中國醫藥大學
醫學檢驗生物技術學系碩士班 碩士學位論文
SDF-1 及 CXC4 在口腔鱗狀上皮細胞癌之癌化過程所扮演的角色
Potential role of SDF-1 and CXCR4 in the tumorigenesis of oral squamous cell carcinoma
指導教授 : 莊淨媛 副教授 Adviser : Jing-Yuan Chuang, Ph. D.
學生姓名 : 陳佳玉
Graduate student : Chia-Yu Chen 學號 : 9473004
中華民國九十六年七月 July 2007
誌謝 誌謝誌謝 誌謝
感謝吾師莊淨媛副教授,除了研究領域上的教導,讓我學術概念有 長足的進步外,對於人生規劃及態度,您都引導我以積極卻又不失縝密 思考的方式前進,問題出現時以沉穩冷靜的態度面對,面對它並解決它;
當我碰到所有困難時,老師總是很關心並耐心的分析許多中肯的見解,
令我獲益良多,由衷地感謝我的老師。
感謝這兩年來幫助過我所有的人:學弟妹怡琳、琮仁、協成,感謝 你們對於實驗的協助以及對我的關心;實驗室同學建豪、惠琳、依伶、
惠如、佳芳、盈如、詩雯、素連、郁芬、瑞辰…感謝你們陪我ㄧ起共渡 這兩年的研究時光;小文、桃子、佳靜、彗怡、琪恩、益誠、Edouard、
穎英、孟芳…感謝你們在我碰到挫折時給我的鼓勵跟支持,那對我真的 很重要;感謝鍾景光老師、湯志昕老師及魏宗德老師、均隆學長、建良 學長、柏村學長熱心給予我實驗上的協助;感謝中台的黃蘭如老師,您 總是熱心的看待學生的每ㄧ件事;感謝系上的儲姐及顏先生還有所有幫 助過我的人;因為有你們,我才得以順利完成此篇論文。
感謝我的家人、我的妹妹佳言、佳足以及我的父母親,在求學期間 以經濟及精神上支持我,你們始終是我最大的動力。感謝所有給我鼓勵 及幫助我的人,再次謝謝你們。謹將此論文獻給我最愛的父母。
陳佳玉 謹致於 中華民國九十六年 七月
摘 摘
摘摘 要要要 要
口腔癌發生率居全國癌症第四位,每年新增罹病人口達 3000 人之多,在 台灣也是最快速成長癌症之ㄧ。趨化激素(chemokine)是ㄧ群可調控細胞 遷移的發炎小分子(約 8~14 kDa), Stromal cell-derived factor-1 (SDF-1) 是趨化激素 CXC 家族的ㄧ員,它對於血球細胞有強烈的化學趨化作用並 刺激細胞的生長發育。有許多文獻指出,SDF-1 及其受器 Chemokine Receptor-4 (CXCR4) 在許多癌症的致癌過程中扮演著重要的角色。據此 觀念,我們著手探討 SDF-1 或 CXCR4 是否與口腔癌相關,並探究 SDF-1 或 CXCR4 在口腔癌的癌化過程中所扮演的角色。首先我們利用免疫組織化學 染色(Immunohistochemical staining)偵測臨床口腔癌組織,發現有相 當高比例病人的癌組織中表達 SDF-1 和受體 CXCR4,雖然強度和有表達之 癌細胞所佔比率不同。六株口腔癌細胞株也強烈表現 CXCR4 蛋白,其中 由台灣培育出之口腔癌細胞株 TW2.6 也同時表達 SDF-1。再者,口腔癌細 胞株經 SDF-1 作用後,除了促進細胞增生(proliferation)能力外,也增 強口腔癌細胞之轉移(migration)以及侵潤(invasion)的能力。以上實驗 結果暗示,在口腔癌的致癌過程中 SDF-1/CXCR4 扮演著相當重要且多重 的角色。未來,SDF-1/CXCR4 在口腔癌癌化之機制將繼續被探索,同時測 試 CXCR4 的佶抗劑 AMD3100 是否可抑制腫瘤細胞的生長和轉移,以尋求臨 床上治療口腔癌新藥。
Abstract
Oral cancer is the fourth most common cancer reported nationwide, with an estimated 3000 new cases annually. Chemokines are a group of small (8-14 kDa) pro-inflammatory molecules that can regulate cell trafficking. Stromal cell-derived factor-1 (SDF-1) is a member of CXC chemokine family and also a potent chemoattractant for hematopoietic cells. Many studies implicated that, in many kinds of cancers, SDF-1 act with its specific receptor CXCR4 may play an important role in tumorigenesis process. Based on this conception, we started to investigate the relationship between SDF-1/CXCR4 and oral squamous cell carcinoma (OSCC), and also explore what role does SDF-1/CXCR4 signaling route play in the tumorigenesis of OSCC. First, we observed that CXCR4 and SDF-1 proteins are expressed in OSCC tissues of clinical oral cancer patients in a high ratio circumstances through immunohistochemical staining. CXCR4 was also expressed in all six OSCC cell lines we investigated. TW2.6 cell line, which was established from Taiwan, also expressed SDF-1 proteins. SDF-1 treatment not only enhanced the proliferation ability but also increased migration and invasion activities in these OSCC cell lines. Collectively, these results imply that the SDF-1/CXCR4 axis may play important and multiple roles in the tumorigenesis of OSCC. In the future, the tumorigenesis mechanism of SDF-1/CXCR4 in oral cancer will be investigated continually. Furthermore, the CXCR4 antagonist, AMD3100, will be tested whether it has the ability to inhibit the growth and metastasis of tumor cells. It may be considered as a potential strategy of the therapy for OSCC patients.
Contents
誌謝………i
摘要………..…...……..ii
Abstract……….……...iii
Contents………iv
Figure index………...…..vi
Table index………. ……vii
Chapter I. Introduction………...………...……1
1.1 Oral cancer………..…………..1
1.2 Chemokines……….….……….2
1.3 Stromal cell-derived factor-1 (SDF-1)………..…..……..3
1.4 Chemokine Receptor-4 (CXCR4)………….………..………..4
1.5 Specific Aim……… ……….5
Chapter II. Reference Investigation……….………….…….6
2.1 SDF-1/CXCR4 in normal cell………...……6
2.2 SDF-1/CXCR4 signaling pathway………...………….8
2.3 SDF-1/CXCR4 associated malignancy……...……….…… …9
2.4 SDF-1/CXCR4 axis inhibitors……….………....11
Chapter III. Materials and Methods………...………….13
3.1 Materials………..13
3.2 Methods………...…………17
3.2.1 Tissue samples………17
3.2.2 Immunohistochemical staining…….………….……….17
3.2.3 Cell culture………..17
3.2.4 RNA Extraction………..18
3.2.5 Reverse transcription-PCR……….…....19
3.2.6 Western blot……….….…….….20
3.2.7 Proliferation assay………..….………21
3.2.8 Migration assay……….…...…...…21
3.2.9 Invasion assay……….…..…...…...22
Chapter IV. Results………..…………..23
4.1 Immunohistochemical staining of CXCR4 and SDF-1 in OSCC tissue………..…….…...23
4.2 CXCR4 and SDF-1 expression in oral cancer cell lines…...24
4.3 SDF-1 increases proliferation of oral cancer cell lines…....…25
4.4 SDF-1 stimulats oral cancer cell migration ……...…….……26
4.5 SDF-1 has the potential to promote invasion of oral cancer cells through matrigel………27
Chapter V. Discussion……….……….….…….28
Reference……….……. ..39
Figure index
Figures in Introduction:
Figure 1. Different levels of regulation of CXCR4 function………...6 Figure 2. SDF-1/CXCR4 signaling pathway……….8 Figure 3. Importance of the CXCR4 chemokine receptor and its ligand,
CXCL12, in the tumor microenvironment and for targeted
metastasis……….9 Figure 4. Chemical structure of the bicyclam AMD3100,
1,10-[1,4-phenylene-bis(methylene)]-bis-1,4,8,11-
azatetradecane……….…. 12 Figures in Results:
Figure 5. Expression of CXCR4 protein in human primary oral cancer
tissue…..………..31 Figure 6. Expression of SDF-1 protein in human primary oral cancer
tissue……… ……..… …32 Figure 7. SDF-1 involved in the tumorigenesis of OSCC……….….….33 Figure 8. Western blot analysis of CXCR4 in six OSCC cell lines…….……34 Figure 9. RT-PCR analysis of the mRNA expression of CXCR4 and SDF-1 in six OSCC cell lines……….…….35 Figure 10. Proliferation assay of TW2.6 and HSC3 after SDF-1
stimulation……….…..36 Figure 11. Migration increased by SDF-1 stimulation………...….37 Figure 12. Invasion of TW2.6 and HSC3 through matrigel after SDF-1
stimulation……….…38
Table index
Table 1. Primer sequences and the length of each PCR products………...…20
Chapter I. Introduction
Oral cancer
Oral cancer is the largest group in the head and neck cancer category.
Common names including mouth cancer, tongue cancer, and throat cancer. It is the fourth most common cancer reported nationwide, with an estimated 3000 new cases annually. Oral cancer is also one of the fastest increasing malignancies in Taiwan (Bureau of health promotion, Taiwan, 2005). Studies have shown that during the last decade in Taiwan, oral squamous cell carcinoma (OSCC) has ranked fifth among cancer-related mortality Betel quid (BQ) consumption is an addictive habit with psychoactive properties, used by an estimated 600 million people worldwide. In Taiwan, 50% of oral cancer patients are chewing betel quid and more than 1000 people have died of OSCC every year. About 90% of patients were male (1). Squamous cell carcinoma (SCC) is the most common type of oral carcinoma. It accounts for about nine of every ten oral malignancies, and it is a major cause of cancer morbidity and mortality.
Oral cancer is associated with a wide range of physical or chemical agents including alcohol, tobacco, betel quid, electrogalvanic stimulation, microbial infection, and ultraviolet radiation (2, 3).There are many studies indicate that environmental factors play an important role in the pathogenesis of oral cancer. The morbidity of oral cancer in the United States and Western Europe, where the major risk factors of oral cancers are thought to be alcohol drinking and tobacco smoking, is rare. In contrast, oral cancer occurs frequently in Taiwan where chewing of betel quid (BQ) with or without
tobacco is popular (4).
Chemokines
Cytokines are soluble extracellular proteins. These glycoproteins are produced constitutively by cells for the purpose of regulating its own functions (autocrine effect) or adjacent cells (paracrine effect). Theses molecules are crucial intercellular regulators and mobilizers of cell growth, differentiation, cell death, angiogenesis, and development and repair processes aimed to the restoration of homeostasis. Cytokines comprise interleukins (initially thought to be produced exclusively by leukocytes), lymphokines (initially thought to be produced exclusively by lymphocytes), monokines (initially thought to be produced exclusively by monocytes), interferons (initially thought to be involved in antiviral responses), colony-stimulating factors (initially thought to support the growth of cells in semisolid media), chemokines (thought to be involved in chemotaxis), and a variety of other proteins (5).
Chemokines are a group of small (8-14 kDa) pro-inflammatory molecules that can regulate cell trafficking through interactions with aG-protein coupled seven-transmembrane receptors (6). To date, there are more than 50 chemokines and 20 chemokine receptors were included in human chemokine systems (7). Chemokines have been divided into four major subfamilies, CC (known as β-chemokines ) , CXC (α-chemokines), CX3C and XC. Subfamilies based upon the arrangement and the number of the N-terminal cysteine residues, with zero, one or three amino acids separating the first two cysteins (7). Chemokines play fundamentantal roles in the homeostasis, functions of the immune systems, angiogenesis (8),
migration, proliferation, lymphocyte homing, cell development, and inflammatory process (28, 43~45, 60).
Stromal cell-derived factor-1 (SDF-1)
SDF-1 is a member of CXC chemokine family. It is a potent chemoattractant for immature and mature hematopoietic cells (9). SDF-1 was cloned from a bone marrow derived stromal cell line and characterized as a pre-B-cell growth stimulating factor (PBSF) (10, 11). It plays crucial roles in embryonic development. Doitsidou. M. et al. reported that SDF-1-deficient mice died in uterus with severe cardiac septal defects and poorly developed marrow (12). SDF-1 is involved in the homing processed of hematopoietic stem cells and promotes human and murine colony-forming progenitor cells survival and proliferation (48). On the other hand, SDF-1 also been demonstrated that has the ability to induce the migration of SCID repopulating cells.
There are already 6 SDF-1 isoforms which have been identified: SDF-1α, β, γ, δ, ε, φ (13). Two major isoforms: SDF-1α and SDF-1β arise from a single gene through alternative splicing and encode 89 and 93 amino acid proteins, respectively (13). The third isoform, SDF-1-γ, is highly expressed in the rat brain and be induced by ischemia injury in rat heart (13). The first three exons in other four isoforms are the same but the fourth exons are different (13). The human SDF-1 gene locates on chromosome 10q11.1 (11).
SDF-1 is widely expressed in many cell types such as dendritic cells, endothelium cells, stromal cells, skeletal muscle cells, heart, liver, kidney, brain and immature osteoblast (13-19). SDF-1 correlates with tissue repair and regeneration such as ischemia, liver damage, myocardial infarction, and
irradiation (20-23).
Chemokine Receptor-4 (CXCR4)
Most chemokines usually bind to multiple receptors and the same receptor can be recognized by many ligands, but CXCR4 is a private receptor for SDF-1. CXCR4 was originally cloned by five separate groups as an G-protein coupled receptor and erroneously classified as a neuropeptide Y receptor (24). 1994, Loetscher et al. named this receptor LESTR (leukocyte-derived seven-transmembrane domain receptor) and suggested it may function in the activation of inflammatory cells (25). After two years, Bleul, C.C et al. identified LESTR was an HIV-1 co-receptor and also a receptor of SDF-1. They designated LESTR to CXC chemokine receptor-4 (CXCR4) (26).
CXCR4 is the only one receptor for SDF-1. This concept implicates that the expression of CXCR4 is critical for the biological functions of SDF-1.
CXCR4 coding region contains 8724 base pairs (bps) and there are two exons separated by an intron sequence (27). Human CXCR4 gene is located on chromosome 2q21. An alternative splicing form of CXCR4 has been reported, termed CXCR4-Lo. The mRNA transcript is about 4.0 kb and differs from the known CXCR4 by the first 9 amino acids in the NH2-terminal extracellular domain of the receptor (28).
CXCR4 is a receptor on T-cell surface for HIV-1 entry. It is also ubiquitously expressed on various cell types including monocyte, B cells, hematopoietic progenitor cells, thymus, brain, spleen, stomach, small intestine, endothelial cells, lung, liver, bone marrow, lymph node, neuron, astrocytes and microglia (29, 30).
Specific Aim
In head and neck squamous cell carcinoma, CXCR4 has been suggested to be involved in metastatic processes by increasing the active matrix metalloproteinase (MMP)-9 secretion (31). SDF-1/CXCR4 system has also been suggested to induce epithelial-mesenchymal transition (EMT) of oral cancer cells and might be involved in the lymph node metastasis (32). T.
Ishikawa et al. reported a highly significant correlation between CXCR4 expression and lymph node metastasis (31). CXCR4 was also detected in squamous cell carcinoma of the tongue, the expression of CXCR4 in metastatic tumor cells of lymph node even more stronger than in cells of primary tumor (37).
In Western and Japanese populations, the main accepted risk factors for OSCC are tobacco smoking and alcohol consumption (65). Nevertheless, chewing betel quid is the leading cause of OSCC in Taiwanese. Because of the etiological and ethnic differences, we are eager to investigate the role of SDF-1/CXCR4 axis in human oral cancer in Taiwanese. Besides analyzing cell migration, we also explored the proliferation induction effects through SDF-1/CXCR4 signaling in OSCC.
Chapter II. Reference Investigation
SDF-1/CXCR4 in normal cell
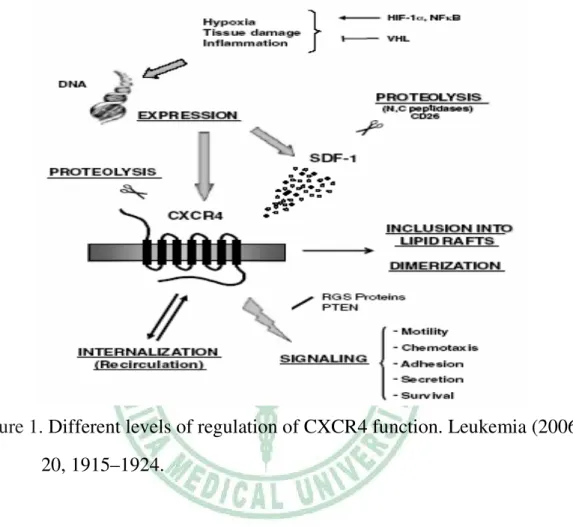
Figure 1. Different levels of regulation of CXCR4 function. Leukemia (2006) 20, 1915–1924.
SDF-1/CXCR4 axis plays an important role in cell development process.
Cell development is regulated by many complex events and overlapping pathways involving growth factors, cytokines and chemokines. When one of the factors defected, the large extent compensate usually started by other motomorphorgens. However, knockout SDF-1/CXCR4 in mice resulted in a significant defect in the hematopoietic stem cells homing to bone marrow as well as developing of brain, heart and large vessels (50). CXCR4 expression is a necessary condition for germ cells migration to another organ during
development. Previous study reported that AMD3100, a CXCR4 antagonist, can affect the rapid mobilization of hematopoietic stem cells, supporting that SDF-1 is essential for hematopoietic stem cell (HSC) retention within the bone marrow (51).
Two sites of hematopoietic activity, the yolk sac and aorta–gonad–mesonephros (AGM) region, function in mouse ontogeny at the pre-liver stage of hematopoiesis (52). During early embryogenesis, primordial germ stem cells (PGC) migrate to AGM and differentiate into haematopoietic stem cells. These PGC are CXCR4-expression and respond toward to an SDF-1 gradient. But hematopoietic stem cells in CXCR4-/- or SDF-1-/- mice still migrate properly from the yolk sac and AGM region to the fetal liver.
This event suggests that there are another chemoattractants are responsible for migration of these cells during this time of development (50). By the end of gestation in fetal liver, hematopoietic stem cells leave and colonization bone marrow tissue. SDF-1 is up-regulated in bone marrow and chemoattracts hematopoietic stem cells. Colonization of bone marrow depends on proper function of SDF-1/CXCR4 (10).
Except the role in cell development, tissue repair and regeneration are other important functions of SDF-1/CXCR4 axis. Hypoxia-inducible factor-1 (HIF-1) is produced by ischemic tissues can induce the expression of SDF-1 (20). SDF-1 expression can attract circulating stem cells and progenitor cells to the damaged tissue area. Briefly, hypoxia induces CXCR4-expressing progenitor cells recruitment for tissue repair. HIF-1 also enhances CXCR4 expression in normal and malignant cells. To regulate oxygen tension, von Hippel-Lindau tumor suppressor protein (pVHL) induces degradation of HIF-1. VHL gene mutations in renal cell carcinoma (RCC) increases CXCR4
expression by HIF-1 accumulation.
SDF-1/CXCR4 signaling pathway
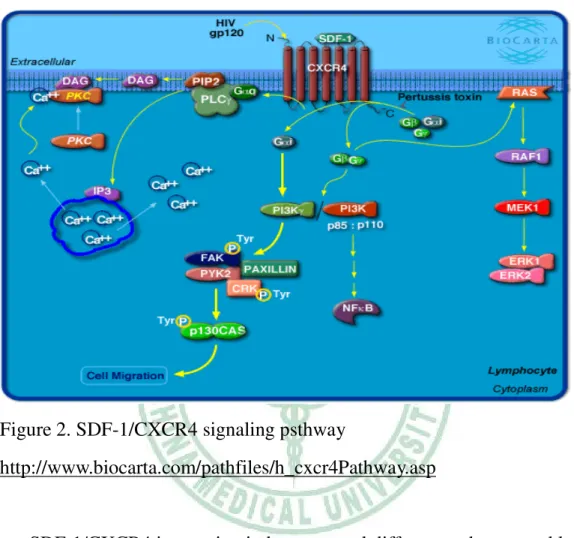
Figure 2. SDF-1/CXCR4 signaling psthway
http://www.biocarta.com/pathfiles/h_cxcr4Pathway.asp
SDF-1/CXCR4 interaction induces several different pathways and leads to act plural functions. CXCR4 binding to SDF-1 activates G-protein mediated signaling, including downstream pathways such as Ras, PLC and PI3 kinase.
PI3 kinase activated by SDF-1 and CXCR4 plays a role in lymphocyte chemotaxis in response to these signals. One endpoint of CXCR4 signaling is the activation of transcription factors such as AP-1 and chemokine regulated genes. JAK/STAT signaling pathway also plays a role in SDF-1/CXCR4 signaling. ERK 1/2 kinases are important mediators that activate the downstream signaling to the nucleus resulting in cell proliferation and
differentiation. Porcile, C. et al. reported that basal ERK 1/2 activity increased after SDF-1 stimulation in ovarian cancer cell lines (33).
SDF-1/CXCR4 associated malignancy
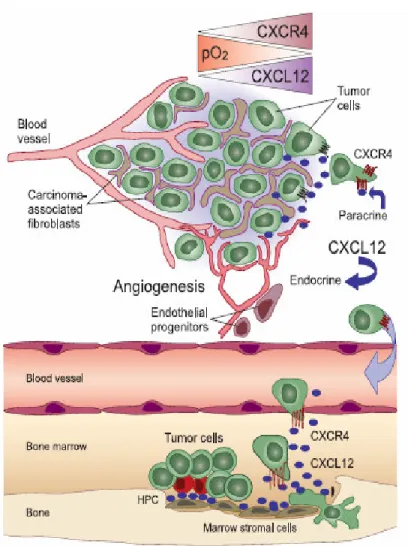
Figure 3. Importance of the CXCR4 chemokine receptor and its ligand, CXCL12, in the tumor microenvironment and for targeted metastasis. Blood.
2006 Mar 1;107 (5)
In addition to function in normal cells, SDF-1/CXCR4 axis is also suggested to play crucial rules in malignant cells. Many studies demonstrated that this ligand/receptor pair acts multiple effects as proliferation, migration,
invasion, and angiogenesis in cancers to facilitate tumor progression.
Cell proliferation is a crucial step involved in early stage of tumorigenesis.
Tumor cells elevate the proliferation rate by decreasing apoptosis or promoting cell cycles attribute to lose their auto-regulatory functions. Many reports demonstrated that SDF-1 increased cell proliferation to promote cancer progression in many kinds of cancers. Marchesi. F. P. et al. reported that addition of SDF-1 increased pancreatic cancer cells proliferation (53).
The same role induced by SDF-1 also found in ovarian cancer, breast cancer, brain cancer, prostate cancer and kidney cancer cell lines (9, 47, 13, 46, 53, 54, 64). Porcile, C. et al. observed that OC314RNAi , a clone from ovarian cell in which the expression of CXCR4 was selectively reduced by RNA interfering technique, was insensitive to the SDF-1-induced cell proliferation (33).
Kang, H. et al. reported that SDF-1 can increase the migration and invasion in breast cancer cell lines. Furthermore, high SDF-1 transcript level from breast cancer tissue was associated with reducing incidence-free survival (34). Under the normal situation, SDF-1 regulates leukocyte trafficking by mediate leukocyte adhesion to endothelium. In malignance cells, this function induced by SDF-1 could lead to transendothelium migration and tissue invasion. In human pancreatic cancer, Mori, T. et al. showed that CXCR4 antagonist inhibited stromal cell-derived factor 1-induced migration and invasion (35). Other cancer cell lines were also found that migration and invasion abilities were increased by SDF-1 such as brain cancer, breast cancer, cervical cancer, ovarian cancer, pancreatic cancer and prostatic cancer (35, 40, 53, 54, 64)
SDF-1/CXCR4 axis inhibitors
There are several SDF-1/CXCR4 axis inhibitors have been reported in many studies. Briefly introduce three major inhibitors: T22, AXL-4C, and AMD3100.
First, CXCR4 antagonist, T22 (Tyr5,12, Lys7]-polyphemusin II) , which is a 18 residue oligopeptide, is used for treatment of T-cell tropic HIV (55). The major disulfide ring of T22 is indispensable for anti-HIV activity and maintenance of the secondary structure. The antiparallel β sheet structure similar to CXC chemokines and provide the inhibition of CXCR4.
Second, AXL-4C is a highly cationic oligopeptide containing 9 arginine.
The peptides had similar activities to SDF-1 but were less potent. It was originally developed to mimic the basic domain of the HIV transactivation protein to inhibit replication (56).
Third, AMD3100 was originally described as a highly potent and selective inhibitor of HIV-1 and HIV-2 replication. It is a heterocyclic bicyclam compound and inhibits binding of SDF-1 and 12G5 to CXCR4 (57).
Bicyclams derivatives composed of two cyclam
(1,4,8,11-tetraazacyclotetradecane) units linked by an aliphatic or aromatic linker. There are two macrocyclic rings are connected by a phenylenebis (methylene) linker (62). In the beginning, the molecular mechanism of action of AMD3100 was believed to be related directly to the viral gp120 protein of HIV (63). Schols D. et al. demonstrated that AMD3100 inhibits HIV fusion and entry into cell membrane by binding to the CXCR4 chemokine receptor, which is the main coreceptor for gp120 used by X4, T-tropic strains of HIV (57). On the other hand, AMD3100 has been demonstrated effectiveness in hematopoietic stem cells mobilization and also inhibits tumor cells growth by
increasing apoptosis and decreasing the proliferation of glioblastoma and medulloblastoma xenografts (58). AMD3100 is successfully used to mobilize HSC from bone marrow into peripheral blood in healthy volunteers by block CXCR4 receptor (59).AMD3100 was performed a phase I study and appears to be a safe and effective agent in multiple myeloma (MM) and non-Hodgkin's lymphoma (NHL) patients who have received prior chemotherapy (59). In phase II trials, combination of AMD3100 and granulocyte colony-stimulating factor (G-CSF) can collect more progenitor cells than G-CSF alone used for autologous and allogeneic stem cell transplantation (61).
Figure 4. Chemical structure of the bicyclam AMD3100,
1,10-[1,4-phenylene-bis(methylene)]-bis-1,4,8,11- azatetradecane (62).
Chapter III. Materials and Methods
Materials
1. Trypsin-EDTA, 0.25% Hyclone 2. Dulbecco’s modified eagle’s medium/nutrient mixture F-12 ham’s (DME/F-12 1:1) Gibco 3. Hydrogen peroxidase , 30% Merck 4. Hematoxylin Merck 5. Citric acid monohydrate Merck 6. Xylene Merck 7. Entellan Merck 8. Silane coating slide
9. NaOH Merck 10. 24-well transwell Corning 11. Bovin serum albumin (BSA) USB 12. Recombinant human CXCL12/SDF-1α (Cat. 350-NS) R&D 13. 12-well plate Orange Scientific 14. Matrigel basement membrane matrix BD 15. Diethyl pyrocarbonate (DEPC) Sigma 16. Formaldehyde solution Qualikems 17. Sodium acetate USB 18. Agarose GIBCOBRL 19. β-mercaptoethanol Merck
20. Ethanol Merck 21. Klen Tag DNA polymerase Protech 22. Ethidium bromide solution Electran 23. Polyoxyethylene sorbitan monolanrate (Tween 20) USB 24. Antifoam A emulsion Sigma 25. Orange G Sigma 26. TEMED Plusone 27. GeneRuler 100bp DNA ladder Fermentas 28. Tris(hydroxymethly)-aminomethane, (Tris) Merck 29. Sodium dodecyl sulfate, (SDS) Merck 30. Acrylamide/bis-acrylamide, 40% solution Sigma 31. Ammonium persulfate, (APS) Merck 32. Glycine Merck 33. Sodium Chloride , (NaCl) Merck 34. non-fat dry milk Ancher 35. Biomax flim Kodak 36. developer Kodak 37. fixer Kodak
38. Polyvinylidene fluoride membrane,(PVDF) member Mersham Life Science
39. Methanol Merck 40. Bradford reagent Sigma 41. PageRuler prestained protein ladder, #SM0679 Fermentas 42. Sodium hydrogen carbonate, (NaHCO3) Merck 43. L-Glutamine Hyclone 44. Penicilline/ streptomycin Hyclone
45. Potassium Chloride, (KCl) Merck 46. Disodium hydrogen phosphate, (Na2HPO4.2H2O2) Merck 47. Potassium dihydrogen phosphate, (KH2PO4) Merck 48. Dimethyl sulfoxide, (DMSO) Sigma 49. Commassie blue Sigma 50. Trypan blue Sigma 51. Cosmic calf serum Hyclone 52. 15 ml tube Nunc 53. Hemacytometer American optical 54. Hypercassette Mersnam life science
Antibodies
1. Anti- Rabbit IgG (Goat), HRP-labeled PerkinElmer 2. Anti- Mouse IgG (Goat), HRP-labeled PerkinElmer 3. Rabbit monoclonal to CCR7 Abcam 4. Monoclonal anti-human/mouse CXCL12/SDF-1 antibody (Cat. MAB350) R&D 5. Mouse anti-actin monoclonal antibody (Cat. MAB1501) Chemicon 6. Rabbit polyclonal to CXCR4 (Cat. Ab2047) Abcam 7. Fusin (H-118) (Cat. Sc-9046) Santa Cruz
Kits
1. IHC select kit, immunoperoxidase secondary detection system Chemicon 2. SuperScript first-strand synthesis system for RT-PCR Invitrogen 3. RNeasy mini kit Qiagen
4. QIA shredder Qiagen 5. Enhanced chemiluminescent kit, (ECL) PerkinElmer
Equipments
1. Upright microscope, BX50 Olympus 2. Inverted microscope, CK2 Olympus 3. Centrifuge, 5100 Kubota 4. Water-jacketed incubater Forma Scientific 5. High-speed refrigerated centrifuge, CR15D Hitachi 6. UV/vis spectrophotometer, Du-650 Beckman 7. Gel imaging system, Eagle eye II Stratagen 8. Electrophoresis power supply, EPS601 Amersham Biosciences 9. Mupid-2 mini-gel electrophoresis system Mupid-2, Japan 10. Lamonar flow Fume Hood, TW 11. Water bath, WB211 Kansin, TW 12. Microwave oven Simpo, TW 13. Stirrer/ hot plate Coring 14. Hoefer semiphor Hoefer pharmacia
Methods
Tissue samples
Tissue samples from patients with OSCC were obtained from China Medical University Hospital in Taiwan. Each specimen was confirmed by histopathologic diagnosis.
Immunohistichemical staining
Formalin-fixed and paraffin-embedded specimens were obtained from surgical biopsies. Before staining, 4 µm-thick sections that were mounted on Silane coating slide. The slides were deparaffinized in xylene and rehydrated in graded ethanol. For the purpose of antigen retrieval, the sections were incubated at 95℃ for 20 minutes. These sections then incubated with 3% , H2O2 for 10 minutes, followed by an blocking step for 5 minutes (DAB150-150 slide kit, Chemicon international Inc.). Sections were incubated with polyclonal Ab against human CXCR4 (abcam) at a dilution of 1 in 50 or monoclonal anti-human/mouse SDF-1 antibody (R&D) at a dilution of 1 in 65 for 30 minutes. For the subsequent steps, the avidin-biotin-peroxidase method was used according to manufacturer’s instruction. The sections were finally counterstained with hematoxylin and mounted.
Cell culture
The following ATCC oral cancer cell lines were used: CA922, Cal27, SAS, HSC3 and SCC4 . TW2.6 oral cancer line was established by S.H. Kok et al.
in Taiwan (School of Dentistry, College of Medicine, National Taiwan
University). Human cervical cancer cell line HeLa was used as a CXCR4-expressing control cells (53). Human breast cancer cell line MCF-7 was used as a SDF-1(+) control cells. Cells were cultured at 37℃ in a humidified atmosphere of 5 % CO2. TW2.6, HSC3 and SCC4 were cultured in Dulbecco’s modified eagle’s medium/nutrient mixture F-12 ham’s (DME/F-12 1:1) culture medium, the other cell lines were in Dulbecco’s modified eagle’s medium/ high glucose (DMEM) culture medium. Medium were all supplemented with 10 % fetal calf serum, 100 U/ml penicillin-streptomycin, and 2 mM L-glutamine.
RNA Extraction
OSCC cells grown in a monolayer (1 x 107) were harvested by typsinization.
Cell lysate underwent centrifugation at 1000 rpm for 5 mins in RNase-free centrifuge tubes. Then the supernatant was completely aspirated. After adding 600 µl RLT buffer (RNeasy mini kit, Qiagen) the lysate was transferred into QIAshredder spin column (QIAshredder , Qiagen) and was centrifuged for 2 mins at 14000 rpm. Then 700 µl 70% ethanol was added to homogenized lysate, and mix well by pipetting. Transferred 700 µl lysate to spin column (RNeasy mini kit, Qiagen) and centrifuged for 30 s at 10000 rpm.
Flow-through was discarded. Added 700 µl RWl buffer (RNeasy mini kit, Qiagen) to the column and centrifuged for 30 s at 10000 rpm. After discarding the flow-through, 500 µl RPE buffer (RNeasy mini kit, Qiagen) was added to the column and was centrifuged for 2 mins at 10000 rpm. The column was placed in a new 2 ml collection tube and then centrifuged for 75 s at 14000 rpm. The column was placed in a new 1.5 ml tube and added 50 µl
DEPC-water directly to the spin column membrane, centrifuged for 75 s at 10000 rpm.
Reverse transcription-PCR
For RT-PCR, 2 µg of total RNA, 1 µl dNTP mix (10 mM), 1 µl oligo dT (0.5 µg/µl) and DEPC-treated water were added in a total volume of 10 µl. After 65℃ for 5 mins, each sample was incubated at 4℃ for 1 min. 2 µl of RT buffer (10 x), 4 µl MgCl2 (25 mM), 2 µl DTT (0.1 M) and 1 µl RNaseOUT recombinant RNase inhibitor (40 units/µl) were added into a reaction mixture and incubated at 42℃ for 2 mins. Then 1 µl (50 units) of SuperScript II reverse transcriptase was added to each sample, and incubated at 42℃ for 50 min. The reaction was terminated with heating at 70℃ for 15 mins. One µl of RNase H (2 units/µl) was added to each sample and incubated for 20 mins at 37℃. The resulting cDNA was subjected to a 50 µl PCR reaction solution containing 5 µl PCR buffer (10x), 1.5 µl MgCl2 (50 mM), 1µl dNTP mix (10 mM), 1 µl sense primer (10 µM), 1 µl anti-sense primer (10 µM), 0.4 µl DNA polymerase (5 units/µl). The following thermal cycle condition was used for β-actin and CXCR4: 94℃ (2 mins), 36 cycles at 94℃ (30 sec), 58℃ (30 sec), 72℃ (1 min), and 72℃ (10 mins). The following thermal cycle condition was used for SDF-1: 94℃ (2 mins), 36 cycles at 94℃ (30 sec), 58℃ (30 sec), 72℃ (1 min): and 72℃ (10 mins). The PCR products were electrophoresised in a 3% agarose gel containing ethidium bromide. The expected sizes of PCR products for β-actin, CXCR4 and SDF-1 are 637 bps, 457 bps and 512 bps, respectively.
Target Sense Primer Antisense Primer
PCR product (base pair) ATCCGCAAAGACCTGTACGC
β-actin
TGTGTGGACTTGGGAGAGGA
637
AATCTTCCTGCCCACCATCT CXCR4
GACGCCAACATAGACCACCT
457
GGGGGAATTCCATGAACGCCAAGGTCGTGGTC SDF-1
GGGGTCTAGAGGGCATGGATGAATATAAGCTGC 512
Table 1. Primer sequences and the length of each PCR products.
Western blot
OSCC cells were cultured to 90% confluence at 37℃ with 5% CO2 . Cell pellet underwent centrifugation at 2000 rpm for 5 mins at 4℃ then the supernatant was discarded. Cell lysate was prepared by adding distilled water and SDS-containing sample buffer. The total protein was quantified with Bradford reagent. 40 µg of protein for each sample was loaded and fractionated by 10% SDS-PAGE. After electrophoresis, the fractionated proteins were transferred onto a PVDF membrane, then the membrane was incubated with mouse monoclonal Ab against β-actin (Chemicon) at 1:10000 dilution or rabbit polyclonal Ab against CXCR4 (Abcam) at 1:1000 dilution for 1 h at room temperature. The specific protein was probed with anti-mouse IgG (PerkinElmer) at 1:10000 dilution or anti-rabbit IgG (PerkinElmer) at 1:3000 dilution for 30 mins at room temperature. Detection was then performed using an enhanced chemiluminescence kit (PerkinElmer).
Proliferation assay
5 x 104 cells for each OSCC cell line were planted in wells of 12-well plate with 1000 µl serum-free medium. After 24 hours incubation under serum-starvation, cells were stimulated with 200 ng/ml SDF-1. After 3, 4, 5, or 6 days of SDF-1 treatment, Cells were harvested by trypsinization and counted by using trypan blue staining and hemacytometer.
Proliferation Index= the cell number in the presence SDF-1 treatment/ the cell number in the absence SDF-1 treatment
Migration assay
The migration potential was evaluated using transwell chamber (24-well, Corning). The upper chamber was separated by an 8-µm pore-size filter.
Serum-free medium containing 4x104 cells was added to the upper chamber.
The lower chamber was loaded with serum-free medium in the absence or presence 100 ng/ml of SDF-1 (R&D). Chambers were incubated for 12 h or 24 h in a humidified atmosphere with 5 % CO2 at 37℃. After 10%
formaldehyde fixation for 10 mins, cells on the upper surface of the filter were removed using cotton swabs. Migrated cells on the lower surface were stained using commassie blue solution for 30 minutes. The number of migrated cells was counted by upright microscopy.
Migration Index= the number of migrated cells in the presence SDF-1 treatment/ the number of migrated cells in the absence SDF-1 treatment
Invasion assay
The invasion potential was also evaluated using transwell chamber (24-well, Corning). Membrane was precoated with 100 µl per inert of matrigel (BD) diluted 1:1 with serum-free medium. Serum-free medium (0.1 ml) containing 2x104 cells was added to the upper chamber. 0.5 ml serum-free medium was loaded into the lower chamber in the absence or presence of 100 ng/ml SDF-1 (R&D). Chambers were incubated for 12 h or 24 h in a humidified atmosphere with 5 % CO2 at 37℃. After 10% formaldehyde fixation for 10 minutes, cells on the upper surface of the filter were removed using cotton swabs. Migrated cells on the lower surface of the filter were stained using commassie blue solution for 30 mins. The numbers of migrated cells were counted by upright microscopy.
Chapter IV. Results
Immunohistochemical staining of CXCR4 and SDF-1 in OSCC tissues
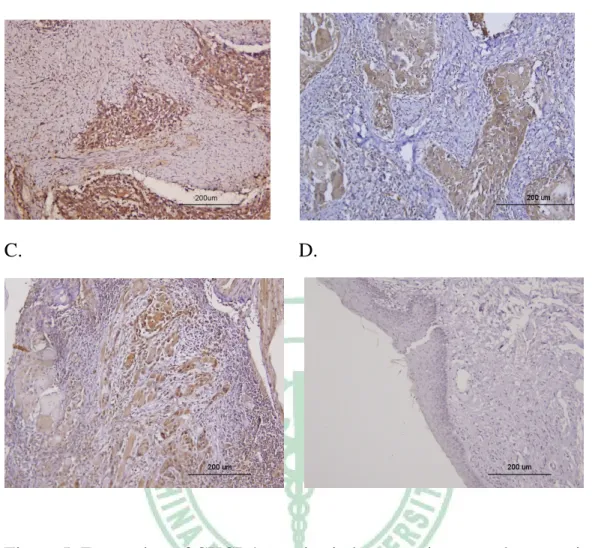
To investigate whether CXCR4 and SDF-1 correlated with OSCC, we did immunohistochemical staining to detect proteins in clinical oral cancer tissue.
The CXCR4 protein was not detected on normal epithelial cell and inflammatory cells (Fig. 5B, 5D), it was mainly expressed in cancer cells (Fig.
5A). Although some cases of OSCC tissues were SDF-1 negative (Fig. 6D), but SDF-1 was expressed in cancer cells in most samples (Fig. 6B, 6C, 6E).
In some cases, the expression of SDF-1 was also found in the inflammatory cells infiltrated into the tumor mass (Fig. 6F). A special case demonstrated that SDF-1 was expressed in malignance region but not in normal epithelia region, and the expression of SDF-1 was appeared from hyperplasia cells (Fig.
7). We also found that the expression of SDF-1 in well-differentiated cancer cells was stronger than in poorly-differentiated cancer cells.
CXCR4 and SDF-1 expression in oral cancer cell lines
To investigate the expression level of CXCR4 and SDF-1 in six oral cancer cell lines, we evaluated the mRNA and protein level by reverse-transcription PCR and Western blot. CXCR4 mRNA was identified in all six oral cancer cell lines (Fig. 9B). The presences of CXCR4 protein in six oral cancer cell lines were also confirmed by Western blot (Fig. 8B). The mRNA expression of SDF-1 was only found in TW2.6 and positive control cells MCF7 (Fig. 9D).
SDF-1 mRNA was not detected in other five oral cancer cell lines (CA922, Cal27, SAS, HSC3 and SCC4).
SDF-1 increases proliferation of oral cancer cell lines
Since we observed that SDF-1 and CXCR4 proteins expressed in OSCC tissues and cell lines, we would like to investigate what roles does this ligand/receptor pair play in OSCC. First, cell proliferation assay was performed to investigate if SDF-1 mediated proliferation of oral cancer cell lines. To determine the optimal induction effect, different SDF-1 doses were used to treat TW2.6 cells. The cell number of TW2.6 treated with or without SDF-1 were counted after 6 days of treatment. Maximal proliferation effect was observed at the concentration of 200 ng/ml SDF-1 (Fig. 10A). Then we did the time-dependent proliferation assay on TW2.6 and HSC3 cell lines.
After stimulation with 200 ng/ml SDF-1, we counted the cell number at 3, 4, 5, and 6 days, respectively (Fig. 10B, 10D). The data showed that cell numbers increased after SDF-1 stimulation in both TW2.6 and HSC cells (Fig. 10C, 10D).
SDF-1 stimulates oral cancer cell migration
To furthermore determine if SDF-1 promote metastasis of oral squamous cell carcinoma, we evaluated cell migration activities after 100 ng/ml SDF-1 stimulation. To investigate the SDF-1-mediated migration of oral cancer cells, the migration assay was performed. First, we tested the optimal culture period for cell migration. TW2.6 cells were incubated in serum-free medium in the absence or presence of 100 ng/ml SDF-1. The migrated cells were counted at 12 h, 24 h, 48 h, and 72 h. After 48 h or 72 h of incubation, migrated cells started to aggregate together. The optimal culture period for cell migration is 12 h or 24 h (Fig 11A). The migration indexes of CA922, TW2.6, HSC3 and SCC4 at 12 h were 1.1, 1.9, 2.26 and 1.8 fold, respectively (Fig. 11D); at 24 h were 1.6, 1.6, 2.5 and 1.7, respectively (Fig. 11E). OSCC cell lines did migrate in response to SDF-1 in a classical chemotaxis assay.
SDF-1 has the potential to promote invasion of oral cancer cells through matrigel.
To investigate whether SDF-1 increases the invasion activity of OSCC cells, the effect of SDF-1-mediate invasion assay was tested. TW2.6 and HSC3 cells were incubated in serum-free medium in the absence or presence of 100 ng/ml SDF-1 and the cell numbers were counted at 12 h and 24 h (Fig. 12C, 12D).
OSCC cells that treated with SDF-1 exhibited greater invasion ability through matrigel than cells not treated. SDF-1 indeed promoted the invasion ability in OSCC cells.
Chapter V. Discussion
Recent studies demonstrated that CXCR4 was highly expressed in malignant breast cancer cells and metastases (40). Hu, J. et al. reported that CXCR4 protein was detected in cancer cells of primary tumor and metastatic cancer cells in lymph node of nasopharyngeal carcinoma (41). Consistently, we also observed that CXCR4 was mainly expressed in oral squamous cell carcinoma but not in normal oral epithelia. Through immunohistochemical staining, we also observed that SDF-1 protein was expressed in cancer cells, infiltrated leukocytes and stroma cells. These results implicate that SDF-1 proteins are secreted into the tumor microenvironment by tumor-infiltrating stroma cells as well as by tumor cells. We suggest the idea that SDF-1 can act an autocrine and paracrine manners in the tumorigenesis of oral cancer.
Although H. Kang et al. did not observed SDF-1 staining in vascular endothelia of breast cancer tissue, but our results consent to most previous studies (13)-SDF-1 was presented in vascular endothelial cells. Endothelia express SDF-1 might attract the CXCR4-expressing cancer cells migrate close to blood vessels then entry into blood fluid, and improve metastasis to distant organs.
Although SDF-1 protein was not detected in OSCC cell lines by Western blot analysis (data not shown). But through RT-PCR, TW2.6 apparently expressed SDF-1 mRNA. This ligand/receptor pair may play important roles in the tumorigenesis of OSCC.
Interestingly, we also observed that SDF-1 expression in well-differentiated OSCC tissues was stronger than in poorly-differentiated tissues. We suggest
mutated cells secreted SDF-1 proteins induce proliferation in the beginning of tumorigenesis. Accompany progression of malignance, the primary cancer cells stop expressing SDF-1 and are attracted by distance organs that secretes SDF-1 proteins. Previous studies showed loss of SDF-1 mRNA expression in malignant tissues such as colon, esophageal and gastric cancers (42). In another thought, SDF-1 was detected in OSCC tissues but not in cultured cancer cells. We suggest this different event could be made by the environment that cancer cells or stroma cells surrounding tumor nest might secret some soluble factors that can induce the expression of SDF-1.
In this study, we showed expression of both CXCR4 mRNA and protein in six oral cancer cell lines. Exogenous SDF-1 promoted proliferation, migration, and invasion of these CXCR4-beared cells. These results suggest that these oral cancer cell lines have CXCR4, which is the specific receptor for SDF-1, plays a possible role in cell proliferation, migration and invasion of oral cancer.
Some studies reported that the role of CXCR4 in cell proliferation is controversial. In 2003, Phillips et al. found that SDF-1 can not promote cell proliferation in non-small cell lung cancer lines. Hwang et al. also reported that SDF-1 didn’t increase proliferation in anaplastic thyroid cancer cell lines.
In contradiction, Porcile, C. et al. showed that SDF-1 can stimulate ovarian cancer cell lines proliferation (33). We suggest the differences of SDF-1-mediate proliferation may be dependent on the cell type or the different signal transduction pathway.
In several cancers, CXCR4 was implicated in tumor metastasis to distant organs. Mori, T. et al. reported that CXCR4 antagonist inhibited SDF-1-induced migration and invasion of pancreatic cell lines (35),
suggesting that the SDF-1-induced migration and invasion was via CXCR4 receptor. Scotton, C.J et al. in 2002, reported SDF-1 can also stimulate both migration and invasion in CXCR4-expressing ovarian cancer cells (64).
Through the transwell chamber, we found SDF-1 can increase the abilities of migration and invasion in oral cancer cell lines. Furthermore, in 2005, Kang, H. et al. reported neutralizing of SDF-1/CXCR4 interaction significantly impaired metastasis to lymph node and lung in mice (34). Taken together, these findings indicate that cancer cells express CXCR4 receptor may induce metastasis toward the organs which secrete its ligand SDF-1. It may explain why oral cancer metastasis is frequently found in lymph node, bone marrow, lung and liver , where SDF-1 is expressed.
In conclusion, our results imply that the SDF-1/CXCR4 axis play important and multiple roles in the tumorigenesis process of OSCC. Developing the antagonists of SDF-1/CXCR4 might provide the opportunities to inhibit tumor growth, distant organ metastasis and even as therapeutic courses for OSCC patients.
A. B.
C. D.
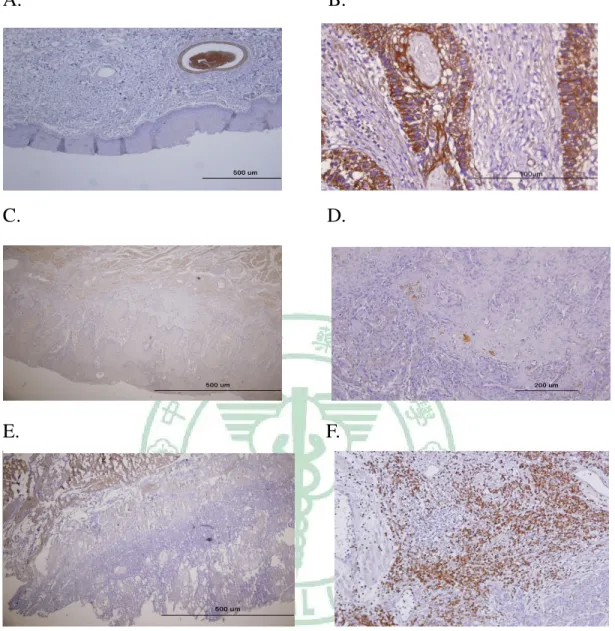
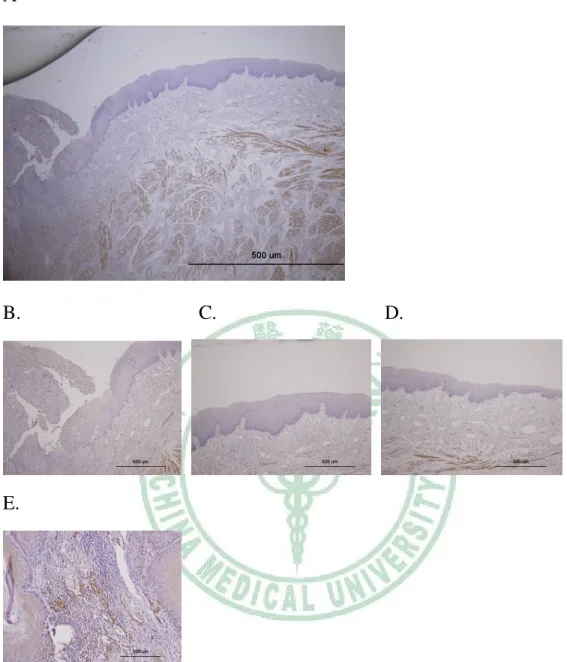
Figure 5. Expression of CXCR4 proteins in human primary oral cancer tissue A, Strong CXCR4 expression was observed in tumor cells. B and C, CXCR4 was expressed in cancer cells but not expressed in infiltrating inflammatory cells. D, Normal epithelia did not express CXCR4.
A. B.
C. D.
E. F.
Figure 6. Expression of SDF-1 proteins in human primary oral cancer tissues.
A, SDF-1 was not expressed in normal oral epithelia. Endothelial cells are positive staining. B, SDF-1 is highly expressed in OSCC cancer cells. C, Tumor cells expressed faintly but extended SDF-1 proteins in some cases. D, Some cases showed strong but sparse staining of SDF-1 proteins in OSCC tissues. E, SDF-1 was not identified in cancer tissue except smooth muscle cells. F, infiltrating inflammatory cells expressed high level of SDF-1 proteins, but cancer cells did not express any SDF-1 proteins in this case.
A
B. C. D.
E.
Figure 7. SDF-1 involved in the tumorigenesis of OSCC. A, In one special case, we observed the OSCC developing since normal epithelia in the right side progressed to malignant cells in the left view. B, Cancer cells expressed SDF-1. C, Hyperplasia epithelia weakly expressed SDF-1. D, SDF-1 was not detected in normal epithelia portion. E, SDF-1 expression was apparent in cancer cells
A.
B.
Figure 8. Western blot analysis of CXCR4 proteins in six OSCC cell lines. A, β-actin ( 42 kDa) was performed as an internal control. B, CXCR4 proteins ( 39 kDa) were expressed in all of the six OSCC cell lines.
CXCR4-presenting HeLa cells were used as a positive control.
A.
B.
C.
D.
Figure 9. RT-PCR analysis of the mRNA expression of CXCR4 and SDF-1 in six OSCC cell lines. A, β-actin transcripts were measured as an internal control. B, CXCR4 mRNA expressed in all of the six OSCC cell lines. C, β-actin transcripts were measured as internal control. D, SDF-1 mRNA only expressed in TW2.6 and positive control MCF-7 cell line.
A.
B. C.
D. E.
Figure 10. SDF-1 induces proliferation of TW2.6 and HSC3 cells. A, Optimal induction effect in TW2.6 cells was induced with 200 ng/ml SDF-1. OSCC cells were grown in serum-free medium in the absence or presence of 200 ng/ml SDF-1. The number of cells was counted after 3, 4, 5, 6 days of incubation periods. B, SDF-1 stimulation increases cell number in TW2.6 cell line. C, proliferation index was increasing of TW2.6 cell line when treatment period prolongs. D, SDF-1 stimulation resulted in increasing cell number in HSC3 cell line. E, proliferation induction index of HSC3 cell line.
TW2.6
ng/ml SDF-1
0 200 300
Cell Number
0 100000 200000 300000 400000
TW2.6
Days
2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5
Cell Number
0 100000 200000 300000 400000 500000 600000
0 ng/ml SDF-1 200 ng/ml SDF-1
TW2.6
Days
2 3 4 5 6 7
Proliferation Index (fold)
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
HSC3
Days
2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5
Cell Number
0 100000 200000 300000 400000 500000 600000 700000 800000
0 ng/ml SDF-1 200 ng/ml SDF-1
HSC3
Days
2 3 4 5 6 7
Proliferation Index (fold)
0.0 0.5 1.0 1.5 2.0 2.5
A.
TW206
Time
12H 24H 48H
Cell Number
100 150 200 250 300 350 400 450 500
0 ng/ml SDF-1
100 ng/ml SDF-1
B. C.
D. E.
Figure 11. SDF-1 induces cell migration in OSCC cells . The data show that the migrated cell number was increased after 100 ng/ml SDF-1 treatment. A, the optimal incubation time of migration for TW2.6 is 12 h and 24 h. B, the migrated TW2.6 cells after incubation with 100 ng/ml SDF-1 for 12 h (100x).
B, the migrated TW2.6 cells after incubated in the absence of SDF-1 for 12 h (100x). D, migration indexes of four OSCC cell lines after SDF-1 treatment for 12 h. E, migration indexes of four OSCC cell lines after SDF-1 treatment for 24 h.
Migration Index= the migrated cell number with SDF-1 treatment/
the migrated cell number without SDF-1 treatment
CA922 TW2.6 HSC3 SCC4
Migration Index (fold)
0.0 0.5 1.0 1.5 2.0 2.5
CA922 TW2.6 HSC3 SCC4
Migration Index (fold)
0.0 0.5 1.0 1.5 2.0 2.5 3.0
A. B.
C. D.
12H
TW2.6 HSC3
Cell Number
0 2 4 6 8 10 12
0 ng/ml SDF-1 100 ng/ml SDF-1
24H
TW2.6 HSC3
Cell Number
0 20 40 60 80 100 120 140 160
0 ng/ml SDF-1 100 ng/ml SDF-1
Figure 12. Invasion of TW2.6 and HSC3 through matrigel after SDF-1
stimulation. A, The migrated TW2.6 cells with 100 ng/ml SDF-1 treatment for 24 h (100x). B, The migrated TW2.6 cells without SDF-1 stimulation (100x).
C, The migrated cell number of TW2.6 and HSC3 after SDF-1 treatment for 12 h. D, The migrated cell number of TW2.6 and HSC3 after SDF-1 treatment for 24 h.