國立臺灣大學醫學院臨床牙醫學研究所 博士論文
Graduate Institute of Clinical Dentistry College of Medicine
National Taiwan University Doctoral Dissertation
人類醛固酮還原酶家族 1 成員 B10
於口腔鱗狀細胞癌之表現
Expression of Aldo-keto reductase family 1, member B10 (AKR1B10)
in oral squamous cell carcinomas
柯惠馨 Hui-Hsin Ko
指導教授:郭彥彬教授、鄭世榮副教授 Advisor: Professor Mark Yen-Ping Kuo Associate Professor Shih-Jung Cheng
中華民國 107 年 6 月
謝誌
首先,誠摯的感謝指導教授郭彥彬教授以及鄭世榮副教授的悉心指導,時 時提醒並指點我正確的方向,讓我能夠順利的完成博士研究論文,也讓我受益良 多。老師們對學問的嚴謹看待,更是我未來在學習上的典範。
感謝研究室的所有同仁們,包括了雅文學姊、淳鈴學妹,在研究技術上的 指導以及糾正,讓我可以克服許多技術上的障礙,順利地完成實驗進度。還有整
個實驗團隊成員:珮瑤、 方瑜 、依青、馨慧 、乃齊、家寧、里奈 等人,因為團
隊成員們的團結,才能幫助我完成這一段小小的里程碑,期許自己未來也能幫助 學弟妹,讓整個團隊能得到更多、更好的成績。
另外,也要感謝臨床上的其他老師:郭生興醫師、李正喆醫師、章浩宏醫 師以及陳信銘醫師,在臨床上給予的所有指導以及包容,我才能排除萬難的讓自 己能順利完成臨床工作以及研究工作;還有一直當我的靠山的伊婷學姊,給予我
各方面的支持,讓我可以順利的拿到學位 。
最後也要感謝我的父母、家人以及我的好友彥宏對我的包容與體諒,讓我 可以順利的完成自己想做的事情,在我透不過氣時,家人與好友總是我最強的後 盾,支撐著我走過一次又一次的挫折,才有這個博士論文的完成。倘若這是一份 美麗的榮耀,也因為有你們,才能夠那麼的絢爛。
中文摘要
背景:
人類醛固酮還原酶家族1 成員 B10 (Aldo-keto reductase family 1, member B10, AKR1B10)的量增加。醛糖還原酶(AKR)家族以 NADPH 為輔酶將來自食物和細胞
內的脂肪類醛、酮和芳香類物質,還原成相對應的醇。AKR1B10 蛋白分子量為
36 kDa,最早在人類肝癌的研究中發現。AKR1B10 是一種分泌型蛋白,可經由分
泌型 lysosome 分泌到細胞外。AKR1B10 在人的大多數組織並不表現或低度表現,
它主要表現於小腸和結腸。最近研究指出,AKR1B10 的表現與肺癌、乳癌、肝癌、
胰臟癌的發生及發展有關。但並無文獻提到有關 OSCC 患者,其癌組織中
AKR1B10 蛋白值濃度及唾液中 AKR1B10 蛋白分泌量,與 OSCC 患者預後之相關
性。因此,本研究進一步檢測 OSCC 患者癌組織中 AKR1B10 蛋白值濃度及唾液
中AKR1B10 蛋白分泌量,是否和 OSCC 患者之腫瘤發展、復發及存活有關。
材料及方法:
本研究第一部分利用口腔癌病患之組織病理切片進行免疫組織化學染色法 (immunohistochemistry(IHC)),測量 77 例 OSCC 癌組織及 32 例正常口腔黏膜 (normal mucosa) 中 AKR1B10 蛋白表現量。AKR1B10 蛋白表現量以染色記分(LSs,
定義為染色強度與染色指標的乘積)定義,染色記分越高表示 OSCC 組織中
AKR1B10 蛋白表現量愈多。ANOVA、卡方檢定(Chi-square test)、Kaplan-Meier 存活率方法及Cox proportional hazard regression model,來分析癌組織中 AKR1B10
蛋白表現量與 OSCC 患者臨床病理參數及存活率之相關性。本研究第二部分利用
酵素連結免疫吸附法(enzyme-linked immunosorbent assay, ELISA),探討 86 位 OSCC 患者及 30 位具正常口腔黏膜者,其唾液中 AKR1B10 蛋白分泌量。同時利 用 ANOVA、 卡 方 檢 定 、 Kaplan-Meier 存 活 率 方 法 及 Cox proportional hazard regression model,來分析唾液中 AKR1B10 蛋白分泌量與 OSCC 患者臨床病理參 數及存活率之相關性,並試圖尋找預測存活時間的獨立預後因子。
結果:
本研究第一部分發現,AKR1B10 蛋白的染色記分(labeling score, LS)較高和
患者有較大腫瘤 (P = 0.041)、有較高臨床分期 (P = 0.037) 及有嚼食檳榔之口腔習
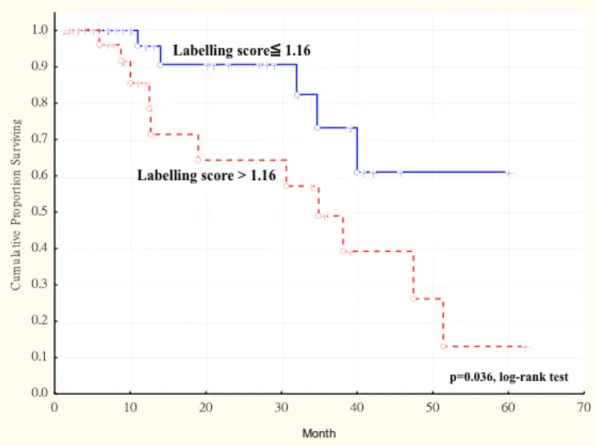
慣 (P = 0.025) ,有統計學上有意義之相關。 以 Cox regression model 進行多變數分 析發現,腫瘤大小 (P = 0.013)、AKR1B10 labeling score> 1.16 (P = 0.001)為影響 OSCC 患者存活時間的獨立預測因子。Kaplan-Meier 存活分析發現, AKR1B10 labeling score> 1.16 比 AKR1B10 labeling score ≤ 1.16 之 OSCC 患者,有較差的無 復發存活率 (log-rank test, P = 0.036)。
本研究第二部分發現,OSCC 患者唾液中 AKR1B10 蛋白分泌量的平均值,
比具正常口腔黏膜者的平均值高,且具統計學上有意義之差別 (25.01 ± 32.96
pg/mL, p < 0.001)。OSCC 患者唾液中 AKR1B10 蛋白分泌量的平均值較高和較大 腫瘤 (P = 0.033)、有較高臨床分期 (P = 0.030) 及有嚼食檳榔之口腔習慣 (P = 0.001)
有統計學上有意義之相關。Cox regression model 進行多變數分析發現,唾液中
AKR1B10 蛋白分泌量為影響存活時間的獨立預測因子(P = 0.002)。Kaplan-Meier 存活分析發現,唾液中AKR1B10 蛋白 > 646 pg/mL 比 AKR1B10 蛋白≦646 pg/mL 者,會有較差的無復發存活率 (log-rank test,P = 0.026)。
結論:
本 研 究 發 現 ,OSCC 患 者 組 織 中 AKR1B10 蛋 白 質 表 現 量 及 唾 液 中 AKR1B10 蛋白量,可以預測 OSCC 之腫瘤大小、臨床分期及其無復發存活率;同
時,組織中AKR1B10 蛋白質表現量及唾液中 AKR1B10 蛋白量與 OSCC 患者嚼食
檳榔之口腔習慣呈現正相關。因此 OSCC 患者組織中 AKR1B10 蛋白質表現量及
唾液中 AKR1B10 蛋白量,可以當作 OSCC 患者預後的指標。
關鍵詞: 人類醛固酮還原酶家族 1 成員 B10、口腔癌、唾液
Abstract
Background/Purpose:Aldo-keto reductase family 1 member B10 (AKR1B10) is a member of the aldo- keto reductase superfamily. This member can efficiently reduce aliphatic and aromatic aldehydes, and it is less active on hexoses. AKR1B10 was originally identified as an up- regulated protein in hepatocellular carcinomas in 1998.
AKR1B10 is a 36-kDa cytosolic reductase and secreted through a lysosome-mediated nonclassical pathway. In normal human tissues, AKR1B10 is primarily expressed in epithelial tissues of digestive tract such as colon, small intestine and stomach and barely detected in other tissues. Elevated expression of AKR1B10 is found in a wide variety of human tumors, such as hepatocellular, non-small cell lung, breast, cervical, and pancreatic carcinomas. However, there have been no previous reports on AKR1B10 protein expression in oral SCCs. In this study, we investigated whether the levels of AKR1B10 in the saliva and cancer tissue of OSCC patients can be used as a prognostic marker.
Materials and methods:
In the first part of this study, we used immunohistochemical staining (IHC) to detect the AKR1B10 protein levels in 77 OSCC and 32 specimens of normal oral mucosa (NOM). The grade of AKR1B10 protein expression was evaluated semiquantitatively according to the proportion of positive-staining cells to the total number of counted cancer or epithelial cells (labeling index) and the staining intensity. In the second part of this study, serum samples were obtained from 72 OSCC patients and from 30 normal control subjects. Salivary AKR1B10 protein levels were determined by enzyme-linked immunosorbent assay (ELISA). The correlation between AKR1B10 LSs and salivary
AKR1B10 level in OSCCs and clinicopathological parameters of OSCC patients was analyzed by Student’s t-test, ANOVA, Kaplan-Meier analyses and Multivariate Cox regression analyses. A p-value of less than 0.05 was considered statistically significant.
Results:
In the first part of this study, we found that the AKR1B10 labeling score (LS) for OSCCs (1.16±1.14) was significantly higher than that for normal oral mucosa (0.10±0.23, p < 0.0001). High expression of AKR1B10 significantly correlated with large tumor size (p = 0.041), advanced TNM stage (p = 0.037) and patient’s areca quid chewing habit (p
= 0.025). Multivariate analysis revealed that high AKR1B10 LS>1.16 (Hazard ratio 3.647, P = 0.001) significantly correlated with mortality.
In the second part of this study, we found that the mean salivary AKR1B10 levels were significantly higher in the OSCC patients than in the normal controls (p < 0.001).
Higher salivary AKR1B10 levels were significantly associated with larger tumor size, more advanced clinical stage and areca quid chewing habit. Result also showed that OSCC patients with a higher salivary AKR1B10 level (> 646 pg/mL) had a significantly poorer survival than those with a lower (≤ 646 pg/mL) salivary AKR1B10 level (p = 0.026).
Conclusion
This study found that AKR1B10 protein level in OSCC tissues and salivary AKR1B20 protein level in OSCC patients could be used to predict the tumor size, clinical stage and the prognosis of OSCC patients. Therefore, we conclude that AKR1B20 protein level in OSCC tissues and salivary AKR1B10 protein level in OSCC patients may be valuable biomarkers for prediction of progression and prognosis of OSCC in Taiwan.
Key words: Aldo-keto reductase family 1 member B10 (AKR1B10)、oral cancer、
Saliva
TABLE OF CONTENTS
口試委員會審定書………... i
謝誌……….. ii
中文摘要………. iii
Abstract……….. v
I. Introduction and literature review ... 1
1.1 Overview of oral squamous cell carcinoma ... 2
1.2 Tumor microenvironment in head and neck cancer ... 11
1.3 ROS homeostasis and metabolism ... 17
1.4 Aldo-Keto Reductase (AKR) family ... 25
II. Materials and methods ... 39
Part I: AKR1B10 level in OSCC tissue ... 39
2.1.1 Patients recruitment and oral cancer specimens ... 39
2.1.2 Immunohistochemistry staining ... 40
2.1.3 Statistical analysis ... 41
Part II: Salivary AKR1B10 secrection ... 41
2.2.1 Patients recruitment and saliva samples ... 41
2.2.2 Saliva collection ... 42
2.2.3 AKR1B10 measurement in saliva ... 43
2.2.4 Statistical analysis ... 43
III. Results ... 44
Part I: Expression of AKR1B10 as an independent marker for poor prognosis in human oral squamous cell carcinoma ... 44
3.1.1 Patient characteristics ... 44
3.1.2 AKR1B10 levels in oral cancer tissues ... 44
3.1.3 Correlation between the AKR1B10 level in OSCC samples and clinicopathological parameters of OSCC patients ... 45
3.1.4 Survival analyses ... 45
Part II: Increased salivary AKR1B10 level is significantly associated with progression and poor prognosis of oral squamous cell carcinoma ... 46
3.2.1 Saliva AKR1B10 levels in normal and cancer patients ... 46
3.2.2 Correlation between the saliva AKR1B10 protein levels and clinicopathological parameters of OSCC patients ... 46
3.2.3 Survival analysis ... 47
IV. Discussion ... 48
Part I: Expression of AKR1B10 as an independent marker for poor prognosis in human oral squamous cell carcinoma ... 48
Part II: Increased salivary AKR1B10 level is significantly associated with progression and poor prognosis of oral squamous cell carcinoma ... 52
V. Conclusion ... 56
VI. Tables ... 58
VII. Figures ... 65
VIII. References ... 73
IX. Appendix ... 101
B1: Publication in Dissertation ... 102 B2: Other Publication related to Dissertation ... 102 B3: Publication (2015-2018) ... 102
Tables Index
Table 1. 1Correlation between the AKR1B10 labelling score (LS) in oral squamous cell carcinoma (OSCC) samples and clinicopathological parameters of 77 OSCC patients ... 58 Table 1. 2Multivariate survival analyses of the AKR1B10 labelling score (LS) and
clinicopathological parameters in patients with oral squamous cell carcinoma by Cox proportional hazard regression model ... 60 Table 2. 1The mean AKR1B10 protein concentration in oral squamous cell carcinoma
(OSCC) patients and normal patient tissue samples ... 61 Table 2. 2Correlation between the mean saliva AKR1B10 expression and
clinicopathological parameters of 86 OSCC patients ... 62 Table 2. 3 Multivariate overall survival analyses of the AKR1B10 protein expression
and clinicopathological parameters in patients with oral squamous cell carcinoma by using Cox proportional hazard regression model ... 64
Figure Index
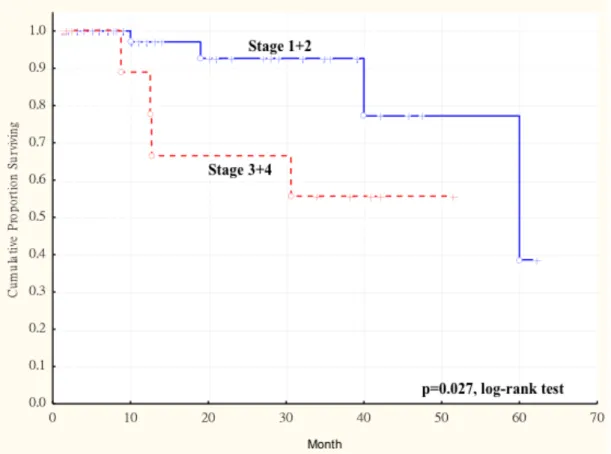
Figure 1. 1 Immunohistochemical staining for AKR1B10. (A, B) Normal buccal mucosa showing negative or weakly positive in few basal layers of the epithelial cells. (C, D) Squamous cell carcinoma demonstrating strongly positive in the peripheral layers of tumor islands with moderately or weak staining cells in the central areas. (Original magnification; A, C 100, and B, D 400 x) ... 65 Figure 1. 2 Kaplan-Meier survival curve showing relation between AKR1B10
expression in primary tumors and survival in 77 patients with oral squamous cell carcinoma (OSCC). The duration of survival was measured from the beginning of treatment to the time of death (complete) or the last follow-up (censored). The cumulative survival for OSCC patients with AKR1B10 LS > 1.16 was significantly lower than that for OSCC patients with AKR1B10 LS ≦ 1.16 (P = 0.036, log-rank test). ... 66 Figure 1. 3 Kaplan-Meier survival curve showing relation between AKR1B10
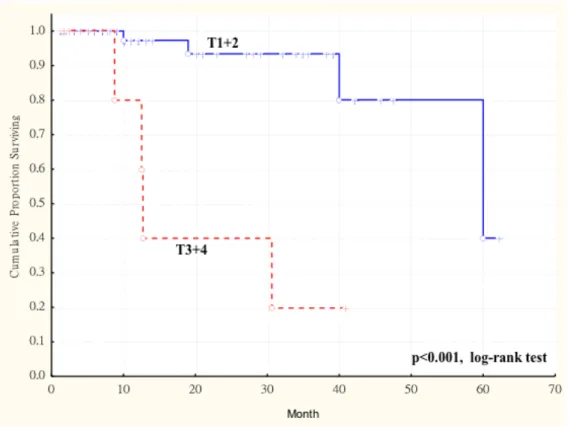
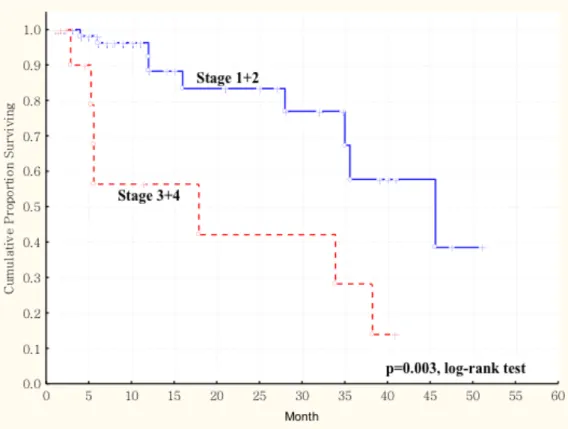
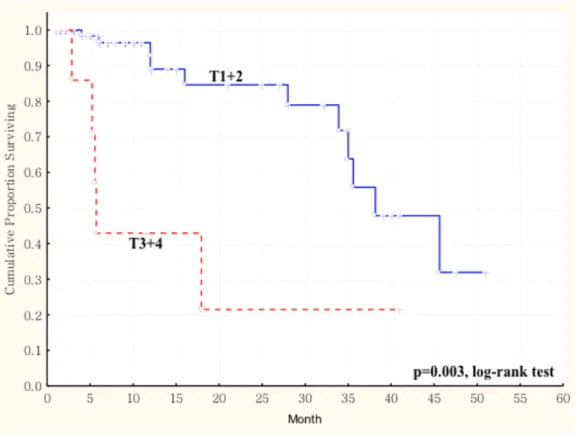
expression in primary tumors and survival in 77 patients with oral squamous cell carcinoma (OSCC). The duration of survival was measured from the beginning of treatment to the time of death (complete) or the last follow-up (censored). The cumulative overall survival was significantly poorer in OSCC patients with higher AKR1B10 LS and larger tumor size (more than 4cm, T3 and T4) than these patients with smaller tumor size (less than 4cm, T1 and T2) (p < 0.003, log-rank test). ... 67 Figure 1. 4 Kaplan-Meier survival curve showing relation between AKR1B10
expression in primary tumors and survival in 77 patients with oral squamous cell carcinoma (OSCC). The duration of survival was measured from the beginning of treatment to the time of death (complete) or the last follow-up (censored). The cumulative overall survival was significantly poorer in OSCC patients with higher AKR1B10 LS and late cancer stage (stage III and stage IV) than these patients with early cancer stage (stage I and stage II) (p < 0.003, log-rank test). ... 68 Figure 2. 1 Kaplan-Meier survival curve showing a significant relation between
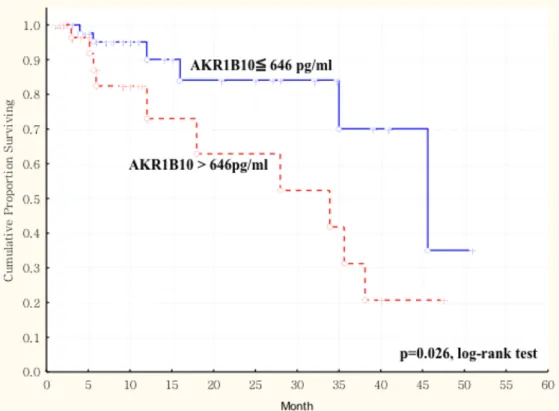
AKR1B10 protein level in 86 patients with oral squamous cell carcinoma (OSCC).
The duration of overall survival was measured from the beginning of surgery to the time of death (complete) or the last follow-up (censored). The cumulative overall survival for OSCC patients with AKR1B10 protein level > 646 pg/mL was significantly poorer than that for OSCC patients with ≦ 646 pg/mL (p = 0.026, log-rank test). ... 69 Figure 2. 2 Kaplan-Meier survival curve showing a significant relation between saliva
AKR1B10 expression in 86 patients with oral squamous cell carcinoma (OSCC) and the tumor size of these 86 OSCC patients. The duration of overall survival was measured from the beginning of surgery to the time of death (complete) or the last follow-up (censored). The cumulative overall survival was significantly poorer in OSCC patients with saliva AKR1B10 expression and larger tumor size (more than 4 cm, T3 and T4) than these patients with smaller tumor size (less than 4cm, T1 and T2) (p < 0.003, log-rank test). ... 70
Figure 2. 3 Kaplan-Meier survival curve showing a significant relation between saliva AKR1B10 expression in 86 patients with oral squamous cell carcinoma (OSCC) and the tumor size of these 86 OSCC patients. The duration of overall survival was measured from the beginning of surgery to the time of death (complete) or the last follow-up (censored). The cumulative overall survival was significantly poorer in OSCC patients with saliva AKR1B10 expression and late cancer stage (stage III and stage IV) than these patients with early cancer stage (stage I and stage II) (p <
0.003, log-rank test). ... 71 Figure 2. 4 Kaplan-Meier survival curve showing relation between salivary AKR1B10
protein levels and overall survival in 76 patients with early stage (T1/T2) OSCC.
The duration of survival was measured from the beginning of surgery to the ti me of death (complete) or the last follow-up (censored). The cumulative survival for patients with a higher salivary AKR1B10 protein level (> 646 pg/ml) was
significantly lower than those with a lower (≦ 646 pg/ml) salivary AKR1B10 protein level (log-rank test, p = 0.046). ... 72
I. Introduction and literature review
Oral squamous cell carcinoma (OSCC) is the eighth most common cancer internationally (World Health Organization 2014). In Taiwan, OSCC ranks as the fifth most common cancer for both sexes and accounts for the fourth most prevalent cancer in males (Taiwan Ministry of Health and Welfare 2016).
Standard treatment for early-stage OSCC is surgical resection of the tumor, whereas advanced-staged tumors are treated with surgery followed by concurrent chemoradiation. Despite recent advances in imaging, surgery, radiation, and chemotherapies, the overall survival has improved by only 15% and 5% in the last 50 and 20 years, respectively (Chinn et al., 2015). This suggests an urgent need for a novel biomarker to predict the progression, recurrence and prognosis of OSCCs.
Aldo-keto reductase family 1 member B10 (AKR1B10), also known as aldose reductase-like-1, belongs to a superfamily of NADPH-dependent aldo-keto reductases, which is implicated in xenobiotic detoxification, cell proliferation, carcinogenesis, and cancer therapeutics (Cao et al., 1998; Penning 2015). Clinically, elevated expression of AKR1B10 is found in non–small cell lung, hepatocellular, nasopharyngeal, cervical, pancreatic, and breastcarcinomas (Fukumoto et al., 2005; Yoshitake et al., 2007; Ma et al., 2012; Chung et al., 2012; Jin et al., 2016; He et al., 2016). Furthermore, it has been considered a valuable prognostic marker for hepatocellular (Jin et al., 2016), cervical (Yoshitake et al., 2007), and breast (Ma et al., 2012) cancers. However, there have been no previous reports on AKR1B10 protein expression in OSCCs.
In this study, we evaluated whether the AKR1B10 level in OSCC surgical specimen measured by immunochemical staining and the salivary AKR1B10 protein
levels in OSCC patients measured by the enzyme-linked immunosorbent assay (ELISA) could be valuable biomarkers to predict the therapeutic effect, progression, recurrence and prognosis of OSCC patients.
1.1 Overview of oral squamous cell carcinoma
1.1.1 Epidemiology of global oral cancers
Oral cancer is the sixth most common cancer worldwide, with a high prevalence in South Asia. In 2012, 300,400 new cases of OSCC were diagnosed worldwide, with 12,170 deaths occurring annually (World Health Organization 2014). The highest incident rates of oral cavity cancer are found in Melanesia, South-Central Asia, and Central and Eastern Europe and the lowest in Africa, Central America, and Eastern Asia for both males and females. Majority (90%) of the cases reported of oral cancer is attributed to tobacco consumption in various forms in these regions (Johnson et al., 2011) Besides smoking, alcohol consumption, smokeless tobacco use, and HPV infections are also the major risk factors for oral cavity cancer, with smoking and alcohol drinking having synergistic effects (Blot et al., 1988). Worldwide, smoking accounts for 42% of deaths from cancers of the oral cavity (including the pharynx) and heavy alcohol consumption for 16% of the deaths; the corresponding percentages in high-income countries are about 70% and 30%, respectively (Danaei et al., 2005) Recently, the rise in the incidence rate of oral cancer in Taiwan may have been in part due to the increased consumption of areca quid and alcohol (Ho et al., 2002). Oral cavity cancer mortality rates among males decreased significantly in most countries, including those of Europe and Asia, over the past decades (Garavello et al., 2010). However, a low 5-year survival rate of 50% is
globally seen among patients with oral cancer (Greenlee et al., 2001). Despite oral cavity being accessible for examination, individuals report to a clinician only in later stages of malignancy, thus no improvement in survival rate for oral cancer over the decades have been observed (Neville et al., 2002). The mortality rate increase in females in most European countries and in several Eastern European countries, including Hungary and Slovakia (Garavello et al., 2010).
This contrasts with the decreasing trends at all ages in both males and females in the United States and United Kingdom, where the tobacco epidemic began and declined earlier (Garavello et al., 2010). A change in trend for incidence rates of oral cancer sites related to HPV infections, such as the oropharynx, tonsil, and base of the tongue, are increasing in young adults in the United States and in some countries in Europe, which is hypothesized to be in part due to changes in oral sexual behavior (D’Souza et al., 2009;
Marur et al., 2010).
1.1.2 Epidemiology of oral cancers in Taiwan
With rapid aging of populations, cancer has become the first leading cause of death in Taiwan since 1982 (Chen et al., 2002). In 1982, the incidence rate of head and neck cancer was 5.12 per 100,000 people in males and 1.54 per 100,000 people in females.
In 1991, the incidence rate of head and neck cancer had not much changed, with the incidence rate being 6.02 and 1.51 per 100,000 people in males and females, respectively.
However, in 2003, the incidence rate of head and neck cancer significantly increased to 35.08 and 3.56 per 100,000 people in males and females, an alarming 5.82-fold increase in men and 2.35-fold increase in woman in a decade was found (Chen et al., 2008). In
100,000 people in males. And since 2006, oral cavity squamous cell carcinoma (OSCC) had become and kept the 6th most common cancer in Taiwan and the 4th most common cancer in Taiwanese men. Similarly, the mortality rate also increased significantly, from 4.25 per 100,000 in 1995 to 9.6 per 100,000 in 2006, a 2.26-fold increase in the past decade.
There has been a trend toward younger age at diagnosis of head and neck cancer over time. From 1989 to 1993, the peak of incidence rate was for people aged 50–59 years, but this shifted to ages 40–49 years between 1993 and 2000. A similar trend was also reported in lower age at diagnosis of head and neck cancer over time. During the period between 1991 and 1994, mortality rate peaked at age 50–59 years, but shifted to age 40–
49 years between 1999 and 2002. These data are consistent with other regional reports from northern and southern Taiwan. In 2006, the median age at death from head and neck cancer was 54 years compared with 69 years in other forms of cancer. Gender differences in head and neck cancer have been described, with a marked male predominance. A study analyzing 703 OSCC patients between 1985 and 1996 in southern Taiwan found a 51:1 male-to-female ratio (Chen et al., 1999) The overall 5-year survival rate in patients with head and neck cancer is one of the lowest among common malignant neoplasms and has not significantly changed during the last two decades. Cancer clinical stage is the major determinant of survival rate. The 5-year survival rates of oral cavity cancer patients in stages I, II, III and IV are 72–90%, 39–85%, 27–70% and 12–50%, respectively (Ko et al., 1995; Liao et al., 2007; Liao et al., 2008). About 50% or more of OSCC patients were found to have their tumours ranked as stage III or stage IV during their first visit to the medical center in Taiwan, leading to a low overall 5-year survival rate, despite recent advances in surgery, radiotherapy and chemotherapy (Lo et al., 2003). However, survival rates for head and neck cancer are significantly influenced by tumor size, lymph node
involvement, distant metastasis, tumor differentiation and areca quid chewing (Liao et al., 2004; 2008). Approximately, 95% of oral cancer is oral squamous cell carcinoma. OSCC is associated with chronic irritant factors such as tobacco, smoking, alcohol and betel quid (BQ) use. While cigarette smoking and alcohol drinking are the major risk factors in Western countries, Areca quid chewing independently contributes to the risk of head and neck cancer, and the estimated prevalence of areca quid chewing in Taiwanese patients (Boyle et al., 1990; Ko et al., 1995). Almost 2.5 million people are BQ users in Taiwan.
A higher rate of incidence of OSCC and its mortality were found to be associated with the increasing prevalence rate of betel chewing in this area. In south Taiwan, betel chewing has been generally accepted as a social custom or behavior. A cumulative effect from areca quid chewing, alcohol drinking and tobacco smoking has been observed, with a 123-fold increased risk of oral cancer when the three risk factors are present (Ko et al., 1995). With regard to the anatomical location of oral cavity cancers, approximately 30–
40% of all cases occur in the tongue or in the buccal mucosa. Altogether, lesions at these sites account for approximately 70% of all oral cavity malignancies (Ko et al., 1995; Chen et al., 1999; Liao et al., 2008).
1.1.3 Risk factors related to oral cancers
Oral cancer was known to be associated with cigarette smoking, excessive alcohol consumption, areca quid chewing, viral or fungal infection (candida, human papilloma virus (HPV), Herpes simplex virus, etc.), nutrition deficiency (Plummer-Vinson Syndrome, Vitamin A deficiency, Vitamin C deficiency, etc.), family hereditary, immunodeficiency, gene mutation, environment factor, such as UV exposure (Pukkala et al., 1997) and so on.
Recent 2003 IARC monograph declared areca quid chewing or even areca nut alone, to be a Group 1 carcinogen. The Taiwanese chewers commonly use fresh, unripe betel fruit with slaked lime as an essential ingredient. The composition of areca quid differs geographically; the areca quid used in Taiwan contains betel leaf, areca nut and slaked lime, and Piper betel inflorescence. Other substances, particularly spices, including tobacco, cardamom, saffron, cloves, aniseed, turmeric, mustard or sweeteners, are added according to local preferences (Yang et al., 2001; Yang et al., 2005). Piper betel inflorescence contains high concentrations of hydroxychavicol and safrole, whereas arecoline, a major areca nut alkaloid, is considered to be the most important carcinogen in the areca nut.
Arecoline has been shown to induce structural chromosomal aberration, sister chromatid exchange and micronuclei formation in different cell types (Shirname et al., 1983; Jeng et al., 2001). Studies in human oral cancer cells have shown that exposure to arecoline or areca nut extract (ANE) results in growth arrest in the late S and G2/M phases (Lee et al., 2006). Piper betel inflorescence, which contains safrole, is a unique ingredient of areca quid in Taiwan. Safrole–DNA adducts have been suggested to play an important role in oral carcinogenesis (Chen et al., 1999). A further report has provided evidence that alkaline-induced DNA damage generated by chewing areca quid may be derived from tobacco, and reactive oxygen species may be involved in generating this DNA damage (Wu et al., 2005). Approximately 50% of patients who were betel quid chewers are also alcohol drinkers and tobacco smokers. (Kuo et al., 1994; Liu et al., 1996). These findings provide a molecular explanation for the synergistic effect of areca quid chewing and tobacco smoking in the development of head and neck cancer in Taiwan.
1.1.4 Precancerous conditions and lesions
Many OSCCs develop from premalignant conditions of the oral cavity (Silverman 1968; 1984). The World Health Organization classifies oral precancerous/potentially malignant disorders into 2 general groups.A precancerous condition is “a generalized state associated with significantly increased risk of cancer.” The precancerous conditions include submucous fibrosis, lichen planus, epidermolysis bullosa, and discoid lupus erythematosus. A precancerous lesion is “a morphologically altered tissue in which oral cancer is more likely to occur than its apparently normal counterpart.” These precancerous lesions include leukoplakia, erythroplakia, and the palatal lesions of reverse smokers.
The Taiwanese national cancer screening program began in 1985 by providing oral mucosa screening programs in oral cancer high-risk groups. At the same time, patients’ self-awareness regarding the prevention of OSCC that had been intensively propagated through the mass media in the recent years (Lo et al., 2003; Chen et al., 2012) . The results strongly suggest the importance and effective outcomes from early diagnosis and treatment of premalignant lesions (Shiau et al., 1979; Speight et al., 1992; Scully 1995; van der Waal et al., 1997) and OSCC.
Intervention should be based on histopathological features of a biopsy of the lesions. At present, histological criteria (presence and degree of dysplasia) represent the gold standard in precancerous lesion risk evaluation (Blot et al., 1988). The latest WHO classification (Barnes et al., 2005) recommends a more objective gradings. The criteria for grading of oral epithelial dysplasia are listed as follows:
Mild dysplasia (grade I) shows proliferation or hyperplasia of cells of the basal
Cytological atypia is generally slight with only mild pleomorphism of cells or nuclei.
Mitoses are not prominent, and when present are usually basally located and normal.
Architectural changes are minimal.
In moderate dysplasia (grade II) there is a proliferation of atypical cells extending into the middle one-third of the epithelium. The cytological changes are more severe than in mild dysplasia and changes such as hyperchromatism, and prominent cell and nuclear pleomorphism may be seen. Increased and abnormal mitoses may be present, but these are usually located in the basal layers. Architectural changes may be seen in the lower half of the epithelium where there may be loss of basal polarity and hyperplasia leading to bulbous rete pegs. However, stratification and maturation are relatively normal, often with hyperkeratosis.
Severe dysplasia (grade III) demonstrates abnormal cell proliferation from the basal layer into the upper third of the epithelium. Cytological and architectural changes can be very prominent. All the changes seen in mild and moderate dysplasia are seen but in addition there is marked pleomorphism often with abnormally large nuclei with prominent or even multiple nucleoli. Prominent and suprabasal mitoses are usually evident and abnormal tripolar or star-shaped forms may be seen. Apoptotic bodies may also be prominent. Architectural changes are severe, often with complete loss of stratification and with deep abnormal keratinization and even formation of keratin pearls.
Abnormal forms of rete pegs are usual and bulbous rete pegs are regarded as particularly significant in the diagnosis of severe dysplasia. Abnormal shaped rete pegs may also be seen, with lateral extensions or small branches. These are quite abnormal and may be the earliest signs of invasion. Occasional lesions may show prominent acantholysis with severe disruption of the architecture. Although the epithelium may be thickened, severe
Carcinoma in situ is the worse form of epithelial dysplasia and is characterized by full thickness cytological and architectural changes. In the oral cavity, such changes are rare, and often, even in the presence of the most severe atypia, there is still an intact keratinized surface layer. Carcinoma in situ is thought by some to be a premalignancy but others regard it as evidence of actual malignant change but without invasion.
These premalignant changes and lesions of the head and neck carry a high risk to progress towards malignant transformation. Severe epithelial dysplasia has an overall malignant transformation rate of about 16% but studies show a wide range of 7% – 50%(Mincer et al., 1972; JBanoczy et al., 1976; Silverman et al., 1984; Vedtofte et al., 1987; Bouquot et al., 1988; Lumerman et al., 1995; Pindborg et al., 1997; Schepman et al., 1998; Jaber et al., 2003). Moderate dysplasia has a malignant transformation potential of 3% – 15%, whereas mild epithelial dysplasia shows a very low risk (less than 5%).
Therefore, all oral premalignant changes and lesions should be submitted to microscopic analysis before any definite treatment. Long-term follow-up of patients with premalignant oral lesions is highly recommended (Chang et al., 2002; Lin et al., 2002).
1.1.5 Clinicopathological classification and treatment modalities of oral squamous cell carcinoma
Currently, TNM staging and pathological grade are the most prevalent and traditional prognostic indicators and basis of individualized treatment. The ‘TNM’
classification of the International Union Against cancer (UICC) relates well to the prognosis and overall survival, earlier the tumor stage, better the prognosis and less complicated is the treatment (Schroeff et al., 2009). Logistic regression analysis revealed
is a growing concern that TNM staging is insufficient to accurately map or classify OSCC, whose biological impact may be related to volume and pathological aggressiveness of disease (Kademani et al., 2005). The WHO grading system (Pindborg et al., 1997) recommends histologic features of OSCC are classified into 3 categories: well- differentiated, moderately-differentiated and poorly-differentiated. This usually depends on the subjective assessment of the degree of keratinization, cellular and nuclear pleomorphism, and mitotic activity (Kademani et al., 2005). The influence of histologic grading as a prognostic factor in OSCC was assessed in 215 patients and was found to be a significant predictor of locoregional failure and tumour recurrence (Kademani et al., 2005).
Surgery is the most well-established mode of initial definitive treatment for a majority of oral cancers, with a longstanding history of being the accepted method of treatment for oral cancers over a century. Introduction of ionizing radiation, following the discovery of radium, became an important means of non-surgical treatment of oral carcinoma. However, in the majority of patients with advanced cancer, radiotherapy is employed in conjunction with surgery, most often offered as post-operative treatment.
Chemotherapy in the management of oral carcinoma was initially considered palliative in the 1950’s, 60’s and 70’s. Nowadays, chemotherapy is also used as a component of multimodality treatment in the neoadjuvant setting, clinical trials of induction chemotherapy with Cis-platinum demonstrated that response to chemotherapy is observed in a significant number of patients (Kademani et al., 2005). In addition, intra- arterial chemotherapy, particularly superselective IA infusion of high-dose cisplatin with systemic neutralization by intravenous sodium thiosulfate(RADPLAT), is a theoretically attractive approach to the treatment of advanced head and neck cancer. intra-arterial infusion of high-dose (Akihiro et al., 2016).
For patients with advanced-staged disease, the current preference for the sequence of combined modality treatment program is surgical resection with immediate appropriate reconstruction followed by post-operative radiation therapy or post-operative concurrent chemoradiotherapy. The observations from two prospective randomized trials of adjuvant chemoradiotherapy have shown that patients who have extracapsular extension of disease in metastatic cervical lymph nodes and those who have positive margins have a significant improvement in local regional control and disease free survival by addition of chemotherapy to postoperative radiation therapy compared to post-operative radiation therapy alone (Bernier et al., 2005). Targeted therapies with epidermal growth factor receptor (EGFR) inhibitors are an active area of investigation at this time. Immunotherapy and gene therapy are also areas of research where further work needs to be done (Shah et al., 2009).
1.2 Tumor microenvironment in head and neck cancer
Squamous cell carcinoma comprises more than 90% of cancers of the head and neck and arises from the squamous lining of the mucosal surfaces of the upper aerodigestive tract, including the oral cavity, pharynx, larynx, and sinonasal tract. Head and neck squamous cell carcinoma (HNSCC) is the sixth most common cancer worldwide, and only 50%–60% of patients are alive at 5 years after diagnosis (Kamangar et al., 2006;
Marur et al., 2008). Treatment can be quite morbid and result in significant functional as well as aesthetic deficits, such as impairment of speech and swallowing and facial deformity. Treatment failure and locoregional recurrence are common and occur in up to 30% of patients and account for the majority of deaths of HNSCC (Leemans et al., 1994). The high rate of local recurrence produced the long-held notion of “condemned
mucosa” or “field cancerization” initially described in the 1950s (Slaughter et al., 1953). It is clear that the notion of the condemned mucosa reflects a “condemned tissue”
composed of the cancerous cells, adjacent epithelial, stromal, and immune cells and their surrounding matrix. Together these elements comprise the tumor microenvironment (TME). From this perspective, the concept that cancer is derived from a single cell type was shifted to carcinogenesis and carcinomas as phenomena that occur in tissues (Albini et al., 2007).
1.2.1 Epithelial-Mesenchymal Transition in tumor microenvironment
The tumor microenvironment plays an important role in facilitating cancer metastasis and may induce the occurrence of epithelial to mesenchymal transition (EMT) in tumor cells. EMT plays crucial roles in the tumor invasion process, causes disruption of cell-cell adherence, loss of apicobasal polarity, matrix remodeling, increased motility and invasiveness in promoting tumor metastasis (Martin et al., 2010; Sun et al., 2010). Recent evidence suggests that a variety of factors in tumor microenvironment can lead to EMT. Inflammation, hypoxia, and stem cells in tumor microenvironment linked with EMT inextricably through complex pathways. Cells that undergo EMT acquire stem cell-like properties and these stem cell-like cells within the tumor known as cancer stem cells (CSCs). CSC may generate tumors through the stem cell processes of self-renewal, tumor initiation and supporting cellular processes, particularly the epithelial-to- mesenchymal transition. The signaling pathways involved in EMT are various, including TGF-b, NF- B, Wnt, Notch, and others. However, a large number of unknown factors and intracellular signaling pathways have been associated with EMT (Kohei 2009).
1.2.2 Metabolism and Cancer
The hallmarks of cancer were the first of the six biological capabilities to be acquired during the multistep development of human tumors. The hallmarks constitute an organizing principle for rationalizing the complexities of neoplastic disease. They include sustaining proliferative signaling, evading growth suppressors, resisting cell death, enabling replicative immortality, inducing angiogenesis, and activating invasion and metastasis. Underlying these hallmarks are genome instability, which generates the genetic diversity that expedites their acquisition, and inflammation, which fosters multiple hallmark functions. Conceptual progress in the last decade has added two emerging hallmarks of potential generality to this list—evading immune destruction and reprogramming of energy metabolism (Hanahan et al., 2000). Metabolism generates oxygen radicals, which contribute to oncogenic mutations. Activated oncogenes and loss of tumor suppressors in turn alter metabolism and induce aerobic glycolysis. Aerobic glycolysis or the Warburg effect links the high rate of glucose fermentation to cancer.
Together with glutamine, glucose via glycolysis provides the carbon skeletons, NADPH, and ATP to build new cancer cells, which persist in hypoxia that in turn rewires metabolic pathways for cell growth and survival. Excessive caloric intake is associated with an increased risk for cancers, while caloric restriction is protective, perhaps through clearance of mitochondria or mitophagy, thereby reducing oxidative stress. Hence, the links between metabolism and cancer are multifaceted, spanning from the low incidence of cancer in large mammals with low specific metabolic rates to altered cancer cell metabolism resulting from mutated enzymes or cancer genes (Dang 2012).
1.2.2.1 Reactive oxygen species in cancer
Based on recent reports, hypoxia may be proposed as a second factor in the initiation of EMT. When tumors grow to a certain size and cancer cells divide uncontrollably, they form larger tumors. As a consequence, there is limited availability of nutrients and oxygen in the microenvironment and cancer cells are exposed to intermittent hypoxic conditions. During hypoxia, mitochondria increase the production of reactive oxygen species (ROS) and evidence indicates that high levels of reactive oxygen species (ROS) in cancer cells can result from increased metabolic activity, mitochondrial dysfunction, peroxisome activity, increased cellular receptor signaling, oncogene activity, increased activity of oxidases, cyclooxygenases, lipoxigenases and thymidine phosphorylase, or through crosstalk with infiltrating immune cells. Accumulation of intracellular ROS in normal cells contributes to the oxidation of various components, including nucleic acids, proteins, and lipids (Gueraud et al., 2010). The oxidative imbalance can cause multiple damages, which usually promote apoptosis in case of overwhelming damages, but also drive to control metabolic reprogramming of cancer cells and other cells in tumor microenvironments. Under normal physiological conditions, the intracellular levels of ROS are steadily maintained to prevent cells from damage. ROS can act as ‘good’ and ‘bad’ molecules, and regulate cellular physiology or induce cytotoxicity depending on the magnitude, duration and site of their generation.
Hence, strategies aimed at altering redox signaling events in tumor cells and intend to disable key antioxidant systems in the presence of ROS inducers might represent promising new anticancer treatments (Harris et al., 2015).
1.2.2.2 Lipid metabolism and cancer progression
In the 1920s, Otto Warburg first published a body of work linking metabolism and cancer through enhanced aerobic glycolysis in (also known as the Warburg effect) that distinguishes cancer from normal tissues (Warburg 1956; Koppenol et al., 2011). The conversion of glucose to lactate, which can occur in hypoxic normal cells, persists in cancer tissues despite the presence of oxygen that would normally inhibit glycolysis. This sustained aerobic glycolysis in certain cancer cells is linked to activation of oncogenes or loss of tumor suppressors (Vander et al., 2009; Koppenol et al., 2011; Levine et al., 2010;
Cairns et al., 2011). Cancer cells also rely on glutamine consumption, which provides carbon and amino-nitrogen needed for amino-acid, nucleotide and lipid biosynthesis (Gaglio et al., 2011). Rewiring of metabolic programmes, such as aerobic glycolysis and increased glutamine metabolism, are crucial for cancer cells to shed from a primary tumor, overcome the nutrient and energy deficit, and eventually survive and form metastases. However, functionally dependent on glucose and glutamine catabolic pathways but commonly disregarded in the past, alterations in lipid-associated pathways encountered in tumors are now well described. Highly proliferative cancer cells show a strong lipid and cholesterol avidity, which they satisfy by either increasing the uptake of exogenous (or dietary) lipids and lipoproteins or over-activating their endogenous synthesis (lipogenesis and cholesterol synthesis). Excessive lipids and cholesterol in cancer cells are stored in lipid droplets(LDs), and high LDs and stored-cholesteryl ester content in tumors (Accioly et al., 2008; Qiu et al., 2015) are now considered as hallmarks of cancer aggressiveness (Bozza et al., 2010; de Gonzalo-Calvo et al., 2015).
1.2.3 Chemoersistance
Chemotherapy is commonly utilized as a therapeutic approach for many cancers.
However, one of major problems occurring during the therapy is the intrinsic or drug resistance of the cancer cells, which leads to malignant transformation and metastasis of tumors. Overcoming intrinsic and acquired drug resistance is a major challenge in treating cancer. Drug refractoriness may exist prior to exposure to chemotherapeutic agents (primary resistance) or may be developed or enhanced during drug treatment (acquired resistance). Previous studies proposed several candidate molecules that play key roles in the mechanisms of chemoresistance, which included: changes in the expression/function of the molecular targets of anticancer drugs, an enhanced ability of cancer cells to repair anticancer drug induced DNA damage, decreased expression/function of pro-apoptotic factors or up-regulation of anti-apoptotic genes, enhanced stress responses and changes in tumor microenvironment(Bozza et al., 2010; de Gonzalo-Calvo et al., 2015). One of the most recognized candidates is an efflux transporter P-glycoprotein. As P-glycoprotein is called multidrug resistance protein, its aberrant expression decreases the metabolic clearance of the drugs through shortening its residence time in the cells (Higgins 2007). The chemoresistance of the cancer cells is also provoked by high expression of an inhibitor of apoptosis (IAP) family members, such as c-IAP1 (Notarbartolo et al., 2004), which is considered to inhibit apoptotic cell death elicited by some chemotherapeutics, such as cisplatin and doxorubicin, through directly interacting with caspases and then abolishing their pro-apoptotic actions (Tirrò et al., 2006). Other resistance-related factors are growth factor receptors and the ubiquitin–proteasome system. Binding of ligands to fibroblast growth factor receptor-1 and -3 activates both signalings of MAP kinase and phosphoinositide 3-kinase (Tomlinson et al., 2012).
Therefore, overexpression of the growth factor receptors may enhance capacities of
resistant cells to proliferate, and protect from fatal damage induced by anticancer drugs.
The ubiquitin–proteasome system, a large multi-subunit complex that cleaves damaged and misfolded proteins, is thought to be activated with cancer chemoresistance. The involvement of proteasomes in the chemoresistance is obvious from data that treatment with proteasome inhibitors MG132 and bortezomib induces multidrug resistance- associated protein 2 in human colon cancers (Loeffler-Ragg et al., 2009; Ebert et al., 2011). In addition, the proteasome modulates the expression of transcription factors, such as NF-κB, p53, c-Jun, and c-Fos, that participate in the mechanisms of cell proliferation and differentiation (Li et al., 2007). Moreover, the ubiquitin–proteasome system appears to regulate cellular susceptibility to death by controlling the balance between pro- apoptotic caspases and anti-apoptotic Bcl and IAPs (Almond et al., 2002). There appear many cross talks between these aspects as shown in the context. For example, the encoding proteins of oncogenes (EGFR-Akt-NF-κB) might modulate the apoptosis- related genes expression, and contribute to EMT, cell stemness and autophagy. The exosome might contain many proteins, which are related to anti-apoptosis and transporter pumps. Additionally, whining tumor microenvironment, cells will appear stem-like once the onset of EMT (EMT-related stemness). Further understanding of chemoresistance signal network would facilitate the establishment of valid therapeutic targets and potential chemosensitivity biomarkers in cancer therapeutics.
1.3 ROS homeostasis and metabolism
Tumor cells harbor genetic alterations that promote a continuous and elevated production of reactive oxygen species. Whereas such oxidative stress conditions would be harmful to normal cells, they facilitate tumor growth in multiple ways by causing DNA
damage and genomic instability, and ultimately, by reprogramming cancer cell metabolism. Both exogenous and endogenous sources of ROS production have been extensively described over the past decade(Panieri et al., 2015).The most biologically relevant are represented by the nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, professional enzymes that catalyze the production of O2−· or H2O2 using NADPH as a reductant (Lambeth 2004) and the mitochondrial electron transport chain (mETC), wherein mainly complexes I and II generate O2− through univalent reduction of molecular oxygen as a consequence of electron leakage during mitochondrial respiration using nicotinamide adenine dinucleotide (reduced form) (NADH) and FADH(Kussmaul et al., 2006; Quinlan et al., 2012). Detoxification from ROS is facilitated by non-enzymatic molecules (such as vitamin A, C, or E) or through antioxidant enzymes which specifically scavenge different kinds of ROS (Liou et al., 2010).
1.3.1 Nonenzymatic antioxidants
Nonenzymatic antioxidants include low-molecular-weight compounds, such as vitamins (vitamins C and E), β-carotene, uric acid, and GSH, a tripeptide (l-γ-glutamyl- l-cysteinyl-l-glycine) that comprise a thiol (sulfhydryl) group. Water-soluble vitamin C (ascorbic acid) provides intracellular and extracellular aqueous-phase antioxidant capacity primarily by scavenging oxygen free radicals. It converts vitamin E free radicals back to vitamin E and Lipid-soluble vitamin E is concentrated in the hydrophobic interior site of cell membrane and is the principal defense against oxidant-induced membrane injury (Mezzetti et al., 1996). β-Carotene inhibits the oxidant-induced NF-κB activation and interleukin (IL)-6 and tumor necrosis factor-α production. Carotenoids also affect
apoptosis of cells. Antiproliferative effects of RA have been shown in several studies (Donato et al., 2005; Niizuma et al., 2006). In addition, GSH is a cofactor for several detoxifying enzymes, such as GSH-Px and transferase. It has a role in converting vitamin C and E back to their active forms. GSH protects cells against apoptosis by interacting with proapoptotic and antiapoptotic signaling pathways (Masella et al., 2005).
1.3.2 Enzymatic antioxidants
The different type of enzymatic antioxidants is superoxide dismutase (SOD), enzymes catalase, and glutathione reductase, members of the aldehyde oxidoreductase and various peroxidases. These enzymes respectively dismutase superoxide radical, breakdown hydrogen peroxides and hydroperoxides to harmless molecules (H2O2/ alcohol and O2), thus they are important in the prevention of lipid peroxidation and maintaining the structure and function of cell membranes.
1.3.2.1 Superoxide dismutase
Superoxide dismutase (SOD) is the first detoxification enzyme and most powerful antioxidant in the cell. It is an important endogenous antioxidant enzyme that acts as a component of first line defense system against reactive oxygen species(ROS).
All 3 forms of SOD, that is, CuZn-SOD, Mn-SOD, and EC-SOD, are widely expressed in the human lung. Mn-SOD is localized in the mitochondria matrix. EC-SOD is primarily localized in the extracellular matrix, especially in areas containing high amounts of type I collagen fibers and around pulmonary and systemic vessels. It has also been detected in the bronchial epithelium, alveolar epithelium, and alveolar macrophages (Kinnula et al.,
2003; Kinnula 2005). Overall, CuZn-SOD and Mn-SOD are generally thought to act as bulk scavengers of superoxide radicals. The relatively high EC-SOD level in the lung with its specific binding to the extracellular matrix components may represent a fundamental component of lung matrix protection (Zelko et al., 2002). SOD enzyme deficiency is common. Hence, the enzyme is indispensable to cellular health, protecting body cells from excessive oxygen radicals, free radicals and other harmful agents that promote aging or cell death. The levels of SODs decline with age, whereas free radical formation increases. It has been suggested that proper daily SOD supplementation will protect the immune system and significantly reduce one’s chances of diseases and ultimately slow down aging process.
1.3.2.2 Catalase (CAT)
Catalase (CAT) is a 240 kilodalton (kDa) tetrameric protein with four similar subunits and is encoded by ctt1 gene mapping to chromosome 11(Radi et al., 1991).Each polypeptide subunit is 60 kDa in weight and contains a single ferriprotoporphyrin (Surai 2006).CAT is a common antioxidant enzyme present almost in all living tissues that utilize oxygen. Degradation of H2O2 is accomplished via the conversion between 2 conformations of catalase-ferricatalase (iron coordinated to water) and compound I (iron complexed with an oxygen atom), consequently completing the detoxification process imitated by SOD (Marklund 1984; Chelikani et al., 2004).It is abundant in cells, where it continuously scouts for hydrogen peroxide molecules. Catalase exists as a tetramer composed of 4 identical monomers, each of which contains a heme group at the active site. Each catalase can break down millions of hydrogen peroxide molecules in one second. CAT also reacts efficiently with hydrogen donors such as methanol, ethanol,
formic acid, or phenols with peroxidase activity. Hydrogen peroxide though at low amounts tends to regulate some physiological processes such as signaling in cell proliferation, cell death, carbohydrate metabolism, mitochondrial function, and platelet activation and maintenance of normal thiol redox-balance (Dröge 2002),however, at high concentrations it has been reported to be very deleterious to cells (Ercal et al., 2001).
Hence, the ability of CAT to effectively limit H2O2 concentration in cells underlines its importance in the aforementioned physiological processes as well as being a first line antioxidant defense enzyme. The deficiency or mutation of the enzyme has been linked with various disease conditions and abnormalities (Góth et al., 2004).
1.3.2.3 Glutathione reductase (GPx)
Glutathione Peroxidase (GPx) is an important intracellular enzyme that breakdown hydrogen peroxides (H2O2) to water; and lipid peroxides to their corresponding alcohols mainly in the mitochondria and sometimes in the cytosol (Góth et al., 2004). Most times, its activity depends on a micronutrient cofactor known as selenium. For this reason, GPX is often referred to as a selenocysteine peroxidase. The enzyme plays a more crucial role of inhibiting lipid peroxidation process, and therefore protects cells from oxidative stress (Gill et al., 2010). The clinical importance of GPx has been underlined by a number of studies. Chabory et al. (Chabory et al., 2009) postulated that individuals with lower GPx activity are predisposed to impaired antioxidant protection, which leads to oxidative damage to membrane fatty acids and functional proteins, and neurotoxic damage. Forgione and colleagues had previously hypothesized that GPX deficiency directly induces an increase in vascular oxidative stress, with attendant endothelial dysfunction (Forgione et al., 2002). Decreased GPx activity has also
been reported in tissues where oxidative stress occurs in several pathological animal models. The accumulation of increased levels of peroxide resulting from inactivation of GPx may act as a second messenger and regulate expression of antiapoptotic genes and the GPx itself to protect against cell damage. Glutathione peroxidases have also been implicated in the development and prevention of many common and complex diseases, including cancer and cardiovascular disease (Rayman 2005).
1.3.2.4 Aldehyde dehydrogenase
Aldehydes are generated during metabolism of various endobiotic and xenobiotic compounds. For example, aldehydes are associated with the metabolism of alcohols, amino acids (e.g., lysine, valine, proline and arginine), anticancer drugs (e.g., cyclophosphamide) and neurotransmitters (e.g., -aminobutyric acid (GABA), serotonin, noradrenaline, adrenaline, dopamine) (Vasiliou et al., 2004; Marchitti et al., 2007). Lipid peroxidation (LPO) of cellular phospholipids induces the formation of more than 200 highly-reactive aldehyde species, including 4-hydroxy 2-nonenal (4-HNE), malondialdehyde (MDA), 4-oxononenal (4-ONE), acrolein, crotonaldehyde and methylglyoxal (Ishitani et al., 1977; Marchitti et al., 2008; Zimniak 2011). Environmental pollutants, such as smog, cigarette smoke, motor vehicle exhaust, pesticides and various food additives, either contain or contribute to the formation of aldehydes, including formaldehyde, acetaldehyde and acrolein (Ishitani et al., 1977; Marchitti et al., 2008;
Zimniak 2011). While some aldehydes play vital roles in normal physiological processes, including vision, embryonic development and neurotransmission, many aldehydes are cytotoxic and carcinogenic (Yokoyama et al., 2001; Marchitti et al., 2008). ALDHs play critical roles in metabolizing these endogenous and exogenous molecules. Their
expression is up-regulated in response to abiotic and biotic stress generated by perturbed endobiotic and/or xenobiotic metabolism. Such stress-responsive expression of ALDHs manifests in a broad range of plant and animal species, underscoring the evolutionary conservation of biological adaptions to oxidative and electrophilic stresses. Aldehydes are strongly electrophilic, highly reactive and relatively long-lived compounds. Reactive aldehydes readily form adducts with DNA, RNA and proteins, leading to impaired cellular homeostasis, enzyme inactivation, DNA damage and cell death (Brooks et al., 2005; Jacobs et al., 2010). They have been implicated in oxidative stress-associated diseases, such as atherosclerosis, cancer, diabetes, chronic alcohol exposure, acute lung injury, and in neurodegenerative diseases like Alzheimer’s and Parkinson’s disease (Jacobs et al., 2010; Yin et al., 2011). The ALDH superfamily contains NAD(P)+- dependent enzymes that oxidize a wide range of endogenous and exogenous aldehydes to their corresponding carboxylic acids (Vasiliou et al., 2004). The ability of ALDHs to act as ‘aldehyde scavengers’ is grounded in the observation that many have broad substrate specificities and can metabolize a wide range of chemically- and structurally-diverse aldehydes.
1.3.2.5 Peroxidases
Hydrogen peroxide and superoxide are also consumed by heme peroxidases, such as eosinophil peroxidase (EPO), lactoperoxidase (LPO), myeloperoxidase (MPO) (Valco M et al., 2007). Paradoxically, these enzymes are sources of more potent oxidants (hypochlorous acid, hydroxyl radical, singlet oxygen, etc.) that can cause a wide range of abnormalities including ischemic- related hypoxia and neuronal death (Everse et al., 2009). Significantly, MPO and EPO have been linked to various allergic, psychiatric, and
neurodegenerative diseases (Klebanoff 2005; Lefkowitz et al., 2008; Davis et al., 2008;
van der Veen et al., 2009).
1.3.2.6 Thioredoxin (Trx) reductase
Thioredoxin (Trx), originally identified as an electron donor for ribonucleotide reductase, functions to regulate various enzymes and trans-activating factors of genes, and is intimately involved in cell growth, differentiation, and death (Nakamura et al., 1997). Trx also functions as a protein disulfide isomerase that corrects disulfide bridges that are formed in error. Moreover, Trx directly donates electrons to peroxiredoxin and, hence, is directly linked to the peroxidase reaction (Fujii et al., 2002). After oxidation, an intramolecular disulfide bond is formed in Trx. Oxidized thioredoxin is reduced by thioredoxin reductase, a selenocystine-containing oxido-reductase, using NADPH as an electron donor. Since Trx-knockout mice are embryonically lethal (Matsui et al., 1996), Trx appears to play essential roles in the reproductive system and/or fetal development.
Among multiple roles of Trx, defect in electron donation to ribonucleotide reductase appears to be the main cause in Trx-knockout mice because DNA synthesis is essential for fetal development.
1.3.2.7 Aldo-keto reductase
Carbonyl compounds are produced by the oxidation of organic compounds, such as unsaturated fatty acids, and are highly reactive. They modify reactive sulfhydryl groups that are commonly present in redox-sensitive molecules, resulting in an impairment of the systems. Mammalians have several enzymatic systems that function to detoxify
carbonyl compounds. The aldo-keto reductase family includes enzymes that reduce carbonyl groups to alcohol using NADPH as an electron donor. Among the members of this family, aldose reductase, the AKR1B gene product, has been the most extensively studied because it is intimately involved in diabetic complications (Yabe-Nishimura 1998). An inhibitor of aldose reductase is one of proposed cures for diabetic complications. Aldehyde reductase, the AKR1A gene product, exhibits the highest similarity to aldose reductase among the family members (Jez et al., 1997) and appears to play a coordinate function (Iuchi et al., 2004). Since steroid hormones and their derivatives contain carbonyl groups and can serve as substrates for aldo-keto reductase (Wermuth et al., 1983), enzymes that are highly expressed in tissues with steroid hormone production may have a role in their elimination. The detoxification of carbonyls is activated by the binding of GSH, which indicates crosstalk between the GSH redox system and aldo-keto reductase system (Dixit et al., 2000).
1.4 Aldo-Keto Reductase (AKR) family
The Aldo-Keto Reductase (AKR) superfamily comprises of several enzymes that catalyze redox transformations involved in biosynthesis, intermediary metabolism and detoxification. Substrates of the family include glucose, steroids, glycosylation end products, lipid peroxidation products, and environmental pollutants. These proteins adopt a (β/α)8 barrel structural motif interrupted by a number of extraneous loops and helixes that vary between proteins and bring structural identity to individual families. The human AKR family differs from the rodent families. Due to their broad substrate specificity, AKRs play an important role in the Phase II detoxification of a large number of pharmaceuticals, drugs, and xenobiotics. Mammalian AKRs fall within 3 well-defined
families (AKR1, 6 and 7). These proteins are widely distributed in tissues and most cells express several AKRs. In human, two human enzymes from the AKR1B subfamily (AKR1B1 and AKR1B10) are of special interest.
1.4.1 AKR1B1
AKR1B1was first identified as a protein with glucose reducing activity in 1956 by Hers et al. (Hers 1956). The AKR1B1 gene lies on the chromosome location of 7q33 and consists of 10 exons. There are a few putative pseudogenes for this gene, and one of them has been confirmed and mapped to chromosome 3. AKR1B1 consists of 316 amino acid residues and weighs 35kDa. with a widely expression in human organs including the kidney, lens, retina, nerve, heart, placenta, brain, skeletal muscle, testis, blood vessels, lung, and liver (O'connor et al., 1999).
1.4.1.1 AKR1B1 function
During hyperglycemia AKR1B1 mediates the first step of the “polyol pathway”
by reducing glucose to sorbitol using NADPH as a co-factor and later sorbitol dehydrogenase (SDH) reduces sorbitol to fructose using NAD as a co-factor. Under normoglycemic conditions only 3% of total glucose is converted into sorbitol through the polyol pathway. Under hyperglycemic conditions, however, the flux of glucose through this metabolic pathway may increase up to 10-fold (30%), which might finally lead to accumulation of excess sorbitol (Yabe-Nishimura 1998; Tang et al., 2012; Hwang et al., 2017). The resulting osmotic stress and imbalances of the pyridine nucleotide redox status decrease the cell’s antioxidative capabilities. In diabetes mellitus, this promotes the
formation of advanced glycation end products (AGEs) (Srivastava et al., 2005; Brings et al., 2017), which themselves lead to diabetic complications, such as microangiopathies, nephropathies, retinopathies, peripheral neuropathies and cataract (Del Corso et al., 2008;
El Gamal et al., 2017). Consequently, aldose reductase inhibitors (ARIs) are at the focus of exploratory pharmaceutical research, as they yield the potential to prevent or control the onset of these diabetic complications. In vitro, AKR1B1 displays poor affinity to glucose and hydrophobicity of the putative substrate-binding domain essentially precludes efficient carbohydrate reduction (Ramana 2011). Therefore, lipid peroxidation- derived hydrophobic aldehydes, such as 4-hydroxynonenal (HNE) and other related nephro- and hepatotoxic compounds are more likely to be physiological substrates of AKR1B1. Finally, high catalytic efficiencies of AKR1B1 towards these compounds suggest an important metabolic role in the detoxification of lipid-derived aldehydes (Barski et al., 2008; Ramana 2011). Physiologically and despite the relatively high KM for glucose, however, selective AKR1B1 inhibition remains an effective treatment option for diabetic complications derived from glucose-related adducts (Ramana 2011).
1.4.1.2 Impact of AKR1B1 in cancer
AKR1B1 expression is increased by high blood glucose via NF-κB (Yang et al., 2008), providing a potential mechanism by which diabetes and elevated risk of developing certain cancers may be linked. AKR1B1 and the polyol pathway also contribute to hyperglycemic pseudohypoxia, which one could imagine linking the Warburg effect to tumor angiogenesis through HIF1a and perhaps bolstering neovascularization at oxygen tensions that would not normally promote it. Consistent with this, VEGF has been linked to diabetic retinopathy and nephropathy (Aiello et al.,
1994; Cha et al., 2000; Ozaki et al., 2000), perhaps downstream of AKR1B1-driven pseudohypoxic effects. Intriguingly, patients with Von Hippel–Lindau disease often develop retinal angiomas and kidney tumors, suggesting that VHL-associated malignancies and diabetic complications may differ primarily by the degree of HIF1a and/or VEGF-dysregulation present. It is possible that diabetics are effectively primed to promote tumorigenesis by virtue of an already abnormally high level of hypoxia/HIF1a signaling.
Therefore, AKR1B1 expression has been reported to be elevated in human cancers (Ramana et al., 2010; Tammali et al., 2011) and drug resistance in cancer-derived cell lines (Dan et al., 2003; Matsunaga et al., 2011; Zhong et al., 2011).
1.4.2 AKR1B10
AKR1B10 is a 36-kDa cytosolic reductase that is similar to AKR1B1 in both amino acid sequence identity (71%) and tertiary structure with the (α/β)8barrel topology (Gallego et al., 2007). Like AKR1B1, AKR1B10 reduces a variety of aromatic and aliphatic aldehydes, dicarbonyl compounds, and some drug ketones using NADPH as the coenzyme (Cao et al., 1998; Spite et al., 2007; Zhong et al., 2009; Endo et al., 2011), and the reaction catalyzed by the enzyme follows an ordered bi bi mechanism with the binding of coenzyme to the free enzyme (Endo et al., 2009). In contrast to AKR1B1, AKR1B10 differs from AKR1B1 not only in its inability to reduce glucose and xylose (Cao et al., 1998; Crosas et al., 2003) and prostaglandin H2 (Kabututu et al., 2009), but also in its high catalytic efficiency for cytotoxic aldehydes (acrolein and 4-hydroxy-2-nonenal;
(Shen et al., 2011)), isoprenyl aldehydes including farnesal and geranylgeranial, which