山村地景對鳥類多樣性的影響初探
—以金山兩湖社區為例
葛兆年1) 許詩涵1) 蘇聲欣2) 鍾智昕3) 王相華1,4)
摘 要
日本里山的鑲嵌地景具有高生物多樣性的維持功能,此一傳統土地管理模式近年來廣為生態學者 所推崇,並引介、推廣至許多國家,包括臺灣。本研究於臺灣北部金山區兩湖里山村的森林、森林邊 緣、鑲嵌棲地設立調查樣區,進行鳥類群聚調查,主要探討的主題為:(1)兩湖山村地景對鳥類多樣性 有何種影響?(2)不同功能群鳥類受到山村地景的相對影響為何?
鳥 類 群 聚 調 查 結 果 顯 示 , 純 森 林 棲 地 的 鳥 類 種 數 、 隻 次 都 比 森 林 邊 緣 及 鑲 嵌 棲 地 低 。 大 多 數 在森林地景出現的鳥種也同時出現在森林—農墾混合棲地(包含森林邊緣及鑲嵌棲地),但有許多鳥 種僅出現在森林—農墾混合棲地,例如喜歡在水域活動的黃頭鷺(Bubulcus ibis)、小白鷺(Egretta garzetta),喜食農墾地穀類的珠頸斑鳩(Streptopelia chinensis)、斑文鳥(Lonchura punctulata)、白腰文 鳥(Lonchura striata),以及外來入侵種白尾八哥(Acridotheres javanicus)。就個別功能群鳥類而言,森 林邊緣型、草生地型、水域型及開墾地型鳥類的鳥類種數、隻次在森林樣區均低於森林邊緣及鑲嵌棲 地樣區,但森林型、聚落型及多樣棲地型鳥類的鳥類種數、隻次在3類型樣區之間都沒有顯著差異。地 景對鳥類多樣性影響的分析結果指出,旱田面積、鑲嵌地景之邊緣長度、棲地多樣性和鳥類多樣性之 間呈現明顯的正相關,顯示兩湖山村現行農墾方式所創造的鑲嵌地景有利於增加鳥類多樣性。兩湖里 山村小面積的農墾作業所形塑的森林—農墾鑲嵌地景,具有開墾面積小、與周邊森林連結度很高(隔離 度低)的特性,可能是導致森林型鳥類及其它非森林型鳥類同時出現在兩湖里山村森林—農墾鑲嵌地景 的重要因素,然而較多非森林型鳥類出現在森林—農墾鑲嵌地景所顯現的生態影響仍有待探討。
關鍵詞:山村、里山、地景、鳥類、多樣性。
葛兆年、許詩涵、蘇聲欣、鍾智昕、王相華。2018。山村地景對鳥類多樣性的影響初探—以金山兩湖 社區為例。台灣林業科學33(4):319-32。
1)林 業 試 驗 所 森 林 保 護 組 ,10066台北市南海路53號 Division of Forest Protection, Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 10066, Taiwan.
2)林業試驗所森林經營組,10066台北市南海路53號 Division of Forest Management, Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 10066, Taiwan.
3 )宜 蘭 大 學 森 林 暨 自 然 資 源 學 系 。2 6 0 宜 蘭 市 神 農 路 一 段 1 號 D e p a r t m e n t o f F o r e s t r y a n d N a t u r a l Resource, Ilan Univ., No.1, Sec. 1, Shennong Rd., Yilan City, Yilan County 260, Taiwan.
4)通訊作者 Corresponding author, e-mail:[email protected]
2018年7月送審 2018年11月通過 Received July 2018, Accepted November 2018.
Research paper
A Preliminary Study of the Effects of a Mountain Village Landscape on Bird Diversity: a Case Study
of Lianghu Village, Jinshan District, Northern Taiwan
Chao-Nien Koh,1) Shih-Han Hsu,1) Sheng-Hsin Su,2) Chih-Hsin Chung,3) Hsiang-Hua Wang1,4)
【Summary】
The mosaic landscape of satoyama (a mountain village) in Japan has the function of maintaining high biodiversity. This traditional land management model has been widely promoted by ecologists in recent years and introduced to many countries including Taiwan. We surveyed bird communities in the forests, forest edges, and mosaic habitats in Lianghu Village, Jinshan District, northern Taiwan, and this study aimed to elucidate: (1) What are the effects of mountain village landscapes on bird diversity?
and (2) What are the effects of mountain village landscapes on different functional groups of birds?
Results showed that bird species richness and numbers of individuals in the forest habitats were lower than those in the mixed forest-agricultural habitats (including both the forest edge and mosaic habitats). Most of the bird species that appeared in the forest habitats also appeared in the mixed forest-agriculture habitats. However, many bird species only appeared in the forest-agricul- tural landscape, such as Bubulcus ibis and Egretta garzetta which prefer wetlands, and Streptopelia chinensis, Lonchura punctulata, and Lonchura striata which prefer to feed on grain in agricultural lands, as well as the invasive species Acridotheres javanicus. For birds in each functional group, the species richness and numbers of individuals of the forest-type, grassland-type, wetland-type, and farmland-type birds were lower in the forest habitat than in the forest-agricultural mosaic habi- tat. However, there was no significant difference in the species richness or numbers of individuals of forest-type, village-type, and diverse habitat-type birds between the forest habitat and forest- agricultural mosaic habitat. Results of the effects of landscape on bird diversity indicated that bird diversity was positively correlated with the area of uplands, the length of mosaic edges, and habitat diversity, so the current mosaic landscape created by the fine-scale farming system in Lianghu Vil- lage may have positive effects on bird species diversity. The fine-scale farming patches and high degrees of connectivity (low isolation) with surrounding forests in this forest-agriculture mosaic landscape may facilitate the co-occurrence of forest-type and non-forest-type bird species in this mountain village. Nevertheless, the ecological effects of the occurrence of some non-forest-type bird species in this forest-agriculture mosaic landscape still need to be further investigated.
Key words: mountain village, satoyama, landscape, bird, diversity.
Koh CN, Hsu SH, Su SH, Chung CH, Wang HH. 2018. A preliminary study of the effects of a mountain village landscape on bird diversity: a case study of Lianghu Village, Jinshan Dis- trict, northern Taiwan. Taiwan J For Sci 33(4):319-32.
緒 言
熱帶及亞熱帶森林在過去數十年來遭受嚴 重的人為擾動(human disturbance),生態學者 開始關注森林擾動對生物多樣性之影響,並針 對此一議題進行不同生物分類群(taxa)的相關研 究。許多研究指出,人為擾動明顯降低物種數 目(Turner 1996, Lewis 2006, Philpott 2008);
但也有部分研究指出,一些物種可以適應一定 程度的人為擾動(Luck and Daily 2003, Peh et al. 2005),甚至因人為擾動而增加其族群數量 (Ranganathan et al. 2008);Lawton等人(1998) 彙整熱帶森林棲地變更對生物多樣性影響的相 關 研 究 , 分 析 鳥 類 、 蝴 蝶 、 甲 蟲 、 螞 蟻 等8種 生物類群,發現不同生物類群受到棲地變更的 影響程度及變化方向都有很大差異,似乎沒有 任何分類群可當作是熱帶森林棲地變更對生物 多樣性影響的指標類群。因此他們建議要釐清 棲地變更對生物多樣性的影響,必須針對不同 分類群物種進行更大規模、更有系統的調查。
更有許多學者發現,同一分類群的不同功能群 (functional group)物種對不同人為擾動強度的反 應會有所不同(Lawton et al. 1998, Diekotter et al. 2008, Farwig et al. 2008)。
聯 合 國2 0 1 0 年 於 名 古 屋 召 開 的 生 物 多 樣 性 公 約 第 十 屆 締 約 方 大 會 , 提 出 里 山 倡 議 (Satoyama Initiative),呼籲世界維護人類與 自 然 之 間 多 年 來 相 互 作 用 形 成 的 山 村 地 景 (mountain village landscape)。相關研究也顯 示,日本里山的傳統土地管理模式所形成的森 林 — 農 墾 地 鑲 嵌 地 景 具 有 高 生 物 多 樣 性 的 維 持功能(Ohara and Ushimaru 2015),而地景的 異質化(heterogeneity)及連結度(connectivity) (Benton et al. 2003, Katoh et al. 2009)被認為 是維持日本山村生物多樣性的重要基礎,因此 日本山村的傳統農林土地經營模式近年來廣為 全世界許生態學者所推崇,並引介、推廣至許 多國家,包括臺灣。水田及其灌溉系統是維持 生物多樣性的優良棲地(Amono 2009, Kadoya 2009) ,尤其是在日本以傳統方式從事耕作、生 產的山村(Washitani 2001, Kobori and Primack 2003, Altieri 2004)。臺灣在日治時期於山村
地區推廣栽植水稻,並將山坡整地為梯田的型 式。但隨著人口老化、年輕人移居城市,多數 山村水稻田轉為栽植旱作或廢耕,現今只剩下 少許水梯田,多數梯田改種旱作,或於廢耕後 自 然 演 替 成 為 草 生 地 、 次 生 林 , 但 道 路 、 房 舍等建物的比例略有增加,也有少許竹林及果 園。上述土地利用形態的變更情形,不僅普遍 發生在小農生產模式的臺灣山村(耆老口述),
也發生在類似生產模式的日本里山地景(Katoh 2009)。
臺灣與日本均是氣候潮濕、多山的島國,
山區原本多為森林所覆蓋,山村居民因應生活 需求開闢農田、水圳等人工地景,造就了山村 的多樣化地景,但由於臺灣與日本在文化、產 業及自然環境上有所不同,山村的土地利用型 態及經營管理方式上也有所差異,可能對生物 的組成、多樣性有不同程度的影響。林業試驗 所於2014~2015年間於金山兩湖里之哺乳類野生 動物調查結果顯示,臺灣獼猴(Macaca cyclopis) 與山羌(Muntiacus reevesi micrurus)在森林樣 區的出現頻度顯著的高於農墾地樣區,但鼬獾 (Melogale moschata subaurantiaca)與白鼻心 (Paguma larvata taivana)在農墾地樣區的出現 率則顯著高於森林樣區,而麝香貓(Viverricula indica pallida)則僅出現農墾地樣區(Yi-Ming Chen, unpubl. data)。Lu and Chen (2006)於屏 東五溝村的研究結果發現,鳥類多樣性與地景 多樣性呈現正相關,為國內少有的調查資料;
但五溝位於屏東縣萬巒鄉,是以水圳棲地為特 色的社區,並非臺灣典型的山村社區,因此國 內目前並沒有針對山村地景對鳥類多樣性影響 的相關研究。鳥類在許多生態系是重要的捕食 者(predators),對小型脊椎動物、無脊椎動物 多樣性及生態功能的維持非常重要(Sekercioglu 2006),它同時也是主要的種子傳播者(seed dispersers),對植物傳播、森林更新有重要影響 (Howe and Miriti 2004)。本研究於金山兩湖里 山村不同地景組合區域設立調查樣區,進行鳥 類群聚調查,主要探討的主題為:(1)山村地景 對鳥類多樣性有何種影響?(2)不同功能群鳥類 受到山村地景的相對影響為何?
材 料 與 方 法
試驗地概述
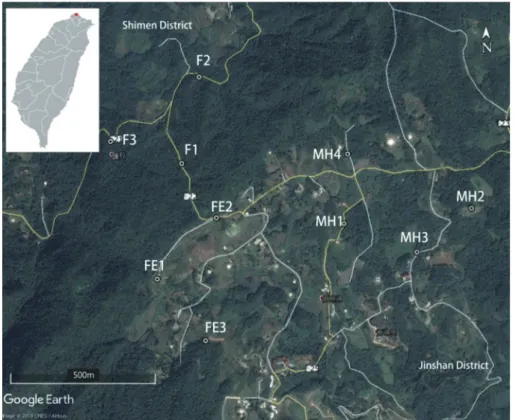
樣區設置於陽明山國家公園東北側,新北 市金山區及石門區交界處,行政區隸屬於新北 市金山區兩湖里(Fig. 1),緯度在北緯25°13′26″
至14′34″,經度在東經121°35′14″至59″之間,
海拔約為200至300 m。本區為森林、農墾地、
人 工 建 物 鑲 嵌 的 山 村 地 景 。 森 林 地 多 為 經 過 擾 動 後 的 次 生 林(secondary forest)或造林地 (plantation),農墾地多數是村民沿著山區森林 等高線開闢而成的梯田。依據在地村民口述,
早期許多梯田都是水田,但目前多數梯田已轉 變為以種植蔬菜與番薯為主之旱田或果園,僅 存少部分梯田維持水田形式,種植水稻、筊白 筍,而水田即使在休耕時仍會維持蓄水狀態。
但也有些梯田呈現廢耕狀態,逐步演替為草生
地或森林覆蓋,且隨著村落人口外移、老化,
廢耕地有逐漸增多的趨勢。
樣區設置及鳥類調查
因 板 塊 擠 壓 引 發 造 山 運 動 , 臺 灣 山 區 的 地形多崎嶇、陡峭,山村地區很少有大面積的 平坦地形,山村居民生活的核心區域多呈現森 林—農墾鑲嵌地景,週邊則多數維持森林的覆 蓋。本試驗參考Google Earth衛星影像,主觀 選擇3種棲地類型進行鳥類調查,包括(1)森林 棲地(forest, F1~F3):樣點周邊半徑50 m範圍 內皆為樹林覆蓋,(2)森林邊緣棲地(forest edge, FE1~FE3):在樣點半徑50 m範圍內,一側為森 林覆蓋,另一側是農墾地,(3)鑲嵌棲地(mosaic habitat, MH1~MH4):樣點周邊半徑50 m範圍 內之農墾地與樹林呈現塊狀鑲嵌狀態。上述森 林邊緣棲地及鑲嵌棲地可合併稱之為森林—農
Fig. 1. Map of the study area (the red asterisk in the inset map) and position of the sampling sites (F, forest plot; FE, forest edge plot; MH, mosaic habitat plot) in Lianghu Village, Jinshan Distinct, northern Taiwan.
墾混合棲地(forest-agricultural mixed habitat)。
本研究在兩湖村設置了3處森林樣點、3處森林 邊緣樣點、4處鑲嵌棲地樣點,合計設置10個樣 點,並將樣點為中心的50 m半徑範圍設定為鳥 類調查樣區。
鳥 類 調 查 於2014年6月至2015年5月間進 行,每個月進行1次調查,合計進行12次調查。
調查方式為選擇天氣良好的日子,於日出至日出 後3小時的時段內,至各調查樣區記錄看到、聽 到的鳥種與其數量;每個樣區的調查時間為10分 鐘,調查結束後再轉移至下一個樣區繼續進行調 查。完成10個樣區的調查,加上樣區間的移動時 間,合計約需要150分鐘。為降低人為觀察偏差 (observer bias),12次調查都由同1人執行;為降 低調查的系統性偏差(systematic bias),每次調查 時都隨機更換樣區的調查次序。
參考Fang et al. (2010)在研究湖山水庫地區 鳥類時,將各鳥種依主要棲地類型(habitat type) 分為7類功能群:(1)森林型( forest type):主要 棲地以森林為主者;(2)森林邊緣型(forest edge type):主要棲息於森林與其他類型棲地交界處;
(3)草生地型(grassland type):以草生地為主要棲 地者;(4)水域型(wetland type):主要棲息於溪流 及湖泊等棲地者;(5)聚落型(village type):以人 類聚落為主要棲息地者;(6)開墾地型(farmland type):主要棲息於果園、菜園及檳榔園等農耕地 者;(7)多樣棲地型(diverse habitat type):利用之 棲地不僅以1種為主,會利用多種棲地者。
地景及鳥類多樣性變數
以2014年1月25日之Google Earth衛星影 像為底圖,將地景區分為樹林、竹林、果園、
草生地、水域(水池、水田)、旱田、道路、房 舍共8種地景單元(landscape unit),使用QGIS 2.10.1地理資訊分析軟體,將以10個樣點為中 心,半徑50及100 m範圍內的地景進行數化處 理,數化後的資料,再依現場實際觀測情形進 行校對、修正。QGIS數化處理的內容包括,計 算各調查樣點周邊半徑50及100 m範圍內不同類 型地景單元之面積、不同地景單元交界處之邊 緣長度(length of edge),以及10個調查樣點(樣
區中心點)與水域、旱田、樹林及建物的最近距 離。此外,為瞭解地景多樣性對鳥類多樣性的 影響,我們採用Simpson及Shannon-Wiener 指 數計算各樣區的棲地多樣性(habitat diversity)。
其中Simpson 指數的計算式為D = 1-Σ(ni/ N)2, n i為 第 i種 地 景 單 元 的 面 積 , N為 所 有 地 景 單 元 的 面 積 ;Shannon-Wiener指數的計算式為 H =-Σ(Pi×lnPi),Pi為第i種地景單元的面積百 分 比 。 鳥 類 多 樣 性 除 了 計 算 各 樣 區 的 物 種 數 (species richness)外,也採用Simpson及Shannon- Wiener 指數計算各樣區的鳥類多樣性。
統計分析
本 研 究 將12個月的鳥調資料加總後進行 下述統計分析。採用Kruskal-Wallis單一等級 變異數分析(Kruskal-Wallis’ one-way ANOVA by ranks),檢測不同棲地在鳥類種數(species richness)及隻次(numbers of individuals)的差異 性是否顯著,採用Dunn多重比較測驗(Dunn’s test of multiple comparisons using rank sum),
檢 定 兩 兩 棲 地 類 型 在 各 功 能 群 鳥 類 種 數 及 隻 次 的 差 異 性 。 此 兩 種 測 驗 均 為 無 母 數 統 計 方 法 , 適 用 於 統 計 分 布 形 式 不 明 或 少 量 之 資 料 (Zar 2010)。鳥類群聚分析則同時使用群集分 析(cluster analysis; CA)及非計量多維標度分析 (non-metric multidimensional scaling; NMDS) 兩 種 多 變 數 分 析 法 , 藉 由 兩 者 互 補 的 方 式 來 探 索 群 聚 資 料 的 變 異 特 性 。 進 行 此 兩 種 分 析 之 前 , 需 先 求 算 樣 區 間 的 鳥 種 組 成 距 離 矩 陣 (distance matrix),本研究採用Bray-Curtis相異 性指數(dissimilarity index)進行計算;此外,
CA分析採用UPGMA (unweighted pair-group method using arithmetic averages)方法完成階 層式群集的聚合演算(Legendre and Legendre 2012)。NMDS的分析維度(dimensions)設定為 2,所得的2維分布序列結果(ordination)再以標 準化(standardization)後的地景及鳥類多樣性資 料進行envfit程式分析,計算出各地景及鳥類多 樣性變數與NMDS空間變化的相關性,並進行 置換排列檢定(permutation test) (Borcard et al.
2011, Oksanen et al. 2018)。最後再將NMDS與
envfit分析結果結合繪製出分布序列圖,以判釋 地景、多樣性因子與鳥類組成之關聯情形。同 時也進行皮爾遜相關(Pearson’s correlation)分 析,檢測地景變數與鳥類多樣性變數間之相關 性。以上統計分析工作皆以R 3.5.0統計語言(R Core Team 2018)完成,其中的多變數及多重比 較分析使用到vegan、dunn.test擴充套件(Dinno 2017, Oksanen et al. 2018)。
結 果
鳥類群聚組成特性
自2014年6月至2015年5月共進行12次調 查,於3類型棲地、10個樣區中共記錄到30科50 種1333隻次鳥類。其中,在3處森林樣區共記 錄到24種202隻次,在3處森林邊緣樣區共記錄 到41種462隻次,在4處鑲嵌棲地樣區共記錄到 38種669隻次(Table 1)。參照Fang et al. (2010) 的鳥類棲地類型分類方式,計有森林型鳥類20 種、森林邊緣型鳥類7種、草生地型鳥類8種、
水域型鳥類5種、聚落型鳥類2種、開墾地型鳥 類7種以及多樣棲地型鳥類1種。
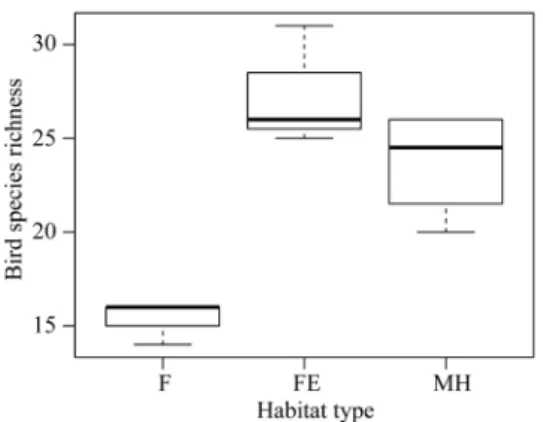
Kruskal-Wallis單一等級變異數分析結果 顯 示 ,3類型樣區的鳥類種數有顯著差異(p = 0.038)、鳥類隻次則有臨界顯著程度的差異(p =
0.055)。Dunn多重比較測驗結果顯示,森林樣區 的鳥類種數明顯低於森林邊緣樣區(p < 0.05),
鳥類隻次明顯低於鑲嵌棲地樣區;森林邊緣樣 區和鑲嵌棲地樣區在鳥類種數及隻次上沒有顯 著差異(Table 1, Figs. 2、3)。就個別功能群鳥 類而言,森林型鳥類、聚落型鳥類、多樣棲地 型鳥類的鳥類種數、隻次在3類型樣區之間都沒 有顯著差異,但森林邊緣型鳥類、草生地型鳥 類、水域型鳥類及開墾地型鳥類的鳥類種數、
隻次在森林樣區均都有低於森林邊緣樣區、鑲 嵌 棲 地 樣 區 的 趨 勢 , 但 在 統 計 檢 測 上 有 些 顯 著、有些不顯著(Table 1)。CA分析結果顯示,
3處森林樣區集中在樹狀圖下方,彼此間的相異 性較高,3處森林邊緣樣區及4處鑲嵌棲地樣區 則散布在樹狀圖的中央及上方,彼此的相異性 較低(Fig. 4);這顯示森林樣區和森林—農墾混 合樣區(森林邊緣樣區及鑲嵌棲地樣區)在鳥類群 聚的組成上有所區隔。NMDS分析結果亦顯示,
3處森林樣區分佈在第一軸的左側空間,3處森 林邊緣樣區及4處鑲嵌棲地樣區則分布在中央及 右側空間(Fig. 5),同樣顯示森林樣區和森林—
農墾混合樣區在鳥類群聚的組成上有所區隔。
將 鳥 種 依 據N M D S 第 一 軸 的 數 值 由 大 至 小 排 序 , 附 加 鳥 種 的 棲 地 類 型 屬 性 及 在 各 類 型 樣 區 的 記 錄 隻 次 , 整 理 成Ta b l e 2 。 表 Table 1. Species richness and numbers of individuals of birds in the 7 habitat types
(according to Fang et al. 2010) recorded in the forest (F), forest edge (FE), and mosaic habitat (MH) plots in Lianghu Village, Jinshan District, northern Taiwan. Values with different superscripts in the same row and item (species richness/numbers of individuals) denote significant differences at p < 0.05 (two-sided Dunn's test)
Habitat type Species richness Numbers of individuals
of birds Total Mean Total Mean
F(3) FE(3) MH(4)* F(3) FE(3) MH(4) F(3) FE(3) MH(4) F(3) FE(3) MH(4) Forest 17 16 14 12.0a 11.7a 11.8a 169 183 295 56.3a 61.0a 73.8a Forest edge 3 6 6 1.7a 3.7ab 4.3b 12 37 75 4.0a 12.3ab 18.8b Grassland 0 5 8 0.0a 3.7b 3.0b 0 48 29 0.0a 16.0b 7.3ab Wetland 0 5 1 0.0a 2.7b 0.5ab 0 11 7 0.0a 3.7b 1.8ab Village 2 1 2 0.7a 0.3a 0.5a 3 2 4 1.0a 0.7a 1.0a Farmland 1 7 6 0.7a 4.7b 3.0ab 16 179 236 5.3a 59.7b 59.0b Diverse habitat 1 1 1 0.3a 0.7a 0.8a 2 2 23 0.7a 0.7a 5.8a All species 24 41 38 15.3a 27.3b 23.8ab 202 462 669 67.3a 154.0ab 167.3b
* Numbers in parentheses denote the numbers of plots in the forests, forest edges, and mosaic habitats.
Fig. 3. Box-plot for comparing numbers of bird individuals among plots of the 3 habitat types in Lianghu Village, Jinshan Distinct, northern Taiwan. F, forest plot; FE, forest edge plot; MH, mosaic habitat plot.
Fig. 2. Box-plot for comparing bird species richness levels among plots of the 3 habitat types in Lianghu Village, Jinshan Distinct, northern Taiwan. F, forest plot; FE, forest edge plot; MH, mosaic habitat plot.
Fig. 4. Dendrogram of the 10 sampling sites in 3 habitat types in Lianghu Village, Jinshan Distinct, northern Taiwan. Here we used the Bray-Curtis dissimilarity index and UPGMA method to perform the cluster analysis. F, forest plot; FE, forest edge plot; MH, mosaic habitat plot.
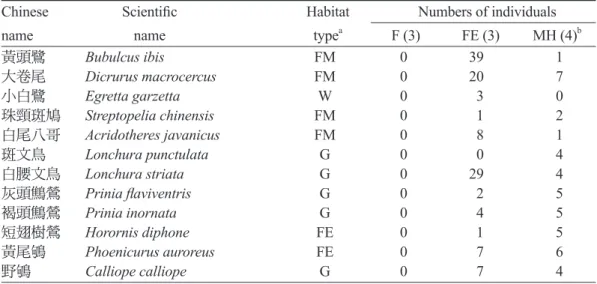
上 側 的 鳥 種 為 僅 出 現 在 森 林 — 農 墾 混 合 樣 區 ( 森 林 邊 緣 及 鑲 嵌 棲 地 樣 區 ) 的 鳥 種 , 都 屬 於 Fang et al. (2010)所區分的非森林型鳥類,包 括 黃 頭 鷺(Bubulcus ibis)、大卷尾(Dicrurus macrocercus)、小白鷺(Egretta garzetta)、
珠頸斑鳩(Streptopelia chinensis)、白尾八哥 (Acridotheres javanicus)、斑文鳥(Lonchura
punctulata)、白腰文鳥(Lonchura striata)、灰 頭鷦鶯(Prinia flaviventris)、褐頭鷦鶯(Prinia inornata)、短翅樹鶯(Cettia canturians)、黃 尾鴝(Phoenicurus auroreus)、野鴝(Calliope calliope)、紅尾伯勞(Lanius cristatus)、黑臉
(Emberiza spodocephala)、中杜鵑(Cuculus saturatus)、灰鶺鴒(Motacilla cinerea);但Table 2下側雖然多屬於Fang et al. (2010)所區分的森 林型鳥類,但並非僅出現在純森林樣區,也同 時出現在森林邊緣及鑲嵌棲地樣區。
地景、鳥類多樣性與群聚的關聯性
將 樣 區 的 地 景 、 鳥 類 多 樣 性 變 數 套 疊 於NMDS的分布序列圖中(Fig. 5),有8項因 子 與NMDS空間變化呈現明顯的相關性(r2 = 0.56~0.83,p < 0.05)。其中物種數(Spp)及調查 樣點周邊半徑100 m範圍內之Shannon棲地多樣 性指數(Hab.shannon100m)、Simpson棲地多樣性 指數(Hab.simpson100m)、邊緣長度(L.edge100m)、
旱田面積(A.upland100m)位於第一軸的右側,與 森林—農墾地鑲嵌地景的樣區同側;與水域距 離(D.wetland)、旱田距離(D.upland)及房舍距 離(D.build)則偏向第一軸的左側,與純森林地 景的樣區同側。這表示森林樣區與農墾地常見 的旱田、水塘、房舍距離較遠,棲地多樣性較 低,所記錄到的鳥類種數也較低;相對的,森
林—農墾地混合樣區的棲地多樣性較高、不同 棲地交界處的邊緣長度較長、旱田開墾面積較 大,其所記錄的鳥類種數也較高。皮爾遜相關 分析結果顯示,調查樣點周邊半徑50、100 m 範圍內之樹林面積(A.wood50m, A.wood100m)與鳥 類種數及Shannon物種多樣性指數呈現明顯的 負相關(p < 0.05) (Table 3)。調查樣點周邊半徑
100 m範圍內旱田面積(A.upland100m)與鳥類種數 及Shannon物種多樣性指數呈現明顯正相關(p <
0.05);相同的,調查樣點周邊半徑50、100 m 範圍內之鑲嵌地景交界處之邊緣長度(L.edge50m
, L.edge100m)以及Simpson、Shannon棲地多樣 性指數(Hab.simpson50m, Hab.shannon50m, Hab.
simpson100m, Hab.shannon100m)也與鳥類種數及 Fig. 5. Biplot of non-metric multidimensional scaling (NMDS) ordination of sampling sites (F, forest plot; FE, forest edge plot; MH, mosaic habitat plot) and bird species (open circles), with arrows indicating the projected landscape and diversity variables using the
“envfit” program (Oksanen et al. 2018). The stress value of NMDS was 0.079. Abbreviations of landscape variables: D.wetland, distance to wetlands; D.build, distance to buildings;
D.upland, distance to uplands; A.upland, area of uplands; L.edge, total length of habitat patch edges; Hab.simpson, Simpson’s habitat diversity; Hab.shannon, Shannon’s habitat diversity; Spp, species richness; and the subscript “100m” denotes the area within a 100-m radius of a sampling point.
Table 2. Habitat type and numbers of individuals of each bird species recorded in the forest (F), forest edge (FE), and mosaic habitat (MH) plots in Lianghu Village, Jinshan District, northern Taiwan. Bird species are ranked decreasingly by NMDS axis 1 scores. Species with fewer than 3 individuals and only recorded in a single plot were excluded from this table Chinese Scientific Habitat Numbers of individuals
name name typea F (3) FE (3) MH (4)b
黃頭鷺 Bubulcus ibis FM 0 39 1
大卷尾 Dicrurus macrocercus FM 0 20 7
小白鷺 Egretta garzetta W 0 3 0
珠頸斑鳩 Streptopelia chinensis FM 0 1 2
白尾八哥 Acridotheres javanicus FM 0 8 1
斑文鳥 Lonchura punctulata G 0 0 4
白腰文鳥 Lonchura striata G 0 29 4
灰頭鷦鶯 Prinia flaviventris G 0 2 5
褐頭鷦鶯 Prinia inornata G 0 4 5
短翅樹鶯 Horornis diphone FE 0 1 5
黃尾鴝 Phoenicurus auroreus FE 0 7 6
野鴝 Calliope calliope G 0 7 4
Shannon物種多樣性指數呈現明顯正相關(p <
0.05) (Table 3)。
討 論
調查資料顯示,樣區記錄到的50鳥種中約 有1/3屬於遷徙性候鳥,如資料許可,理應將調 查資料細分為不同季節予以分析,但受限於樣 區數量及調查頻率,本研究所取得的調查資料相 對有限,故僅將整年的調查資料合併後進行初步 的分析、探討,並未論及季節性的差異,亦或繁 殖季與非凡繁殖季的差異。然而,許多鳥類有遷 徙特性,故建議後續進行類似研究時,應適度加 強現場調查強度,所累積的資料方足以進行更深 入、完整的分析。此外,本研究於樣區類型選擇 時將森林—農墾混合地景區分成森林邊緣棲地及 鑲嵌棲地兩種類型,但在資料分析時發現,從地 景單元之比例看不出兩者的差異,因此在無母數 及多變量統計分析結果中,兩類型棲地的鳥類多 樣性均未能檢驗出明顯差異。
山村地景與鳥類多樣性
本研究發現,縱使沒有大面積水田,兩湖
山村的森林—農墾混合棲地(森林邊緣及鑲嵌棲 地 樣 區)鳥類多樣性仍然顯著高於森林棲地,
NMDS分析及相關分析的結果亦顯示,棲地多 樣性與鳥類多樣性呈現明顯的正相關,表示兩 湖山村的小規模塊狀農墾方式所創造的鑲嵌地 景 有 利 於 增 加 鳥 類 多 樣 性 。 國 外 許 多 研 究 顯 示,森林在經歷大面積、高強度的伐木、農墾 作業後,會對生物多樣性造成明顯的負面影響 (Moran et al. 2004, Kirika et al. 2008),但這情形 似乎並未在日本里山及本研究金山區兩湖里的 山村地景發生。基本上,日本傳統山村盛行小 面積的農墾及森林作業,創造農墾地與非農墾 地鑲嵌地景(mosaic landscape);相較於近代大 面積伐木後採行施用化學肥料、農藥的農墾作 業方式,屬於相對輕微的人為擾動(Kadoya and Washitani 2011)。棲地破碎化對生物多樣性所產 生的影響取決於擾動區塊的面積(patch size)及擾 動區塊的隔離度(isolation) (Andren 1994, Katoh et al. 2009),兩湖里山村盛行小面積的農墾作 業,形塑的森林—農墾鑲嵌地景與周邊未開墾 區森林的連結度很高(隔離度低),可能是導致鳥 類多樣性在森林—農墾混合樣區高於森林樣區 的重要因素。
近 年 來 , 許 多 探 討 地 景 變 化 對 生 物 多 樣 性 影 響 之 研 究 會 擴 及 不 同 尺 度 的 生 物 多 樣 性 , 多 數 國 外 學 者 採 用 分 割 多 樣 性(diversity p a r t i t i o n i n g ) 方 式 分 析 生 物 多 樣 性 在 樣 區 (plot)、棲地(habitat)、區域(region)等不同尺 度(scale)上的變化(Clough et al 2007, Cabra- Garcia et al 2010, Neuschulz et al 2011, Pacheco and Vasconcelos 2012, Carrara et al 2015),即把γ diversity分割成α及β兩種層面來 分析。Carrara等(2015)研究發現,與未砍伐的 森林比較,林地砍伐造成的破碎化森林並未降 低鳥類α多樣性,但會明顯損及鳥類β多樣性;
Neuschulz等(2011)在不同類型森林擾動棲地的
鳥類多樣性研究指出,β多樣性占整體多樣性的 比例明顯高於α多樣性,Clough等(2007)針對植 物及昆蟲多樣性的研究也發現同樣的趨勢。現 今,生物多樣性保育學者已有共識,β多樣性在 大區域層級的生物保育上非常重要,也是中、
大尺度生物多樣性保育計畫必須顧及的重點,
以及檢視保育計畫成功與否的關鍵因子(Socolar et al 2016)。本研究僅分析α多樣性在兩湖社區 3類型棲地(森林樣區、森林邊緣樣區、破碎森 林樣區)的差異,以及哪些地景單元及環境因子 會 對α多樣性造成影響,但如要深入了解臺灣 山村地景對生物多樣性的影響,需要有更大空 間尺度的研究,涵蓋多個山村社區及其周邊森 con’t
白頭翁 Pycnonotus sinensis FM 16 107 222
紅尾伯勞 Lanius cristatus FM 0 3 3
樹鵲 Dendrocitta formosae F 13 31 40
紅嘴黑鵯 Hypsipetes leucocephalus FE 9 23 49
大冠鷲 Spilornis cheela FE 1 2 1
黑臉 鵐 Emberiza spodocephala G 0 6 5
中杜鵑 Cuculus saturatus F 0 1 6
黑枕藍鶲 Hypothymis azurea FE 2 3 13
綠繡眼 Zosterops japonicus F 22 39 56
臺灣藍鵲 Urocissa caerulea F 5 9 0
金背鳩 Streptopelia orientalis D 2 2 23
灰鶺鴒 Motacilla cinerea W 0 4 7
洋燕 Hirundo tahitica V 1 2 1
黃眉柳鶯 Phylloscopus inornatus F 4 6 10
山紅頭 Cyanoderma ruficeps F 7 8 16
白腹鶇 Turdus pallidus F 5 2 10
小彎嘴 Pomatorhinus musicus F 54 50 80
五色鳥 Psilopogon nuchalis F 15 12 33
大彎嘴 Megapomatorhinus erythrocnemis F 11 8 13
竹雞 Bambusicola thoracicus F 9 9 11
小雨燕 Apus nipalensis V 2 0 3
樹鷚 Anthus hodgsoni F 2 3 8
赤腹鶇 Turdus chrysolaus F 2 1 1
繡眼畫眉 Alcippe morrisonia F 14 1 10
鳳頭蒼鷹 Accipiter trivirgatus F 1 0 1
a Habitat type of birds: F, forest type; FE, forest edge type; G, grassland type; W, wetland type; V, vil- lage type; FM, farmland type; D, diverse habitat type.
b Numbers in parentheses denote the number of plots in the forests, forest edges, and mosaic habitats.
林,方可擴及β及γ多樣性層面的分析。本研究 發現山村的小規模擾動會增加α多樣性,但α多 樣性增加導因於出現許多干擾地常見鳥種,若 將調查範圍擴增至周邊區域的數個不同山村,
有可能會出現α多樣性增加,但β、γ多樣性持 平或降低的情形,此部分尚有待進一步的研究 予以釐清。
不同功能群鳥類對地景改變的適應性
Clough等人(2009)研究森林轉變成可可 樹混農林生產地(cocoa agroforestry)對於鳥類 多 樣 性 之 影 響 時 , 將 鳥 類 依 食 性(diet)、棲地 (habitat)、植生層次(stratum)、重量(weight)區 分為不同的功能群,發現棲地轉變對不同功能 群鳥類的影響有很大差異。例如就食性而言,
食果型(frugivores)鳥類種數隨著樣區與森林距 離的增加而遞減,食穀型(granivores)鳥類則隨 著樣區與森林距離的增加而遞增;就棲地喜好 而言,森林型(forest specialists)鳥類種數隨著 樣區與森林距離的增加而遞減,廣域型(habitat generalists)鳥類則隨著樣區與森林距離的增加 而遞增。許多其他研究也發現,人為擾動對不
同棲地喜好的鳥類會產生不同的影響,森林型 鳥類對人為擾動的影響較敏感,在擾動區域會 轉變為以廣域型鳥類為主(generalist species) (Bender et al. 1998, Peh et al. 2005, Farwig et al.
2008)。Neuschulz等人(2011)探討森林棲地改變 對鳥類群聚影響,依據主要棲地類型將鳥種區 分為森林型、森林廣域型(forest generalists)、灌 木型(shrubland species)、開闊型(open country species)4類功能群,也同樣發現不同棲地功能群 鳥類對森林棲地改變的反映有很大差異。
本 研 究 比 較 兩 湖 山 村 森 林 — 農 墾 鑲 嵌 地 景與周邊森林的鳥類群聚差異,結果顯示,森 林型鳥類的物種數、隻次在上述兩類型棲地間 沒有顯著差異,但部分森林型以外的鳥類物種 數、隻次在森林—農墾鑲嵌地景顯著高於純森 林 地 景 , 例 如 森 林 邊 緣 型 、 草 生 地 型 、 水 域 型及開墾地型的鳥類,不論是在森林邊緣棲地 或鑲嵌棲地都有增加的趨勢,顯示森林—農墾 鑲嵌地景增加了鑲嵌棲地邊緣長度及草生地、
水域、開墾地的棲地比例,故可以提升以上述 類型鳥類之種類及隻次。反觀森林型鳥類,其 物種數及隻次在純森林地景與森林—農墾鑲嵌 Table 3. Pearson’s correlation coefficients between landscape variables and bird diversity variables. Only landscape variables significantly correlated with both bird diversity variables are listed
Bird diversity variable
Landscape variablea Species richness Shannon diversity index
r p r p
A.wood50m -0.759 0.011 -0.835 0.003
A.wood100m -0.761 0.011 -0.781 0.008
A.upland100m 0.696 0.025 0.725 0.018
L.edge50m 0.775 0.009 0.755 0.012
L.edge100m 0.956 0.000 0.724 0.018
Hab.simpson50m 0.825 0.003 0.829 0.003
Hab.simpson100m 0.943 0.000 0.769 0.009
Hab.shannon50m 0.858 0.002 0.802 0.005
Hab.shannon100m 0.965 0.000 0.770 0.009
a Abbreviations of landscape variables: A.wood, area of woods; A.upland, area of uplands; L.edge, total length of habitat patch edges; Hab.simpson, Simpson’s habitat diversity; Hab.shannon, Shan- non’s habitat diversity; and the subscripts “50m” and “100m” denote area within 50-m and 100-m radii of sampling points, respectively.
地景之間沒有差異,代表並沒有受到農墾地與 森林鑲嵌的影響。Carrara等人(2015)的研究發 現,森林棲地減少會導致森林型鳥類多樣性的 明顯減少,但此一現象似乎並未在兩湖山村出 現,應該與此地開墾面積不大,森林雖被切割 破碎但不至於完全孤立隔離有關。簡而言之,
兩湖山村森林—農墾鑲嵌地景同時提供了森林 型、非森林型鳥類的棲地需求,但周邊的純森 林地景主要提供森林型鳥類所需要的棲地,只 有相對少量的非森林型鳥類被記錄到。然而,
較多的非森林型鳥類在森林—農墾鑲嵌地景被 記 錄 到 , 是 否 全 然 顯 現 正 向 效 益 , 仍 有 待 斟 酌、探討。例如,入侵種白尾八哥的出現、聚 集,長期而言可能會與本地鳥種競爭,帶來負 面 的 生 態 影 響 ; 另 依 據 當 地 耆 老 口 述 , 有 一 些水鳥如磯鷸(Actitus hypoleucos)、白腰草鷸 (Tringa ochropus)等並非長期穩定的出現,只是 偶發性的出現在兩湖山村所剩不多的水田中;
此外,出現在兩湖森林—農墾地景的許多非森 林型鳥類屬於廣域型鳥類,也廣泛出現在淺山 至平地的各類型受擾動棲地,例如黃頭鷺、大 卷尾、白腰文鳥等,這些鳥種在山村聚集所代 表地生態意涵有待進一步地探究,例如對其它 鳥種族群、昆蟲族群,以及植物傳播、更新會 產生的相關影響。
結 論
棲 地 破 碎 化 對 生 物 多 樣 性 所 產 生 的 影 響 取決於擾動的面積及擾動區塊的隔離度。新北 市金山區兩湖里山村的小面積農墾作業所形塑 的 森 林 — 農 墾 鑲 嵌 地 景 , 有 利 於 增 加 鳥 類α多 樣 性 。 此 一 森 林 — 農 墾 鑲 嵌 地 景 具 有 開 墾 面 積小、與周邊森林連結度很高(隔離度低)的特 性,可能是森林型鳥類及其它非森林型鳥類同 時出現在此一山村地景,導致鳥類α多樣性高於 純森林地景樣區的重要因素。兩湖山村森林—
農墾鑲嵌地景的α多樣性增加導因於出現大量干 擾地常見鳥種,若將調查範圍擴增,有可能會 出現α多樣性增加,但β、γ多樣性持平或降低 的情形。
引 用 文 獻
Altieri MA. 2004. Linking ecologists and tra- ditional farmers in the search for sustainable agriculture. Front Ecol Environ 2:35-42.
Amano T. 2009. Conserving bird species in Japanese farmland: past achievements and fu- ture challenges. Biol Conserv 142(9):1913-21.
Andrén H. 1994. Effects of habitat fragmenta- tion on birds and mammals in landscapes with different proportions of suitable habitat: a re- view. Oikos 71(3):355-66.
Bender DJ, Contreras TA, Fahrig L. 1998.
Habitat loss and population decline: a meta- analysis of the patch size effect. Ecology 79(2):517-33.
Benton TG, Vickery JA, Wilson JD. 2003.
Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol Evol 18:182-8.
Borcard D, Gillet F, Legendre P. 2011. Nu- merical ecology with R. New York: Springer.
306 p.
Cabra-García J, Chacón P, Valderrama- Ardila C. 2010. Additive partitioning of spider diversity in a fragmented tropical dry for- est (Valle del Cauca, Colombia). J Arachnol 38(2):192-205.
Carrara E, Arroyo-Rodríguez V, Vega- Rivera JH, Schondube JE, de Freitas SM, Fahrig LF. 2015. Impact of landscape com- position and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biol Conserv 184:117-26.
Clough Y, Gabriel D, Holzschuh A, Purtauf T, Kruess A, Steffan-Dewenter I, et al. 2007.
Alpha and beta diversity of arthropods and plants in organically and conventionally man- aged wheat fields. J Appl Ecol 44:804-12.
Clough Y, Putra DD, Pitopang R, Tscharn- tke T. 2009. Local and landscape factors de- termine functional bird diversity in Indonesian
cacao agroforestry. Biol Conserv 142:1032-41.
Diekötter T, Billeter R, Crist TO. 2008.
Effects of landscape connectivity on the spa- tial distribution of insect diversity in agri- cultural mosaic landscapes. Basic Appl Ecol 9(3):298-307.
Dinno A. 2017. dunn.test: Dunn’s test of mul- tiple comparisons using rank sums. R package, vers. 1.3.5. Available from https://CRAN.R- project.org/package=dunn.test.
Fang W, Lin RS, Fan MW. 2010. An avian conservation priority system for the Hushan Reservoir area. Taiwan J Biodivers 12(2):143- 66. [in Chinese with English summary].
Farwig N, Sajita NK, Bohning-Gaese K.
2008. Conservation value of forest plantations for bird communities in western Kenya. For Ecol Manage 258(11):3885-92.
Howe HF, Miriti MN. 2004. When seed dis- persal matters. BioScience 54(7):651-60.
Kadoya T, Suda S, Washitani I. 2009. Crisis of agricultural habitat complexes for dragon- flies in Japan: an assessment based on natural history data. Biol Conserv 142:1899-905.
Kadoya T, Washitani I. 2011. The Satoyama Index: a biodiversity indicator for agricultural landscapes. Agric Ecosyst Environ 140:20-6.
Katoh K, Sakai S, Takahashi T. 2009. Fac- tors maintaining species diversity in satoyama, a traditional agricultural landscape of Japan.
Biol Conserv 142:1930-6.
Kirika JM, Farwig N, Böhning-Gaese K.
2008. Effects of local disturbance of tropi- cal forests on frugivores and seed removal of a small‐seeded Afrotropical tree. Conserv Biol 22(2):318-28.
Kobori H, Primack RB. 2003. Participatory conservation approaches for satoyama, the traditional forest and agricultural landscape of Japan. Ambio 32(4):307-11.
Lawton JH, Bignell DE, Bolton B, Bloemers GF, Eggleton P, Hammond PM, et al. 1998.
Biodiversity inventories, indicator taxa and ef- fects of habitat modification in tropical forest.
Nature 391:72-6.
Legendre P, Legendre LF. 2012. Numerical ecology. 3rd ed. Amsterdam, the Netherlands:
Elsevier. 1006 p.
Lewis SL. 2006. Tropical forests and the changing earth system. Phil Trans R Soc B 361:195-210.
Lu HM, Chen JH. 2006. Study on the bird diversity and landscape diversity: a case study of ditch landscape in Wugou Village, Pingtung County J Archit 57:123-41. [in Chinese with English summary].
Luck GW, Daily GC. 2003. Tropical country- side bird assemblages: richness, composition, and foraging differ by landscape context. Ecol Appl 13(1):235-47.
Moran C, Catterall CP, Green RJ, Olsen MF. 2004. Functional variation among fru- givorous birds: implications for rainforest seed dispersal in a fragmented subtropical land- scape. Oecologia 141:584-95.
Neuschulz EL, Botzat A, Farwig N. 2011.
Effects of forest modification on bird com- munity composition and seed removal in a het- erogeneous landscape in South Africa. Oikos 120:1371-9.
Ohara RG, Ushimaru A. 2015. Plant beta- diversity is enhanced around grassland-forest edges within a traditional agricultural land- scape. Appl Veg Sci 18:493-502.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, et al. 2018. veg- an: Community Ecology Package. R package, vers. 2.5-1. Available from https://CRAN.R- project.org/package=vegan.
Pacheco R, Vasconcelos HL. 2012. Habitat diversity enhances ant diversity in a naturally heterogeneous Brazilian landscape. Biodivers Conserv 21:797-809.
Peh KSH, Jong J, Sodhi NS, Lim SLH, Yap
CAM. 2005. Lowland rainforest avifauna and human disturbance: persistence of primary forest birds in selectively logged forests and mixed-rural habitats of southern Peninsular Malaysia. Biol Conserv 123(4):489-505.
Philpott SM.2008. Biodiversity loss in Latin American coffee landscapes: review of the evi- dence on ants, birds, and trees. Conserv Biol 22(5):1093-105.
R Core Team. 2018. R: a language and envi- ronment for statistical computing, vers. 3.5.0.
Vienna, Austria: R Foundation for Statistical Computing. Available at http://www.r-project.
org.
Ranganathan J, Ranjit Daniels RJ, Subash Chandran MD, Ehrlich PR, Daily GC. 2008.
Sustaining biodiversity in ancient tropical
countryside. PNAS 105(46):17852-4.
Sekercioglu CH. 2006. Increasing awareness of avian ecological function. Trends Ecol Evol 21(8):464-71.
Socolar JB, Gilroy JJ, Kunin WE, Ed- wards DP. 2016. How should beta-diversity inform biodiversity conservation? Trends Ecol Evol 31(1):67-80.
Turner IM. 1996. Species loss in fragments of tropical rain forest: a review of the evidence. J Appl Ecol 33(2):200-9.
Washitani I. 2001. Traditional sustainable ecosystem ‘SATOYAMA’ and biodiversity cri- sis in Japan: conservation ecological perspec- tive. Glob Environ Res 5:119-33.
Zar JH. 2010. Biostatistical analysis. 5th ed.
Upper Saddle River, NJ: Prentice Hall. 944 p.