Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Coactivator TIF1b Interacts with Transcription Factor C/EBPb
and Glucocorticoid Receptor To Induce
a1-Acid Glycoprotein
Gene Expression
CHING-JIN CHANG,
1YA-LING CHEN,
2,3ANDSHENG-CHUNG LEE
1,2,3*
Institute of Biological Chemistry, Academia Sinica,
1and Institute of Molecular Medicine
2and H. L. Tsai Memorial
Laboratory,
3College of Medicine, National Taiwan University, Taipei, Taiwan
Received 12 March 1998/Returned for modification 12 June 1998/Accepted 14 July 1998
The transcription of the
a1-acid glycoprotein gene is induced by inflammatory cytokines and glucocorticoids.
C/EBP
b is a major transcription factor involved in the induction of the agp gene by some cytokines. In this
report, we have identified a novel transcriptional intermediary factor, TIF1
b, which could enhance the
transcription of the agp gene by the glucocorticoid receptor (GR) and C/EBP
b. TIF1b belongs to a subgroup
of RING (really interesting new gene) finger proteins that contain a RING finger preceding two B box-type
fingers and a putative coiled-coil domain (RBCC domain). Immunoprecipitation experiments showed that the
interaction between GR and TIF1
b is ligand independent. The overexpression of the TIF1b gene enhances
GR-regulated expression in a ligand- and glucocorticoid-responsive element (GRE)-dependent manner. TIF1
b
can also augment C/EBP
b-mediated activity on wild-type and GRE-mutated agp genes, but this augmentation
is diminished when all three C/EBP
b-binding elements are mutated. Functional and biochemical
character-izations indicated that the bZIP domain of C/EBP
b and the RBCC domain, plant homeodomain finger, and
bromodomain of TIF1
b are crucial for the interactions of these proteins. Taken together, these results suggest
that TIF1
b serves as a converging mediator of signal transduction pathways of glucocorticoids and some
inflammatory cytokines.
The acute-phase reaction to inflammatory stimuli is
ac-companied by an increase in a variety of serum proteins,
col-lectively named acute-phase proteins. The synthesis of these
proteins is regulated by glucocorticoids and inflammatory
cy-tokines, such as interleukin 1 (IL-1), IL-6, and tumor necrosis
factor alpha (5–7, 62). C/EBPb was initially identified as the
key transcription factor involved in the regulation of the
a1-acid glycoprotein (AGP) gene during the acute-phase response
(termed AGP/EBP) (18). C/EBPb was also shown to be
in-volved in the regulation of a number of other genes, such as
those for IL-6 and albumin (termed NF-IL-6, LAP, IL-6DBP,
or CRP2) (2, 14, 20, 49, 61). In addition to C/EBPb-binding
motifs, a glucocorticoid-responsive element (GRE) also exists
between
2120 and 2107 in the 59-flanking region of the agp
gene (8, 18). Previous reports showed that maximal induction
of the agp gene by glucocorticoids also requires another
C/EBPb-binding element located downstream of GRE (34, 50,
60). The synergistic interaction between cytokines and
glu-cocorticoids has been attributed to protein-protein interactions
between C/EBPb and the glucocorticoid receptor (GR) (45).
GR belongs to a family of nuclear receptors that function as
ligand-dependent transcription factors (9, 48). Transcriptional
activation of target genes by nuclear receptors is mediated by
two activation regions, AF1, located in the N terminus, and
AF2, located in the C terminus of the hormone-binding
do-main of the receptor. GR-mediated transcription is promoter
dependent and cell specific (for a review, see reference 24).
Results from studies of transcriptional interference or
squelching between AF1 and AF2 of steroid receptors
sug-gested the existence of coactivators or transcriptional
interme-diary factors which interact specifically with the AF1 and AF2
domains (3, 39, 55). Recent studies have led to the
identifica-tion of several proteins that interact with nuclear receptors in
a ligand-dependent manner and play essential roles in
medi-ating their transcriptional activities. These proteins include
RIP140 (15), TIF1 (36), Trip1/SUG1 (38, 59), SRC-1/p160 (26,
31, 47), TIF2/Grip1 (29, 58), ARA70 (63), and CBP/p300 (16,
27, 31). Several of these factors showed markedly different
affinities for various nuclear receptors (56, 59). CBP and p300
are large nuclear proteins and have been demonstrated to
interact functionally with a number of sequence-specific
tran-scriptional activators (for a review, see reference 30). Previous
data indicated that competition for limiting amounts of CBP
may account for many of the inhibitory actions of both GR and
the retinoic acid receptor on AP1 activation (31).
Genes for two related TIF1 proteins, TIF1a and TIF1b,
have been cloned and shown to be members of the RING
(really interesting new gene) finger family (for reviews, see
references 12 and 52). The RING finger motif can be defined
simply as Cys3-His-Cys4, a new class of the zinc finger. At least
80 members of the RING finger family have been identified.
Many members, including the tumor suppressor BRCA-1 (42),
the oncogene product Mel18 (32), and the mediator of the
tumor necrosis factor receptor, TRAF2 (51), have been
impli-cated as being in control of cell growth, cell differentiation, and
development. The functions of these RING fingers remain to
be defined, although some reports have suggested that they are
the interface for protein-protein interactions (4, 10).
To delineate the mechanisms of transcriptional regulation of
the agp gene by C/EBPb, we have initiated studies on proteins
that interact with C/EBPb by purifying them using a number of
procedures, including anti-C/EBPb antibody immunoaffinity
chromatography (40, 41). In this report, we present results on
the identification and characterization of the roles of TIF1b in
the activation of the agp gene. These results indicate that the
* Corresponding author. Mailing address: Institute of Molecular
Medicine, College of Medicine, National Taiwan University, #7 Chun
Shan South Rd., Taipei, Taiwan. Phone: 2356-2982. Fax:
886-2-2321-0977. E-mail: [email protected].
5880
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
enhancement of GR or C/EBPb activity by TIF1b occurs
through direct protein-protein interactions.
MATERIALS AND METHODS
Plasmids and constructs.The EST clone containing partial human TIF1b cDNA (from nucleotides 1882 to 2673) was obtained from Research Genetics. An 0.8-kb DNA fragment insert isolated from the plasmid was used as a probe for screening the day-16 mouse embryo cDNA library (Novagen). A cDNA clone with a 2.8-kb insert containing the complete open reading frame of TIF1b was obtained. Mammalian expression plasmids were constructed by cloning the fol-lowing TIF1b fragments into cytomegalovirus (CMV) expression vector pcDNA3 (Invitrogen): the full-length EcoRI-HindIII fragment (pcDNA3-TIF1b), and EcoRI-SacI fragment (residues 1 to 563), an EcoRI-PvuII fragment (residues 1 to 372), and a fragment resulting from BamHI deletion (residues 80 to 383 deleted) of the full-length EcoRI-HindIII fragment. An SfiI-HindIII fragment (residues 14 to 834), a BamHI fragment (residues 80 to 383), a BamHI-SacI fragment (residues 383 to 563), and a BamHI-HindIII fragment (residues 383 to 834) were cloned into the pGEX-1 vector (Pharmacia) for the production of glutathione S-transferase (GST) fusion proteins. The full-length EcoRI frag-ment of C/EBPb, an N-terminal NcoI fragfrag-ment (amino acids 21 to 151 [C/EBPb-N), or a C-terminal NcoI-HindIII fragment (amino acids 151 to 296 [C/EBPb-C]) was ligated to the pRSET vector (Invitrogen) for recombinant protein production.
Other plasmid constructs, such as pCMV-C/EBPb, rat AGP (wild type [WT])-CAT, AGP (C mutant)-[WT])-CAT, AGP (D mutant)-[WT])-CAT, AGP (E mutant)-[WT])-CAT, AGP (CDE mutant)-CAT, and AGP (GRE mutant)-CAT, were as described previously (40, 59). Briefly, the plasmids were obtained by ligation of the wild-type or mutant (see below) rat agp gene promoter sequence from2736 to 11 to the chloramphenicol acetyltransferase (CAT) reporter gene. The C, D, E, and GRE mutants correspond to serial 3-base substitutions at positions274 to 272 (ACA to GTG),296 to 294 (CAA to TGG), 2106 to 2104 (AGA to GAG), and2118 to 2116 (ACA to GTG), respectively. pMMTV-CAT is the mouse mammary tumor virus long terminal repeat ligated to the CAT reporter gene. The mammalian cell expression vector (pRSV-hGR) and recombinant baculo-virus containing GR were kindly provided by M.-J. Tsai of Baylor College of Medicine. pRSV-CREB and pCMV-PKAc were obtained from Susan Taylor.
Recombinant proteins and antibodies.Human TIF1b (from nucleotide 1882 to 2673) was cloned into the pRSET vector and expressed in Escherichia coli BL21(DE3)(pLysS). This recombinant protein was purified on a nickel column and used for rabbit immunization. Monoclonal and polyclonal antibodies to C/EBPb were as described previously (18). Anti-GR antibody was purchased from Santa Cruz Biotech.
Cell cultures, transient transfection, and CAT assay. BHK, HeLa, and P388D1 cells were cultured in Iscove’s modified Dulbecco’s medium supple-mented with 10% fetal calf serum. DNA transfection was performed by the calcium phosphate precipitation method. BHK cells were grown in 6- or 3.5-cm-diameter petri dishes to 30 to 40% confluence. The amounts of CAT reporter plasmid DNA and expression plasmid DNA used in each experiment are de-scribed in the figure legends. pcDNA3 plasmid DNA was used to adjust the total amount of DNA for each transfection to be equal. pCMV/SEAP (Tropix) (0.5 mg) was included in each transfection as an internal control for transfection efficiency. During the 24-h posttransfection period, the cells were placed in fresh medium and, in some experiments, induced with 1mM dexamethasone (water soluble; Sigma). Cells were harvested 24 h later and extracted with 100ml of 0.25 M Tris-HCl (pH 7.8). The acetylated forms of chloramphenicol were separated by thin-layer chromatography and quantified with an image analyzer (BAS 1000; Fuji). All transfection experiments were repeated two to four times.
Preparation of whole-cell extracts, immunoprecipitation, and Western blot-ting.Whole-cell extracts from P388D1 cells were prepared by lysing the cells with buffer containing 25 mM HEPES (pH 7.6), 0.3 M NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5% Nonidet P-40 (NP-40), and 0.5 mM dithiothreitol (DTT). For immunoprecipitation analysis, 1 mg of whole-cell extracts was precleaned with preimmune serum and protein A-Sepharose in 0.5 ml of immunoprecipitation buffer (25 mM HEPES [pH 7.6], 0.25 M NaCl, 1 mM EDTA, 0.1% NP-40, 0.5 mM DTT, 6% glycerol) at 4°C for 2 h. The precleaned supernatants were incubated with 5mg of anti-TIF1b or anti-C/EBPb antibody and protein A-Sepharose in the presence or absence of 1mM dexamethasone at 4°C for 90 min. After extensive washes, the protein complex was dissolved in sodium dodecyl sulfate (SDS) loading buffer and subjected to SDS-polyacrylamide gel electro-phoresis (PAGE). The separated polypeptides were blotted onto a Hybond-C membrane (Amersham) and probed with anti-TIF1b, anti-C/EBPb, or anti-GR antibody. The results were detected with an enhanced chemiluminescence kit (Amersham).
Protein-protein interaction assay.Glutathione-Sepharose 8A beads (Pharma-cia) were mixed with 3mg of wild-type or deletion mutant recombinant GST-TIF1b fusion protein or GST only in 500 ml of phosphate-buffered saline con-taining 1% Triton X-100 on a rotary shaker for 20 min at room temperature. The beads were washed three times with phosphate-buffered saline, combined with 100 ng of recombinant full-length C/EBPb, truncated C/EBPb-N, or truncated C/EBPb-C in a final volume of 500 ml of binding buffer (25 mM HEPES [pH 7.6], 0.25 M NaCl, 1 mM EDTA, 0.1% NP-40, 0.5 mM DTT, 6% glycerol), and
incubated on a rotary shaker for 2 h at 4°C. The beads were washed three times with binding buffer, and the bound proteins were subjected to SDS-PAGE and Western blot analysis.
RESULTS
Isolation and characterization of cDNA clones for TIF1
b.
The RING finger protein family consists of members found in
animals, plants, and viruses, but the function of the RING
finger domain remains to be defined. By comparison with
se-quences of RING finger domains similar to those of inhibitors
of apoptosis or RING-1, a number of human EST clones
carrying putative RING finger domains were identified.
Se-quence analysis revealed that one of these clones, containing
an 0.8-kb insert, was highly homologous to a mouse protein,
TIF1b (37). Rabbit antibodies were generated by use of a
recombinant protein derived from the EST clone. In
serendip-itous Western blot experiments for identifying
C/EBPb-inter-acting proteins, we used a rabbit anti-TIF1b antibody as a
control. Surprisingly, it reacted with a protein of
;100 kDa
that appeared in the eluent of the anti-C/EBPb immunoaffinity
column (data not shown). This observation prompted us to
study the possible physical and functional interactions between
TIF1b and C/EBPb. Using the 0.8-kb DNA fragment as a
probe to screen the mouse cDNA library, a 2.8-kb cDNA clone
that could encode a protein of 834 amino acids was obtained.
TIF1b and TIF1a are strongly homologous in the N- and
C-terminal regions. The N-terminal region is a RING finger
preceding two B box-type fingers and a putative coiled-coil
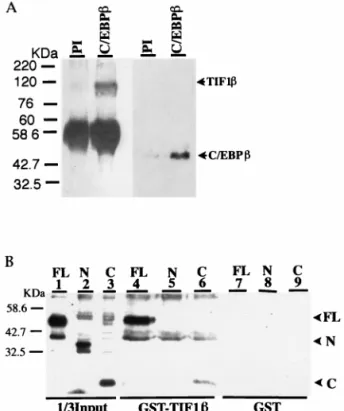
FIG. 1. Protein-protein interactions between C/EBPb and TIF1b. (A) P388D1 whole-cell extracts were immunoprecipitated with anti-C/EBPb poly-clonal antibody (C/EBPb) or preimmune serum (PI). The precipitated proteins were subjected to SDS-PAGE followed by Western blotting with anti-C/EBPb monoclonal or anti-TIF1b polyclonal antibodies. (B) Several recombinant C/EBPb constructs were incubated with glutathione bead-immobilized GST-TIF1b (lanes 4 to 6) or GST (lanes 7 to 9). After extensive washes, the protein complexes were subjected to SDS-PAGE and immunoblotted with anti-C/EBPb antibody. Lanes 1 to 3 represent direct loading of different recombinant C/EBPb constructs. FL, N, and C represent full-length C/EBPb, C/EBPb-N, and C/EBPb-C, which are described in Materials and Methods.at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
domain (RBCC motif) (36). The C-terminal region consists of
a plant homeodomain (PHD) finger followed by a
bromodo-main (36).
TIF1b appears to be widely expressed, since Northern
blot-ting reveals a major 3-kb TIF1b transcript in all human tissues
(data not shown). The subcellular localization of TIF1b was
then determined by indirect immunofluorescence, which
showed granular staining of TIF1b only in the nucleoplasm
and not in the nucleolus (data not shown).
TIF1
b stimulates the C/EBPb-mediated activation of the
agp gene.
The physical interaction of C/EBPb and TIF1b was
examined by an immunoprecipitation assay. P388D1 is a
mouse macrophage cell line which expresses C/EBPb
consti-tutively. Anti-C/EBPb antibody can bring down TIF1b in
P388D1 whole-cell extracts (Fig. 1A). Direct protein-protein
interactions were studied by pull-down assays with GST-TIF1b
(full length) and both full-length C/EBPb and truncated forms
of C/EBPb, C/EBPb-N and C/EBPb-C (Fig. 1B). The results
showed that the bZIP domain of C/EBPb (i.e., C/EBPb-C) is
sufficient for its direct interaction with TIF1b.
To characterize the functional role of TIF1b in the
activa-tion of the agp gene by C/EBPb, we performed cotransfecactiva-tion
experiments. TIF1b augmented the activation of the agp gene
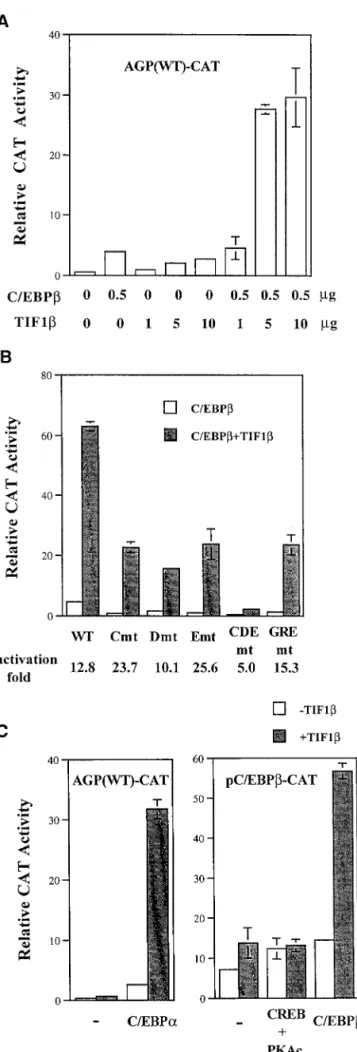
by C/EBPb in a dose-dependent manner (Fig. 2A). To further
elucidate the roles of C/EBPb-binding motifs in the activation
effect between C/EBPb and TIF1b, we performed transfection
assays with reporters containing mutated C/EBPb-binding
mo-tifs (C, D, E, and CDE mutants). Compared to the results
obtained with the wild-type reporter gene, there was
stimula-tion of C/EBPb activity by TIF1b with the C-, D-, or
E-site-mutated reporter gene (13-fold for the wild type and 26-, 10-,
and 24-fold for the C, D, and E mutants, respectively).
Induc-tion by these factors was dramatically reduced when a reporter
containing mutations of all three C/EBPb-binding sites (i.e.,
the CDE mutant) was tested (Fig. 2B). Transactivation of the
GRE-mutated agp gene by C/EBPb and TIF1b was
compara-ble to that of the wild-type gene (Fig. 2B). Thus, the
augmen-tation effect of TIF1b on the activation of the C/EBPb gene is
exclusively C/EBPb-binding motif dependent, and GR is
ap-parently not required.
To further assess the activation specificity of TIF1b and
other factors, we conducted transient transfection assays with
expression vectors for C/EBPa, CREB, and TIF1b. As shown
in Fig. 2C, left panel, TIF1b also augmented the
C/EBPa-mediated transactivation of the agp gene. The pC/EBPb-CAT
reporter contains the promoter from the c-ebpb gene
(nucle-otides
2390 to 182) (17). It has been reported that there are
C/EBP- and CREB-responsive elements in the regulatory
re-gion of c-ebpb (17, 44). TIF1b potentiated C/EBPb activity but
FIG. 2. Stimulation of C/EBPb-mediated gene activation by TIF1b. (A) BHK cells were cotransfected with 2mg of AGP (WT)-CAT, 0.5 mg of pCMV-C/EBPb (pCMV-C/EBPb), and increasing amounts of pcDNA3-TIF1b (TIF1b) (1, 5, and 10mg). (B) In 3.5-cm-diameter petri dishes, BHK cells were cotransfected with 0.5mg of AGP (WT)-CAT, AGP (C mutant)-CAT, AGP (D mutant)-CAT, AGP (E mutant)-CAT, AGP (CDE mutant)-CAT, or AGP (GRE mutant)-CAT and 0.1mg of pCMV-C/EBPb with or without 2 mg of pcDNA3-TIF1b. The data represent the average activity of two independent duplicate experiments. The fold induction by TIF1b is indicated below the panel. mt, mutant. (C) (Left panel) In 3.5-cm-diameter petri dishes, BHK cells were cotransfected with 0.5mg of AGP (WT)-CAT and 0.05mg of pCMV-C/EBPa in the presence or absence of 2mg of pcDNA3-TIF1b. (Right panel) BHK cells were transfected with 0.25 mg of pC/EBPb-CAT, and 0.1 mg of pCMV-C/EBPb or both 0.1 mg of pRSV-CREB and 0.1 mg of pCMV-PKAc in the absence or presence of 2 mg of pcDNA3-TIF1b. Error bars indicate standard deviations. 2, pcDNA3 vector control.
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
not CREB activity in the c-ebpb gene promoter (Fig. 2C, right
panel).
To define the regions of TIF1b that could interact with
C/EBPb physically and functionally, we constructed
mamma-lian cell expression vectors and prepared recombinant GST
fusion proteins of various deletion mutants of TIF1b (Fig. 3A
and B, upper panels). Mutants of TIF1b with either the PHD
finger and the bromodomain deleted (amino acids 1 to 563) or
the RBCC domain deleted (amino acids 80 to 383 deleted)
failed to potentiate the activation of the agp gene by C/EBPb
(Fig. 3A, lower panel). Physical interaction experiments with
recombinant proteins derived from C/EBPb and deletion
mu-tants of TIF1b indicated that the RBCC domain of TIF1b was
sufficient to interact with C/EBPb (Fig. 3B, lower panel). The
region of amino acids 383 to 563 seemed to interact weakly
with C/EBPb. Taken together, these results suggest that TIF1b
interacts with C/EBPb through the RBCC domain and
en-hances C/EBPb transcriptional activity through the PHD
fin-ger and the bromodomain.
TIF1
b enhances the transcriptional activity of GR. TIF1a
was identified as a protein that interacts directly with the
li-gand-binding domains of several nuclear receptors in a
ligand-and AF2-dependent manner both in vivo ligand-and in vitro. It was
suggested that TIF1a mediates the transcriptional activation of
the target gene by the AF2 domain of nuclear receptors (36).
TIF1a and TIF1b share highly conserved domains. To further
investigate the role of TIF1b in nuclear receptor-mediated
transactivation of target gene expression, we performed
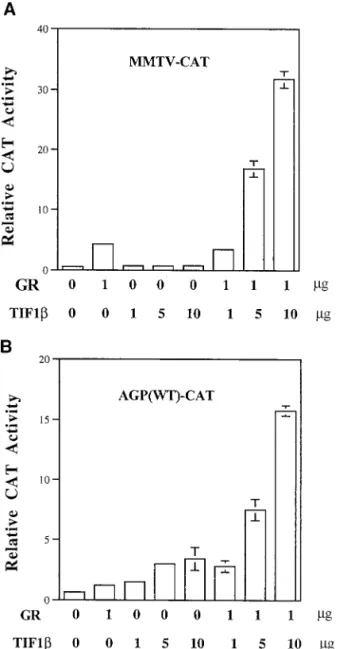
tran-sient transfection assays. As shown in Fig. 4, in the presence of
exogenous GR and dexamethasone, TIF1b stimulated the
transcription of mouse mammary tumor virus and the agp
promoter in a dose-dependent manner. TIF1b did not
aug-ment the transcriptional activity of GR in the absence of
dexa-methasone (data not shown).
To determine the molecular basis of target gene activation
by TIF1b and GR, we conducted an analysis of the interaction
between TIF1b and GR by an immunoprecipitation assay.
Polyclonal antibody to TIF1b but not preimmune serum
im-munoprecipitated GR from P388D1 whole-cell extracts in the
absence or presence of dexamethasone (Fig. 5A; the anti-GR
antibody detected two isoforms, 95 and 90 kDa). This result
indicates that TIF1b and GR coexist in a complex. Direct
protein-protein interactions were examined by an
immunopre-cipitation assay with GST-TIF1b fusion protein and
recombi-nant GR. TIF1b interacted with GR in the absence or
pres-ence of dexamethasone (Fig. 5B).
Previous studies indicated that maximal induction of the agp
gene by glucocorticoid requires the downstream
C/EBPb-bind-ing sequences (34, 50, 60). To further study the TIF1b-mediated
GR induction of the agp gene, we performed transfection assays
using agp promoters containing mutated C/EBPb-binding sites or
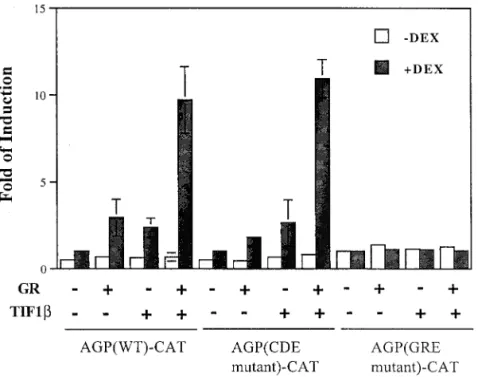
mutated GRE. As indicated by the previous results, the
activa-tion of the agp gene by TIF1b and GR was observed only in the
presence of dexamethasone (Fig. 6). Reporters containing
tated C/EBPb-binding elements (namely, C, D, and E
mu-tants) remained responsive to TIF1b and GR. In contrast, the
reporter containing mutated GRE was unresponsive to TIF1b
and GR (Fig. 6). The activation of the agp gene by TIF1b alone
in the presence of dexamethasone was likely due to the effect
of endogenous GR. These results show that the transcriptional
activation of the agp gene by TIF1b and GR is dependent on
GRE but independent of C/EBPb-binding motifs.
FIG. 3. Functional and biochemical characterization of C/EBPb-interacting domains of TIF1b. (A) (Upper panel) Schematic representation of several TIF1b expression vectors; numbers denote amino acid positions. (Lower panel) Transient transfection assays. BHK cells (in 3.5-cm-diameter petri dishes) were transfected with 0.5mg of AGP (WT)-CAT, 0.1 mg of pCMV-C/EBPb, and 2 mg of pcDNA3-TIF1b (full length [fl] or from amino acid 1 to 563 or 1 to 372, or full length but with amino acids 80 to 383 deleted). (B) (Upper panel) Schematic representation of several GST-TIF1b fusion proteins. (Lower panel) Protein pull-down assay. Glutathione bead-immobilized recombinant GST-TIF1b (fl or from amino acid 80 to 383, 383 to 563, or 383 to 834) incubated with full-length recombinant C/EBPb (100 ng). After extensive washes, the protein complex was analyzed by immunoblotting with anti-C/EBPb antibody. 2, pcDNA3 vector control.
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
We further examined the effect of TIF1b on the agp gene by
cotransfecting pRSV-GR and pCMV-C/EBPb simultaneously.
As shown in Fig. 7, the net effect of the transactivation of the
agp gene by cotransfection of pCMV-C/EBP
b, pRSV-GR, and
pCMV-TIF1b seemed to be the result of pRSV-GR plus
pCMV-TIF1b. Taken together, these results suggest that there
is no additive or synergistic activation of the agp gene by the
overexpression of GR, TIF1b, and C/EBPb under these
exper-imental conditions.
DISCUSSION
TIF1b was originally identified as a protein that interacts
directly with two chromosomal proteins, mHP1a and mMOD1
(37). In this report, TIF1b was identified as a coactivator for
C/EBPb and GR in the activation of the agp gene. The
spec-ificity of TIF1b for GR or C/EBPb was demonstrated by
trans-fection assays with wild-type or mutant reporter plasmids
con-taining GRE and/or C/EBPb-binding elements. Direct
protein-protein interactions between TIF1b and GR or
C/EBPb were apparently responsible for the activation of the
agp gene by these proteins. These results suggest that TIF1
b
may act as an integrator or coactivator for both glucocorticoid
and cytokine signaling pathways leading to the activation of
C/EBPb. The identification of TIF1b as a coactivator provides
further clues about the mechanisms of transactivation by GR
and C/EBPb and the regulation of their target genes, such as
acute-phase response genes.
The competition for a common coactivator, CBP, by AP-1
and the nuclear receptor provides an example of how genes
that contain either an AP-1- or a nuclear receptor-binding site
could be regulated (31). However, a more complex pattern of
regulation was observed for genes containing composite
re-sponse elements. One GRE and three C/EBPb-binding sites
are located in the upstream regulatory region of the agp gene.
The regulation of this gene by interactions between TIF1b,
GR, and C/EBPb seems to be complex and depends on the
steady-state levels of these factors. When pRSV-GR,
pCMV-C/EBPb, and pCMV-TIF1b were cotransfected into cells, no
apparent changes in the transactivation of the agp gene were
seen compared to those seen with pRSV-GR and
pCMV-TIF1b (Fig. 7). In fact, the slight decrease in activation
ob-served could have been attributed to competition between
C/EBPb and GR for TIF1b.
Results from the deletion analysis indicated that the RBCC
domain and the PHD domain-bromodomain are essential for
the function of TIF1b. The RBCC domain is the
C/EBPb-interacting domain. Although the actual functional significance
of these domains is unknown, it is currently assumed that they
are involved in protein-protein interactions. The RBCC motif
has been found in the N-terminal part of several putative
transcriptional factors, ribonucleoproteins, and
proto-onco-gene products, including PML, RFP, RPT-1, SS-A/Ro, XNF7,
and PWA33 (for a review, see reference 21). Three RBCC
domain-containing proteins, PML, RFP, and TIF1a, have
been identified in the context of fusion oncoproteins resulting
from chromosomal translocations. The RBCC domain is fused
to truncated products of other genes (23, 36, 54). Mutations in
the RBCC domain of PML prevent PML nuclear body
forma-tion (11). Many PHD finger-containing proteins have been
FIG. 4. TIF1b potentiates GR-activated gene expression. (A) HeLa cells were transiently transfected with 2mg of pMMTV-CAT reporter plasmid, 1 mg of pRSV-hGR (GR), and increasing amounts of pcDNA3-TIF1b (TIF1b) (1, 5, and 10mg). Each assay was done in the presence of 1 mM dexamethasone. (B) BHK cells were transiently transfected with 2mg of AGP (WT)-CAT reporter plasmid. Other plasmids and conditions of treatment are as described for panel A. Relative CAT activity normalized with an internal control represents an average of two independent duplicate experiments. Error bars indicate standard deviations.
FIG. 5. Protein-protein interactions between GR and TIF1b. (A) Immuno-precipitation of GR by anti-TIF1b antibody. P388D1 whole-cell extracts were immunoprecipitated with anti-TIF1b or control (PI) antibody in the presence (1) or absence (2) of 1 mM dexamethasone (DEX) and subjected to SDS-PAGE and Western blotting with anti-GR polyclonal antibody. (B) Recombi-nant GST-TIF1b was incubated with recombiRecombi-nant GR in the presence (1) or absence (2) of 1 mM dexamethasone and then immunoprecipitated with anti-TIF1b antibody or preimmune serum. After extensive washes, the protein com-plex was analyzed by Western blotting with anti-GR antibody. Recombinant GR used for the interaction assay (1/5 input) was included as a control.
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
implicated in interactions between chromosomal proteins (1).
These include products of the Drosophila genes trithorax and
polycomblike. It is interesting to note that bromodomains are
found in the adaptor proteins p300, CBP, and GCN5, as well as
in SWI/SNF2 (13, 30, 35). These proteins reside in large
mul-tiprotein complexes. Thus, the overall structure of TIF1b
im-plies that it activates gene transcription by taking part in the
formation of multiprotein complexes.
A TIF1b-related protein, TIF1a, was found to interact with
several nuclear hormone receptors. TIF1a contains a nuclear
receptor-binding motif, LXXLL (28, 37). However, there is no
motif resembling LXXLL in the TIF1b sequence. Inhibition of
RXRa activity was observed as a result of ectopic expression of
TIF1a in the transient transfection assays. The RBCC
domain-containing protein PML exerts a very powerful enhancing
ef-fect on the transactivating properties of several steroid
hor-mone receptors (25). It is likely that a specific functional
interaction exists between coactivators and nuclear receptors.
The activation effect of TIF1a or TIF1b on GR- or
RXRa-induced gene expression, respectively, remains to be studied.
In addition to GR and C/EBPb, TIF1b has been reported to
interact with the transcriptional silencing domain of the
Dro-sophila Kruppel-related KRAB proteins and to serve as a
core-pressor (22, 33, 43). The mechanism of repression by TIF1b
remains elusive. To test the possibility that TIF1b is a general
mediator of various transcriptional factors, we performed
co-transfection assays with mammalian cell expression vectors for
CREB and the protein kinase A catalytic subunit in the
pres-ence of the pC/EBPb-CAT reporter. TIF1b could not enhance
CREB activity (Fig. 2C). Another member of the C/EBP
fam-ily, C/EBPa, was also tested, and the results showed that
TIF1b could augment the C/EBPa-activated expression of
AGP (WT)-CAT (Fig. 2C). Thus, in addition to C/EBPb, the
transcriptional activity of another member of the C/EBP family
may be modulated by TIF1b.
How could TIF1b function as a coactivator? TIF1b seems to
be a bifunctional protein involved in the remodeling of the
chromatin template in both the repression and the activation
of transcription (37). Thus, the interaction between GR and
TIF1b or between C/EBPb and TIF1b may promote the
con-version of a transcriptionally inactive heterochromatin-like
FIG. 6. GRE-dependent and C/EBPb-binding-element-independent augmentation of the activation of AGP-CAT by GR and TIF1b. AGP (WT)-CAT, AGP (CDE mutant)-CAT, or AGP (GRE mutant)-CAT was used as a reporter (see Materials and Methods). In 3.5-cm-diameter petri dishes, BHK cells were cotransfected with one reporter plasmid (1mg) and 0.5 mg of pRSV-hGR (GR), 5 mg of pcDNA3-TIF1b (TIF1b), or both in the presence or absence of 1 mM dexamethasone (DEX). The fold induction for each experiment is shown. The values are the averages of at least two independent experiments. Error bars indicate standard deviations.
FIG. 7. Activation of the agp gene by C/EBPb, GR, and TIF1b. BHK cells (grown in 6-cm petri dishes) were transfected with 2mg of AGP (WT)-CAT and 0.5mg of pCMV-C/EBPb (C/EBPb), 1 mg of pRSV-hGR (GR), or 5 mg of pcDNA3-TIF1b (TIF1b) in various combinations in the absence or presence of 1mM dexamethasone (DEX). Normalized relative CAT activity represents an average of two independent experiments. Error bars indicate standard devia-tions.
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
structure to an active euchromatin-like open structure by
trig-gering the release of HP1 and MOD1 (37). We previously
identified a nucleolar phosphoprotein, Nopp140, that
func-tions as a mediator between C/EBPb and the general
tran-scription factor TIFIIB (40). In light of the analogous features
shared by the interactions between C/EBPb and Nopp140 or
TIF1b, we also tested the physical interaction between TIF1b
and TFIIB. Our results did not offer conclusive evidence on
any physical interaction between TIF1b and TFIIB (data not
shown). Thus, the mechanism of activation of the agp gene by
C/EBPb and TIF1b is different from that of C/EBPb and
Nopp140.
The present results revealed a direct protein-protein
inter-action between TIF1b and GR and showed that this
interac-tion is ligand independent. However, the funcinterac-tional interacinterac-tion
between GR and TIF1b is ligand dependent. It is speculated
that TIF1b may participate in the formation of a coactivator
complex to activate target gene expression. CBP has been
demonstrated to interact with other nuclear receptor
coactiva-tors (SRC-1/ACTR/pCIP family) to form a functional complex
and to result in a synergistic response to the nuclear receptors
(19, 31, 57). These coactivators have been identified as histone
acetyltransferases that remodel chromatin structure to
facili-tate transcriptional activation (19, 46, 53). Further experiments
to identify other TIF1b-interacting proteins may yield
impor-tant mechanistic insights.
ACKNOWLEDGMENTS
This research was supported by grants NSC86-2311-B001-089 and
NSC88-2311-B001-114 (to C.-J.C.) and NSC86-2311-B001-094-Y (to
S.-C.L.) from the National Science Council.
We thank Susan Taylor, Sophia Tasi, and Ming-Jer Tasi for plasmids
and Bertrand Chin-Ming Tan for critical reading of the manuscript.
REFERENCES
1. Aasland, R., T. J. Gilson, and A. F. Stewart. 1995. The PHD finger: impli-cations for chromatin-mediated transcriptional regulation. Trends Biochem. Sci. 20:56–59.
2. Akira, S., H. Isshiki, T. Sugita, O. Tanabe, S. Kinoshita, Y. Nishio, T. Nakajima, T. Hirano, and T. Kishimoto.1990. A nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP family. EMBO J. 9:1897–1906. 3. Barettino, D., M. D. M. Vianco-Ruiz, and H. G. Stunnenberg. 1994. Char-acterization of the ligand-dependent transactivation domain of thyroid hor-mone receptor. EMBO J. 13:3039–3049.
4. Barlow, P. N., B. Luisi, A. Milner, M. Elliott, and R. Everett. 1994. Structure of the C3HC4 domain by 1H-nuclear magnetic resonance spectroscopy. A new structural class of zinc-finger. J. Mol. Biol. 237:201–211.
5. Baumann, H., G. L. Firestone, T. L. Burgess, K. W. Gross, K. R. Yamamoto, and W. A. Held.1983. Dexamethasone regulation ofa1-acid glycoprotein and other acute phase reactants in rat liver and hepatoma cells. J. Biol. Chem. 258:563–570.
6. Baumann, H., and J. Gauldie. 1990. Regulation of hepatic acute phase plasma protein genes by hepatocyte stimulating factors and other mediators of inflammation. Mol. Biol. Med. 7:147–159.
7. Baumann, H., G. P. Jahreis, and K. K. Morella. 1990. Interaction of cyto-kine- and glucocorticoid-response elements of acute-phase plasma protein genes. J. Biol. Chem. 265:22275–22281.
8. Baumann, H., and L. E. Maquat. 1986. Localization of DNA sequences involved in dexamethasone-dependent expression of the rata1-acid glyco-protein gene. Mol. Cell. Biol. 6:2551–2561.
9. Beato, M., P. Herrlich, and G. Schutz. 1995. Steroid hormone receptors: many actors in search of a plot. Cell 83:851–857.
10. Borden, K. L. B., M. N. Boddy, J. Lally, N. J. O’Reilly, S. Martin, K. Howe, E. Solomon, and P. S. Freemont.1995. The solution structure of the RING finger domain from the acute promyelocytic leukemia proto-oncoprotein PML. EMBO J. 14:1532–1541.
11. Borden, K. L. B., J. M. Lally, S. R. Martin, N. J. O’Reilly, E. Solomon, and P. S. Freemont.1996. In vivo and in vitro characterization of the B1 and B2 zinc-binding domains from the acute promyelocytic leukemia protoonco-protein PML. Proc. Natl. Acad. Sci. USA 93:1601–1606.
12. Borden, K. L. B., and P. S. Freemont. 1996. The RING finger domain: a recent example of a sequence-structure family. Curr. Opin. Struct. Biol. 6:395–401.
13. Brownell, J. E., J. Zhou, T. Ranalli, R. Kobayashi, D. G. Edmondson, S. Y. Roth, and C. D. Allis.1996. Tetrahymena histone acetyltransferase A: a homology to yeast Gcn5p linking histone acetylation to gene activation. Cell 84:843–851.
14. Cao, Z., R. M. Umek, and S. L. McKnight. 1991. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 5:1538–1552.
15. Cavailles, V., S. Dauvois, F. L’Horset, G. Lopez, S. Hoare, P. J. Kushner, and M. G. Parker.1995. Nuclear factor RIP140 modulates transcriptional acti-vation by the estrogen receptor. EMBO J. 14:3741–3751.
16. Chakravarti, D., V. J. LaMorte, M. C. Nelson, T. Nakajima, I. G. Schulman, H. Juguilon, M. Montminy, and R. M. Evans.1996. Role of CBP/p300 in nuclear receptor signalling. Nature 383:99–103.
17. Chang, C. J., B. J. Shen, and S. C. Lee. 1995. Autoregulated induction of the acute-phase response transcription factor gene, agp/ebp. DNA Cell Biol. 14:529–537.
18. Chang, C. J., T. T. Chen, H. Y. Lei, D. S. Chen, and S. C. Lee. 1990. Molecular cloning of a transcription factor, AGP/EBP, that belongs to mem-bers of the C/EBP family. Mol. Cell. Biol. 10:6642–6653.
19. Chen, H., R. J. Lin, R. L. Schiltz, D. Chakravarti, A. Nash, L. Nagy, M. L. Privalsky, Y. Nakatani, and R. M. Evans.1997. Nuclear receptor coactivator ACTR is a novel histone acetyltransferase and forms a multimeric activation complex with P/CAF and CBP/p300. Cell 90:569–580.
20. Descombes, P., M. Chojkier, S. Lichtsteiner, E. Falvey, and U. Schibler. 1990. LAP, a novel member of the C/EBP gene family, encodes a liver-enriched transcription activator protein. Genes Dev. 4:1541–1551. 21. Freemont, P. S. 1992. A novel zinc finger coiled-coil domain in a family of
nuclear proteins. Trends Biochem. Sci. 17:344–345.
22. Friedman, J. R., W. J. Fredericks, D. E. Jensen, D. W. Speicher, X.-P. Huang, E. G. Neilson, and F. J. Rauscher III.1996. KAP-1, a novel core-pressor for the highly conserved KRAB repression domain. Genes Dev. 10:2067–2078.
23. Goddard, A. D., J. Borrow, P. S. Freemont, and E. Solomon. 1991. Charac-terization of a zinc finger gene disrupted by the t(15;17) in acute promyelo-cytic leukemia. Science 254:1371–1374.
24. Gronemeyer, H. 1991. Transcription activation by oestrogen and progester-one receptors. Annu. Rev. Genet. 25:89–123.
25. Guiochon-Mantel, A., J. F. Savouret, F. Quignon, K. Delabre, E. Milgrom, and H. D. The.1995. Effect of PML and PML-RAR on the transactivation properties and subcellular distribution of steroid hormone receptors. Mol. Endocrinol. 9:1791–1803.
26. Halachmi, S., E. Marden, G. Martin, H. MacKay, C. Abbondanza, and M. Brown.1994. Estrogen receptor-associated proteins: possible mediators of hormone-induced transcription. Science 264:1455–1458.
27. Hanstein, B., R. Eckner, J. DiRenzo, S. Halachmi, H. Liu, B. Searcy, R. Kurokawa, and M. Brown.1996. p300 is a component of an estrogen recep-tor coactivarecep-tor complex. Proc. Natl. Acad. Sci. USA 93:11540–11545. 28. Heery, D. M., E. Kalkhoven, S. Hoare, and M. G. Parker. 1997. A signature
motif in transcriptional co-activators mediates binding to nuclear receptors. Nature 387:733–736.
29. Hong, H., K. Kohli, A. Trivedi, D. L. Johnson, and M. R. Stallcup. 1996. GRIP1, a novel mouse protein that serves as a transcriptional coactivator in yeast for the hormone binding domains of steroid receptors. Proc. Natl. Acad. Sci. USA 93:4948–4952.
30. Janknecht, R., and T. Hunter. 1996. Transcriptional control: versatile mo-lecular glue. Curr. Biol. 6:951–954.
31. Kamei, Y., L. H. Xu, T. Heinzel, J. Torchia, R. Kurokawa, B. Gloss, S.-C. Lin, R. Heyman, D. Rose, C. Glass, and M. Rosenfeld.1996. A CBP inte-grator complex mediates transcriptional activation and AP-1 inhibition by nuclear receptors. Cell 85:403–414.
32. Kanno, M., M. Hasegawa, A. Ishida, K. Isono, and M. Taniguchi. 1995. Mel-18, a polycomb group-related mammalian gene, encodes a transcrip-tional negative regulator with tumor suppressive activity. EMBO J. 14:101– 107.
33. Kim, S.-S., Y.-M. Chen, E. O’Leary, R. Witzgall, M. Vidal, and J. V. Bon-ventre.1996. A novel member of the RING finger family, KRIP-1, associates with the KRAB-A transcriptional repressor domain of zinc finger proteins. Proc. Natl. Acad. Sci. USA 93:15299–15304.
34. Klein, E. S., D. DiLorenzo, G. Posseckert, M. Beato, and G. M. Ringold. 1988. Sequences downstream of the glucocorticoid regulatory element me-diate cycloheximide inhibition of steroid induced expression from the rat a1-acid glycoprotein promoter: evidence for a labile transcription factor. Mol. Endocrinol. 2:1343–1351.
35. Laurent, B. C., I. Treich, and M. Carlson. 1993. The yeast SNF2/SWI2 protein has DNA-stimulated ATPase activity required for transcriptional activation. Genes Dev. 7:583–591.
36. Le Douarin, B., C. Zechel, J.-M. Garnier, Y. Lutz, L. Tora, B. Pierrat, D. Heery, H. Gronemeyer, P. Chambon, and R. Losson.1995. The N-terminal part of TIF1, a putative mediator of the ligand-dependent activation function (AF-2) of nuclear receptors, is fused to B-raf in the oncogenic protein T18. EMBO J. 14:2020–2033.
37. Le Douarin, B., A. L. Nielsen, J.-M. Garnier, H. Ichinose, F. Jeanmougin, R.
at NATIONAL TAIWAN UNIV MED LIB on June 17, 2009
mcb.asm.org
Losson, and P. Chambon.1996. A possible involvement of TIF-1a and TIF-1b in the epigenetic control of transcription by nuclear receptors. EMBO J. 15:6701–6715.
38. Lee, J. W., F. Ryan, J. C. Swaffield, S. A. Johnston, and D. A. Moore. 1995. Interaction of thyroid-hormone receptor with a conserved transcriptional mediator. Nature 374:91–94.
39. Meyer, M.-E., H. Gronemeyer, B. Turcotte, M.-T. Bocquel, D. Tasset, and P. Chambon.1989. Steroid hormone receptors compete for factors that medi-ate their enhancer function. Cell 57:433–442.
40. Miau, L. H., C. J. Chang, W. H. Tsai, and S. C. Lee. 1997. Identification and characterization of a nucleolar phosphoprotein, Nopp140, as a transcription factor. Mol. Cell. Biol. 17:230–239.
41. Miau, L. H., C. J. Chang, B. J. Shen, W. H. Tsai, and S. C. Lee. 1998. Identification of heterogeneous nuclear ribonucleoprotein K (hnRNP K) as a repressor of C/EBPb-mediated gene activation. J. Biol. Chem. 273:10784– 10791.
42. Miki, Y., J. Swensen, D. Shattuck-Eidens, P. A. Futreal, K. Harshman, S. Tavtigian, Q. Liu, C. Cochran, L. M. Bennett, W. Ding, et al.1994. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science 266:66–71.
43. Moosmann, K., O. Georgiev, B. Le Douarin, J.-P. Bourquin, and W. Schaf-fer.1996. Transcriptional repression by RING finger protein TIF1b that interacts with the KRAB repressor domain of KOX1. Nucleic Acids Res. 24:4859–4867.
44. Niehof, M., M. P. Manns, and C. Trautwein. 1997. CREB controls LAP/C/ EBPb transcription. Mol. Cell. Biol. 17:3600–3613.
45. Nishio, Y., H. Isshiki, T. Kishimoto, and S. Akira. 1993. A nuclear factor for interleukin-6 expression (NF-IL6) and the glucocorticoid receptor synergis-tically activate transcription of the rata1-acid glycoprotein gene via direct protein-protein interaction. Mol. Cell. Biol. 13:1854–1862.
46. Ogryzko, V. V., R. L. Schiltz, V. Russanva, B. H. Howard, and Y. Nakatani. 1996. The transcriptional coactivators p300 and CBP are histone acetyltrans-ferases. Cell 89:373–380.
47. Onate, S. A., S. Y. Tsai, M.-J. Tsai, and B. W. O’Malley. 1995. Sequence and characterization of a coactivator for the steroid hormone receptor super-family. Science 270:1354–1357.
48. Parker, M. G. 1993. Steroid and related receptors. Curr. Opin. Cell Biol. 5:499–504.
49. Poli, V., F. P. Mancini, and R. Cortese. 1990. IL-6DBP, a nuclear protein involved in interleukin-6 signal transduction, defines a new family of leucine zipper proteins related to C/EBP. Cell 63:643–653.
50. Ratajczak, T., P. M. Williams, D. DiLorenzo, and G. M. Ringold. 1992. Multiple elements within the glucocorticoid regulatory unit of the rata1-acid
glycoprotein gene are recognition sites of C/EBP. J. Biol. Chem. 267:11111– 11119.
51. Rothe, M., M.-G. Pan, W. J. Henzel, T. M. Ayres, and A. V. Goeddel. 1995. The TNFR2-TRAF signaling complex contains two novel proteins related to baculoviral inhibitor of apoptosis proteins. Cell 83:1243–1252.
52. Saurin, A. J., K. L. B. Borden, M. N. Boddy, and P. S. Freemont. 1996. Does this have a familiar RING? Trends Biochem. Sci. 21:208–214.
53. Spencer, T. E., G. Jenster, M. M. Burcin, C. D. Allis, J. Zhou, C. A. Mizzen, N. J. McKenna, S. A. Onate, S. Y. Tsai, M.-J. Tsai, and B. W. O’Malley.1997. Steroid receptor coactivator-1 is a histone acetyltransferase. Nature 389:194– 198.
54. Takahashi, M., Y. Inaguma, H. Hiai, and F. Hirose. 1988. Developmentally regulated expression of a human “finger”-containing gene encoded by the 59 half of the ret transforming gene. Mol. Cell. Biol. 8:1853–1856.
55. Tasset, D., L. Tora, C. Formental, E. Scheer, and P. Chambon. 1990. Distinct classes of transcriptional activating domains function by different mecha-nisms. Cell 62:1177–1187.
56. Thenot, S., C. Henriquet, H. Rochefort, and V. Cavailles. 1997. Differential interaction of nuclear receptors with the putative human transcriptional coactivator hTIF1. J. Biol. Chem. 272:12062–12068.
57. Torchia, J., D. W. Rose, J. Inostroza, Y. Kamei, S. Westin, C. K. Glass, and M. G. Rosenfeld.1997. The transcriptional co-activator pCIP binds CBP and mediates nuclear-receptor function. Nature 387:677–684.
58. Voegel, J. J., M. J. S. Heine, C. Zechel, P. Chambon, and H. Gronemeyer. 1996. TIF2, a 160 KDa transcriptional mediator for the ligand-dependent activation function AF-2 of nuclear receptors. EMBO J. 15:3667–3675. 59. vom Baur, E., C. Zechel, D. Heery, M. J. S. Heine, J. M. Garnier, V. Vivat,
B. Le Douarin, H. Gronemeyer, P. Chambon, and R. Losson.1996. Differ-ential ligand-dependent interactions between the AF-2 activating domain of nuclear receptors and the putative transcriptional intermediary factors mSUG1 and TIF1. EMBO J. 15:110–124.
60. Williams, P., T. Ratajczak, S. C. Lee, and G. M. Ringold. 1991. AGP/EBP (LAP) expressed in rat hepatoma cells interacts with multiple promoter sites and is necessary for maximal glucocorticoid induction of the rat a1-acid glycoprotein gene. Mol. Cell. Biol. 11:4959–4965.
61. Williams, S. C., C. A. Cantwell, and P. F. Johnson. 1991. A family of C/EBP-related proteins capable of forming covalently linked leucine zipper dimers in vitro. Genes Dev. 5:1553–1567.
62. Won, K.-A., and H. Baumann. 1990. The cytokine response element of the rata1-acid glycoprotein gene is a complex of several interacting regulatory sequences. Mol. Cell. Biol. 10:3965–3978.
63. Yeh, S., and C. Chang. 1996. Cloning and characterisation of a specific coactivator, ARA70, for the androgen receptor in human prostate cells. Proc. Natl. Acad. Sci. USA 93:5517–5521.