行政院國家科學委員會補助專題研究計畫
行政院國家科學委員會補助專題研究計畫
行政院國家科學委員會補助專題研究計畫
行政院國家科學委員會補助專題研究計畫 ■ ■ ■ ■ 成 果 報 告 成 果 報 告 成 果 報 告 成 果 報 告
□ □
□ □期中進度報告 期中進度報告 期中進度報告 期中進度報告
蘭科植物B群花部發育基因之研究
計畫類別:■ 個別型計畫 □ 整合型計畫 計畫編號:98-2311-B-006-003-MY2
執行期間: 2009 年 8 月 1 日至 2011 年 7 月 31 日
計畫主持人:陳虹樺 計畫參與人員:潘昭君
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列 管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位:國立成功大學生命科學系
中 華 民 國 100 年 5 月 31 日
目錄目錄 目錄目錄
目錄---Ⅱ 中文摘要---Ⅲ 英文摘要---Ⅳ
前言、文獻探討/ introduction---1
研究目的/propose---2
研究方法/material and method---3
結果/result---5
討論/discussion---9
參考文獻/reference---12
附表及附圖/table and figure---15
附錄---28
附件 supporting text---29
中文摘要 中文摘要 中文摘要 中文摘要
蘭花的數量約佔被子植物近 10%,其物種數量為開花植物最大的科之ㄧ。蘭花 與其花媒者以共同演化關係緊密相連,因此大多數蘭花物種都具有高度特化的唇 瓣。蘭花以合蕊柱和唇瓣為最重要的花形特徵,其中唇瓣為中間花瓣所特化而成。
蘭科植物的分類共分為五個亞科,含擬蘭亞科、拖鞋蘭亞科、蘭亞科、梵尼蘭亞科 及樹蘭亞科。蝴蝶蘭屬於樹蘭亞科,目前是台灣主要花卉外銷產品之一,而且為因 應加入世界貿易組織對我國農業所造成的衝擊,蝴蝶蘭更被國家列為重點培育的精 緻農業之一。有關蘭花形發育的相關研究雖然在本實驗室已經以蝴蝶蘭為模式物 種,共提出五個姬蝴蝶蘭 B 群基因(PeMADS2-6),含四個 AP3-like 的基因(PeMADS2-5) 和一個 PI-like 的基因(PeMADS6)。其中四個 AP3-like 的 B 群基因會差異性地分別參 與不同花器的發育。由於一般單、雙子葉植物只含有一個 AP3-like 基因,而且目前 仍缺乏對於其他蘭花物種花形發育相關的研究,特別是較原始的擬蘭亞科和高度歧 異化的拖鞋蘭亞科,因此本計畫擬研究 B 群 MADS-box 基因在五個不同蘭亞科中的 基因是否皆具有四個 AP3-like 的基因,並進一步釐清 AP3-like 基因複製現象,探討 由於複製所產生的 B 群基因在不同花器扮演調控的功能是否相同,進而了解調控蘭 科植物花形發育的分子機制。
關鍵詞: B 群 MADS-box 基因、花部發育、花器決定、基因複製、蘭科
Abstract
Orchidaceae is an excellent model to examine the perianth development given its sophisticated floral architecture. In this study, 24 AP3-like and 13 PI-like genes were identified from 11 species of orchids and characterized into four AP3- and two PI-duplicated homologs. The first duplication event in AP3 homologs occurred in early evolutionary history of the Orchidaceae gave rise to AP3A and AP3B clades. Further duplication events resulted in four subclades, namely AP3A1, AP3A2, AP3B1, and AP3B2, during the evolution of Orchidaceae. The AP3 paralogous genes were expressed throughout inflorescence and floral bud development. The transcripts of PeMADS2, 4, 5 were widely detected at both developmental stages of floral primordia and floral organ primordia.
Distinct expression of these transcripts I specific floral organs were not observed until the late inflorescence and floral bud stages. In contrast, the transition point of distinguished expression of PeMADS3 (AP3A1 subclade) in lateral petal, lip, and column primordia was observed at late floral organ primordia stage. In addition, the AP3 orthologous genes carried out diverse expression patterns in various species of orchids. However, the PI homologs uniformly expressed in all floral whorls. AP3A2 orthologs play a noticeable role in lip formation. This was strongly supported by their sole expression in lip, and further evidence come from that their expression was extended from the lip to the lip-like petals in four sets of peloric mutants. Finally, a homeotic orchid perianth (HOT) model is proposed, in which dualistic characters of duplicated B-class MADS-box genes were involved in orchid perianth development and growth.
Keywords: B-class MADS-box, floral growth, floral organ identity, gene duplication, Orchidaceae
前言、文獻探討/introduction
Although the floral ABCDE model is generally thought to be ubiquitous in the core eudicots, studies on basal angiosperms, monocots, and basal eudicots have demonstrated great variations in the patterns of gene expression (Kramer and Hall 2005; Stellari et al.
2004; Zahn et al. 2005). A “sliding-boundary” model has been proposed to explain the development of petaloid sepals caused by an outward shift of gene expression (Kanno et al.
2003; Kramer and Jaramillo 2005). However, in some cases, variations in gene expression cannot be easily explained. For example, the expression of B-class genes extends to whorl 1 of the flowers in Asparagales species such as Agapanthus praceox (Nakamura et al. 2005), Muscari armeniacum (Nakada et al. 2006), and Phalaenopsis equestris (Tsai et al. 2004), whereas AOGLO (a B-class gene) is expressed only in whorl 2 in Asparagus that produces undifferentiated tepals (Park et al. 2004).
Orchidaceae is divided into five subfamilies based on the robust molecular evidence:
Apostasioideae, Vanilloideae, Cypripedioideae, Orchidoideae, and Epidendroideae (Cameron 2004). Because members of the Apostasioideae largely develop lily-like flowers with three anthers, but without distinct lips, it is sometimes considered to be a more primitive and separate subfamily. Orchids make up nearly 10% of angiosperms, and the high species diversity in orchids is largely due to their adaptation to specialized insect pollination (Dressler 1993b). The family Orchidaceae is characterized by a fused gynoecium and androecium (the gynostemium or the column). Besides, two whorls of the perianth are sometimes undistinguishable, although in most cases one of the inner three petals becomes a highly modified organ known as a labellum or lip. Lips offer a landing platform for pollinators or function in mimicry (Cozzolino and Widmer 2005; Dressler 1993a, b; Rudall and Bateman 2002).
The highly specialized and diverse forms of flowers in orchids have served as excellent models for examining the complicated network of regulatory genes in floral morphogenesis. Three types of perianth of orchid flowers have been discovered, including normal (non-peloria), semi-pseudopeloria, and pseudopeloria (Fig. 1) according to the similarities between the sepals and laterial petals in both natural occurring and by micropropagation. Although the perianth of orchid should be generally called tepal, sepal and petal can be easily distinguished in morphology in normal orchid perianth (Fig. 1A). A complete homeotic transformation and only a mild change can be observed in pseudopeloria (Fig. 1C) and semi-pseudopeloria (Fig. 1B), respectively. In pseudopeloric flower, sepals are transformed into organs that resemble lateral petals (Fig. 1C) that defined as Type D pseudopeloria previlously (Mondragon-Palomino and Theissen 2009). However, the perianth organs shown a mild transformation of sepal to lateral petal in a gradually changing level are found in the semi-pseudopeloric flower (Fig. 1B). The morphologies of sepal and petal are extremely similar, moderate differences in details of coloration, corrugation, and shape recognize these two floral organs. In orchids, a floral mutant in which the morphology of lateral petals with a keeled callus is similar to that of lip is referred to a peloric mutant (Cubas 2004; Rudall and Bateman 2002).
A set of biological processes, such as organ positioning, organ identity specification, cell proliferation, cell expansion, and cell differentiation and maturation, are coordinated by many regulators, including auxin, MADS, and TCP (Dornelas et al. 2010). Members of the B-class MADS-box genes are reported to involve in the continuously developmental program in orchids (Chang et al. 2010; Tsai et al. 2004; Tsai et al. 2005; Xu et al. 2006). So far, orchid B-class MADS-box genes have been characterized for P. equestris (Epidendroideae) (Tsai et al. 2004; Tsai et al. 2005), Dendrobium crumenatum (Epidendroideae) (Xu et al. 2006), Habenaria radiata (Orchioideae) (Kim et al. 2007) and
Oncidium Gower Ramsey (Chang et al. 2010). Members of MADS-box genes in P.
equestris participate in various processes related to organ development. A PI-like gene, PeMADS6, is ubiquitously expressed in petaloid sepals, petals, column, and ovary (Tsai et al. 2005). In contrast, four AP3-like genes, PeMADS2, PeMADS3, PeMADS4, and PeMADS5 exhibit different expression patterns, i.e. PeMADS4 expresses in lip of orchid perianth (Tsai et al. 2004). These differences provide the first indication that at least four AP3-like paralogs are involved in a single species of monocots (Tsai et al. 2004). One PI-like (DcOPI) and two AP3-like (DcOAP3A/B) homologs were identified in D.
crumenatum (Xu et al. 2006), and two PI-like (HrGLO1, HrGLO2), one AP3-like (HrDEF) homologs in H. radiata (Kim et al. 2007) and one PI-like (OMADS8) and three AP3-like (OMADS3, OMADS5, OMADS9) homologs were in O. Gower Ramsey (Chang et al. 2010;
Hsu and Yang 2002). Extended expression of HrDEF was detected in petaloid-sepals of the petaloid-sepal mutant, but not in wild-type sepals, indicating that ectopic expression of a B-class MADS-box gene can change the organ identity of whorl 1 into whorl 2 in orchid flowers (Kim et al. 2007). In flowers of the peloric mutant P. equestris, the appearance of the two lateral petals is converted into lip-like petals (Tsai et al. 2004). The changed expression of PeMADS4 in the lip-like petals of peloric mutants is always accompanied by the conversion of petals into lips within the same whorl in Phalaenopsis (Tsai et al. 2004).
Moreover, an ‘orchid code’ was proposed, theorizing the combinational interaction of four DEF-like genes for the organ identity of the different perianth organs (Mondragon-Palomino and Theissen 2008, 2009). It is reported that the AP3B2 orthologous gene in O. Gower Ramsey, OMADS3 plays a potential role in cell proliferation and repressing lip development (Chang et al. 2010). However, the function of these genes in orchid floral organ development can only be revealed by addressing their expression patterns at early stages of development by using in situ hybridization. Yet, the expression of B-class genes per se cannot be simply equated with their functions.
Previous studies have uncovered multiple paralogs and complex expression patterns of B-class genes in Phalaenopsis orchids, which lead us to wonder what the role for these genes really are. To assess whether the duplication of B-class genes detected in P. equestris is also true for all orchids for dictating their spectacular floral morphology, orthologs of B-class genes were identified and characterized in subfamilies of Orchidaceae. AP3-like and PI-like MADS-box genes from four subfamilies of Orchidaceae were identified and characterized. A total of 37 orchid B-class genes were isolated from 11 species collected from the field or from orchid nurseries and subjected to extensive phylogenetic analyses.
Four AP3 paralogs in orchids resulted from two runs of duplication events in the ancestral AP3-like genes. Strong purifying selection was detected in both B-class genes. A dualistic model of perianth development addressing differences in the expression patterns of the homeotic B-class genes may have influenced the identity and growth of the floral organs, which is required if evolution in orchids is to keep pace with that in their pollinators.
研究目的/purpose
Previous studies have uncovered multiple paralogs and complex expression patterns of B-class genes in Phalaenopsis orchids, which lead us to wonder what the role for these genes really are. To assess whether the duplication of B-class genes detected in P. equestris is also true for all orchids for dictating their spectacular floral morphology, orthologs of B-class genes were identified and characterized in subfamilies of Orchidaceae.
研究方法/material and method Plant materials
Eleven species of orchids were selected from four subfamilies of Orchidacea collected from orchid nurseries and from the wild for de novo identification of B-class MADS box genes: Vanilloideae, VA (Galeola falconeri, Vanilla pilifera Holttum); Cypripedioideae, CY (Paphiopedilum Macabre); Orchidoideae, OR (Anoectochilus formosanus, Ludisia discolor, and Habenaria petelotii); and Epidendroideae, EP (Oncidium Gower Ramsey, Liparis distans, Phaius tankervilliae, Brassavola nodosa, and Dendrobium Spring Jewel). Each of the species is shown in Figure S1 and voucher information is given in Supplementary Table S1.
Normal plants of Phalaenopsis equestris and its somaclonal variants of peloric mutant were analyzed for gene expression patterns among the five B-class MADS box genes.
Three pairs of wild-type and peloric mutant plants were included, namely a diploid wild-type P. equestris (S82-159), with red petals and an orange lip, and its peloric mutant (S85-355); a tetraploid P. equestris ‘King Car’ and its peloric mutant, and P. spp. ‘A3947’
and its peloric mutant (Fig. 8A-8F). In addition, one pair of normal plant of Cymbidium ensifolium and its peloric mutant were analyzed for AP3A2 ortholog expression (Fig. 8G and 8H). All the peloric mutant flowers show the phenotypic conversion of lateral petals into lip-like petals. Detailed information on plant collections and growth conditions is given in the supporting text. All plants were grown in a greenhouse at National Cheng Kung University (NCKU) under natural light (photosynthetic photon-flux density of 90 μmol m–2 s–1) and controlled temperature (23-27 °C).
Cloning and characterization of the orchid AP3-like and PI-like homologs
Total RNA was extracted as described earlier (Tsai et al. 2004). For the small samples of plant materials such as A. formosanus, L. discolor, and H. petelotii, RNA was extracted by using RNeasy Plant Mini Kit (QIAGEN, Stanford, CA). To identify B-class MADS-box genes from various orchids, rapid amplification of cDNA ends (RACE) and RT-PCR were employed with degenerated primers (Table S3) to isolate partial or full-length cDNAs. The methods and primers used in 3’-RACE and PCR are described in the supporting text. The PCR-amplified products were cloned into a pGEM-T Easy vector (Promega, Madison, WI, USA). For DNA sequencing, 10–12 positive clones were randomly selected.
Phylogenetic analyses
Two datasets for B-class genes were constructed. The PI dataset comprised 74 sequences and the AP3 dataset comprised 63 sequences (Table S2). Full-length (and some partial length) DNA sequences were aligned by using CLUSTALX (Thompson et al. 1997) (Fig. S4) with the default options and then used for nucleotide alignment justification by RevTrans (Wernersson and Pedersen 2003). Phylogenetic analyses involved Bayesian inference (BI) with MrBayes 3.0b4 (Huelsenbeck and Ronquist 2001), and neighbor-joining (NJ) and maximum parsimony (MP) methods involved the use of PAUP*
4.0b10 (Swofford 2002). Detailed information is provided in the supporting text.
Maximum-likelihood tests of selection
The branch lengths with maximum likelihood were calculated with PAUP* 4.0b10 (Swofford 2002) under the GTR + I + G model on the basis of the BI tree obtained. A tree-base maximum likelihood approach was used, as implemented in phylogenetic analysis by maximum likelihood (PAML) (Yang 1997), to test changes in selection in Orchidaceae.
Comparative sets are described in the supporting text.
Fluorescence in situ hybridization (FISH)
Chromosome preparation is described in the supporting text. FISH was performed as described earlier (Chung et al. 2008), with minor modifications. Chromosome preparation consisted of treatment with pepsin (12.5 mg/ml in 10 mM HCl) at room temperature for 60 min to remove the cell debris covering chromosomes. Individual BAC clones (1045F09, 2050C15, 2053A18, 2016E10-B, and 1031M24) representing each AP3 paralog (PeMADS2, PeMADS3, PeMADS4, PeMADS5, and PeMADS6) respectively were used as probes (provided by Dr. W. L. Wu, Department of Life Sciences, NCKU). Detailed information on FISH is provided in the supporting text.
RNA in situ hybridization
Phalaenopsis equestris inflorescence excluded floral buds were fixed in fixation buffer (4% paraformaldehyde and 0.5% glutaraldehyde) at 4°C for 16-24 h. Inflorescence materials were dehydrated through a graded ethanol series, embedded in Histoplast and sectioned at 6-8 µm with use of a rotary microtome (MICROM, HM 310, Walldorf, Germany). Tissue sections were deparaffinized with xylene, rehydrated through an ethanol series, and pre-treated with proteinnase K (2 μg/ml) in 1x phosphate-buffered saline (PBS) at 37°C for 30 min. Prehybridization and hybridization were performed following the protocols in previous study (Tsai et al. 2005). DNA substrates containing a partial C-terminal region and 3’-UTR were used for riboprobe synthesis. PCR products amplified with primers were listed in Table S3. The resulting PCR fragments were used as templates for synthesis of both antisense and sense riboprobes with digoxigenin-labled employing UTP-DIG (Roche Applied Science, Penzberg, Germany) and T7/SP6 Riboprobe® in vitro Transcription System (Promega, Madison, WI, USA) following the manufacturer’s instructions.
RT-PCR analyses
RNA samples were extracted from different floral organs at floral bud stage. Total RNA was treated with RNase-free DNase (NEB, Hertfordshire, England) to remove the contamination of genomic DNA. First-strand cDNA was synthesized by using SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA) from total RNA of the dissected floral organs, including sepals, lateral petals, lip, and column. Semi-quantitative PCR amplification involved 25 and 30 cycles with specific forward and reverse primers located within the partial C-terminal region (Table S3).
Real-time quantitative RT-PCR
The primers for real-time RT-PCR were designed by using Primer Express (Applied Biosystems, Foster City, CA, USA) and gene-specific primers for PeMADS2, PeMADS3, PeMADS4, PeMADS5, and PeMADS6 are listed in Table S3. Real-time RT-PCR reaction used SYBR GREEN PCR Master Mix (Applied Biosystems, Foster City, CA, USA) by using ABI 7500 Real-Time PCR Instrument (Applied Biosystems, Foster City, CA, USA).
Experiments were carried out in triplicates and repeated at least three times independently.
Data were analyzed by using the sequencing detection software (Version 1.2.2, Applied Biosystems).
結果/result
Identification of four subclades of AP3-like genes and two clades of PI-like genes in Orchidaceae.
A total of 24 AP3-like and 13 PI-like genes were identified from 11 species of orchids distributed in four subfamilies (Table S1, Fig. S1). Multiple alignments of protein sequences of AP3 and PI homologs from Orchidaceae, Poaceae, Liliaceae, Arecaceae, and the Eudicots showed that they have distinct C-terminal domains (Fig. S2). The orchid AP3 homologs were divided into four clades, namely AP3A1 (containing PeMADS3), AP3A2 (containing PeMADS4), AP3B1 (containing PeMADS2), and AP3B2 (containing PeMADS5) (Table 1, Fig. S2A). Except the AP3B2 orthologs with truncated ends, all the AP3 homologs contained both a PI-derived motif and a paleoAP3 motif at the end of the C-terminal region (Fig. S2A). In contrast, the PI homologs shared a highly conserved sequence identity (86.0%–99.0%) with the PI motif (Fig. S2B). An insertion/deletion phenomenon was detected in four PI homologs in the subfamily Orchidoideae: a deletion of one amino acid at residue 166 of AfPI-1, LudPI-1, and HpPI-1, and an insertion of one amino acid at residue 193 of HpPI-2 and HrGLO2 (Fig. S2B). Together, these results demonstrate the presence of four subclades of the AP3 homologs and two PI homologs in Orchidaceae.
Phylogenetic analyses of B-class MADS-box genes in Orchidaceae.
To verify the phylogenic relationships of orchid B-class genes, the newly identified orchid B-class genes were analyzed along with the homologs of other seed plants (mainly monocots) reported earlier (Table S2) for resolving their phylogenetic relationships.
Different analytical procedures, including Bayesian inference, maximum parsimony, and neighbour-joining analyses, yielded consistent results (Fig. 2). The orchid AP3 homologs were separated into two major clades, AP3A and AP3B (Fig. 2A), and each subdivided further into two subclades, AP3A1/AP3A2 and AP3B1/AP3B2 (Fig. 2A), in line with the results of sequence analysis shown in Fig. S2. The four paralogous subclades could be the result of two duplication events throughout the evolution of the monocots. It shows that no robust support for the Monocot AP3A and Monocot AP3B clades. The phylogenetic tree indicates that the AP3 duplications predated the Orchidaceae family and may have predated the origin of Asparagales. The secondary duplication probably occurred at or after the diversification of the orchids. The PI ortholog was present as a single-copy gene in Vanilloideae (GalPI and ViPI), and a duplication on the node gave rise to all other Orchidaceae PIs (orchid PI-1 and PI-2), after which both Cypripedioideae and Epidendroideae lost one duplicated PI copy (orchid PI-1) (Fig. 2B). The Cypripedioideae PI (PaphPI) appeared as a group within the orchid PI-2 clade, most members of which were PI orthologs of Orchidoideae and Epidendroideae (Fig. 2B).
Maximum-likelihood tests of selection for the B-class genes in orchids.
To assess selection pressure during the evolution of the lineage, we performed statistical tests for the ratio of non-synonymous (dN) substitutions to synonymous (dS) substitutions (ω = dN/dS). Maximum-likelihood tests reveal that the ω ratio has changed significantly between the Orchidaceae and the rest of the angiosperm lineage. The ω ratio is close to one under neutral evolution, and divergence of the expected ratio points to purifying selection (ω <1) or positive selection (ω >1) (Yang and Bielawski 2000). For the AP3 lineage, we performed the maximum-likelihood test between the clades AP3A and AP3B. The model that assumed different selection pressures for AP3A, AP3B, and all other
AP3 genes performed better than the model that assumed the same ω value (p<0.001) for all. To be specific, the AP3B clade evolved under a more relaxed selection pressure (ωorchidAP3B=0.2275) than the AP3A clade (ωorchidAP3A=0.0353) did. Within clade AP3A, both AP3A1 and AP3A2 do not differ significantly in their ω ratios (Table 2). Similar results were obtained with the two subclades of the AP3B homologs, orchid AP3B1 and orchid AP3B2. Furthermore, the two-ratio model test, which assumes different values of ω between orchids and other PI-like genes, fitted the data significantly better than the one-ratio model, which assumes a constant selection pressure across all the B-class genes (Table 2). There was a significant difference in ω ratios (p<0.001) between the two orchid PI subclades, orchid PI-1 (ωorchidPI-1=0.1175) and orchid PI-2 (ωorchidPI-2=0.0361) (Table 2).
Fluorescence in situ hybridization of the four subclades of orchid AP3 homologs on chromosomes of the Phalaenopsis genome.
To study whether the four duplicated AP3 homologs were located on the same or different chromosomes, each subclade of the orchid AP3 homologs was localized by using fluorescence in situ hybridization (FISH) in Phalaenopsis equestris genome. FISH was carried out with BAC probes, and each probe corresponded to a distinct BAC representing one PeMADS gene. In the nucleus at the G2 stage, signals are presented as paired dots (Fig.
3). The results of FISH revealed that the four subclades of the orchid AP3 homologs were located on different chromosomes because various PeMADS genes were detected as separate pairs of signals in the bi-colored FISH (PeMADS2 and PeMADS3, Fig. 3A;
PeMADS2 and PeMADS4, Fig. 3B; PeMADS2 and PeMADS5, Fig. 3D; and PeMADS4 and PeMADS5, Fig. 3G). In addition, AP3 and PI homologs were located on different chromosomes (PeMADS5 and PeMADS6, Fig. 3J and 3K). These results suggest that the four orchid AP3 homologs may result from genome duplications and yet tandem duplications cannot be rule out as their original source during the evolutions of orchids.
in situ hybridization of PeMADS2-5 at early floral development stage of Phalaenopsis.
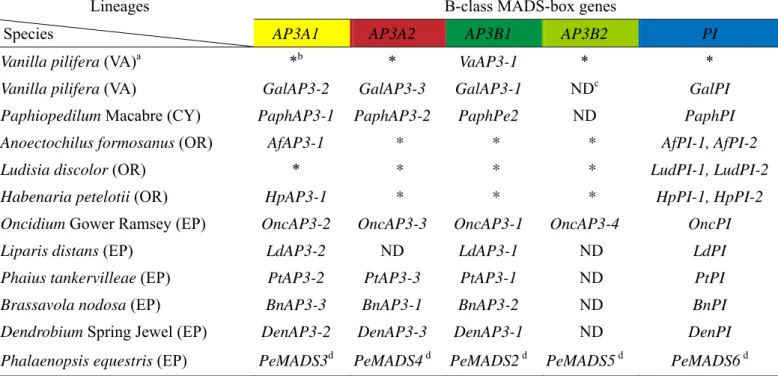
For the purpose of studying spatial expression of these genes, the inflorescence tip of P. equestris was dissected and detail analysis various floral development stages has been defined. In accord with the floral organ developmental program described in Arabidopsis, there is a fundamental similarity in inflorescence and floral development from early to late stages in Phalaenopsis. Flowers originally initiate from the shoot apical meristem (SAM) and undergo an exceedingly long period of identity, development, and growth in most orchids. The Phalaenopsis inflorescence is racemose, and each flower that subtended by its bract is axially on the rachis from the base upward (Fig. 4A). Floral primordia, floral buds, and flowers are arranged in a spiral pattern on the twig (Fig. 4B). Dissecting the tip of the inflorescence, about 11 floral promordia and floral buds constitute the inflorescence in P.
equestris and three major stages of inflorescence development are defined according to their various traits (Fig. 4C, 4D). Stage 1 corresponds to the floral primordia stage in which floral primordia initiate from the inflorescence meristem (Im). Stage 2 accords with the early floral primordia stage, and the formation of sepal, petal, lip and column primordia occurs gradually (Fig. 4C, 4D). Stage 3 corresponds to the late stage of floral organ primordia formation. At stage 3 of inflorescence development, pollinia and callus have developed on column and lip primordia, respectively (Fig. 4C, 4D). After the maturation of floral organ primordia, floral organs enlarge in size and protruding out of their companioning bracts due to cell proliferation and expansion (Fig. 4A, 4B) at Stage 4. Five sub-stages of floral bud stage are assigned according to sizes of floral buds as previously described (Tsai et al. 2004).
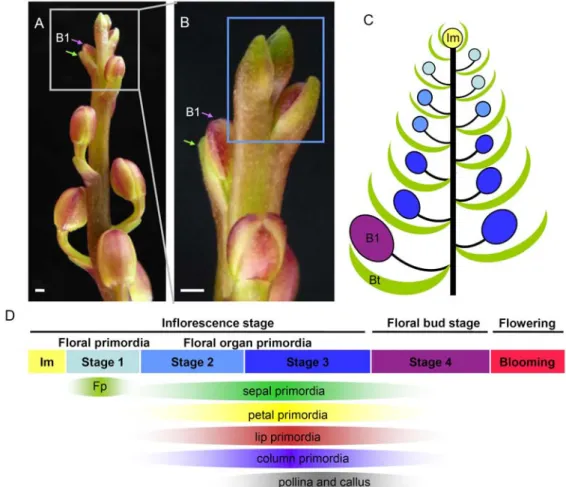
The spatial expression patterns of PeMADS2-5 were studied by in situ hybridization
on longitudinal or transverse sections during floral primordia and floral organ primordia stages of Phalaenopsis inflorescence (Fig. 5E-5P). Unexpectedly, we found that the distinct expression pattern in late floral bud stage (Tsai et al. 2004) was not the case for this experiment in floral organ primordia stage that initiated from the inflorescence meristem and the early floral primordial stage. Instead, the four duplicated AP3 paralogous genes expressed throughout the inflorescence development and floral growth from Stage 1 to Stage 3 (Fig. 5E-5G, 5I-5P). Signals of PeMADS genes were detected after the earliest stage of floral primordia emergence (Stage 1) (Fig. 5I and 5K). Then, the transcripts of PeMADS2, PeMADS4 and PeMADS5 were widely detected in both floral primordia and floral organ primordia developmental stages of Stage 2 and 3 (PeMADS2: Fig. 5E, 5F, and 5G; PeMADS5: Fig. 5I and 5J; and PeMADS4 Fig. 5N, 5O, and 5P). At Stage 2, PeMADS3 expressed in each floral organ primordia including sepal, petal, and lip (Fig. 5L).
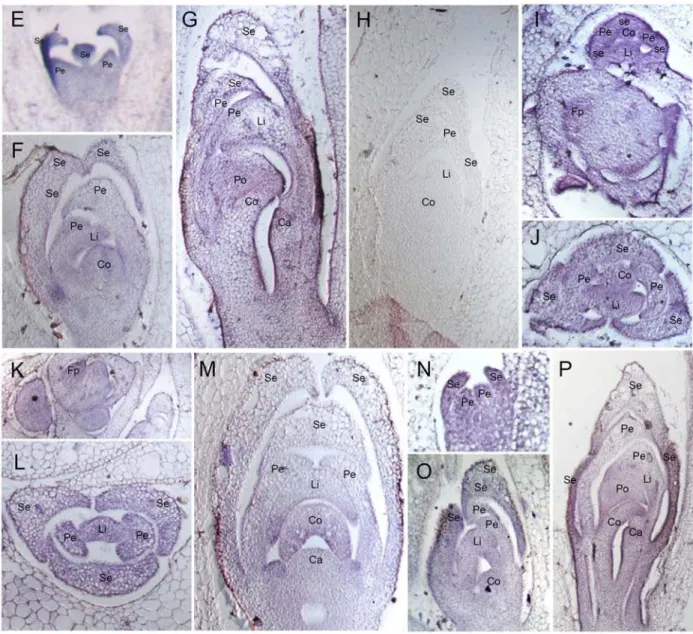
Distinguished expression of PeMADS3 (AP3A1 subclade) in lip and column primordia was first observed at late floral organ primordia stage (stage 3) (Fig. 5M) and it was very weakly expressed in sepal. Moreover, we noticed that the localization of PeMADS2-5 transcripts accumulated predominantly in the epidermal cells during late floral organ primordial stage (Fig. 5G, 5K, 5N, and 5Q). The rearranged expression patterns of AP3 genes in Phalaenopsis showed asynchronous transition points from floral formation through floral organ differentiation and growth and finally floral organ extension (Fig. 6).
The transition point of PeMADS3 gene took place at floral organ primordia stage (Stage 3) (Fig. 6C) while that of PeMADS2, PeMADS5, and PeMADDS4 (Fig. 6A, 6B and 6D) was at floral bud stage (Stage 4). The dualistic transcriptional patterns may indicate the differentiated characters of AP3 homologs in floral organ identities and in floral development and growth.
Spatial expression profiles of orchid B-class MADS-box homologs at floral bud stagae.
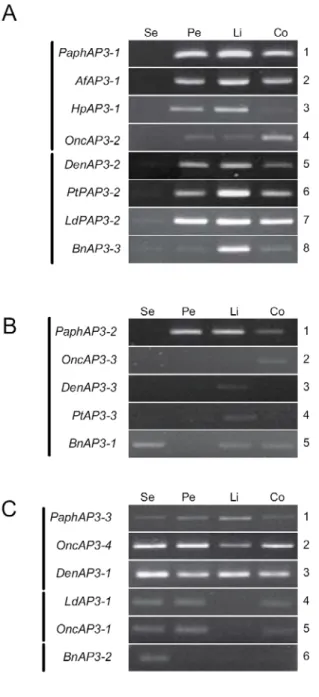
It is interesting to known that whether the duplicated orchid B-class MADS box genes that belong to the same subclade showed the similar expression profiles across subfamilies as those expression in the floral bud stage of Phalaenopsis equestris (Tsai et al. 2004). For this purpose, expression patterns of the four subclades of AP3-like (AP3A1/AP3A2 and AP3B1/AP3B2) and PI-like genes were examined by using semi-quantitative RT-PCR in floral bud stages. As expected, the orchid AP3 orthologous genes displayed diverse expression patterns at late floral bud stages. The expression patterns of genes in AP3A1 subclade were correlated with the perianth types (Fig. 7). Perianth types of Paphiopedilum Macabre, Anoectochilus formosanus, Habenaria petelotii, Oncidium Gower Ramsey are classified as normal (non-peloria), while that of the rest species in this study are semi-pseudopeloria, or pseudopeloria (Fig. S1). according to the similarities between the sepals and lateral petals in First, most AP3A1 orthologs (PaphAP3-1, AfAP3-1, HpAP3-1 and OncAP3-1) were strongly expressed in lateral petals, lip, and column, but not in sepals of normal (non-peloric) perianth (Fig. 7A, rows 1-4). In some cases, extreme faint signals of AP3A1 orthologs (DenAP3-2, PtAP3-2, LdAP3-2 and BnAP3-2) were present in the sepals of semi-pseudopeloric, and pseudopeloric tepal (Fig. 7A, rows 5-8). This result indicated that AP3A1 orthologs may play a part in the homeotically transformed petaloid sepals of semi-pseudopeloric and pseudopeloric flowers, as well as in lateral petals and lip but not in sepals of normal plants. Second, only two AP3A2 orthologs, such as DenAP3-3 and PtAP3-3, limitedly expressed in the lip and absent from all the other floral organs (Fig.
7B, rows 3 and 4).
However, the other AP3A2 homologs show varied expression profiles, they either in column or in sepals, petals (Fig. 7A, rows 1, 2, and 5). Moreover, three AP3B homologs, LdAP3-1, OncAp3-2, and BnAP3-2, were expressed in sepals and/or petals (Fig. 7C, rows
4-6), while the expression of the rest three AP3B homologs (PaphAP3-3, OncAP3-4 and DenAP3-1) were relatively constant in all floral whorls during the development of floral buds (Fig. 7C, rows 1-3). Alternatively, it demonstrated that AP3B clade genes expressed mainly in sepals and lateral petals with occasional exceptions (Fig. 7C, rows 4-6). For PI orthologs (Fig. 7D), since they display extremely high similarity (86.0% - 99.0%), all the PI homologs showed uniformly and highly conserved expression profiles in all four floral organs, similar to that of PeMADS6 (Tsai et al. 2005).
Analyses of homeotic changes in expression profiles of B-class MADS-box homologs associated with the development of prloric flowers.
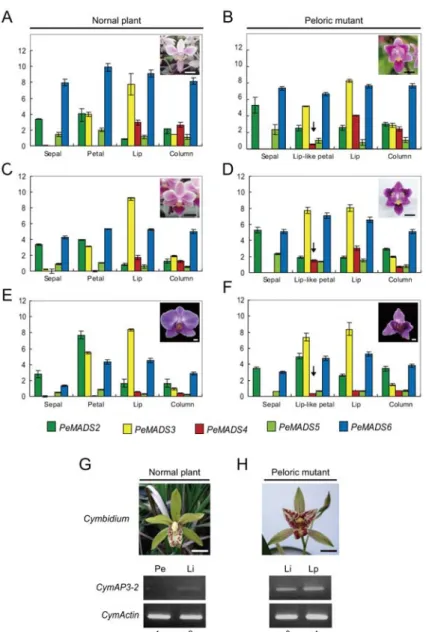
Our earlier study has shown complex protein–protein interactions among the four AP3 and one PI in various floral organs of P. equestris (Tsai et al. 2008). It is intriguing to know whether the expression profiles were altered and lead to the conversion of lateral petals into lip-like petals in the peloric mutants (Fig. 8). In three different pairs of the normal plants and their peloric mutants of Phalaenopsis, expressions of PeMADS2~6 in various floral organs of floral bud stages were compared using real-time quantitative RT-PCR analysis (Fig. 8A-7F). In normal plants, the expressions of PeMADS2 and PeMADS5 were expressed in all floral organs while signals of PeMADS5 were weaker than those of PeMADS2. In addition, slightly higher transcripts can be detected in sepals and petals than in lip and column. The expressions of PeMADS3 were in lateral petals, lip, and column, while undetectable or negligible in sepals. PeMADS4 was expressed in lip and column, while undetectable or negligible in the first two whorls. Moreover, transcripts of PeMADS6 were approximately ubiquitous in all floral organs. Although all the PeMADS2-6 genes were expressed in lip and column, the expression level of PeMADS3 was significantly higher in lip than in column. In the Phalaenopsis peloric mutants, the transcriptional patterns of these five genes in sepals, lip, and column were not affected if any, consistent with the observation that the morphologies of these organs were nearly unaffected in the peloric mutants. Nevertheless, the expression of PeMADS4 is detected in lip-like petals (arrows, in Fig. 8B, 8D, and 8F). In summary, even though the morphologies of sepals and lateral petals are quite similar in orchids, the differential expression of PeMADS3 turned out to be crucial between sepals and petals. In addition, the modified petal of lip shows the differential expression of PeMADS4 in lateral petals to distinct from lip.
To test the strong correlation of AP3A2 genes and lip formation and development in Orchidaceaea, we examined the expression patterns of AP3A2 ortholog in normal plant Cymbidium and its homeotic mutant of peloria (Fig. 8G and 8H). The AP3A2 clade gene was only expressed in the lip of normal plant of Cymbidium tepal (Fig. 8G, lane 2).
Expectedly, the expression of AP3A2 clade genes was extended to the lip-like petals in the peloric mutant of Cymbidium (Fig. 8H, lane 3). Taken together, the fact that AP3A2 orthologs have played a noticeable role in lip formation was strongly supported by further evidence of their specific transcripts in both lip and lip-like petals during floral bud growth in Phalaenopsis and Cymbidium peloria mutant species.
討論/discussion
Detecting the selection of orchid B-class MADS genes.
The orchid PI-like homologs show greater similarities in their sequences than AP3-like homologs. Both PI-like and AP3-like homologs have evolved under a different selection pressure (measured by ω ratios) (data not shown), observations that are consistent with earlier studies on PI-like genes in general (Hernández-Hernández et al. 2007).
Furthermore, both orchid AP3A and AP3B clades show significantly different selection pressures, suggesting that these two duplicated lineages are selected for different functions.
However, no differential selection pressure was detected within the two subclades of either orchid AP3A (AP3A1 and AP3A2) or AP3B (AP3B1 and AP3B2), suggesting that the two pairs of AP3 subclade paralogs may have a high functional redundancy. Alternatively, it may due to their overlapping expression domains. For this reason, further functional analysis of genes involved in regulating the identity of floral organs in orchids by using loss-of-function mutations or virus-induced gene silencing (VIGS) will be necessary to elucidate the regulatory network at the molecular level that governs morphogenesis in orchids and to provide a physical basis for genetic engineering in orchids. Knock down of floral MADS-box genes simultaneously in Phalaenopsis by inoculating of PeMADS6 at a conserved region among MADS-box genes with CymMV transcripts indicate that flower organ become greenish streaks and patches in the silenced plants suggesting a conversion of reproductive tissue to vegetative tissue (Lu et al. 2007). Moreover, down-regulation of PeMADS6 expression leads to severe phenotypic effects in floral perianth of Phalaenopsis by using a modified optimal method in the VIGS plants (Hsieh et al., unpublished data).
Homeotic developmental changes of epidermal cell types on both adaxial and abaxial surfaces of sepals and lateral petals with a tendency toward phyllody may indicate an alternation of floral organ identities. The differential silencing levels of PeMADS2-6 genes floral organs in the PeMADS6-silenced plants were examined with real-time RT-PCR. In PeMADS6-silenced Phalaenopsis flowers, the transcription level of target B-class MADS-box gene (PeMADS6) was reduced significantly, while that of non-target B-class genes (PeMADS2-5) were knocked down with varied degrees (Hsieh et al., unpublished data).
Dualistic transcriptional patterns of AP3-like genes in orchids.
The fact that the combinational expression profiles of orchid AP3-like genes span the whole floral organ development program may indicate their roles in biological processes, through the organ identity specification, cell proliferation, cell expansion, and cell differentiation and maturation (Dornelas et al. 2010). It is noticeable that the transition points of transcripts from ubiquitous expression patterns to confined floral organs are different among the duplicated AP3 paralogs in Phalaenopsis orchid. In this paper, results from in situ hybridization and Northern blots (Tsai et al. 2004) revealed that Phalaenopsis AP3-like genes expressed regionally in particular floral organs to determine floral organ identity at the floral organ primordia stage and to promote floral growth and development at floral bud stage. The confined expression of AP3A1 gene was observed at inflorescence stage when the floral formation begins, while that of AP3A2 and AP3B homologs were detected at floral bud stage that occur after the onset of floral organ differentiation and volume expansion. The major critical transition points represented by the asynchrony of relocated expressions in duplicated AP3 paralogs not only imply their dualistic roles in floral organ specification, but also indicate that the shifting patterns of AP3 genes in both temporal and spatial may determine the fate of orchid perianth growth and development
conspicuously.
The specific tendency of AP3 genes in orchid perianth development.
Morphology of orchid perianth has developed into five tepal-like floral organs and a distinguished lip during its evolution. The obscuring molecular mechanism in regulation of specific floral organ formation may be uncovered by gene duplication and subsequent sub- and/or neo-functionalization. The duplicated and divergent genes may result from alteration of the promoter region and/or in changes in coding sequence by mutation following a shift in expressional regulation (Force et al. 1999; Moore et al. 2005). Our results revealed that orchid duplicated AP3A and AP3B show significantly different selection pressure and distinct expression profiles. Through this, gene functions can evolve to regulate floral organs morphogenesis, such as the formation of lip may be contributed by the duplication of B-class MADS-box genes in orchids. Although the orchid tepal may seem extremely similar organs, but results from in situ hybridization display that dualistic patterns of AP3A1 genes had discriminated the intrinsic qualities between the sepals and the petals since early floral development stage. In addition to the shifted homeotic and distinct expression of AP3A2 homologs in lip-like petals of peloric mutants of P. equestris and C.
ensifolium in this study, similar observation has been reported that greenish sepals are changed to white lip-like organs due to the extending expression of HrDEF, an AP3A2 homolog, in the petaloid-sepal mutant of Habenaria radiate (Kim et al. 2007). Complete peloric mutants have also been reported in natural populations of both Paphiopedilium rothschildianum (Cypripedioideae), and Catteleya (Epidendroideae) (Rudall and Bateman 2003) as well. Peloric mutants seem to broadly exist in natural populations of the above subfamilies of Orchidaceae, and they may indeed result from ectopic expression of the single orchid AP3A2 homologs. These observations set the foundations of the sub-functionalization of orchid AP3A genes by changing their expressions. It is plausible that the diversification of PeMADS4 (AP3A2 subclade) during evolution was a major driving force behind the development of different types of petal organs. Therefore, the orchid AP3A2 gene plays an essential role in specifying the development of the lip. The zygomorphic expressions of the AP3A2 homologs correlate with the highly modified perianth development in Orchidaceae. These results indicated that the duplicated factors modify the expression patterns of original B-class genes in species with tepal flowers and represent the restoration of their function in orchid perianth development.
“Homeotic Orchid Tepal Model”: a molecular regulatory mechanism of orchid floral development.
In model plants, such as Arabidopsis and Antirrihnum, transcripts of AP3 and PI are typically expressed in petals and stamen, following the well-known “ABC model” where homologs of B-class genes are broadly expressed in tepals, stamens, and carpals in basal angiosperm (Soltis et al. 2007). The AP3-like genes from tepal-like monocots such as lily and tulips also show similar expression patterns in the outer three whorls supporting the
“modified ABC model” (Tzeng and Yang 2001). However, floral development of all angiosperm species can not be commonly and perfectly explained by these two models. For investigating the evolutionary history of the perianth, developmental genetic data was used to infer the ancestral perianth character state for different angiosperm clades (Hileman and Irish 2009). In orchids, so far, SQUAMOSA (SQUA)-like genes, DOMADS2 from Dendrobium madame Thong-In (Yu and Goh 2000), DthyrFL1, 2 and 3 from D.
thyrsiflorum (Skipper et al. 2005), and ORAP11 and ORAP13 from Phalaenopsis (Chen et al. 2007) have been identified, but these genes do not perform the A-function role in determining the identities of floral organs. Instead, they maintain their ancestral role of
determining the identity of the meristem (Chen et al. 2007).
The likelihood that the monocot ancestor of AP3 homolog was either regional, but weak expressed in perianth organs or ubiquitous in perianth organs from early through late stages of floral development (Hileman and Irish 2009). Results of our study appear that the expression profiles of AP3A1-like genes in orchids were restricted in specific floral organs and different stages whereas AP3A1 and AP3B-like gene transiently show ubiquitous expression in inflorescence stage, and expressed in distinguished floral organs only in floral bud stage. This raised the discrepancy between the transition timing for determination of floral organ identity (Stage 2 of inflorescence stage) and that for the expression of AP3 genes. Two possibilities can be evoked to elucidate the asynchronous phenomena of re-localized transcripts of AP3-like genes following the various floral identity stages. First, except for AP3-like MADS-box genes, other factors specifically determined the floral organ identities in orchid perianth. Based on this concept, an AP1/AGL9 group of MADS-box gene, OMADS1, showing high homology to AGL6, is expressed in the apical meristem, lip, and carpel (Hsu et al. 2003). Moreover, OMADS1 can interact with OMADS3, which belongs to AP3B2 subclade (Hsu et al. 2003). In addition, AGL6-like gene also showed specific expression pattern in lip and it can be involved in the dimer protein complex composed of AP3A and AGL6 proteins (Luo et al., unpublished data).
Further complications arise from the regulatory functions of the five B-class PeMADSs involved in the formation of complexes with various combinations of the duplicated homologs (Tsai et al. 2008). These reports suggest that specific expression patterns for the duplicated AP3 and PI homologs and other AGL6-like genes may account for the multiple protein–protein interactions, which specify the identity of different floral organs, especially petals and lips, in orchids. Thus, the lip growth in floral bud stage may not be completely determinated by AP3A2 genes.
The other possibility explains that the floral organ identities of orchid perianth were determinated by two gradual progresses from inflorescence through floral bud stages. The floral primordia are not discriminated into sepal promordia and inner tepal primordia until the first transition point (AP3A1) during late inflorescence stage, while the petal and lip tissue are categorized after the second transition point (AP3A2 and AP3B) in the floral bud stage. If this is the case, it is possible that the differences in morphology among sepals, lateral petals, and the lip in Orchidaceae lie in the combination of the duplicated B-class genes and other factors. Thus, a “Homeotic Orchid Tepal (HOT) Model” is proposed to explain the regulation of perianth morphogenesis in Orchidaceae in both spatial and temporal condition (Fig. 9A and 9B). In this model, PI and AP3B clades determine the formation of sepals. The combination of PI and both AP3A1 and AP3B clades control the lateral petal formation at inflorescence stage, whereas PI and AP3A1, AP3A2 clade genes and an AGL6-like gene contribute to lip morphogenesis in floral bud stage. In addition to B-class genes, other floral homeotic genes such as C- and D-class MADS-box proteins is also involved in the formation of multimeric protein complexes to define the organ identities of the column (Xu et al. 2006). According to the proposed HOT model, the combinations of homeotic protein complexes can form high-order complexes of multimers with more components than the quartet model (Theissen 2001). This novel multimerization model appears to demonstrate the extraordinary evolutionary history of floral zygomorphic morphogenesis in orchids.
參考文獻/reference
Cameron, K.M. (2004) Utility of plastid psaB gene sequences for investigating intrafamilial relationships within Orchidaceae. Mol Phylogenet Evol 31: 1157-1180.
Chang, Y.Y., Kao, N.H., Li, J.Y., Hsu, W.H., Liang, Y.L., Wu, J.W. and Yang, C.H. (2010) Characterization of the possible roles for B class MADS box genes in regulation of perianth formation in orchid. Plant Physiol 152: 837-853.
Chen, D., Guo, B., Hexige, S., Zhang, T., Shen, D. and Ming, F. (2007) SQUA-like genes in the orchid Phalaenopsis are expressed in both vegetative and reproductive tissues.
Planta 226: 369-380.
Chung, M.C., Lee, Y.I., Cheng, Y.Y., Chou, Y.J. and Lu, C.F. (2008) Chromosomal polymorphism of ribosomal genes in the genus Oryza. Theor Appl Genet 116: 745-753.
Cozzolino, S. and Widmer, A. (2005) Orchid diversity: an evolutionary consequence of deception? Trends Ecol Evol 20: 487-494.
Cubas, P. (2004) Floral zygomorphy, the recurring evolution of a successful trait. Bioessays 26: 1175-1184.
Dornelas, M.C., Patreze, C.M., Angenent, G.C. and Immink, R.G. (2010) MADS: the missing link between identity and growth? Trends Plant Sci 16: 89-97.
Dressler, R.L. (1993a) Orchid Phylogeny In Phylogeny and classification of the orchid family. Edited by Dudley, T.R. pp. 59-82. Dioscorides Press, Portland, Oregon.
Dressler, R.L. (1993b) Phylogeny and classification of the orchid family. Dioscorides Press, Portland, Or.
Force, A., Lynch, M., Pickett, F.B., Amores, A., Yan, Y.L. and Postlethwait, J. (1999) Preservation of duplicate genes by complementary, degenerative mutations. Genetics 151: 1531-1545.
Hernández-Hernández, T., Martínez-Castilla, L.P. and Alvarez-Buylla, E.R. (2007) Functional diversification of B MADS-Box homeotic regulators of flower development:
Adaptive evolution in protein-protein interaction domains after major gene duplication events. Mol Biol Evol 24: 465-481.
Hileman, L.C. and Irish, V.F. (2009) More is better: the uses of developmental genetic data to reconstruct perianth evolution. American Journal of Botany 96: 83-95.
Hsu, H.F., Huang, C.H., Chou, L.T. and Yang, C.H. (2003) Ectopic expression of an orchid (Oncidium Gower Ramsey) AGL6-like gene promotes flowering by activating flowering time genes in Arabidopsis thaliana. Plant Cell Physiol 44: 783-794.
Hsu, H.F. and Yang, C.H. (2002) An orchid (Oncidium Gower Ramsey) AP3-like MADS gene regulates floral formation and initiation. Plant Cell Physiol 43: 1198-1209.
Huelsenbeck, J.P. and Ronquist, F. (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754-755.
Kanno, A., Saeki, H., Kameya, T., Saedler, H. and Theissen, G. (2003) Heterotopic expression of class B floral homeotic genes supports a modified ABC model for tulip (Tulipa gesneriana). Plant Mol Biol 52: 831-841.
Kim, S.-Y., Yun, P.-Y., Fukuda, T., Ochiai, T., Yokoyama, J., Kameya, T. and Kanno, A.
(2007) Expression of a DEFICIENS-like gene correlates with the differentiation between sepal and petal in the orchid, Habenaria radiata (Orchidaceae). Plant Sci 172:
319-326.
Kramer, E.M. and Hall, J.C. (2005) Evolutionary dynamics of genes controlling floral development. Curr Opin Plant Biol 8: 13-18.
Kramer, E.M. and Jaramillo, M.A. (2005) Genetic basis for innovations in floral organ identity. J Exp Zoolog B Mol Dev Evol 304: 526-535.
Lu, H.C., Chen, H.H., Tsai, W.C., Chen, W.H., Su, H.J., Chang, D.C. and Yeh, H.H. (2007)
Strategies for functional validation of genes involved in reproductive stages of orchids.
Plant Physiol 143: 558-569.
Mondragon-Palomino, M. and Theissen, G. (2008) MADS about the evolution of orchid flowers. Trends Plant Sci 13: 51-59.
Mondragon-Palomino, M. and Theissen, G. (2009) Why are orchid flowers so diverse?
Reduction of evolutionary constraints by paralogues of class B floral homeotic genes.
Ann Bot 104: 583-594.
Moore, R.C., Grant, S.R. and Purugganan, M.D. (2005) Molecular population genetics of redundant floral-regulatory genes in Arabidopsis thaliana. Mol Biol Evol 22: 91-103.
Nakada, M., Komatsu, M., Ochiai, T., Ohtsu, K., Nakazono, M., Nishizawa, N.K., Nitta, K., Nishiyama, R., Kameya, T. and Kanno, A. (2006) Isolation of MaDEF from Muscari armeniacum and analysis of its expression using laser microdissection. Plant Sci 170:
143-150.
Nakamura, T., Fukuda, T., Nakano, M., Hasebe, M., Kameya, T. and Kanno, A. (2005) The modified ABC model explains the development of the petaloid perianth of Agapanthus praecox ssp. orientalis (Agapanthaceae) flowers. Plant Mol Biol 58: 435-445.
Park, J.H., Ishikawa, Y., Ochiai, T., Kanno, A. and Kameya, T. (2004) Two GLOBOSA-like genes are expressed in second and third whorls of homochlamydeous flowers in Asparagus officinalis L. Plant Cell Physiol 45: 325-532.
Rudall, P.J. and Bateman, R.M. (2002) Roles of synorganisation, zygomorphy and heterotopy in floral evolution: the gynostemium and labellum of orchids and other lilioid monocots. Biol Rev Camb Philos Soc 77: 403-441.
Rudall, P.J. and Bateman, R.M. (2003) Evolutionary change in flowers and inflorescences:
evidence from naturally occurring terata. Trends Plant Sci 8: 76-82.
Skipper, M., Pedersen, K.B., Johansen, L.B., Frederiksen, S., Irish, V.F. and Johansen, B.B.
(2005) Identification and quantification of expression levels of three FRUITFULL-like MADS-box genes from the orchid Dendrobium thyrsiflorum (Reichb.f.). Plant Science 169: 579-586.
Soltis, D.E., Ma, H., Frohlich, M.W., Soltis, P.S., Albert, V.A., Oppenheimer, D.G., Altman, N.S., dePamphilis, C. and Leebens-Mack, J. (2007) The floral genome: an evolutionary history of gene duplication and shifting patterns of gene expression. Trends Plant Sci 12:
358-367.
Stellari, G.M., Jaramillo, M.A. and Kramer, E.M. (2004) Evolution of the APETALA3 and PISTILLATA lineages of MADS-box-containing genes in the basal angiosperms. Mol Biol Evol 21: 506-519.
Swofford, D.L. (2002) PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Sinauer Associates, Sunderland, Massachusetts, USA.
Theissen, G. (2001) Development of floral organ identity: stories from the MADS house.
Curr Opin Plant Biol 4: 75-85.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F. and Higgins, D.G. (1997) The CLUSTAL-X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25: 4876-4882.
Tsai, W.C., Kuoh, C.S., Chuang, M.H., Chen, W.H. and Chen, H.H. (2004) Four DEF-like MADS box genes displayed distinct floral morphogenetic roles in Phalaenopsis orchid.
Plant Cell Physiol 45: 831-844.
Tsai, W.C., Lee, P.F., Chen, H.I., Hsiao, Y.Y., Wei, W.J., Pan, Z.J., Chuang, M.H., Kuoh, C.S., Chen, W.H. and Chen, H.H. (2005) PeMADS6, a GLOBOSA/PISTILLATA-like gene in Phalaenopsis equestris involved in petaloid formation, and correlated with flower longevity and ovary development. Plant Cell Physiol 46: 1125-1139.
Tsai, W.C., Pan, Z.J., Hsiao, Y.Y., Jeng, M.F., Wu, T.F., Chen, W.H. and Chen, H.H. (2008)
Interactions of B-class complex proteins involved in tepal development in Phalaenopsis orchid. Plant Cell Physiol 49: 814-824.
Tzeng, T.Y. and Yang, C.H. (2001) A MADS box gene from lily (Lilium Longiflorum) is sufficient to generate dominant negative mutation by interacting with PISTILLATA (PI) in Arabidopsis thaliana. Plant Cell Physiol 42: 1156-1168.
Wernersson, R. and Pedersen, A.G. (2003) RevTrans - Constructing alignments of coding DNA from aligned amino acid sequences. Nucleic Acids Res 31: 3537-3539.
Xu, Y., Teo, L.L., Zhou, J., Kumar, P.P. and Yu, H. (2006) Floral organ identity genes in the orchid Dendrobium crumenatum. Plant J 46: 54-68.
Yang, Z. (1997) PAML: a program package for phylogenetic analysis by maximum likelihood. Comput Appl Biosci 13: 555-556.
Yang, Z. and Bielawski, J.P. (2000) Statistical methods for detecting molecular adaptation.
Trends Ecol Evol 15: 496-503.
Yu, H. and Goh, C.J. (2000) Identification and characterization of three orchid MADS-box genes of the AP1/AGL9 subfamily during floral transition. Plant Physiol 123:
1325-1336.
Zahn, L.M., Leebens-Mack, J., DePamphilis, C.W., Ma, H. and Theissen, G. (2005) To B or Not to B a flower: the role of DEFICIENS and GLOBOSA orthologs in the evolution of the angiosperms. J Hered 96: 225-240.
附表及附圖/table and figure
Table 1. AP3 and PI lineages identified in this study.
Lineages B-class MADS-box genes
Species AP3A1 AP3A2 AP3B1 AP3B2 PI
Vanilla pilifera (VA)a *b * VaAP3-1 * *
Vanilla pilifera (VA) GalAP3-2 GalAP3-3 GalAP3-1 NDc GalPI Paphiopedilum Macabre (CY) PaphAP3-1 PaphAP3-2 PaphPe2 ND PaphPI Anoectochilus formosanus (OR) AfAP3-1 * * * AfPI-1, AfPI-2
Ludisia discolor (OR) * * * * LudPI-1, LudPI-2
Habenaria petelotii (OR) HpAP3-1 * * * HpPI-1, HpPI-2
Oncidium Gower Ramsey (EP) OncAP3-2 OncAP3-3 OncAP3-1 OncAP3-4 OncPI
Liparis distans (EP) LdAP3-2 ND LdAP3-1 ND LdPI
Phaius tankervilleae (EP) PtAP3-2 PtAP3-3 PtAP3-1 ND PtPI Brassavola nodosa (EP) BnAP3-3 BnAP3-1 BnAP3-2 ND BnPI Dendrobium Spring Jewel (EP) DenAP3-2 DenAP3-3 DenAP3-1 ND DenPI Phalaenopsis equestris (EP) PeMADS3d PeMADS4 d PeMADS2 d PeMADS5 d PeMADS6 d
a Abbreviations are of subfamilies of Orchidaceae: CY, Cypripedioideae; EP, Epidendroideae;
OR, Orchidoideae; and VA, Vanilloideae.
b Orthologs did not survey in this study.
c Orthologs cannot be identified by RACE and RT-PCR with degenerated primers.
d Sequence identified by Tsai et al., 2004, 2005.
Table 2. Maximum likelihood tests of selection for the B-class genes.
-ln likelihood Γ=2(lnL0-lnL1) Probability
AP3 lineage
One ratio -2924.85 (ω=0.103)
Two ratio
(Orchid vs all others) -2923.204 3.29
(vs. AP3 one ratio) <0.1 All others (ωAllotherAP3=0.1162)
Orchid AP3 (ωorchidAP3=0.0746) Orchid AP3 A vs B
A = B ≠ all others -2923.204
All others (ωAllotherAP3=0.1162)
Orchid AP3 A = B (ωorchidAP3A&B=0.0746) A ≠ B ≠ all others -2913.718 18.97
(vs. A=B≠ all others) <0.001a All others (ωAllotherAP3=0.1148)
OrchidAP3A (ωorchidAP3A=0.0353) OrchidAP3B (ωorchidAP3B=0.2275) Orchid AP3 A1 vs A2
A1 = A2 ≠ all others -2915.670 18.36
(vs. AP3 one ratio) <0.001a All others (ωAllotherAP3=0.1258)
OrchidAP3A1=A2 (ωorchidAP3A=0.0353) A1 ≠ A2 ≠ all others -2915.518 0.304
(vs. A1=A2≠ all others) >0.5 All others (ωAllotherAP3=0.1258)
OrchidAP3A1 (ωorchidAP3A1=0.0396) OrchidAP3A2 (ωorchidAP3A2=0.0276) Orchid AP3 B1 vs B2
B1 = B2 ≠ all others -2921.231 7.238
(vs. AP3 one ratio) <0.01b All others (ωAllotherAP3=0.0922)
OrchidAP3B1=B2 (ωorchidAP3B1=0.2315)
B1 ≠ B2 ≠ all others -2920.639 1.184
(vs. B1=B2 ≠ all others) <0.5 All others (ωAllotherAP3=0.0922)
OrchidAP3B1 (ωorchidAP3B1=0.1866) OrchidAP3B2 (ωorchidAP3B2=0.3965) PI lineage
One ratio -9578.499 (ωPI = 0.082) Two ratios
Orchid ≠ Monocots = all others -9574.21 8.578
(vs. PI one ratio) <0.01b All others (ωAllotherPI=0.0875)
Orchid PI (ωorchidPI=0.0515) Three ratios
Monocots ≠ Orchids PI-1 ≠ Orchids PI-2 ≠ Vanilloideae
-9566.119
(ωMonocotsPI=0.0874) (ωVanilloidPI=0.0085) (ωorchidPI-1=0.1175) (ωorchidPI-2=0.0361)
16.18
(vs. PI one ratio for orchids) <0.001a ap value < 0.001 level
bp value < 0.01 level
Figure 1. Schemes illustrate perianth of flowers types in orchids. The normal perianth (A), semi-pseudopeloria (B), and pseudopeloria (C). The perianth organs shown here are
represented with three colours: green, sepal; yellow, lateral petal; yellow-green grid, mild transformation of sepal to lateral petal, and red, lip. Se, sepal; Pe, lateral petal; Li, lip; Co, column.
Fig. 1
Fig. 2A
Fig. 2B
Figure 2. Phylogenetic trees of B-class MADS-box genes for the AP3 and PI lineages from Orchidaceae. Phylogenetic trees based on Bayesian inference (BI) were constructed for (A) AP3 and (B) PI genes, from Orchidaceae, other monocots, basal angiosperms, and gymnosperms. The numbers above the branches are posterior probabilities from BI analysis (only posterior probabilities above 0.50 are shown) and the numbers below the branches are Neighbor-joining (NJ) analysis (only bootstrap values over 50 are shown). Circles mark the lineages, clades, and subclades of AP3-like genes as well as the clades of PI-like genes.
Abbreviations to the right of the tree are of subfamilies of Orchidaceae: CY, Cypripedioideae;
EP, Epidendroideae; OR, Orchidoideae; and VA, Vanilloideae.
Fig. 2
Figure 3. Identification of genome relationships of PeMADS2-6 by using FISH. The four duplicated AP3 homologs (PeMADS2, PeMADS3, PeMADS4, and PeMADS5) and the PI homolog (PeMADS6) were localized in P. equestris genome by using FISH. The green signals of PeMADS2 are distinguishable from the red signals of PeMADS3 (A), PeMADS4 (B), or PeMADS5 (D). Chromosomes are counterstained in blue with DAPI. (C) The image extracted from (B) shows that chromosomes (arrows) containing red or green signals are different. The distribution of PeMADS4 (E) and PeMADS5 (F) alone is presented as red signals. (G) The positions of PeMADS4 (green) and PeMADS5 (red) are different. (H and I) The distribution of PeMADS6 (red) alone is shown. (J and K) The positions of PeMADS5 (green) and PeMADS6 (red) are different. The scale bar in each panel is 8 μm.
Fig. 3
Figure 4. Inflorescence and flower development in Phalaenopsis. Photograph of P.
equestris inflorescence and developing floral buds (A and B). Blue box represents the
inflorescence stage (Stage1-3), White arrow indicates the first floral bud (B1) at Stage 4, and green arrow indicated its bract. (Scale bar=1 mm) (C) Schematic representation of
Phalaenopsis inflorescence. The light yellow color symbolizes inflorescence meristem, the light blue, blue, dark blue, and purple colours represent Stage 1, 2, and 3 at floral primordial or floral organ primordial sub-stages and floral bud stage, respectively. Im, inflorescence meristem; Fp, floral primordium. (D) Major stages defined for Phalaenopsis inflorescence and floral bud development before flowering. Except for inflorescence meristem, three sub-stages (Stage 1-3) are assigned for the inflorescence stage on the basis of differentiation of the floral primordia and floral organ primordia. The floral primordia stage (Stage 1) is defined as the formation of floral primordia. Within the floral organ primordia stage (Stage 2-3), sepal, lateral petal, lip, and column primordia are formed at Stage 2, while pollina and callus develop on column and lip, respectively at Stage 3. Floral bud stage (Stage 4) is assigned according to the extending floral buds out of their bracts.
Fig. 4
Figure 5. RNA in situ hybridization analysis of PeMADS2~5 transcripts in inflorescence stage of Phalaenopsis. (A-L) Longitudinal and transverse sections of the tip of inflorescence were hybridized with antisense RNA probes of PeMADS2 (A, B, C), PeMADS5 (E, F), PeMADS3 (G, H, I), and PeMADS4 (J, K, L). Probe with sense RNA fragment of PeMADS2 was severed as a negative control (D). Primordia in (E, G), (A, B, E, H, J, K), and (C, D, F, I, L) are at Stage 1, 2, and 3, respectively. Im, inflorescence meristem; Fp, floral primordium;
Bt, bract, Se, sepal; Pe, lateral petal; Li, lip; Co, column; Ca, callus; Po, pollinia.
Fig. 5
Figure 6. Dualistic expressions of MADS-box genes for both floral organ identity and growth in orchids. Symbolized spatial expression patterns of AP3-like MADS-box genes during floral organ development and developing floral bud stages. Schematic representation was derived from both combined results of in situ hybridization at inflorescence stage (this study) and Northern blot at floral bud stage (Tsai et al. 2004). The two stages can be
distinguished with the floral bud protruding from its bract observed at floral bud stage. Floral zones shown in colour are dependent on the expression profiles of PeMADS2-5. (A) Green, PeMADS2, (B) grass green, PeMADS5; (C) yellow, PeMADS3; and (D) red, PeMADS4. The transition points from ubiquitous expression in the entire floral primordia and floral organ primordia to constrained floral organs are seemingly different, for PeMADS2, 4 and 5 are at floral bud stage, while that of PeMADS3 gene is at the late floral organ primordia stage.
Arrows indicate the transition points. Se, sepal; Pe, lateral petal; Li, lip; Co, column.
Fig. 6
Figure 7. Expression patterns of AP3- and PI-like genes in orchids. Semi-quantitative RT-PCR was performed for the analysis of expression levels of the B-class homologs of AP3A1 (A), AP3A2 (B), AP3B1/2 (C), PI (D), in various floral organs, Actin (E) served as an internal control. Se, sepal; Pe, lateral petal; Li, lip; Co, column.
Fig. 7
Figure 8. Ectopic expression of AP3A2 orthologs in prloric flowers of Phalaenopsis and Cymbidium. (A-F) Three pairs of normal plants and their peloric mutant flowers of
Phalaenopsis equestris. (A, B) A diploid P. equestris (S82-159) and its peloric mutant (S85-355); (C, D) a tetraploid P. equestris ‘King Car’ and its peloric mutant; (E, F) a P. spp.
‘A3947’ and its peloric mutant. (D) One pair of Cymbidium ensifolium and its peloric mutant flower. (Scale bar = 1 cm) Real-time quantitative RT-PCR analysis of PeMADS2~6 in three pairs of dissected floral organs at floral bud stage in Phalaenopsis. Green bar: PeMADS2, grass green bar: PeMADS5, yellow bar: PeMADS3, red bar: PeMADS4, blue bar: PeMADS6.
Arrows in (B), (D), and (F) indicate the ectopic expressions of PeMADS4 in the lip-like petals of peloric mutants. (G, H) By using RT-PCR analysis, the transcript of AP3A2 clade gene expressed in lip (lane 2) of wild-type of Cymbidium ensifolium flower, and it extends to the lip-like petals in the peloric mutant of Cymbidium (lanes 3-4). Pe, lateral petal; Li, lip; Lp, lip-like petal.
Fig. 8
Figure 9. Proposed “Homeotic Orchid Tepal (HOT) Model” for orchid floral organ identity and growth. The expression patterns of the B-, C-, D- class and other MADS-box genes in Orchidaceae led to the HOT model. Orchid floral organ identity and growth are determined at the inflorescence stage (A), while the follow-up growth is at the floral bud stage (B). PI and AP3B clade determine the formation of sepals. The combination of PI and both AP3A1 and AP3B controls the formation of lateral petals. Both PI and AP3A2 clade genes (and/or other MADS-box genes) control the formation of lip. Green, AP3B1; grass green, AP3B2; yellow, AP3A1; Red, AP3A2; blue, PI-like genes; purple, C- and D-class genes; and pink, unknown MADS genes.
Fig. 9
附錄
發表刊物名稱、卷期及出版日期
Plant cell and Physiology (2011) July (in press)