UAS-OdsHRNAi strains obtained from stock centers. OdsH 3′ UTR is underlined.
Strain Length (bp) Sequence (5′ to 3′)
GD51289 328 CGCAGCAAGCGGATGAAGAAAGCCATCGATCGGCAGGCGAAGAAGCTACAGGACAAGGGATTGGAAGTGGACTATGCCCGTCTGGAGGCCGAGTACCTAG CTGCCCACCAGGAGAACGGAGTGGATGAGAATAACTGGCTGGATGATGATGGCTACGATGATCTGCACATCGATGTGGTGGGCGTTGAACCGGAGTACGT GACCGGCGACAGTTTGGATCACTCGTTCTGCTCCTCCAGGACGTACCAGACGAAGAGCACCAGCAGCGAACTGGATTCCAATGATATGGGACTCCAGGGA AGAGTGGAAACTCCGCCACCACCACAGC
KK103949 331 GACAAGGGATTGGAAGTGGACTATGCCCGTCTGGAGGCCGAGTACCTAGCTGCCCACCAGGAGAACGGAGTGGATGAGAATAACTGGCTGGATGATGATG GCTACGATGATCTGCACATCGATGTGGTGGGCGTTGAACCGGAGTACGTGACCGGCGACAGTTTGGATCACTCGTTCTGCTCCTCCAGGACGTACCAGAC GAAGAGCACCAGCAGCGAACTGGATTCCAATGATATGGGACTCCAGGGAAGAGTGGAAACTCCGCCACCACCACAGCCACCGATGCAGAATAAGACCCTC TACAATTCGCCCTTCAGCATCGAATCCCTGT
JF02198 405 AGCTACAGGACAAGGGATTGGAAGTGGACTATGCCCGTCTGGAGGCCGAGTACCTAGCTGCCCACCAGGAGAACGGAGTGGATGAGAATAACTGGCTGGA TGATGATGGCTACGATGATCTGCACATCGATGTGGTGGGCGTTGAACCGGAGTACGTGACCGGCGACAGTTTGGATCACTCGTTCTGCTCCTCCAGGACG TACCAGACGAAGAGCACCAGCAGCGAACTGGATTCCAATGATATGGGACTCCAGGGAAGAGTGGAAACTCCGCCACCACCACAGCCACCGATGCAGAATA AGACCCTCTACAATTCGCCCTTCAGCATCGAATCCCTGTTGGGATCGTAACGAATATCGAATGTTGTACTATAATTCTACTATGATTTCGGGTGGTTAGC TAAGC
HMS01554 21 CAGCAGCGAACTGGATTCCAA
Table 5. Homology of the target sequences of mir-6-1-based RNAi constructs in Drosophila melanogaster, D. simulans and D. mauritiana.
The designed sequences are shown in bold letters. Mismatched nucleotides are indicated in red. Gaps are denoted as “-.” Percentages in parentheses refer to the level of sequence homology in comparison with the designed sequence (number of identical matches divided by the sequence length).
mir-6-1-OdsHmauA mir-6-1-OdsHsimA mir-6-1-OdsHmauB mir-6-1-OdsHsimB
D. melanogaster GGAGAGAGTCTTCCAGGGAAGT(86.4%) ATAGCGGTTTGGTTCCAGAATC(81.8%) ATATGGGACTCCAGG--GAAGA(63.6%) AATTCGCCCTTCAGCATCGAAT(86.4%) D. simulans GGAGAGAGCCTTCCAGGACAAT(90.9%) ATAGCGGTTTGGTGTCAAAATA ATATGGAAGTCCAGG--GAATA(77.3%) AAATCGTCCTTCAGCATCAAAT
D. mauritiana GGAGAAAGCCTTCCAGGAAAAT ATAAAGATATGGTTCCAAAATA(72.7%) ATATGGAAGTGGAAGTGGAATA AAATCGTCCTTCAGCATCAAAT (100%)
35
His-GFP expression implies GSC loss or disruption of subsequent spermatogenic development in young OdsH0 flies
Spermatogenesis in D. melanogaster is a series of cellular divisions and
metamorphosis events. At the apex of a testis tub lies a group of somatic support cells termed hub cells, which are surrounded by 6-12 germline stem cells (GSCs) (SPRADLING
et al. 2011). Each GSC is flanked by a pair of cyst progenitor cells (i.e. somatic stem cells, SSCs); both GSCs and SSCs are connected to the hub cells. As a GSC divides asymmetrically into two daughter cells, one maintains the GSC identity, and the other becomes a spermatogonium encapsulated by two somatic cyst cells. While these two cyst cells undergo no further division, the spermatogonium enters four rounds of synchronous mitotic division with incomplete cytokinesis, producing a cyst of 16 interconnected spermatogonia. The spermatogonia then embark on premeiotic DNA replication, and switch to an extended G2 phase for cell growth as spermatocytes (LIM et al. 2012). During the G2 phase, each spermatocyte substantially increases 25 times in volume. This premeiotic G2 phase continues for more than 3 days before two meiotic divisions proceed, which generates 64 round spermatids (WHITE-COOPER 2010). Still interconnected by a cytoplasmic bridge (ring canal), these round spermatids
subsequently enter the elongation program and become individual, mature sperms (Figure 15).
At day 0, His-GFP expression patterns were various in OdsH+ and OdsH0 testes.
This is probably due to the individual variation. Because the development of testes starts in the late larval stage (CHENG et al. 2008), it is likely that flies collected on the same day differ in sexual maturation. The quantification difference of GFP signal indicates there are more cells (predominantly germ cells) in OdsH+ testes. One explanation
36
(“explanation A”) is that OdsH0 flies fail to maintain as many GSCs as OdsH+ flies do, and GSC loss in OdsH0 testes causes a decrease in germ cell number. An alternative (“explanation B”) is that the GSCs are intact, but the subsequent development in OdsH0 testes is disrupted. However, at day 32, the quantification of GFP shows no difference between OdsH+ and OdsH0 testes. How does the age factor coordinate with these two explanations? One hypothesis is that the age effect on GSC maintenance, or the subsequent developmental stages, is accumulative and relatively minor than OdsH0 in young flies. In aged flies, OdsH0 effect is masked by the increasing age effect, so the germ cell number decreases in OdsH+ and OdsH0 testes without significant difference.
Both explanations are contradictory to the previous discovery that the GSC number is significantly less in 10-day-old, but not in 3-day-old, OdsH0 testes (CHENG et al. 2012).
37
Spermatogonium Spermatocyte Spermatid
Figure 15. Male germline differentiation in Drosophila melanogaster. The schema is not in scale. Expression periods of three GFP reporter lines used in this study, His2AvD-GFP, sa-GFP and bamP-GFP, are indicated. Note that the various spermatid “stages” are defined by the nuclear shape, not mitochondrial.
GSC Sperm
His2AvD-GFP
bamP-GFP
sa-GFP