棲蘭山環境溫度對親屍性甲蟲多樣性的影響
黃文伯1) 賴韋翰1) 葛兆年2,3)
摘 要
本研究以懸掛型誘餌掉落式陷阱調查棲蘭山親屍性甲蟲的群聚結構,並探討環境溫度變化對物種 豐度、豐量與多樣性的影響。於2013年1月至12月,共捕獲親屍性甲蟲6科70種1772隻,其中以隱翅蟲 科、球蕈蟲科與埋葬蟲科在個體豐量上較佔優勢。親屍性甲蟲的組成結構在人工林與天然林之間,具 有36.81% (Wainstein’s Similarity Index, Kw)的相似性。天然林的月均溫顯著比人工林低,但在每月捕 獲的個體豐量上,在天然林則顯著較高。無論在人工林或天然林,物種豐度、豐量與Shannon-Wiener 多樣性指數三者以二次多項式分析,皆與月均溫有顯著的正相關,環境溫度越高,親屍性甲蟲的物種 豐度、豐量與Shannon-Wiener多樣性指數亦越高。此外,與人工林相比,天然林的二次多項式曲線 則往較低溫的方向偏移。此顯示中海拔霧林帶棲蘭山的環境溫度,對親屍性甲蟲的群聚結構有顯著的 影響。
關鍵詞:親屍性甲蟲、多樣性、溫度、森林、棲蘭山。
黃 文 伯 、 賴 韋 翰 、 葛 兆 年 。2014。棲蘭山環境溫度對親屍性甲蟲多樣性的影響。台灣林業科學 29(Supplement):S55-64。
1)國立臺南大學生態科學與技術學系,70005台南市中西區樹林街二段33號 Department of Ecoscience and Ecotechnology, National Univ. of Tainan, 33 Shulin St., Sec. 2, West Central District, Tainan 70005, Taiwan.
2)林業試驗所森林保護組,10066台北市南海路53號 Department of Forest Protection, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
3)通訊作者 Corresponding author, e-mail:[email protected]
2014年8月送審 2014年11月通過 Received August 2014, Accepted November 2014.
Research paper
The Effect of Ambient Temperature on the Biodiversity of Necrophilous Beetles at Chilanshan, Northeastern Taiwan
Wenbe Hwang,1) Wei-Han Lai,1) Chao-Nien Koh2,3)
【Summary】
Baited hang-fall traps were used to research the community structure of necrophilous beetles at Chilanshan, northeastern Taiwan and the effects of changes in the ambient temperature on the species richness, abundance, and diversity. In total, 1772 individuals of 70 species and 6 fami- lies of necrophilous beetles were collected in January to December 2013. Three families, i.e., the Staphylinidae, Leiodidae, and Silphidae, were dominant in terms of individual abundances. The community similarity of necrophilous beetles was 36.81% (Wainstein’s similarity index, Kw) be- tween the plantation and the natural forest. Average monthly temperatures were significantly lower in the natural forest than on the plantation, while monthly individual abundances were higher in the natural forest. Species richness, individual abundances, and the Shannon-Wiener index were significantly correlated with the average monthly temperature in a quadratic relationship. The high- er the ambient temperature was, the higher were the species richness, individual abundance, and Shannon-Wiener index of the necrophilous beetle community. In addition, the quadratic polyno- mial curve of the natural forest was shifted in the direction of lower temperatures compared to that of the plantation. These results indicate that the ambient temperature had a significant impact on the community structure of necrophilous beetles in the cloud forest at mid-elevation Chilanshan.
Key words: necrophilous beetles, biodiversity, temperature, forest, Chilanshan.
Hwang W, Lai WH, Koh CN. 2014. The effect of ambient temperature on the biodiversity of necro- philous beetles at Chilanshan, northeastern Taiwan. Taiwan J For Sci 29(Supplement):S55- 64.
緒 言
動物屍體的分解過程中,不同分解者會在 不同的腐敗期前來(Payne 1965, Catts and Goff 1992),而其相關的捕食者亦會出現,使屍體如 同微生態系統,質能營養流動於各營養層的物 種間。屍體上出現的類群主要為雙翅目、鞘翅 目與膜翅目昆蟲(Payne 1965, Johnson 1975),
雙翅目雖然經常是最早到的類群(Bourel et al.
2003),但物種數較少,幼蟲個體數較多,膜 翅目以蟻科為主,小型屍體常有單一蟻窩獨佔 的 現 象(Hwang 2011),鞘翅目則多樣性最高
(Hwang and Shiao 2012),除了屍食性物種外 (Hanski 1983, Boonrotpong et al. 2004, Hwang and Shiao 2008),尚有蕈食者(Chandler and Peck 1992)與捕食者(Peschke and Fuldner 1977, Klimaszewski 1984, Hu and Frank 1997),可架 構出一個以屍體為生產者的食物網。屍體存在 的時間經常受所在棲地的環境溫度影響,環境 溫度越高,蠅蛆發育越快,常使屍體分解速率 加快(Swift et al. 1979, Nishida 1984),縮短屍 體微生態系統的存續時間。
不 同 緯 度 或 海 拔 的 環 境 氣 候 , 甚 至 是 不 同 的 植 被 結 構 , 都 會 影 響 環 境 溫 度 的 季 節 性 變化,進而影響分解速率和屍體微生態系統的 時效性(Lundt 1964, Nabaglo 1973, Swift et al.
1979)。環境溫度的高低,也會影響屍體上物 種間的競爭壓力(Sun et al. 2014),在競爭排斥 下,不同物種有各自出現的季節性(Hwang and Koh 2013a),從而影響群聚結構。在高溫環境 下,屍體分解速率快,可利用的時間較短,競 爭力強的物種即可能佔有優勢,使其他物種沒 有 機 會 獲 得 資 源 而 消 失 , 例 如 熱 帶 屍 體 上 昆 蟲 群 聚 的 低 多 樣 性 , 過 低 的 氣 溫 條 件(如北台 灣 的 冬 季),亦僅有少數耐寒性的物種可以活 動 , 其 群 聚 多 樣 性 亦 低 , 然 而 在 適 溫 的 環 境 中,屍體被使用的時效較長,昆蟲演替使利用 屍體的物種增加,群聚即有較高的多樣性,例 如北台灣夏季親屍性甲蟲高多樣性(Hwang and Shiao 2011, Hwang and Shiao 2012, Novak et al. 2014)。有鑑於過低或過高的環境溫度下,
親屍性甲蟲多樣性皆低,而在中間適溫,則具 有高多樣性,可歸納出親屍性甲蟲的物種多樣 性對應環境溫度的變化,可能會以常態分佈曲 顯呈現兩者的關係。親屍性甲蟲在屍體上所形 成的食物網結構完整,其物種多樣性較其他類 群高,各物種對環境敏感度的差異也大,在進 行環境溫度變遷監測時,其物種組成的結構變 化可作為環境衝擊的指標(Halffter and Favila 1993, Hwang and Koh 2013b)。
環 境 變 遷 對 親 屍 性 甲 蟲 群 聚 結 構 應 有 不 同 程 度 的 衝 擊(Hanski and Cambefort 1991, Martin-Piera and Lobo 1993)。在以往的研究 中,台灣南北部低海拔山區的親屍性甲蟲群聚 結構已有初步的了解,例如南北台灣緯度差異 下,福山與南仁山的親屍性甲蟲存有顯著的物 種組成與多樣性差異(Hwang and Shiao 2012),
在 不 同 棲 地 環 境 中 , 如 北 台 灣 福 山 林 地 與 草 地,除了有顯著的物種組成與多樣性差異外,
當環境溫度上升,親屍性甲蟲組成結構在草地 所面臨的衝擊則較林地為大,推論原因在於草 地總生物量較林地為低,熱蓄積能力亦較低,
熱量透過草地生物量緩衝的效率較林地差,氣
溫變化較為劇烈,升溫亦較林地為快,此影響 屍體的分解速率,進而使物種組成與多樣性受 到較大的衝擊(Hwang and Koh 2013b)。因此了 解不同緯度與海拔的親屍性甲蟲組成,未來可 以建立環境變遷對緯度與海拔二維的生態衝擊 模式,而林相的差異與生態熱容穩定性亦應考 慮在影響的因子內。棲蘭山位在台灣中海拔山 區霧林帶,本研究即調查棲蘭山親屍性甲蟲的 群聚結構,補充二維生態衝擊的基礎資料,並 比較人工林與天然林棲地在環境溫度變化時,
對物種豐度、豐量與多樣性的影響。
材 料 與 方 法
一、研究地點
地點位於台灣宜蘭縣大同鄉棲蘭神木園區 至鴛鴦湖一帶(24°36’N, 121°29’E),海拔高度 約在1200-1900 m,沿林道設置兩類棲地,分別 為100線林道兩旁天然下種林類型棲地(簡稱人 工林)以及130線林道自然林類型棲地(簡稱天然 林)。沿100線林道人工林棲地間隔1 km以上設 置一個樣點,而在130線林道天然林棲地,因範 圍較小,故至少間隔200 m以上設置一個樣點。
兩類型棲地分別設置8個樣點,每個樣點設置一 個懸掛式陷阱(hang-fall trap)。
二、陷阱設計
因 人 工 林 與 天 然 林 林 底 經 常 樹 根 盤 錯 , 難 以 於 地 面 設 置 陷 阱 捕 捉 甲 蟲 , 故 以Hwang and Koh (2013a, b)的地面型掉落式陷阱(Pitfall trap)為模式,改造成懸掛型掉落式陷阱(hang- fall trap)。懸掛型掉落式陷阱以45×45×0.3 cm 的壓克力板做為昆蟲降落平台,表面噴以綠漆 增加摩擦力,平台中央鑿有一直徑15 cm的圓 孔,圓孔下方設置一個深21 cm的漏斗,上緣 直徑約19 cm包覆平台圓孔,下緣出口端內徑 2 cm,插入500 ml的集蟲罐中,罐底鑽有直徑 約1 mm的小孔做為排水孔。為避免其他腐食 性脊椎動物取食誘餌,平台圓孔上方安置一個 16×16×8 cm的下開鐵籠,網目為2×2 cm,鐵 籠和平台間以鐵絲固定,並於鐵籠下方、漏斗
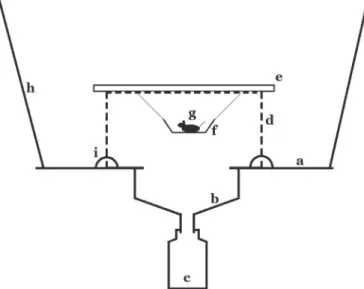
圓孔正上方懸掛一個免洗碗,碗內放置鼠屍做 為誘餌。鐵籠上方置有一塊16×16×0.3 cm的壓 克力板做為擋雨之用。平台四角鑿有小孔綁以繩 索,懸掛於樹枝上,陷阱底緣離地至少1 m以上 (Fig. 1)。
由2013年1月至12月,每月下旬於每個樣 點陷阱中各放置一隻18-22 g的新鮮解凍之ICR 品系小鼠鼠屍,於第4天收集集蟲瓶中的甲蟲。
誘 餌 集 蟲 期 間 , 在 兩 條 林 道 的 頭 、 中 、 尾 陷 阱處,各放置一個溫度記錄器(HOBO UA002 型),每30 min記錄一次環境溫度。
三、統計分析
所有採集到的甲蟲皆分科鑑定至種或形態 種,統計兩種林相親屍性甲蟲的物種豐度與豐 量,並援用Hwang and Koh (2013b)的分析方 法,每月的物種多樣性以Shannon-Wiener Index 呈現,使用Wilcoxon signed-ranks test (WSRT) 比較兩棲地類型物種豐度、豐量與多樣性,以 Wainstein的相似性指標(Wainstein’s Similarity I n d e x , Kw) 比 較 兩 種 林 相 親 屍 性 甲 蟲 的 物 種 組成結構相似性,並以Pearson’s correlation coefficient分析物種豐度、豐量與多樣性各別與
環境溫度二次多項式的相關性,使用之統計軟 體為SPSS 20,p < 0.05視為顯著。
Shannon-Wiener Index (H’) = –Σ pi loge pi
Kw (0~100) = Re x J
Re(%)(Renkonen’s coefficient) = Σ min (pi1, pi2) J(Jaccard’s coefficient) = c x 100 / (S1 + S2 – c) pi:i物種於群聚中的相對豐量
pi1:i物種於物種組成1中的相對豐量 pi2:i物種於物種組成2中的相對豐量 c:兩物種組成共有的物種數目 S1:物種組成1之物種豐度 S2:物種組成2之物種豐度
結 果
2013年1-12月於棲蘭山人工林與天然林 研 究 期 間 , 除7月因颱風導致天然林無法調查 外,共捕獲親屍性甲蟲6科70種1772隻,其中 以隱翅蟲科(Staphylinidae)捕獲44種699隻為各 科中物種豐度與豐量最高的科別,其次為球蕈 蟲科(Leodidae) 7種佔相對豐量33.63%,埋葬 蟲科(Silphidae) 3種16.20%佔居豐量第三位,
而牙蟲科(Hydrophilidae) 7種8.18%、金龜子科
Fig. 1. Diagram of a baited hang-fall trap. a: 45×45×0.3 cm green acrylic plate, b: funnel, c:
500-ml plastic jar, d: 16×16×8-cm metal cage, e: transparent acrylic plate, f: bait bowl, g:
18-22 g mouse carcass, h: rope, i: iron wire.
(Scarabaeidae)的3種糞金龜1.92%與閻魔蟲科 (Histeridae) 6種0.62%三科的豐量較低,其他非 親屍性甲蟲科別全年捕獲數量皆僅個位數,並 不列入統計(Table 1)。以Wainstein的相似性指 標(Wainstein’s Similarity Index, Kw)比較所捕獲 的親屍性甲蟲兩地物種豐度、共有物種數與物 種的相對豐量,甲蟲的組成結構在人工林與天 然林具有36.81%的相似性。
置放誘餌期間,以每30 min所自動記錄的 環境溫度,統計得月均溫,在人工林月均溫顯 著高於天然林,月均溫相差1.52℃(WSRT: p = 0.003) (Fig. 2),但天然林在每月所捕獲的個體 豐量則顯著高於人工林(WSRT,p = 0.047),兩 種林相的物種豐度(WSRT,p = 0.123)與多樣性 (Shannon-Wiener Index, WSRT,p = 0.236)則無 顯著性差異。
Table 1. Numbers of species and individuals of necrophilous beetle families captured by baited hang-fall traps in the plantation and in the natural forest at Chilanshan in January to December 2013
Plantation Natural forest Total
Species Individuals Species Individuals Species Percent (%)
Histeraidae 3 4 5 7 6 0.62
Hydrophilidae 7 47 6 98 7 8.18
Leiodidae 7 309 7 287 7 33.63
Scarabaeidae 2 27 2 7 3 1.92
Silphidae 2 158 3 129 3 16.20
Staphylinidae 29 289 34 410 44 39.45
Fig. 2. Average monthly temperatures in the plantation (P) and in the natural forest (NF) at Chilanshan in January to December 2013. Data for July in the natural forest were not included because of a typhoon. Average monthly temperatures in the plantation were significantly higher than those in the natural forest (WSRT,p = 0.003).
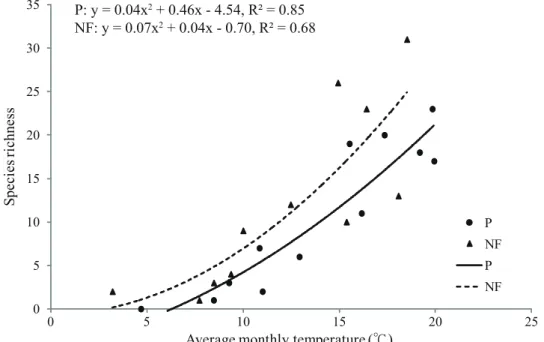
Fig. 3. Effects of average monthly temperatures on the species richness of necrophilous beetles collected in the plantation (P) and in the natural forest (NF) at Chilanshan in 2013.
There was a significant positive correlation between the species richness and average monthly temperatures (Pearson’s correlation coefficient for a quadratic relationship: p <
0.001 on the plantation and p = 0.010 in the natural forest).
以 二 次 多 項 式 分 析 每 月 捕 獲 的 甲 蟲 物 種 豐度與當月平均溫度發現,無論在人工林或是 天然林,皆顯示物種豐度與環境溫度有顯著的 相關性(Pearson’s correlation coefficient:人工 林,p < 0.001;天然林,p = 0.010),在棲蘭山 當環境溫度越高,可捕獲的親屍性甲蟲物種數 便越高,在較低溫的天然林,此二次多項式曲 線則往低溫方向偏移(Fig. 3)。
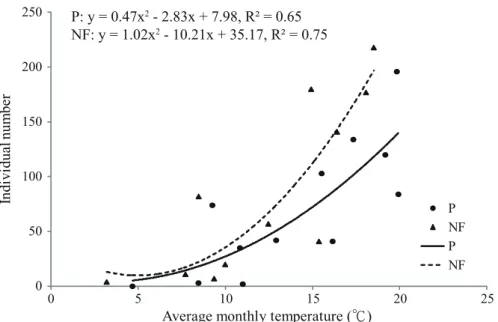
在人工林或天然林每月捕獲的甲蟲個體豐 量,皆與月平均溫度呈顯著的正相關(Pearson’s correlation coefficient:人工林,p = 0.009;天 然林,p = 0.004),結果顯示棲蘭山的環境溫度 越高,親屍性甲蟲個體出現的頻率便越高,二 次多項式曲線在較低溫的天然林亦往低溫方向 偏移(Fig. 4)。
月 平 均 溫 度 對 親 屍 性 甲 蟲 多 樣 性 的 影 響,無論在人工林或天然林亦有顯著的相關性 (Pearson’s correlation coefficient:人工林,p <
0.001;天然林,p = 0.016),隨著環境溫度升 高,多樣性指數亦增加,人工林約在18.5℃時 達到高峰,較低溫的天然林在低溫條件下具有 較高的多樣性(Fig. 5)。
討 論
在 探 討 全 球 暖 化 對 生 態 系 統 影 響 的 程 度 時 , 陸 域 生 物 在 海 拔 或 緯 度 間 的 移 動 , 經 常 拿 來 做 為 環 境 變 遷 對 生 態 系 統 衝 擊 的 證 據 (Konvicka et al. 2003, Chen et al. 2011)。以屍 體為微棲地的親屍性甲蟲,其物種多樣性高,
加上屍體分解速率深受環境溫度影響,在環境 溫 度 變 化 時 , 其 群 聚 結 構 也 會 跟 著 改 變 , 相 當 適 合 做 為 環 境 監 測 的 指 標 類 群(Hwang and Koh 2013b)。然而在不同的氣候條件下,親屍 性甲蟲的群聚結構並不相同(Hwang and Shiao 2012),因此若要建立環境變遷對緯度與海拔二
Fig. 4. Effects of average monthly temperatures on individual numbers of necrophilous beetles collected in the plantation (P) and in the natural forest (NF) at Chilanshan in 2013.
There was a significant positive correlation between individual numbers and average monthly temperatures (Pearson’s correlation coefficient for a quadratic relationship: p = 0.009 in the plantation and p = 0.004 in the natural forest).
Fig. 5. Effects of average monthly temperatures on the Shannon-Wiener index (Hs) of necrophilous beetles collected in the plantation (P) and in the natural forest (NF) at Chilanshan in 2013. There was a significant positive correlation between the Shannon- Wiener index and average monthly temperatures (Pearson’s correlation coefficient for a quadratic relationship: p < 0.001 in the plantation and p = 0.016 in the natural forest).
維的生態衝擊模式,則須選定在不同緯度與不 同海拔的氣候條件,先分析親屍性甲蟲的群聚 結構,再依環境溫度對其多樣性的影響,進行 長期監測。
與以往在宜蘭哈盆自然保留區及屏東南仁 山生態保護區的調查相互比較,在棲蘭山的親 屍性甲蟲,同樣是以隱翅蟲科與球蕈蟲科在個 體豐量上最為優勢;於南仁山不出現,且在哈 盆相對豐量不高的埋葬蟲科(2.18%),在棲蘭 山則有較高的捕獲量(16.20%);金龜子科的糞 金龜(1.92%)與牙蟲科(8.18%)兩類群的相對豐 量,亦較哈盆(糞金龜9.69%、牙蟲科11.31%) 與 南 仁 山(糞金龜25.34%、牙蟲科10.51%)低 (Hwang and Shiao 2012),這顯示中海拔霧林帶 棲蘭山的親屍性甲蟲與宜蘭哈盆及屏東南仁山 有相當大的差異,其具有獨特的群聚結構。
基 於 親 屍 性 甲 蟲 在 高 溫 環 境(如熱帶南仁 山、哈盆草地)與低溫環境(如冬季)皆有較低的 多 樣 性 , 適 溫 的 環 境 條 件 有 較 高 的 多 樣 性(如 哈盆林地春至秋季) (Hwang and Shiao 2012, Hwang and Koh 2013b),藉此可假設在較大尺 度的氣候差異下,親屍性甲蟲多樣性應依環境 溫度變異呈常態分布曲線。棲蘭山的海拔較哈 盆高,緯度與海拔亦比南仁山高,故棲蘭山12 個月份的溫度曲線皆較哈盆與南仁山低(Hwang 2006, Hwang and Koh 2013b)。若以較大尺度環 境溫度變異下的常態分布曲線來評估,棲蘭山 的氣候條件,應落在常態分布圖的左側部分,
其親屍性甲蟲群聚多樣性與環境溫度會存在正 相關的現象。本研究結果,無論是在人工林或 是天然林,以二次多項式分析物種豐度、個體 豐 量 和Shannon-Wiener多樣性指數三者和環 境溫度的關係,皆呈現顯著的正相關,隨著環 境溫度的增加,親屍性甲蟲群聚的多樣性亦增 高,表現出原假設常態分布曲線的左側部分。
植 物 的 物 種 豐 度 常 間 接 影 響 消 費 營 養 層 的物種多樣性(Chey et al. 1997, Scherber et al.
2010),也使動物屍體數量的可利用性因植物 相產生差異。同樣的,棲地的異質性越高也會 使上層消費者有較高的物種多樣性(Tews et al.
2004)。在相同緯度與海拔條件下,人工林與天
然林的植物相並不相同,人工林以檜木和柳杉 為主,樹齡相近,且林下無明顯的次冠層與灌木 層,天然林則為各種闊葉樹所組成,林下遍布灌 木與草本植物。以植物多樣性和異質性來看,天 然林皆較人工林為高,結果顯現在所捕獲昆蟲的 個體豐量上,天然林每月捕獲的個體豐量顯著高 於人工林。但物種豐度與多樣性指數在兩種林相 間,並無顯著差異,此則須考慮不同棲地環境溫 度變化幅度所帶來的影響,當棲地生物量高,可 緩衝環境溫度變化幅度,例如福山在年均溫升 高下,林地親屍性甲蟲多樣性變化的幅度則較草 地為低(Hwang and Koh 2013b)。人工林雖為植 被多樣性低的針葉林,但林木高度皆在10 m以 上,天然林林木高度約在10 m以下,在植物生 物量熱蓄積能力上,人工林則較天然林為優,面 對低溫環境,人工林有較佳的氣溫緩衝效果,此 外,人工林腹地較為平坦,天然林則位在較為傾 斜裸露的坡地,坡地與平地的地勢方位可能影響 熱對流與傳導,如本研究在相同的月份,天然林 的月均溫低於人工林,而使天然林的高植被多樣 性與異質性,可能被較低的環境溫度所抵銷。人 工林異質性低,但高生物量可緩衝環境低溫,而 天然林異質性高,但地勢與低生物量使環境具有 較嚴苛的低溫,此可能是親屍性甲蟲物種豐度與 多樣性指數在兩種林相間無顯著差異的原因。
棲 地 的 類 型 往 往 影 響 生 活 在 其 中 的 昆 蟲 群 聚 組 成 , 同 類 型 的 棲 地 間 , 也 具 有 較 相 似 的物種組成結構(Hughes et al. 2000),若探討 不同棲地間親屍性甲蟲群聚結構的相似性,比 較Hwang and Koh (2013b)在宜蘭哈盆自然保 留區的數據,林地和草地兩種不同類型的棲地 間,無疑是相當低的(3.95%, Kw),而同一林地 在相鄰兩個年度相互比較,則可高達61.7%的 相似度。棲蘭山人工林與天然林親屍性甲蟲群 聚結構的相似性為36.81%,此數值符合同為林 地但不同林相的範疇內。儘管兩種林相在六個 甲蟲科別間有許多相同的物種重疊,且每月的 Shannon-Wiener多樣性指數亦無法顯現兩者的 差異,但以Wainstein的相似性指標(Wainstein’s Similarity Index)仍能判別兩種林相親屍性甲蟲 群聚結構的不同。同樣的,天然林中親屍性甲
蟲物種豐度、個體豐量和Shannon-Wiener多樣 性指數的二次多項式曲線皆往較低溫的方向偏 移,且天然林在較低溫的條件下有較高的多樣 性指數,亦證明了天然林較低溫的環境存在著 與人工林不同的生態特性。
目前在台灣以相同的方法調查,且已可供 比較的生態環境有宜蘭哈盆、屏東南仁山,探 討環境變遷所造成的生態衝擊,在親屍性甲蟲 群聚部分,已呈現基本的模式(Hwang and Shiao 2012, Hwang and Koh 2013b)。在建立緯度與海 拔二維的生態基礎資料上,棲蘭山親屍性甲蟲 群聚的調查,補充了北台灣中海拔的數據,且 提供較大尺度的氣候模式下,親屍性甲蟲多樣 性常態分布曲線於低溫部分的趨勢。然而,較 低溫的環境是否對環境變遷有更高的敏感性,
或是當暖化時,是否多樣性會因為適應溫度範 圍較窄的狹溫性物種往較高緯度或較高海拔播 遷(Parry and Carter 1989, Chen et al. 2011)而提 高,還需未來以相同的方法進行監測比較,才 能了解環境變遷對棲蘭山生態的衝擊程度。
誌 謝
本文承蒙行政院農委會林業試驗所科技計 畫(編號:102農科-13.2.1-森-G1)經費補助,
以及退輔會榮民森林保育事業管理處之行政協 助,特此感謝。
引 用 文 獻
Boonrotpong S, Sotthibandhu S, Pholpunthin C. 2004. Species composition of dung beetles in the primary and secondary forests at Ton Nga Chang Wildlife Sanctuary. Sci Asia 30:59-65.
Bourel B, Callet B, Hedouin V, Gosset D.
2003. Flies eggs: a new method for the estima- tion of short-term post-mortem interval? Fo- rensic Sci Int 135:27-34.
Catts EP, Goff ML. 1992. Forensic entomol- ogy in criminal Investigations. Annu Rev En- tomol 37:253-72.
Chandler DS, Peck SB. 1992. Diversity and seasonality of leiodid beetles (Coleoptera:
Leiodidae) in an old-growth forest and a 40- year-old forest in New Hampshire. Environ Entomol 21:1283-93.
Chen IC, Hill JK, Ohlemüller R, Roy DB, Thomas CD. 2011. Rapid range shifts of spe- cies associated with high levels of climate warming. Science 333 (6045):1024-6.
Chey VK, Holloway JD, Speight MR. 1997.
Diversity of moths in forest plantations and natural forests in Sabah. B Entomol Res 87(4):371-85.
Halffter G, Favila ME. 1993. The Scarabaei- nae an animal group for analysing, inventory- ing and monitoring biodiversity in tropical rainforest and modified landscapes. Biol Int 27:15-21.
Hanski I. 1983. Distributional ecology and abundance of dung and carrion-feeding beetles (Scarabaeidae) in tropical rain forests in Sara- wak, Borneo. Acta Zool Fenn 167:1-45.
Hanski I, Cambefort Y. 1991. Ecology of dung beetles. Princeton, NJ: Princeton Univ.
Press. 481 p.
Hu GY, Frank JH. 1997. Predation on the horn fly (Diptera: Muscidae) by five species of Philonthus (Coleoptera: Staphylinidae). Envi- ron Entomol 26:1240-6.
Hughes JB, Daily GC, Ehrlich PR. 2000.
Conservation of insect diversity: a habitat ap- proach. Conserv Biol 14(6):1788-97.
Hwang W. 2006. Konkurrenz und Aasnutzung necrophager und necrophiler Käfer in Nord- und Sudtaiwan mit einem Beitrag zur Biologie von Nicrophorus nepalensis Hope (Coleoptera:
Silphidae) [PhD dissertation]. Freiburg, Ger- many: Albert-Ludwigs-Univ. 108 p.
Hwang W, Shiao SF. 2008. Use of carcass by burying beetles Nicrophorus nepalensis Hope (Coleoptera: Silphidae). Formosan Entomol 28:87-100.
Hwang W. 2011. Environmental variables
affecting community composition and carcass utilization of necrophagous ants (Hymenoptera:
Formicidae) in southern and northern Taiwan.
BioFormosa 46(1):21-31
Hwang W, Koh CN. 2013a. Distribution of three carrion beetles and population dynamics of Nicrophorus nepalensis (Coleoptera: Silphi- dae) in the Fushan area of Taiwan. BioFormosa 48:1-11.
Hwang W, Koh CN. 2013b. Application of necrophilous beetles to long-term monitoring of a forest ecosystem associated with climatic change. Taiwan J For Sci 28(2):83-96.
Hwang W, Shiao SF. 2011. Dormancy and the influence of photoperiod and temperature on sexual maturity in Nicrophorus nepalensis (Coleoptera: Silphidae). Insect Sci 18:225-33.
Hwang W, Shiao SF. 2012. Comparative study of species composition and seasonal activity of necrophilous beetles with emphasis on Apho- diinae and Scarabaeinae (Coleoptera: Scara- baeidae) in subtropical and tropical Taiwan.
Formosan Entomol 32:331-44.
Johnson MD. 1975. Seasonal and microseral variations in the insect population on carrion.
Am Midl Nat 93(1):79-90.
Klimaszewski J. 1984. A revision of the genus Aleochara Garvenhorst of America north of Mexico (Coleoptera: Staphylinidae: Aleochari- nae). Mem Entomol Soc Can 129:211.
Konvicka M, Maradova M, Benes J, Fric Z, Kepka P. 2003. Uphill shifts in distribution of butterflies in the Czech Republic: effects of changing climate detected on a regional scale.
Glob Ecol Biogeogr 12:403-10.
Lundt H. 1964. Ökologische Untersuchun- gen über die tierische Besiedlung von Aas im Boden. Pedobiologia 4:158-80.
Martin-Piera F, Lobo JM. 1993. Altitudi- nal distribution patterns of cropo-necrophage
Scarabaeidea in Veracruz, Mexico. Coleopts Bull 47:321-34.
Nabaglo L. 1973. Participation of inverte- brates in decomposition of rodent carcass in forest ecosystem. Ekol Polska 21:251-69.
Nishida K. 1984. Experimental studies on the estimation of postmortem intervals by means of fly larvae infesting human cadavers. Jpn J Legal Med 38:24-41.
Novak T, Šajna N, Antolinc E, Lipovšek S, Devetak D, Janžekovič F. 2014. Cold toler- ance in terrestrial invertebrates inhabiting sub- terranean habitats. Int J Speleol 43(3):265-72.
Parry ML, Carter TR. 1989. An assessment of the effects of climatic change on agriculture.
Climatic Change 15:95-116.
Payne JA. 1965. A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology 46:592-602.
Peschke K, Fuldner D. 1977. Übersicht und neue Untersuchungen zur Lebensweise der par- asitoiden Aleocharinae (Coleoptera: Staphylini- dae). Zool Jahrbucher Syst 104:242-62.
Scherber C, Heimann J, Köhler G, Mitschu- nas N, Weisser WW. 2010. Functional identity versus species richness: herbivory resistance in plant communities. Oecologia 163:707-17.
Sun SJ, Rubenstein DR, Chen BF, Chan SF, Liu JN, Liu M, et al. 2014. Climate-mediated cooperation promotes niche expansion in bury- ing beetles. eLife 3:e02440. DOI: 10.7554/
eLife.02440.
Swift MJ, Heal OW, Anderson JM. 1979.
Decomposition in terrestrial ecosystems. Lon- don: Blackwell Scientific. 372 p.
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F.
2004. Animal species diversity driven by habi- tat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79-92.