Investigation of Camellia Tree Diseases in Taiwan and Establishment of a Rapid Detection Method
for Colletotrichum spp.
Tse-Yen Liu,1,2) Chao-Han Chen,1) Liang-Jong Wang,1) Ting-Hsuan Hung,2) Meng-Hao Hsu,1) Meng-Ling Wu3,4)
【Summary】
Camellia is commonly used for afforestation and as an important oil-producing crop with high economic value. Diseases that occur on aboveground parts of Camellia are considered to be major reasons causing declines in the quality and quantity of tea oil, but little is known about them. In this study, we investigated disease occurrence of Camellia in Taiwan and discovered that blister blight, leaf spot, and soft rot were major symptoms on Camellia farmlands in the north, while leaf spot and anthracnose were found on Camellia farmlands in central Taiwan. Through tis- sue isolation, fungal pathogen candidates of Camellia diseases were identified; 15 fungal isolates were obtained, subjected to nucleic acid extraction, and sequenced. Sequence blast results showed that these isolates included Colletotrichum spp. (known as an anthracnose disease pathogen), Al- ternaria sp., Botryosphaeria dothidea, Botryotinia fuckeliana (Botrytis cinerea), Lasiodiplodia pseudotheobromae, Neofusicoccum parvum, Pestalotiopsis spp., Phomopsis sp., and Sclerotinia sclerotiorum. All isolates were reported to be plant pathogens, but Camellia-related research is still lacking. A phylogenetic tree constructed with internal transcribed spacers (ITSs) of the 15 isolates in this study showed that pathogens of Camellia anthracnose were clustered into 2 species com- plexes of C. gloeosporioides and C. acutatum. Colletotrichum spp. isolates from different regions were all classified into the same group and were evolutionally closer than other potential Camellia leaf pathogens. Anthracnose is considered to be the most important Camellia disease with 2 species complexes, and a rapid detection method was first established in this study. Three primer sets were designed for the rapid detection by a polymerase chain reaction (PCR) based on our ITSs to iden- tify anthracnose disease on Camellia. The primer sets designed in this study showed high specific- ity and sensitivity and could successfully detect Colletotrichum spp. and distinguish between the C.
gloeosporioides and C. acutatum species complexes. This will contribute to achieving prevention strategies and early disease prevention of Camellia anthracnose. In this study, Colletotrichum spp.
and 8 other potential fungal pathogens were isolated from Camellia leaves, and results will be use- ful in understanding potential pathogens. With the establishment of a rapid detection method for serious anthracnose disease of Camellia, our study can be used as an important reference for early diagnosis, which will be valuable for disease control and management.
Key words: Camellia, Colletotrichum spp., molecular rapid detection, polymerase chain reaction.
Liu TY, Chen CH, Wang LJ, Hung TH, Hsu MH, Wu ML. 2021. Investigation of Camellia tree diseases in Taiwan and establishment of a rapid detection method for Colletotrichum spp..
Taiwan J For Sci 36(1):1-19.
研究報告
臺灣油茶病害調查及油茶炭疽病菌 快速檢測方法之建立
劉則言1,2) 陳昭翰1) 汪澤宏1) 洪挺軒2) 徐孟豪1) 吳孟玲3,4)
摘 要
油茶為臺灣早期的造林樹種,也是近年重要油料作物。油茶地上部病害為造成油茶產量與品質下 降的主要原因之一,但卻缺少相關之研究。本研究經由臺灣北部及中部油茶園的病害調查,發現在北 部油茶園葉部病害以餅病、葉斑病和軟腐病為主,中部則以葉斑病和炭疽病最為常見。進一步將感病 油茶葉片進行組織分離,獲得真菌分離菌株,並依菌落型態進行分群後,從中選取15株真菌分離株,
萃取核酸並定序與序列比對,發現油茶葉片及果實的植物病原真菌,以炭疽病菌屬(Colletotrichum spp.)真菌所占比例最高。此外,自油茶罹病葉上分離而得的菌株還包含Alternaria sp.、Botryosphaeria dothidea、Botryotinia fuckeliana (Botrytis cinerea)、Lasiodiplodia pseudotheobromae、Neofusicoccum parvum、Pestalotiopsis sp.、Phomopsis sp.、Sclerotinia sclerotiorum。這些真菌分離株皆有被報導為 植物病原菌,但在油茶上的研究仍相當缺乏。進一步將真菌分離株的ITS序列進行親緣關係分析,可以 發現感染油茶的炭疽病菌,主要來自於C. gloeosporioides和C. acutatum這兩個種群,並與自其他作物 上分離到的常見炭疽病菌歸為同一系群,與其他葉部潛在病原真菌在類緣關係上有所區別。有鑑於炭 疽病菌的兩個種群為油茶主要病原菌,本研究首次針對油茶上此兩個重要炭疽病菌種群發展快速檢測 方式。透過真菌核酸ITS區域,設計3組油茶炭疽病菌之專一性引子對,利用分子檢測技術(聚合酶鏈鎖 反應,PCR)來進行快速檢測。結果顯示本研究設計之引子對油茶炭疽病菌具良好專一性及靈敏度,並 可成功區分在油茶上主要造成炭疽病菌的不同種群。透過快速檢測方式的建立,未來將有助於病原菌 在潛伏感染時期的檢測。本研究自臺灣油茶罹病葉片上分離出炭疽病菌屬及其他8種真菌,有助於了解 油茶葉部之潛在病原菌,同時針對危害嚴重之油茶炭疽病發展快速檢測技術,可作為未來油茶病害防 治與健康管理之用,提供病害早期發現早期防治的管理模式。
關鍵詞:油茶、炭疽病菌、分子快速檢測、聚合酶鏈鎖反應。
劉則言、陳昭翰、汪澤宏、洪挺軒、徐孟豪、吳孟玲。2021。臺灣油茶病害調查及油茶炭疽病菌快速 檢測方法之建立。台灣林業科學36(1):1-19。
1) Division of Forest Protection, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan. 林業試驗所森林保護組,10066臺北市南海路53號。
2) Department of Plant Pathology and Microbiology, National Taiwan Univ., 1 Roosevelt Rd., Sec. 4, Taipei 10617, Taiwan. 國立台灣大學植物病理與微生物學系,10617台北市羅斯福路四段1號。
3) Office of Deputy Director, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan. 林業試驗所副所長室,10066臺北市南海路53號。
4) Corresponding author, e-mail:[email protected] 通訊作者。
Received January 2021, Accepted April 2021. 2021年1月送審 2021年4月通過。
INTRODUCTION
The Camellia tree is described as a small-sized evergreen tree of the genus Ca- mellia which is one of the four largest woody oil crops in the world. After the fruits are harvested and exposed to the sun, the seeds are hulled out and then squeezed to produce a slightly bitter oil. It is commonly known as the “bitter tea oil” and also referred to “Orien- tal olive oil” based on its beneficial effects on human health and its fatty acid composition which is comparable to that of olive oil (Chen et al. 1996, Chen and Sun 2011).
The planting of Camellia in Taiwan has mainly been for afforestation instead of seed harvesting and oil extraction. Extensive field cultivation and management are commonly adopted. In recent years as food safety issues have become of greater concern coupled with the low ratio of domestic self-produced oil materials, Camellia trees have the potential to be developed into an important local eco- nomic oil crop. Nowadays, the main species of domestic Camellia trees in Taiwan are Camellia oleifera, C. tenuifolia, and C. brevi- styla. The total planted area is about 1000 ha, and groves are mainly distributed in cen- tral, southern, and eastern counties, among which Chiayi County has the largest planted area, with the rest mainly distributed in Nan- tou, Hualien, Miaoli, and Taoyuan (Xie and Huang 2013, Zhang 2014).
Camellia trees prefer a warm and humid growing environment, mildly acidic soil with abundant organic matter and good drainage, and sufficient light. The applicable planting period of Camellia trees is from November to March. As planted Camellia seedlings grow slowly, good weed management, fertilizer management, pruning, and field pest manage- ment are required to promote the growth and improve yields and the quality of tea fruits (Chen and Sun 2011).
Generally, the first harvest of C. oleifera occurs in about the fourth year after planting.
In the early stage after planting, the severity of pests and diseases can affect seedling sur- vival, which requires disease and pest control.
Mature Camellia trees show stronger resis- tance to pests and diseases. If we better un- derstood the disease occurrence on Camellia farms, we could identify major diseases and develop preventive strategies through good field management. According to the monitor- ing and investigation of the occurrence of dis- eases on domestic Camellia farms, the main occurrence seasons and months of various diseases and pests can be determined which can be used as references for preventing dis- eases and pests. Common diseases on Camel- lia farms include anthracnose (also known as brown blight), net blister blight, leaf blight, and sooty mold. Among them, Camellia an- thracnose is considered to be dominant (Wu et al. 2013, Lu et al. 2017).
Anthracnose is a widespread plant dis- ease. Anthracnose pathogens are classified to the genus Colletotrichum which contains many different species and can infect the leaves and fruits of many crops. In the early phase of leaf infection, brown round or irregular spots can be observed, and sometimes concentric round patterns can also emerge. Late-phase leaf le- sions are often grayish-white with black dots in the central area; infections can produce ne- crotic spots or cracking on the fruit. In severe cases, it can cause premature fruit drop (Can- non et al. 2012, da Silva et al. 2020). Camellia infected by Colletotrichum spp. shows many symptoms, such as blackening of the branches, a tendency to break, tree trunk cancer, fruit cracking, and large-scale dropping of leaves and fruits. It also affects the growth vigor of trees and fruit yields; the entire plant may wilt and die (Wu et al. 2013).
The disease mainly occurs in warm and
humid environments and can be observed throughout the year in Taiwan. The peak pe- riod of disease occurrence is June to October.
The pathogen can be spread by wind, and in severe cases, it can reduce production by more than 50% in planted areas resulting in serious economic losses (Liu et al. 2019).
In previous studies, the main pathogen causing anthracnose was reported to be C.
gloeosporioides according to morphologi- cal characters. With the rapid progress of sequencing technology, the alignment of multiple gene sequences and bioinformatics analytical techniques are widely used to ef- fectively classify and identify Colletotrichum spp., and many new species of anthracnose pathogens have been reported. The concept of
“species” has been further developed into dif- ferent clades based on results of phylogenetic analyses of DNA sequences. In recent years, Colletotrichum spp. were identified into at least two different groups as C. gloeospori- oides and C. acutatum “species complexes”, which contain many similar species (Damm et al. 2012, Weir et al. 2012).
Although Camellia trees are important domestic afforestation tree species and oil- bearing crops, related disease research is still limited. In this study, we observed the disease occurrence and investigated the pathogen community on Camellia farms to understand dominant and potential pathogenic threats.
While anthracnose caused by Colletotrichum spp. was considered the most important aboveground disease of Camellia in previ- ous studies, we also investigated isolates of Colletotrichum spp. Although there have been several PCR primer pairs designed for detecting anthracnose on different crops with acceptable sensitivities, more-practical and -useful primer pairs were developed for Col- letotrichum spp. detection in this study. With latent infection and destructive characters
of Colletotrichum spp., specific primer pairs were designed to develop a sensitive and specific method for the rapid detection of anthracnose. These primer pairs can also be used to distinguish between the C. gloeospo- rioides and C. acutatum species complexes that cause anthracnose on Camellia trees. In addition, rapid detection would be valuable for monitoring Camellia anthracnose dis- eases, and potentially benefit disease control in the future.
MATERIALS AND METHODS Investigation of the disease occurrence of Camellia
To evaluate the disease occurrence and identify the most important diseases of Ca- mellia in Taiwan, 3 Camellia farming sites were selected. The first sample plot was lo- cated in Longtan, Taoyuan (in northwestern Taiwan), and the 2 other sample sites were in Lienhuachih, Nantou and Dongshi, Taic- hung (both in the central area of Taiwan). All samples were collected in 2017 to 2018, and leaf symptoms observed in the field were re- corded to identify the most common diseases on Camellia farms. Several diseased leaves were collected for further pathogen isolation to identify fungal pathogen groups infecting Camellia.
Isolation and incubation of fungal pathogens Fresh Camellia leaves with disease symp- toms were collected in the field. Potential fungal pathogens were isolated from infected plant tissues of samples by the following pro- cess. About 0.5~1 cm2 of plant tissues contain- ing healthy-diseased tissue junctions were cut out. The excised samples were immersed in 1%
NaClO for 90 s for surface sterilization. Then, leaf segments were transferred to sterilized distilled water 3 times for 30 s each to remove
any residual NaClO. These leaf segments were then placed on sterilized filter paper to remove any residual distilled water and finally placed on 2% water agar. The water agar plates were incubated at room temperature for 2~4 d. A single slice of fungal hyphae was cut from the edge of a leaf segment and transferred to pota- to dextrose agar (PDA) for further pure-culture incubation. After 2~4 d of incubation, hyphae from the edge of each mycelium were cut and transferred to new PDA plates for incubation to produce pure fungal isolates. All fungal isolates were stored in sterilized distilled water for further use.
Fungal genomic DNA extraction
The DNA extraction method was modi- fied from that of Hung et al. (1999). Fungal mycelium (0.16 g; about 0.4×0.4-cm agar plate covered with growing fungal hyphae of isolates) was scraped and placed into a 1.5-ml plastic tube. Hyphae were then homogenized with a plastic pestle. DNA extraction buffer (0.9 ml at pH 8.0; 100 mM Tris-HC1, 100 M EDTA, and 250 mM NaC1) and 0.1 ml 20%
Sarkosyl buffer were added. The suspension was vortexed and incubated at 55℃ for 1 h.
The tube was centrifuged at 6000 rpm for 5 min. The supernatant (0.8 ml) was saved, and 100 μl of 5 M NaC1 and 100 μl of cetyltri- methylammonium bromide (CTAB)/NaC1 (10% CTAB in 0.7 M NaC1) were added and incubated at 65℃ for 10 min. An equal volume of chloroform/isoamyl alcohol (24 : 1) was added, mixed thoroughly, and centrifuged at 11,000 rpm for 5 min. The aqueous suspension was saved. Equal volumes of phenol/chloro- form/isoamyl alcohol (25 : 24 : 1) were added, mixed thoroughly, and centrifuged at 11,000 rpm for 5 min. The aqueous suspension was saved. Isopropanol (0.6 volume) was added to precipitate nucleic acids, and the mixture was centrifuged at 12,000 rpm for 30 min. The pel-
let was washed with 70% ethanol to remove any residual CTAB. Briefly, the pellet was dried and resuspended in 100 μl of TE buffer (pH 8.0 : 10 mM Tris and 1 mM EDTA) and stored at -20℃. Genomic DNA (gDNA) of all 15 fungal isolates was extracted, and the inter- nal transcribed spacer (ITS) sequences were amplified by a PCR using the universal ITS4 and ITS5 primers to identify each isolate.
Fungal ITS sequence amplification Fungal ITS sequences were am- plified by a PCR using the primer sets ITS5 (5’-GGAAGTAAAAGTCGTAA- CAAGG-3’) and ITS4 (5’-TCCTCCGCT- TATTGATATGC-3’). A PCR was conducted in 25-μl reaction mixtures (10 mM Tris-HCl at pH 8.7, 50 mM KCl, 2 mM MgCl2, 0.25 mM of each dNTP, 1.25 units of Taq DNA polymerase, 0.2 μM each primer, and 2 μl of extracted DNA). All PCR reagents were procured from Promega (Madison, WI, USA). The PCR was carried out with the following setup: initial incubation at 94℃
for 2 min, followed by 30 cycles of a dena- turing step at 94℃ for 30 s, an annealing step at 56℃ for 30 s, and extension at 72℃
for 1.5 min, and a final extension of 5 min at 72℃. The amplified DNA was sequenced for further identification.
Phylogenetic analysis
ITS regions of fungal isolates derived from diseased Camellia leaves were amplified to examine their molecular characteristics.
Newly obtained sequences of each fungal isolate in this study were used to blast on the National Center for Biotechnology Informa- tion (NCBI) database. Sequences of model species with 99~100% similarity to our se- quences were derived from the database for further phylogenetic tree construction. Molec- ular Evolutionary Genetic Analysis (MEGA)
software vers. 6.0 (Tamura et al. 2013) was used for the sequence analysis. ClustalW implemented in MEGA vers. 6 was used for multiple sequence alignments of each ITS sequence which were manually adjusted to al- low for maximum sequence similarity (Tamu- ra et al. 2013). The phylogenetic analysis was inferred using the maximum-likelihood meth- od based on the Kimura 2-parameter model (Kimura 1980). Each clade was assessed us- ing a bootstrap analysis with 1000 replicates.
All positions containing gaps were eliminated (Tamura et al. 2013).
Colletotrichum sp. primer pair design and PCR amplification
Representative Colletotrichum ITS se- quences derived from the NCBI database (GeneBank) were used to evaluate specific regions for primer design (Appendix Fig. 1).
ITS sequences were aligned with ClustalW software, and regions for primer design were artificially selected. Conserved se- quences between the ITS5 and ITS4 regions were used for designing 3 primer sets, in- cluding 1 genus-specific primer pair and 2 species complex-specific primer pairs.
Sequences were as follows: genus-specific primer pair of CgG-F1: 5’-CGAAATGCGAT AAGTAATGT-3’ and CgG-R1: 5’-TACT ACGCAAAGGAGGCT-3’; 1st species complex-specific primer pair of CgS-F1:
5’-CTCTTGGTTCTGGCATCG-3’ and CgS- R1: 5’-TACCTTTGAGGGCCTACAT-3’; and 2nd species complex-specific primer pair of CgS-F2: 5’-GCAGCGAAATGCGATAAG-3’
and CgS-R2: 5’-CCCAGTGCGAGACGTAA A-3’. Two genus-specific primer pairs from previous studies were used for comparison:
Col1: 5’-AACCCTTTGTGAACRTACC- TA - 3 ’ a n d C o l 2 : 5 ’ - T TA C TA C G - CAAAGGAGGCT-3’ (Martinez-Culebras et al. 2003) and Col-F: 5’-AACCCTTTGT-
GAACATACCT-3’ and Col-R: 5’-CCACT- CAGAAGAAACGTCGTT-3’ (Cano et al.
2004). All primers used in this study are listed in Table 1. PCR conditions were the same as those described above.
PCR specificity and sensitivity tests of Colletotrichum sp. primer pairs
Fifteen fungal isolates obtained from Camellia leaves were subjected to primer pair specificity tests. The 5 primer pairs examined included CgG-F1/CgG-R1, CgS-F1/CgS-R2, CgS-F2/CgS-R2, Col1/Col2, and Col-F/
Col-R. Nucleic acids of pure cultured Col- letotrichum sp. isolates were serially diluted 10, 102, 103, 104, 105, 106, and 107 times, and sensitivities of individual primer pairs were tested. PCR conditions were the same as those described above.
Practical application tests of Colletotri- chum disease on Camellia
Camellia leaves suspected of being in- fected were collected from the field. Lesioned leaf tissues were cut out and used for total nu- cleic acid extraction following the instruction manual of the Plant Genomic DNA Mini Kit (Geneaid Biotech, New Taipei City, Taiwan).
Three primer pairs designed in this study were tested for practical application. Nucleic acids of pure-cultured Colletotrichum sp. were used as a positive control, whereas nucleic acids of healthy leaves and sterile water were used as negative controls. PCR conditions were the same as those described above.
RESULTS
Disease occurrence on Camellia
In this study, 3 sampling sites were se- lected to investigate the disease occurrence on Camellia in northern and central areas in Taiwan. Symptoms of leaves were observed
and recorded to represent Camellia disease occurrences. All different disease symptoms of Camellia diseases observed in the 2-yr investigation were classified and are shown in Fig. 1. Blister blight, algal spots, and soft rot were major symptoms observed at the sampling site in Longtan, Taoyuan. Anthrac- nose and leaf spots were major symptoms at Lienhuachih, Nantou, while anthracnose was the major disease in Dongshi, Taichung. The findings reflected that anthracnose and leaf spots were commonly observed in the central area, while blister blight and soft rot were only observed in northern Taiwan with its cooler climate.
Isolation, incubation, and identification of fungal pathogens
Fungal isolates derived from Camel- lia leaves exhibiting different symptoms included anthracnose, leaf spot, and soft rot.
All fungal isolates were first classified based on the colony morphology. Then we grouped all of the fungal isolates into 15 different groups. Representative morphologies of pure fungal isolates are shown in Fig. 2. Based on ITS sequences, 5 fungal isolates were identi- fied as Colletotrichum spp., 3 isolates were Pestalotiopsis spp., and 7 other isolates were Neofusicoccum parvum, Lasiodiplodia pseu-
dotheobromae, Alternaria sp., Phomopsis sp., Botryotinia fuckeliana, Botryosphaeria dothi- dea, and Sclerotinia sclerotiorum (Table 2).
Overall, Colletotrichum spp. causing anthrac- nose in the field occupied the largest propor- tion of total fungal isolates. Together with results of field disease occurrence investiga- tions, anthracnose was generally considered one of the dominant diseases on Camellia farms we investigated in Taiwan. While Pestalotiopsis spp. were considered to be potential fungal pathogens causing leaf spots, their pathogenicity still needs to be evaluated.
However, several potential fungal pathogens were also considered according to harmful records on other plants. It was mentioned that Sclerotinia sclerotiorum was the only fungus isolated from Camellia leaves with soft rot symptoms. These symptoms were observed on the northern Camellia farm in Taoyuan and could potentially be regarded as a new Camellia disease in Taiwan. Although numer- ous symptoms and potential pathogenetic fungal isolates were observed in our study, the following experiments focused on an- thracnose because of its high occurrence rate and characteristics of latent infections.
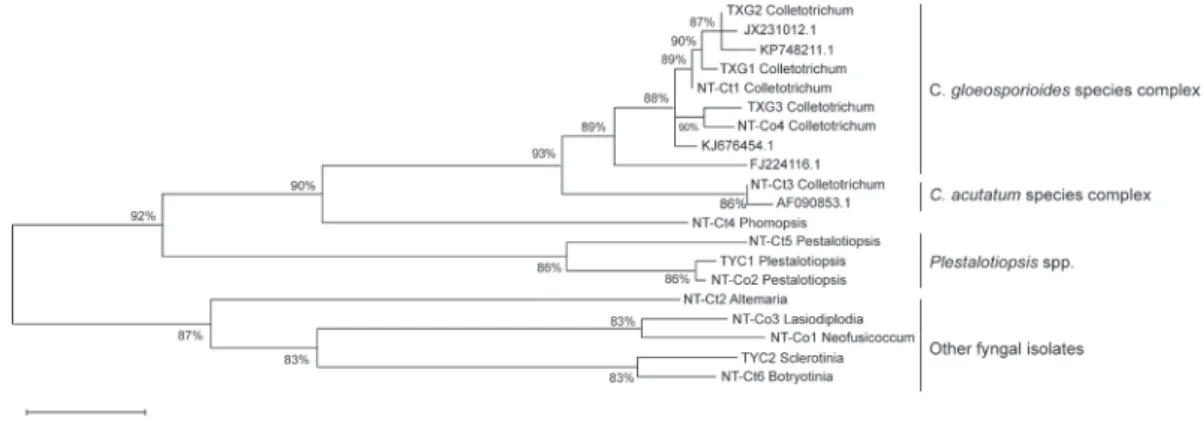
A phylogenetic tree of all the aforemen- tioned 15 fungal isolates was constructed with other representative fungal ITS se- Table 1. Primers used in this study
Genus-specific primer pair Reference
CgG-F1 5’- CGA AAT GCG ATA AGT AAT GT -3’
This study CgG-R1 5’- TAC TAC GCA AAG GAG GCT- 3’
Col1 5’-AAC CCT TTG TGA ACR TAC CTA-3’
Martinez‐Culebras et al. 2003 Col2 5’ -TTA CTA CGC AAA GGA GGC T-3’
Col-F 5’-AAC CCT TTG TGA ACA TAC CT-3’
Cano et al. 2004 Col-R 5’-CCA CTC AGA AGA AAC GTC GTT-3’
Species complex-specific primers This study CgS-F1 5’-CTC TTG GTT CTG GCA TCG-3’
CgS-F2 5’-GCA GCG AAA TGC GAT AAG-3’ This study CgS-R2 5’-CCC AGT GCG AGA CGT AAA-3’
acutatum species complex. We considered that besides C. gloeosporioides, C. acutatum could also be a potential anthracnose patho- gen to Camellia in Taiwan.
Specific PCR established for the rapid detection of Camellia anthracnose
Three primer sets of CgG-F1/CgG- R1, CgS-F1/CgS-R1, and CgS-F2/CgS-R2, were first designed. After basic PCR ampli- fication tests, the CgS-R1 primer showed less specificity and was eliminated from further studies. Since CgS-R2 showed good specificity with CgS-F1 and CgS-F2, the 3 primer sets of CgG-F1/CgG-R1, CgS-F1/
CgS-R2, and CgS-F2/CgS-R2 were selected as the genus- or species-specific primers.
While CgG-F1/CgG-R1 was designed as the Colletotrichum genus-specific primer set, quences derived from the NCBI, with a spe-
cial focus on Colletotrichum spp. According to the maximum-likelihood tree constructed with MEGA vers. 6, the fungal isolates were classified into different clades with differ- ent taxonomic statuses. Among all fungal isolates from Camellia, the Colletotrichum spp. clade was closest to the Phomopsis spp.
clade, then to the Pestalotiopsis spp. clade (Fig. 3). Alternaria sp., Botryotinia fuckeli- ana, and Sclerotinia sclerotiorum were sepa- rated into a quite-distinct group compared to other isolates. Alternaria sp. showed the highest difference with other fungi and was considered an outer group for phylogenetic tree construction. We identified that there were at least 2 Colletotrichum groups found:
one was grouped as the C. gloeosporioides species complex and the other was the C.
Fig. 1. Investigation of the disease occurrence of Camellia in Taiwan. Symptoms of blister blight, algal spots, and soft rot were observed in Longtan, Taoyuan (A to C); anthracnose (D to F) and leaf spot were observed in Lienhuachih, Nantou (G and H); and anthracnose was observed in Dongshi, Taichung (I).
CgS-F1/CgS-R2 and CgS-F2/CgS-R2 were designed as species-specific primers which could only amplify the DNA region of the C.
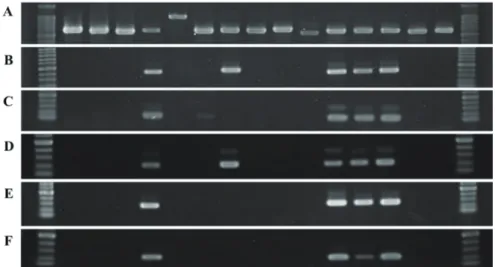
gloeosporioides species complex. To dem- onstrate the specificity of each primer set, 2 Colletotrichum spp.-specific primer sets, Col1/Col2 and Col-F/Col-R designed in a previous study, were used for comparison with the newly developed primer sets in this study, and ITS4/ITS5 were used as positive controls. All 15 representative fungal DNAs were used as targets to evaluate the speci- ficity of each primer set. DNA fragments amplified by Col1/Col2, Col-F/Col-R, CgG- F1/CgG-R1, CgS-F1/CgS-R2, and CgS-F2/
CgS-R2 were 416, 138, 203, 254, and 227 bp, respectively (Fig. 4B-F). Specificities of
the genus-specific primer sets of Col1/Col2 and CgG-F1/CgG-R1 were demonstrated when only Colletotrichum isolates showed the 416- and 203-bp DNA bands, while the remaining 10 isolates of other species had no PCR products with the 2 primer sets (Fig.
4B, D). Otherwise, CgS-F1/CgS-R2 and CgS-F2/CgS-R2 respectively amplified 254- and 227-bp DNA fragments of all C. go- leosporioides isolates (Fig. 4E, F). Also no other signal of PCR products of C. acutatum or other fungal isolates was detected by the 2 primer sets. However, the Col-F and Col- R primers considered to be a Colletotrichum genus-specific primer set in a previous study (Cano et al. 2004) showed no identification ability with the C. acutatum isolate with
Fig. 2. Morphologies of fungal isolates from diseased Camellia leaves. Fungal isolates of Neofusicoccum parvum, Pestalotiopsis sp., Lasiodiplodia pseudotheobromae, Colletotrichum gloeosporioides, Botryosphaeria dothidea, Alternaria sp., Colletotrichum acutatum, Phomopsis sp., Pestalotiopsis sp., and Botryotinia fuckeliana from Lienhuachih, Nantou (A to J);
isolates of Colletotrichum gloeosporioides from Dongshi, Taichung (K to M); isolates of Plestalotiopsis mangiferae and Sclerotinia sclerotiorum from Longtan, Taoyuan are shown (N and O). Sclerotinia sclerotiorum was the only fungal isolate derived from soft rot leaves and was considered to be its pathogen.
Table 2. List of investigated sites and fungal isolates in this study No.Fungal isolateHostCollection siteSymptomsColletotrichum detection* 1Neofusicoccum parvumCamellia oleiferaLienhuachih, NantouAnthracnose/Leaf spots- 2 Pestalotiopsis sp.C. oleiferaLienhuachih, NantouAnthracnose/Leaf spots- 3 Lasiodiplodia pseudotheobromaeC. oleiferaLienhuachih, NantouAnthracnose/Leaf spots- 4 Colletotrichum gloeosporioidesC. oleiferaLienhuachih, NantouAnthracnose+ 5 Botryosphaeria dothideaC. brevistylaLienhuachih, NantouLeaf spots- 6 Alternaria sp.C. brevistylaLienhuachih, NantouAnthracnose/Leaf spots- 7 Colletotrichum acutatumC. brevistylaLienhuachih, NantouAnthracnose+ 8 Phomopsis sp.C. brevistylaLienhuachih, NantouAnthracnose/Leaf spots- 9 Pestalotiopsis sp.C. brevistylaLienhuachih, NantouAnthracnose/Leaf spots- 10Botryotinia fuckelianaC. brevistylaLienhuachih, NantouAnthracnose/Leaf spots- 11 Colletotrichum gloeosporioidesCamellia sp.Dongshi, TaichungAnthracnose+ 12Colletotrichum gloeosporioidesCamellia sp.Dongshi, TaichungAnthracnose+
13 Colletotrichum gloeosporioidesCamellia sp.Dongshi, TaichungAnthracnose+ 14Pestalotiopsis mangiferaeCamellia sp.Longtan, TaoyuanAnthracnose/Leaf spots- 15Sclerotinia sclerotiorumCamellia sp.Longtan, TaoyuanSoft rot/Water-soaked spots-* Detection of Colletotrichum spp. was conducted by a PCR with CgG-F1/CgG-R1. +/- refers to positive or negative signals after PCR amplification.
Fig. 3. Phylogenetic tree of fungal isolates from Camellia leaves. Internal transcribed spacer (ITS) sequences derived from the NCBI database are shown with their accession numbers, and other ITS sequences with isolate numbers were derived in this study.
KP748211.1, JX231012.1, KJ676454.1, and FJ224116.1 were representative sequences of the Colletotrichum gloeosporioides species complex while AF090853.1 was considered to be the C. acutatum species complex. The phylogenetic analysis was conducted using the maximum- likelihood method with MEGA 6. Associated taxa clustered together in the bootstrap test (with 1000 replicates) are shown as percentages.
no amplified PCR products observed but showed a non-specific band with Alternaria sp. isolates (Fig. 4C).
Results showed that the CgG-F1/CgG- R1, CgS-F1/CgS-R2, and CgS-F2/CgS- R2 primer sets developed in this study had good specificities according to their original Colletotrichum genus-specific or C. gloeo- sporioides species-specific purpose. By combining the CgG-F1/CgG-R1 Colletotri- chum genus-specific primer set and one of the C. gloeosporioides species-specific sets of CgS-F1/CgS-R2 or CgS-F2/CgS-R2, we could also generally identify fungal isolates belonging to the C. gloeosporioides or C.
acutatum species complex (Fig. 4, Table 2).
While anthracnose is regarded as the most serious Camellia disease and usually occurs with a period of latent infection, we conclud- ed that the specific PCR established in this study with 3 newly developed primer sets will be valuable for Camellia anthracnose detection based on their high specificity. The
sensitivity and practical application of the PCR-based rapid detection method were fur- ther evaluated in this study.
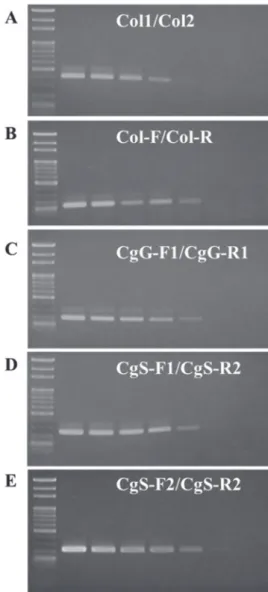
Sensitivities of the PCR primer sets gDNA extracted from 1 pure C. gloeo- sporioides isolate was used to verify the sen- sitivity of each aforementioned primer set (including 2 previous and 3 newly designed primer sets). The original DNA concentra- tion was approximately 11.3 ng/μl and was 10-fold serially diluted from 10× to 107× with sterile double-distilled H2O. Each con- centration of C. gloeosporioides DNA was amplified by the 5 primer sets. Sensitivities of the Col1/Col2, Col-F/Col-R, and CgG- F1/CgG-R1 primer sets were quite similar and could amplify C. gloeosporioides gDNA with the 10-4 dilution, i.e., about 1.13 pg/
μl gDNA in a 20-μl PCR. Comparing the above 3 primer sets, Col-F/Col-R and CgG- F1/CgG-R1 showed stronger PCR-amplified signals under lower concentrations and also
demonstrated better sensitivities than Col1/
Col2 (Fig. 5A-C).
The 2 C. gloeosporioides species- specific primer sets designed in this study were demonstrated to have higher sensitivi- ties than all 3 above Colletotrichum genus- specific primer sets. The sensitivity limita- tions of the 2 primer sets were certified to be able to detect about 0.1 pg/μl gDNA of C.
gloeosporioides (Fig. 5D, E). It was exciting to discover that the primer sets designed in our study had good specificities and sensitiv- ities that could be further used in Camellia anthracnose disease detection in the field. In addition, they can be used for early disease diagnosis of harvested fruit.
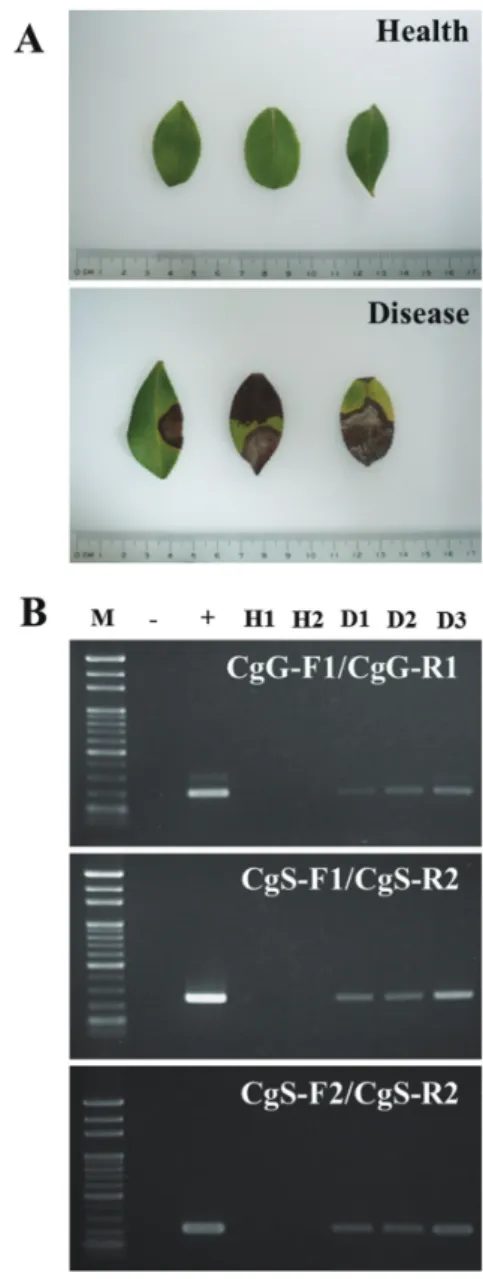
Practical applied validation
Anthracnose-diseased Camellia leaves collected from Camellia farms were used for validation of the practical rapid detec- tion method. Total DNA was extracted from both healthy and diseased leaves and was amplified by the 3 primer sets designed in this study. gDNA extracted from pure C. gloeosporioides mycelium was used as a positive control (Fig. 6).
PCR results demonstrated that all 3 primer sets successfully amplified target fragments from diseased samples without false-positive signals from healthy samples. Considering the specificity and sensitivity of the practi- cal tests, the 3 primer sets were judged to
Fig. 4. Rapid detection and specificity test of primers designed for Camellia anthracnose disease detection. All 15 fungal isolates from Camellia leaves were used to evaluate the specificity of each primer set. The fungal universal primer set ITS4/5 was used as a positive control (A). Amplification results of 2 Colletotrichum genus-specific primer sets, Col1/
Col2 (with a 416-bp amplicon) and Col-F/Col-R (with a 138-bp amplicon), from previous studies are shown (B, C). One Colletotrichum genus-specific primer set (CgG-F1/CgG-R1) and 2 C. gloeosporioides species complex-specific primer sets (CgS-F1/CgS-R2 and CgS-F2/
CgS-R2) designed in this study were estimated by PCR to have 203-, 254-, and 227- bp amplicons, respectively (D-F). Samples of each lane from left to right are molecular marker, Neofusicoccum parvum, Pestalotiopsis sp., Lasiodiplodia pseudotheobromae, C. gloeosporioides, Botryosphaeria dothidea, Alternaria sp., C. acutatum, Phomopsis sp., Pestalotiopsis sp., Botryotinia fuckeliana, C. gloeosporioides, C. gloeosporioides, C.
gloeosporioides, Pestalotiopsis mangiferae, Sclerotinia sclerotiorum, and molecular marker.
be reasonably reliable for the rapid detec- tion of Colletotrichum and for distinguish- ing between the C. gloeosporioides and C.
acutatum species complexes. This may also contribute to anthracnose disease control in the future.
DISCUSSION
Disease occurrences of Camellia in the field in Taiwan during 2017 to 2018 were in- vestigated in this study. Based on our inves- tigation, we demonstrated that the aboveg- round diseases of Camellia were similar to diseases on other tea plants, including blister blight, leaf spot, soft rot, and anthracnose (TPS 2019). Among the diseases caused by fungal pathogens on Camellia leaves, soft rot caused by S. sclerotiorum could potentially be considered a newly recorded disease on Camellia, and the disease usu- ally occurred in cool weather. Anthracnose is considered the most serious disease on Camellia, causing a decline in the quality of camellia oil, and it was also reported to be one of the most destructive disease on other crops, fruits, and afforestation trees, e.g., pepper, strawberry, tea trees, mango trees, stout camphor trees, etc. (Carlos et al. 2008, Kamle et al. 2013, Chung et al. 2020, Lin et al. 2020). According to a field investigation of leaf symptoms, anthracnose was a more- dominant disease than leaf spot; soft rot was only observed in several regions during specific seasons, i.e., late winter or early spring (Lu et al. 2019). Clearly, our findings showed that anthracnose and leaf spot were major Camellia foliage symptoms observed on Camellia farmlands in the northern and central areas of Taiwan. Both of these dis- eases seemed to be closely connected to tem- perature, with higher occurrences observed with higher temperatures in central Taiwan.
Fig. 5. Sensitivity test of specific primers designed for Camellia anthracnose disease detection. The DNA of Colletotrichum gloeosporioides was used with 10-fold serial dilution to evaluate the sensitivity of each primer set. All 5 primer sets of Col1/Col2 (A), Col-F/Col-R (B), CgG-F1/CgG-R1 (C), CgS-F1/CgS-R1 (D), and CgS-F2/
CgS-R2 (E) used in this study were tested at 7 different concentrations. Lanes 1 to 8 represent a molecular marker and PCR products of different concentrations with a series of 10-fold dilutions (at 1×, 10-1×, 10-
2×, 10-3×, 10-4×, 10-5×, and 10-6×).
Since some anthracnose and leaf spot symp- toms were atypical, further isolation was conducted to identify the compositions of the fungal pathogens on diseased leaves.
These findings generally corresponded to the pathogenic fungi isolated from dis- eased Camellia leaves. In the identifica- tion of the fungal compositions of poten- tial pathogens, at least 15 different fungal isolates were selected according to their colony types and further specified into 9 different species according to their micro- morphological structure and ITS sequences (Table 2). Fungal isolates included Colletot- richum spp., Pestalotiopsis spp., N. parvum, L. pseudotheobromae, Alternaria sp., Pho- mopsis sp., Botryotinia fuckeliana, Botryos- phaeria dothidea, and S. sclerotiorum. Some of these fungi were previously verified to be pathogenic to Camellia, i.e., Colletotrichum spp., Pestalotiopsis spp. and S. sclerotiorum (Vogelgsang et al. 1998, Thomma 2003, Lisa et al. 2006). Other fungi were also consid- ered to be potential pathogens or secondary pathogens according to their pathogenicity to at least 1 other host plant (Vogelgsang et al. 1998, Thomma 2003, Lisa et al. 2006).
Since the study involved only disease occur- rence and fungal isolation, further inocula- tion experiments are still needed to clarify the pathogenicity of each fungal isolate.
Despite the uncertified pathogenicity tests of potential fungal pathogens, our findings provide the first systemic study of the fungal composition of the field disease occurrence in Camellia in Taiwan. These results could help us infer potential and dominant patho- gens on Camellia farms. These results can serve as a valuable reference for Camellia disease management in Taiwan.
On the other hand, isolation of Colleto- trichum from Camellia anthracnose-infected leaves revealed that both C. gloeosporioides
Fig. 6. Practical application test of Camellia anthracnose disease-specific primers.
Healthy and diseased samples used for the practical application test (A). The 3 primer sets designed in this study were used for the practical application test (B). Lane 1, molecular marker; Lane 2, negative control (H2O as a template); Lane 3, positive control (DNA of Colletotrichum gloeosporioides as a template); Lanes 4 and 5, DNA of healthy leaves; Lanes 6 to 8, DNA of diseased leaves.
and C. acutatum were anthracnose-causing pathogens. While fungi of the genus Col- letotrichum are distributed into several spe- cies complexes, C. gloeosporioides and C.
acutatum are the 2 major species complexes (Damm et al. 2012, Weir et al. 2012). Our results expanded our previous knowledge and revealed that there could be additional Colletotrichum species causing anthracnose on Camellia (Chen et al. 2017). However, the pathogenicities of different Colletotri- chum isolates especially belonging to differ- ent Colletotrichum species complexes have still not been determined. It would be inter- esting to clarify the different pathogenicities or virulences of isolates among different Colletotrichum species complexes derived from Camellia, which would also be useful for evaluating plant resistance. We intend to continue pursuing this investigative lead in further experimental studies.
Since early symptoms and various spots caused by Colletotrichum species are some- times hard to diagnose by traditional meth- ods, one of the major objectives of our study was to develop a rapid detection method with higher sensitivity and specificity for detecting Colletotrichum spp. Among the 3 primer sets we designed in this study, one was used for Colletotrichum genus-specific detection and the other two were used for C.
gloeosporioides species complex-specific detection. All 3 primer sets showed good specificity and sensitivity and were quite useful for detecting diseased plant samples.
It is also worth noting that the 2 species complex-specific primer sets of C. gloeospo- rioides, CgS-F1/CgS-R2 and CgS-F2/CgS- R2, demonstrated higher sensitivities. The 2 primer sets showed 10-fold sensitivity com- pared to all other primer sets tested in this study and could be used to detect pathogens under as low a DNA concentration as 0.1
pg. Although application of the primer sets in this study still needs further evaluation, preliminary results indicated their possibili- ties for long-term monitoring of changes in pathogen populations and disease manage- ment in the early stage.
The primer sets developed in our study can be applied to identify Colletotrichum spp. (Torres-Calzada et al. 2011). Using different combinations of primer sets, the genus-specific primer set (CgG-F1/CgG- R1) together with 1 species complex-specific primer set (CgS-F1/CgS-R2 or CgS-F2/CgS- R2), we could detect Colletotrichum spp.
and distinguish between the C. gloeospori- oides and C. acutatum species complexes.
When fungal DNA could be amplified by both genus-specific and species complex- specific primer sets, the fungus belonged to the C. gloeosporioides species complex. In contrast, if the fungal DNA was only ampli- fied by the Colletotrichum genus-specific primer, the fungus should belong to other Colletotrichum spp. such as those in the C.
acutatum species complex.
While anthracnose widely occurs in different plants and Colletotrichum is com- posed of many species complexes (Armit- age et al. 2020), the genus-specific CgG- F1/CgG-R1 primer pair was supposed to be capable of widely amplifying all Col- letotrichum spp. even from different plants.
Similar applications could also be extended to the CgS-F1/CgS-R2 and CgS-F2/CgS-R2 primer sets for detecting Colletotrichum spe- cies. Although amplification was only tested on fungal isolates from Camellia and many more Colletotrichum isolates should be ac- quired for further assessment, our study still provides a crucial step toward anthracnose rapid detection and Colletotrichum species identification.
CONCLUSIONS
This research first revealed Camellia disease occurrences in northern and central Taiwan. According to leaf symptoms, we clas- sified the symptoms into blister blight, algal spot, leaf spot, soft rot, and anthracnose. These symptoms were quite similar to those of dis- eases on tea trees. To understand the fungal composition of potential pathogens, fungal isolates from diseased Camellia leaves were identified as Colletotrichum spp., Pestalotiop- sis spp., Neofusicoccum parvum, Lasiodiplodia pseudotheobromae, Alternaria sp., Phomopsis sp., Botryotinia fuckeliana, and Sclerotinia sclerotiorum. Among all the aboveground dis- eases, anthracnose was considered the most important one with its high observation and isolation rates that were consistent with previ- ous considerations. It is worth noting that C.
gloeosporioides was not the only species of Colletotrichum causing anthracnose on Ca- mellia, while C. acutatum was also isolated from diseased Camellia leaves. Traditionally, anthracnose disease control has been quite dif- ficult based on time-consuming identification and latent infection by Colletotrichum spp.
In this study, a rapid detection PCR method was established with newly designed genus- specific and species complex-specific primer sets. All the 3 primer sets showed good speci- ficity and sensitivity and could be applied to detect pathogens on plant tissues. They can be further used for field disease management even with a large number of samples. On the other hand, combinations of our primer sets were also useful in distinguishing between the C. gloeosporioides and C. acutatum species complexes. To our knowledge, there has been no comprehensive study of Camellia diseases.
In this study, we investigated potential fungal pathogens of Camellia and established a rapid detection method for Colletotrichum identifica-
tion which can be helpful for Camellia disease management in Taiwan.
LITERATURE CITED
Armitage AD, Nellist CF, Bates HJ, Zhang L, Zou X, et al. 2020. Draft genome sequence of the strawberry anthracnose pathogen Col- letotrichum fructicola. Microbiol Resourc An- nounce 9(12):e01598-619.
Cannon PF, Damm U, Johnston PR, Weir BS. 2012. Colletotrichum--current status and future directions. Stud Mycol 73(1):181-213.
Cano J, Guarro J, Gené J. 2004. Molecular and morphological identification of Colletotri- chum species of clinical interest. J Clin Micro- biol 42(6):2450-4.
Garrido C, Carbú M, Fernández-Acero FJ, Budge G, Vallejo I, Colyer A, et al. 2008. Iso- lation and pathogenicity of Colletotrichum spp.
causing anthracnose of strawberry in southwest Spain. Eur J Plant Pathol 120:409-15.
Chen JR, Sun WZ. 2011. Camellia cultiva- tion and utilization. Tainan Dist Agric Newsl 76:5-7.
Chen YJ, Wang LL, Chen XP, Li Y. 2011.
Oil content and fatty acid composition of Camellia oleifera seed in Guangxi. Food Sci 32:172-6.
Chen Y, Qiao W, Zeng L, Shen D, Liu Z, Wang X, et al. 2017. Characterization, patho- genicity, and phylogenetic analyses of Colleto- trichum species associated with brown blight disease on Camellia sinensis in China. Plant Dis 101(6):1022-8.
Chen YZ, Wang DB, Su YQ, Wang B. 1996.
Camellia oleifera: selective breeding of “cold dew seed” type of cultivar clones and analysis of their fatty acid composition. Econ For Res 14:1-5.
Chung PC, Wu HY, Wang YW, Ariyawansa HA, Hu HP, Hung TH, et al. 2020. Diversity and pathogenicity of Colletotrichum species
causing strawberry anthracnose in Taiwan and description of a new species, Colletotrichum miaoliense sp. nov. Sci Rep 10:14664.
Crouch JA, Clarke BB, White JFJ, Hillman BI. 2009. Systematic analysis of the falcate- spored graminicolous Colletotrichum and a de- scription of six new species from warm-season grasses. Mycologia 101(5):717-32.
da Silva LL, Moreno HLA, Correia HLN, Santana MF, de Queiroz MV. 2020. Colletot- richum: species complexes, lifestyle, and pecu- liarities of some sources of genetic variability.
Appl Microbiol Biotechnol 104(5):1891-904.
Damm U, Cannon PF, Woudenberg JH, Crous PW. 2012. The Colletotrichum acutatum species complex. Stud Mycol 73(1):37-113.
Hung TH, Wu ML, Su HJ. 1999. Develop- ment of a rapid method for the diagnosis of citrus greening disease using the polymerase chain reaction. J Phytopathol 147:599-604.
Kamle M, Pandey BK, Kumar P, Kumar MM. 2013. A species-specific PCR based assay for rapid detection of mango anthrac- nose pathogen Colletotrichum gloeospori- oides Penz. and Sacc. J Plant Pathol Microb 4(6):1000184.
Kimura M. 1980. A simple method for esti- mating evolutionary rate of base substitutions through comparative studies of nucleotide se- quences. J Mol Evol 16:111-20.
Lin SR, Yu SY, Chang TD, Lin YJ, Wen CJ, Lin YH. 2020. First report of anthracnose caused by Colletotrichum fructicola on tea in Taiwan. Plant Dis 10.1094/PDIS-06-20-1288- PDN.
Lisa MK, Maile EV, Francis TZ. 2006. Iden- tification and characterization of Pestalotiopsis spp. causing scab disease of guava, Psidium guajava, in Hawaii. Plant Dis 90(1):16-23.
Liu TY, Wang LJ, Hsu MH, Wu ML. 2018.
Research on health management to the major disease and pests of Camellia. Taiwan Forestry Research Institute Annual Report 2018. Taipei,
Taiwan: Taiwan Forestry Research Institute. p 44-5.
Lu PK, Chen ZS. 2017. Illustration of pests of Camellia oleifera in Yilan and Hualien.
Hualien, Taiwan: Hualien District Agricultural Research and Extension Station. p 27-33.
Lu PK, Hsieh WT, Pan HR, Ou JH. 2019.
First report of Haradamyces foliicola causing leaf blight on oil tea (Camellia oleifera) in Tai- wan. Plant Dis 103(5):1027.
Martinez-Culebras PV, Querol A, Suarez- Fernandez MB, Garcia-Lopez MD, Barrio E. 2003. Phylogenetic Relationships among Colletotrichum pathogens of strawberry and design of PCR primers for their Identification.
J Phytopathol 151(3):135-43.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolu- tionary genetics analysis version 6.0. Mol Biol Evol 30(12):2725-9.
Thomma BPHJ. 2003. Alternaria spp.: from general saprophyte to specific parasite. Mol Plant Pathol 4:225-36.
TPS. 2019. List of plant diseases in Taiwan (5th edition). Taipei, Taiwan: Taiwan Phytopathol- ogy Society (TPS). p 52-4.
Torres-Calzada C, Tapia-Tussell R, Quijano- Ramayo A, Martin-Mex R, Rojas-Herrera R, Higuera-Ciapara I, et al. 2011. A species- specific polymerase chain reaction assay for rapid and sensitive detection of Colletotrichum capsici. Mol Biotechnol 49:48-55.
Vogelgsang S, Watson A, DiTommaso A.
1998. Effect of moisture, inoculum production, and planting substrate on disease reaction of field bindweed (Convolvulus arvensis L.) to the fungal pathogen, Phomopsis convolvulus.
Eur J Plant Pathol 104:253-62.
Weir BS, Johnston PR, Damm U. 2012. The Colletotrichum gloeosporioides species com- plex. Stud Mycol 73(1):115-80.
Wu ML, Zhuang LM, Chen YZ, Xie WW.
2013. Disease and control of Camellia. TFRI
Extension Folder. Taipei, Taiwan: Taiwan For- estry Research Institute. No. 107.
Xie JM, Huang YX. 2013. Investigation of the current development of Camellia industry
in Taiwan. For Res Newsl 20(5):13-22.
Zhang TW. 2014. Overview of Camellia In- dustry in Taiwan. Hualien Dist Agric Newsl 89:14-7.
Appendix Fig. 1. Sequences used for Colletotrichum-specific primer design. Representative internal transcribed spacer (ITS) sequences of Colletotrichum spp. were derived from the NCBI. Sequences belonging to the C. gloeosporioides species complex are KF836743.1, DQ084497.1, EF063686.1, and AY902476.1. Sequences belonging to the C. acutatum species complex are AF090853.1, KC790936.1, and LC194228.1. Other sequences of Colletotrichum spp. included C. fioriniae (MT492141.1), C. boninense (MK311221.1, JQ926743.1, and JF683320.1), C. crassipes (KP050648.1 and JN198430.1), and C. fuscum (KF498868.1, JQ005762.1, and EU400144.1).