國立臺灣大學醫學院分子醫學研究所遺傳諮詢組 碩士論文
Genetic Counseling Program, Graduate Institute of Molecular Medicine College of Medicine
National Taiwan University Master Thesis
胚胎細胞中粒線體 DNA 含量與胚胎發育能力之關係 The investigation of mitochondrial DNA copy number as a
biomarker of embryo developmental competency
吳劭穎 Shao-Ying Wu
指導教授:李妮鍾 醫師
Advisor: Ni-Chung Lee, MD, PhD
中華民國 108 年 7 月 July, 2019
目 錄
口試委員會審定書………...
I誌謝………...
II中文摘要………...………
III英文摘要…...………
V頁數 第一章 緒論
第一節 研究背景... 1
第二節 研究目的………. 3
第二章 研究設計與方法 第一節 研究架構……… 4
第二節 研究對象……… 4
第三節 試管嬰兒療程、胚胎胚培養與切片流程……… 5
第四節 全基因體放大流程……… 5
第五節 次世代定序 (NGS) 流程………...…… 6
第六節 胚胎染色體套數計算……… 7
第七節 粒線體 DNA 相對含量計算……… 8
第八節 臨床懷孕指標……… 9
第九節 研究指標分析方式……… 9
第十節 統計學方法……… 10
第三章 研究結果
第一節 胚胎染色體套數與粒線體 DNA 含量之關係…… 11 第二節 胚胎發育速度與粒線體 DNA 含量之關係……… 11 第三節 母體年齡與粒線體 DNA 含量之關係…………... 12 第四節 第五天和第六天囊胚之比較………...…… 12 第五節 粒線體 DNA 含量用於預測胚胎之懷孕率……… 13
第四章 研究討論
第一節 胚胎染色體套數與粒線體 DNA 含量之關係…… 15 第二節 於染色體套數異常或品質不佳胚胎中粒線體
DNA 含量增加之原因與意義…….……… 17 第三節 粒線體異質性與試管嬰兒成功率之關係………... 18 第四節 “越少越好?” 粒線體 DNA 含量多寡與胚胎
發育速度之關係………...…… 19 第五節 粒線體 DNA 量與母體年齡無顯著關聯之原因… 21
第五章 研究限制………..…… 22
第六章 研究結論………..…… 23
參考文獻……….….… 24
圖表目錄
圖 1 胚胎染色體套數與粒線體 DNA 含量之關係….…. 32
圖 2 胚胎發育速度與粒線體 DNA 含量之關係.……… 32
圖 3 母體年齡與粒線體 DNA 含量之關係…….……… 33
圖 4 第五天和第六天囊胚之染色體正常比例以及植入 後臨床懷孕率之比較………...……… 33 圖 5 植入後有成功達成臨床懷孕的囊胚與植入後未能
達成臨床懷孕的囊胚中粒腺體含量之比較……….. 34 圖 6 粒線體 DNA 的含量用於預測胚胎之懷孕率... 35
表 1 植入後胚胎的粒腺體 DNA 含量之個別百分位比….36
國立臺灣大學碩博士學位論文
口試委員會審定書
胚胎細胞中粒線體 DNA 含量與 胚胎發育能力之關係
The investigation of mitochondrial DNA copy number as a biomarker of embryo developmental
competency
本論文係 吳劭穎 君(P06448003)在國立臺灣大學醫學 系、分子醫學所完成之碩士學位論文,於民國 108 年 7 月 29 日承下列考試委員審查通過及口試及格,特此證明
口試委員:
(簽名)
(指導教授)
系主任、所長 (簽名)
誌謝
偉大的科學家牛頓曾經說過:“如果我能看得更遠,那是因為站在巨人的肩膀 上。”不過我認為更重要是我們要感謝所有在這段路途上幫助我們,讓我們能順利 爬上巨人肩膀的人。因為沒有他們,我們必然會走的更辛苦、更孤單、也許更不會 成功。
在此首先我想感謝我的指導教授李妮鍾醫師。儘管老師您平常都是酷酷的,不 過我真的覺得您講話十分幽默。而且您每次上課的都超級精彩,內容深入淺出,讓 我們在歡樂的氣氛中充分學習。在門診中除了跟您學習臨床知識之外,也深深感受 您對每一位患者及家屬的用心與關心,除了敬佩之餘更是我日後與人相處互動的 榜樣。在論文方面也由衷感謝老師您在研究思考上給予我無限發揮空間及包容,同 時也一直引導我去突破我現有的知識範疇,結合我過去所學,讓我更能學以致用。
另外我想謝謝所有在課堂、實驗室和門診指導過我們的師長們。除了知識上 的傾囊相授之外,更讓我從重溫了當學子的美好,純粹的享受知識的學習。
我也想謝謝我的家人,在唸書和工作夾擊之最容易被犧牲的就是和你們相處 的時間。我先謝謝我的爸媽,從小他們都一直教我不要畏懼挑戰,更要活到老學到 老。第二我想謝謝我兩個可愛的孩子,這兩年陪伴著你們的時間真的少了,真的很 感謝你們的體諒。另外我想謝謝我的太太,每當我感到心力交瘁快撐不下去時,你 都會用你獨特的方式激勵我,讓我能發揮潛力堅持下去。
我還想特別謝謝四個人。第一位是愛群婦產科診所的翁紹評執行長,很謝謝您 願意讓我在忙碌的臨床業務之餘重返校園進修。第二位是我們美麗大方的助教杜 惠桑小姐,很謝謝您兩年來每次都不厭其煩的提醒我這位迷糊的“老學生”各項大小 事情。第三位是胡醫師實驗室的劉雅芬女士 (雅芬姐),很感謝您協助我這位 IRB 菜鳥順利完成所有繁複的申請步驟。第四位是愛群婦產科診所基因醫學組的吳柏 寬 (阿寬),很謝謝您在許多實驗原理步驟的解惑,每次和您討論完問題都讓我有 恍然大悟、茅塞頓開的感覺。
最後我想謝謝在班上所有的好同學好夥伴們。跟你們在一起讓我永遠都不會 覺得自己很老。雖然不常見面每次總有聊不完的話題。上課考試壓力很大,一起互 相鼓勵打氣度過重重難關。每逢課程遭遇困難時,永遠都會有人馬上挺身而出,讓 我能無後顧之憂的快樂學習。這兩年的點點滴滴我永遠會銘記在心 ~謝謝大家!
中文摘要
研究背景: 全世界首例試管嬰兒問世至今已超過 40 年。雖然目前人工生殖的技 術已十分發達,傳統試管嬰兒的成功率仍僅有三成左右。造成胚胎植入後著床失敗 的主因為胚胎染色體套數異常。植入前胚胎染色體套數檢查 (Preimplantation Genetic Screening; PGS) 是一種在植入前對胚胎進行切片和染色體數目分析的 技術,目的為挑選出染色體數目正常的胚胎來進行植入。即使如此,植入經由 PGS 篩選過的的胚胎仍有近三成左右仍無法順利著床。由此可見除了染色體的數目是 否正確之外,應還有其他可用於輔助判斷胚胎著床能力的因素存在。粒線體是一種
存在於人體大部分細胞內的胞器。它是細胞內能量分子「三磷酸腺苷」(Adenosine
Triphosphate; ATP)合成製造的主要場所,所以粒腺體可被視為「細胞能量中心」。
近年來已陸續有研究發現胚胎細胞內的粒線體 DNA 含量等對胚胎品質是有影響 的,不過亦有其他研究結果抱持的相反結果。所以胚胎內的粒腺體 DNA 含量是否 可用於預測胚胎品質、著床率、懷孕率等目前仍具爭議性。
研究目的: 探討胚胎滋養層細胞內的粒線體 DNA 含量與 (1)胚胎染色體套數 [euploid vs. aneuploid]、(2)胚胎發育速度 [Day 5 囊胚 vs.Day 6 囊胚] 及 (3) 母體年齡之間的關係。另外亦有針對染色體套數正常的但是具有不同發育速度的 胚胎之間的懷孕率及粒線體含量進行分析。希望此研究結果能讓粒線體 DNA 含量 在臨床上成為篩選胚胎著床能力的另一重要指標。
研究方法: 我們進行回溯性病例研究,收集自 2016 年 11 月至 2018 年 04 月在愛 群婦產科診所進行人工生殖 (試管嬰兒與植入前胚胎染色體數目篩檢) 的胚胎及 母 親 年 齡 之 資 訊 。 我 們 從 NGS 數 據 中 分 析 滋 養 層 細 胞 內 的 粒 腺 體 DNA (mitochondrial DNA copy number; mtDNA) 與細胞核 DNA (nuclear DNA copy number; nDNA) 的相對含量 (mtDNA score)。同時也探討胚胎染色體套數、胚胎 發育速度、胚胎植入後結果以及母親年齡等資料與 mtDNA score 之相關性。
結果: 總共有來自 460 名女性不孕症患者中共 1301 顆有接受次世代定序 PGS 的胚胎被分析。染色體套數異常的胚胎和染色體套數正常的胚胎相比在滋養層細
胞中含有量較多的 mtDNA,且此差異在統計學上具有顯著性。在第五天發育為成熟 囊胚的胚胎 [Day 5 囊胚] 和於第六天才發育為成熟囊胚的胚胎 [Day 6 囊胚]
相比在滋養層細胞中含有量較多的 mtDNA,且此差異在統計學上具有顯著性。Day 5 囊胚不論是在胚胎染色體正常比例,臨床懷孕率都顯著優於 Day 6 囊胚。母體 年齡對於胚胎滋養層細胞中的 mtDNA 含量無直接影響。根據本研究結果所界定用 於預測胚胎著床力的相對 mtDNA 含量為 0.002,即所有 mtDNA 高於此數值的胚胎在 植入後均未著床成功。
結論: 雖然在染色體套數異常的胚胎中 mtDNA 的量會顯著上升,不過這並非代表 在染色體套數正常的胚胎中 mtDNA 含量越高胚胎品質是越差的。更大型且採用雙 盲隨機分配的相關研究仍須持續進行來使我們能更了解粒線體量的多寡在胚胎中 的意義以及臨床運用於挑選植入成功率的最高的胚胎之可行性。
關鍵字: 粒線體、試管嬰兒、胚胎品質、次世代定序、染色體
Abstract
Introduction: The primary goals of modern-day assisted reproductive technology (ART) is to increase pregnancy and live birth rates, while at the other hand decrease complications related to preterm births from multiple pregnancies. Scientists have tested many novel technologies that could reliably assess embryo viability and quality, which ultimately would enable transferring only a single competent embryo that is suffice in creating a healthy live birth. Recently, mitochondria DNA quantity within the embryonic trophectoderm cells have been suggested to be a predictor of embryo viability in several studies, while others have found no such correlation.
Objective(s): To investigate the relationship between the quantity of embryonic trophectoderm cells’ mitochondria DNA and (1) embryo ploidy (euploid vs. aneuploid) 、
(2) embryo developmental rate (Day 5 vs. Day 6 blastocyst formation) and (3) maternal age. Secondly, to explore the difference in mtDNA quantity and transfer outcomes between euploid embryos of different development rates. cells have implications in respect to the reproductive potential of the embryos. Lastly, to assess the accuracy of mtDNA quantity in predicting implantation potential of euploid embryos.
Study Design: This is an observational retrospective analysis consisted of medical records review of infertile patients that underwent in-vitro fertilization and preimplantation genetic screening at IHMED Fertility Center from 2016/11 to 2018/04.
Primary intervention was the calculation of “mtDNA score”, which was the relative quantification of embryo mitochondria DNA copy number with regard to nuclear DNA copy number extracted from the NGS data for PGS. The relationship between mtDNA score to embryo ploidy, embryo developmental rate, maternal age, and embryo transfer outcomes were also analyzed.
Result(s): A total of 1,301 embryos from 460 infertile women undergoing in-vitro fertilization (IVF) with intracyoplasmic sperm injection (ICSI) and next generation sequence (NGS) based pre-implantation genetic screening (PGS) were included for the analysis. Aneuploid embryos had significantly greater quantity of mtDNA compared to eupoid embryos. When considering the embryo development rate, day 5 blastocysts had a significantly higher mtDNA quantity than day 6 blastocysts, independent of embryo ploidy. Thirdly, when factoring the effects of maternal age, no differences in the mtDNA quantity was noted when analyzing blastocysts pooled from subjects aged < 40 and > 40 years-old, irrespective of embryo ploidy and development rate. The comparison of embryo euploidy rates, clinical pregnancy rates, and mtDNA score of euploid embryos that resulted in clinical pregnancy between day 5 blastocysts and day 6 blastocysts also showed significant difference favoring day 5 blastocysts. The optimal cut-off values of mtDNA score that predicts the viability of euploid blastocysts was 0.002.
Conclusion(s): mtDNA quantity is a useful indicator that accurately predicts the implantation potential of euploid blastocysts. Increased mtDNA content was frequently observed in aneuploid blastocysts or euploid blastocysts of reduced implantation potential, indicating possible embryonic stress or intrinsic mitochondrial dysfunction. Nevertheless, a minimal mtDNA quantity must also be maintained in order for proper embryonic development and establishment of viable pregnancy. The delicate balance between mtDNA quantity and embryo viability warrants further large scale, randomized controlled studies.
Keywords: mitochondria, in-vitro fertilization, embryo viability, next-generation sequencing, chromosomes
第一章 緒論
第一節 研究背景
Embryo aneuploidy has long been regarded as one of most important contributor to implantation failure and miscarriage [1,2]. Therefore, preimplantation genetic screening (PGS) with comprehensive chromosome screening (CCS) was implemented as a mean to eliminate the chance of transferring embryos with abnormal chromosome copy number, at the same time reducing the complications related to multiple pregnancies. Nevertheless, there are still nearly one-third of embryos that were deemed to be euploid by PGS but failed to implant or achieve live birth [3]. Uterine structural anomalies, presence of fibroids or endometrial polyps, hydrosalpinx, underlying autoimmune disorders have all been implicated as possible culprits that led to implantation failure of euploid embryo [4].
However, even when all of above factors are controlled, the overall implantation rate of euploid embryos still remained far from perfect.
Mitochondria, also best known as “the powerhouse of cell”, is one of the most abundant cellular components in the mammalian oocytes and early embryos. In addition to being the main organelle responsible for ATP production through oxidative phosphorylation (OXPHOS), mitochondria also serve many other regulatory roles in cells, such as calcium homeostasis, apoptosis, and generation of reactive oxygen species (ROS) [5-7]. Despite the importance of proper mitochondrial functions to human reproductions have long been recognized, it was only until the recent advancements in PGS technology which prompted scientists to further examine their relationship clinically under the settings of IVF [8-12].
There have been several recent large scale studies, with the largest analysis consisted of over 1,500 embryos, implicating the quantity of mitochondria DNA (mtDNA) in
embryos was intrinsic to their implantation potential and pregnancy outcome [8-11].
These studies all consisted of subjects that underwent in-vitro fertilization (IVF) with PGS, with simultaneous determination of mtDNA quantity in the trophectoderm biopsy.
Not only did these studies consistently revealed a significant elevation of mtDNA in aneuploid embryo compared to their euploid counterparts, an unique phenomenon of
“mtDNA threshold” was also observed, above which none of the euploid embryo had succeeded in implantation. Thus the concept of utilizing mtDNA quantity as an adjuvant marker for the viability of euploid embryos have been suggested [8-11].
The findings of the above studies were nevertheless challenged by others. In an analysis by Victor et al., no significant difference in blastocysts’ trophectoderm mtDNA levels was found after applying a “correction factor” for each embryo’s chromosomal composition, irrespective of the corresponding embryo viability, ploidy or maternal age [12]. In another interesting study by Treff et al., under the setting of double euploid embryo transfers that resulted in only singleton pregnancies, no significant difference in the trophectoderm mtDNA levels was found between the sibling embryos [13]. Adding to these conflicting results, a recent study revealed a significantly higher mtDNA quantity in blastocysts that was biopsied on day 5 compared to the ones biopsied on day 6 [14].
This was the first ever study that made such a distinction, but unfortunately the transfer outcome for these groups were not analyzed independently, thus the potential effect of blastocyst development rate was not adequately considered.
第二節 研究目的
The purposes of the present study are to investigate:
(1) The relationship between mtDNA quantity to embryo ploidy
(2) The relationship between mtDNA quantity to embryo development rate (3) The relationship between mtDNA quantity to maternal age
(4) The transfer outcomes of Day 5 euploid blastocysts vs. Day 6 euploid blastocysts with respective to mtDNA quantity
(5) The optimal cut-off mtDNA quantity for predicting clinical pregnancy of euploid embryos
第二章 研究設計與方法
第一節 研究架構
Study Design
The is a retrospective cohort analysis consisted of de-identified NGS data from infertility patients consenting to PGS during IVF procedures. The study protocol was approved by the institutional review board of National Taiwan University Hospital (IRB
#201905053RIND).
第二節 研究對象
Study Participants
A total of 460 subjects, between the age of thirty to forty-six were enrolled in the present study. Among them, a total of 1,301 blastocysts were obtained and analyzed, with 496 designated as euploid and 805 as aneuploid after PGS. The blastocysts were further divided into four groups according to developmental rate: euploid day 5 blastocyst (ED5; n= 333), aneuploid day 5 blastocysts (AD5; n= 496), euploid day 6 blastocyst (ED6; n=
163), aneuploid day 6 blastocysts (AD6; n= 309). Main exclusion criteria were cases which PGS was not performed either due to poor embryo quality, failed amplification, or by patients’ informed choice. Other exclusion criteria were cases with uterine structural anomaly, untreated hydrosalpinx, and autoimmune disorders such as systematic lupus erythematous and antiphospholipid syndrome. Lastly, all embryos designated as mosaic were also excluded.
第三節 試管嬰兒療程
、胚胎胚培養與切片流程
Ovarian Stimulation and Embryo Processing
All IVF and embryo processing took place at IHMED Fertility Center using standardized techniques. Intracytoplasmic sperm injection (ICSI) was used for all oocytes, and the derived zygotes were culture with a continuous single culture medium (Global Total;
LifeGlobal Inc.). Five to eight-cell trophectoderm biopsies were performed with laser- assisted micromanipulator (LYKOS; Hamilton-Thorne Inc.) on either day 5 or day 6 post fertilization, depending on the developmental rate and morphological grade of individual blastocyst. Specifically, Gardner’s blastocyst grading system was used to score each blastocyst, and only the ones with grade of 3CC or above were biopsied for PGS. The embryos were then cryopreserved with an open vitrification system (Cryotop; Kitazato Inc.). Each biopsied sample was then suspended in 1× phosphate-buffered saline (PBS) (HyClone, GE Healthcare Life Sciences) and placed in a tube containing 2.5μl of 0.5%
polyvinylpyrrolidone.
第四節 全基因體放大流程
Whole Genome Amplification Workflow
Whole genome amplification (WGA) of each biopsied sample was performed using the SurePlex WGA Kit (Illumina Inc.) in accordance to manufacturer’s instructions with minor in-house modifications. Sureplex utilizes a fragment-amplification based polymerase chain reaction (PCR) approach with self-insert degenerative primers that facilitates annealing of target genome sites [15]. The biopsied samples are processed under a hood equipped with laminar flow in a laboratory specifically used for DNA amplification. The first step consisted of cell lysis / extractions (45 minutes), followed by the pre-amplification steps (70 minutes) and the final amplification steps (60 minutes).
Upon completion, agarose gel electrophoresis of the PCR products is performed to determine the extent of amplification success. The specifications and details of all the above steps is accessible from manufacture’s website (SurePlex Summary Protocol Reference Guide – Illumina) [16]. Sureplex was the chosen WGA method since it has been validated for use in single cell amplification, with additional advantages of protocol simplicity, high amplification reproducibility, and low allele drop-out rate (<10%) [15,17].
第五節 次世代定序 (NGS) 流程
Next Generation Sequencing (NGS) Workflow
For the present study, we utilized a “low-pass” NGS protocol that was first proposed by Wells et al. [18], with in-house modifications made in accordance to our laboratory standards. With the low-pass protocol, the WGA products are sequenced with low coverage depth of 0.01x. Therefore, in theory less than 1% of the embryonic genome is sequenced per run. As one may wonder whether such low genomic coverage would be sufficient for aneuploidy screening, previous works have validated this method by testing samples from well-characterized cell lines and embryo biopsies from another validated platform (array comparative genomic hybridization; aCGH) that demonstrated 100%
sensitivity and specificity for ploidy calling, as well as a high concordance rate (>99%) for each evaluated chromosome [18,19].
Libraries for the WGA samples were prepared in accordance to the standard VeriSeq PGS protocol (Illumina Inc.) [20]. The input DNA was first “tagmented” (tagged + fragmented) with adaptor sequence by Nextera XT Transposome, followed by addition of index adaptor oligos with Nextera XT 96-Index Kit (Illumina, Inc.) that enabled multiplexing up to 24 samples. Contaminant DNA, RNA, and oligonucleotides were removed with
commercial DNA purification kit (Zymo DNA Clean & Concentrator; Zymo Research Inc.) and quantified with target-selective fluorescence dsDNA assay kit (Qubitw Assays;
Thermo Fisher Scientific Inc.). Single-end, 36 base pairs reads dual-index sequencing was performed on a MiSeq sequencer with MiSeq Reagent Kit v2 kit (Illumina, Inc.).
Sequence quality score (Q-score) was used to assess the probability of an error in base calling during sequencing. A score of Q30 was equivalent to 1 error in 1,000 base calls (99.9% accuracy). In the present study, greater than 90% base calls had scores higher than Q30 at 1 × 36 bp. Samtools (Genome Research Limited) was used to remove unmapped, duplicated, and low scoring reads. The mapped reads were also normalized for GC- content. In the present study, a 24-sample multiplex generated an average of 500,000 filtered reads mapped to the reference genome per sample analyzed.
第六節 胚胎染色體套數計算
Determination of Embryo Ploidy
The generated reads were mapped to the reference human genome (hg19) using Burrows- Wheeler Alignment (BWA) tool [21] and analyzed with BlueFuse Multi Software (Illumina Inc.) [22]. Individual chromosomes from the reference genome were divided into non-overlapping bins, each with an approximate size of one megabyte. Reads from each sample were then aligned to the corresponding bins.
Using the NGS data from an euploid embryo cohort, a set of reference value for each individual chromosome was obtained by averaging the percentage of mapped reads allocated to regions on the same chromosome. The embryo ploidy was then determined by dividing the percentage of reads for each chromosome regions from the test sample to the corresponding reference value of the same chromosome regions. For our laboratory, the range of reference ratio for euploid (disomy) state was from 0.7 to 1.3 for each
individual chromosome. Therefore, an embryo would be assigned as trisomy if the reads for any given chromosome exceeded the reference value of the same chromosome by a ratio of 1.3x. Vice versa, an embryo would be assigned as monosomy if the reads for any given chromosome had a ratio less than 0.7x in respect to the reference value of the same chromosome.
第七節 粒線體 DNA 相對含量計算
Determination of mtDNA quantity (mtDNA score)
For the present study, the “relative” quantity of mtDNA (mtDNA score) in individual trophectoderm biopsy sample was inferred from the NGS data using the sequencer on- board software. The rationale for applying this “indirect” method of calculation rather than directly quantifying the actual amount of mtDNA underscored the importance of basic understanding of mitochondria biology, as well as the methodology consideration for embryo biopsy techniques and analytical platform.
For most human cell types, a single copy of the nuclear genome, comprised of approximately three billion DNA base pairs packaged into 23 pairs of chromosomes, resides in the cell’s nucleus. In contrast, variable number of mitochondria are present in the cytoplasm, each with multiple copies of the double-stranded circular mitochondrial genome containing 16,569 DNA base pairs [23]. Since the samples collected during the embryo biopsy were from a cluster of trophectoderm cells that randomly herniated through the pre-breached zona pellucida, the actual number of cells collected was impossible to control and could vary greatly among different samples. Consequently, by directly quantifying the actual mtDNA content in a given sample, one would not be able to distinguish whether the was in fact a “true” elevation of mtDNA per individual cell, or simply as the result from increased number of cells obtained during the embryo biopsy
procedure. Therefore, the total detected mtDNA reads must be normalized against the total detected the nuclear DNA reads, under the assumption that they both were sequenced with equal opportunities. The calculated “mtDNA score” would in fact be the ratio between the number of mtDNA reads to the number nuclear DNA reads, which translated to the average mtDNA content among the cells of a biopsy sample. Given their marked size difference, the disproportion between the reported nuclear and mitochondria genome ratio was expected, even when mtDNA is usually presented in multiple copies. For the present study, the coverage depths reported for the mitochondrial genome is between 20x- 50x per analyzed sample.
第八節 臨床懷孕指標
Determination of Pregnancy
Serum β-hCG was measured nine to ten days after the frozen embryo transfer, and a value above 5 IU/mL was considered to be a positive pregnancy. Clinical pregnancy was defined as a pregnancy confirmed by ultrasound visualization of the gestational sac between the 5th to 6th weeks of gestation. All cases of biochemical pregnancy, defined as transient elevation serum beta-hCG level without visualization of gestation sac, was allocated to non-pregnant group.
第九節 研究指標分析方式
Outcome Measures
The primary outcomes were the relationship of mtDNA scores to maternal age, embryo ploidy, and blastocyst development rate. The secondary outcomes are the comparison of mtDNA scores for blastocysts of similar ploidy (ED5 vs. ED6 ; AD5 vs. AD6) and similar development rate (ED5 vs. AD5; ED6 vs. AD6). The tertiary outcome measure was the
calculation of the optimal cut-off value for the mtDNA score that distinguished implanted euploid blastocysts from non-implanted blastocysts. The differences in embryo euploidy rate, clinical pregnancy rate were also analyzed, as well as the difference in the mtDNA content between day 5 euploid blastocysts and day 6 euploid blastocysts that resulted in clinical pregnancy.
第十節 統計學方法
Statistical Analysis
The mean with standard deviation for the continuous variables were calculated and the between-group differences were analyzed by Student’s t-test. Categorical variables were presented as raw frequencies with corresponding percentages, and the between-group differences were analyzed by Chi-square test with Yates correction if indicated. The results were considered statistically significant at p < 0.05. The strength of correlation between two independent variables was calculated with Pearson product-moment correlation coefficient. All the continuous and categorical variable analysis were performed with GraphPad QuickCalcs, and all the figures and tables were created with Microsoft Excel 2016.
第三章 研究結果
第一節 胚胎染色體套數與粒線體 DNA 含量之關係
The relationship between mtDNA score to embryo ploidy
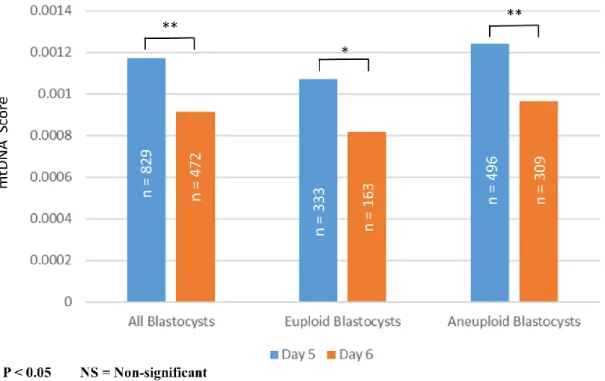
In term of embryo ploidy, a significance difference in mean mtDNA score was observed between all euploid and aneuploidy blastocysts, irrespective of development rate (0.00098966347 ± 0.00071762096 vs. 0.00113364289 ± 0.00129129378; p = 0.023). When further analyzing only blastocysts of identical development rate, day 5 euploid blastocysts showed a significantly lower mean mtDNA score compared to day 5 aneuploid blastocysts (0.001072905 ± 0.000714348 vs. 0.001239826 ± 0.001475416; p = 0.03). On the contrary, though day 6 euploid blastocysts also showed a trend of lower mean mtDNA score compared to day 6 aneuploid blastocysts, the difference was not statistically significant (0.000819607 ± 0.000696004 vs. 0.000963201 ± 0.00089825; p = 0.05). The results from the above analysis are outlined in Figure 1.
第二節 胚胎發育速度與粒線體 DNA 含量之關係
The relationship between mtDNA score to embryo development rate
In term of embryo development rate, a significance difference in mean mtDNA score was observed between all day 5 and day 6 blastocysts, irrespective of ploidy (0.001172775 ± 0.00122991626 vs. 0.000913612 ± 0.00083600924; p = 0.001). When sub-analyzing only embryos of identical ploidy, day 6 euploid blastocysts showed a significantly lower mean mtDNA score compared to day 5 euploid blastocysts (0.000819607 ± 0.000714348 vs.
0.001072905 ± 0.000696004; p = 0.022). Similarly, day 6 aneuploid blastocysts also showed a significantly lower mean mtDNA score compared to day 5 aneuploid
blastocysts (0.001239826 ± 0.001475416 vs. 0.000963201 ± 0.00089825; p = 0.001). The results from the above analysis are outlined in Figure 2.
第三節 母體年齡與粒線體 DNA 含量之關係
The relationship between mtDNA score to maternal age
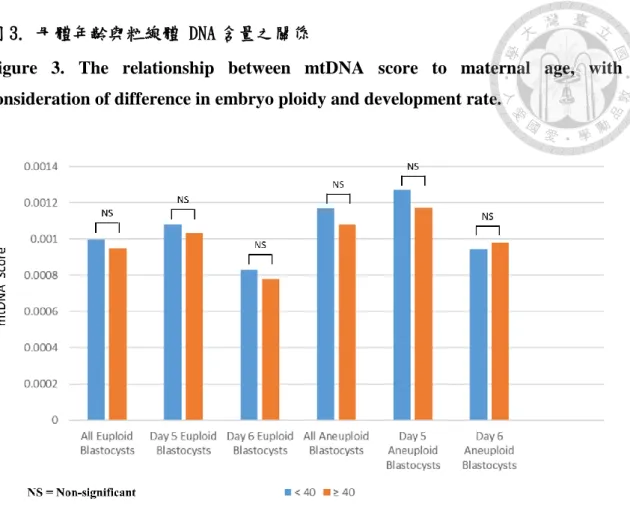
A significant correlation failed to exist between mtDNA score and maternal age (Pearson product-moment correlation coefficient, r = -0.002). In order to further investigate the effects of maternal age, the difference of mtDNA score between patients <40 year-old and ≥ 40 year-old at the time of oocyte retrieval was also analyzed. No significant difference in mean mtDNA score was found between the two maternal age groups.
Furthermore, when considering the blastocysts of similar ploidy and development rate, a significant correlation also failed to exist between maternal age and mtDNA score. The results from the above analysis are summarized in Figure 3.
第四節 第五天和第六天囊胚之比較
The outcomes of Day 5 euploid blastocysts vs. Day 6 euploid blastocysts
The embryo that developed into mature blastocyst on day 5 had a significantly higher proportion being euploid compared to day 6 blastocysts (40.1% vs. 34.5%, p = 0.044).
When comparing the clinical pregnancy rate of day 5 euploid blastocysts versus day 6 euploid blastocysts, a significant difference was noted favoring the day 5 embryos (69.7%
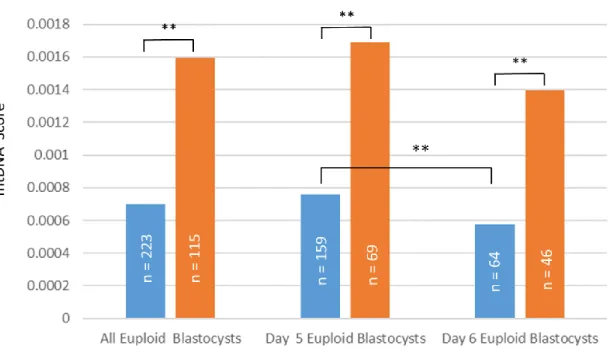
vs. 58.1%; p = 0.038). The euploid blastocysts that resulted in clinical pregnancy have significant lower mean mtDNA score than the euploid blastocysts that did not result in clinical pregnancy (0.00069757754 ± 0.00034627866 vs. 0.00159407610 ±
0.00092511726; p < 0.0001). When inspecting blastocysts of different development rate, the day 5 euploid blastocysts that resulted in clinical pregnancy have significant lower mean mtDNA score than the day 5 euploid blastocysts that did not result in clinical pregnancy (0.00075879001 ± 0.00037345518 vs. 0.00168701665 ± 0.00079820584; p <
0.0001). The same phenomenon was also observed when comparing the mean mtDNA score between Day 6 euploid blastocysts that resulted in clinical pregnancy and the one that did not (0.00057267100 ± 0.00024040394 vs. 0.00139528658 ± 0.00113822518; p <
0.0001). The day 5 euploid blastocysts that resulted in clinical pregnancy after transfer had significantly higher mean mtDNA level than day 6 euploid blastocysts that resulted in clinical pregnancy (0.00075879001 ± 0.00037345518 vs. 0.00057267100 ± 0.00024040394; p < 0.0001). Theresults from the above analysis are outlined in Figure 4 and Figure 5, respectively.
第五節 粒線體 DNA 含量用於預測胚胎之懷孕率
The optimal cut-off mtDNA score for predicting clinical pregnancy of euploid embryos
The absolute mtDNA threshold score for the prediction of clinical pregnancy after transfer of an euploid embryo was 0.002, above which no clinical pregnancy was attained.
(100% negative predictive rate). Among all the transferred euploid embryos (n = 338), none with mtDNA score greater than the absolute threshold (n = 25) achieved clinical pregnancy. On the other hand, the transferred embryos with mtDNA score below the absolute threshold value (n = 313), 71.8% achieved clinical pregnancy (n = 225).
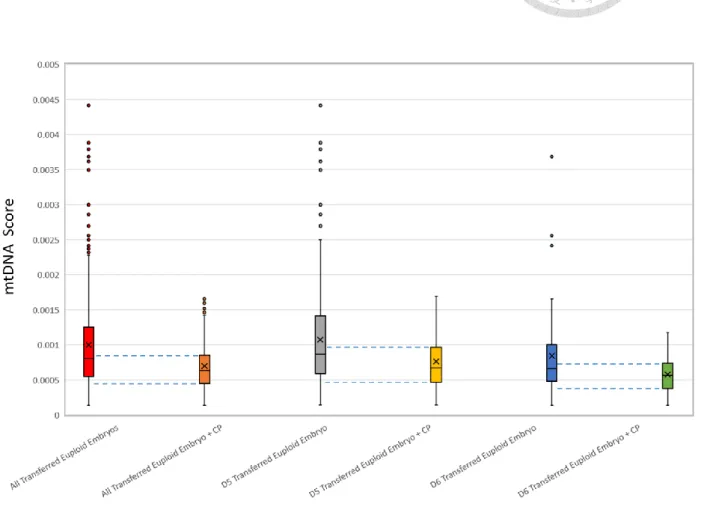
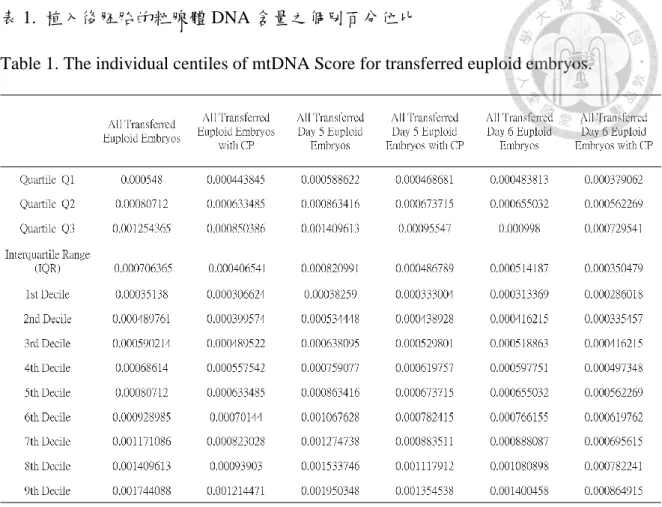
In order to determine the optimal upper and lower threshold of mtDNA score that predict clinical pregnancy for euploid embryo, we performed an analysis of the mtDNA score for all the transferred euploid embryos. Specifically, individual mtDNA score for all the
transferred euploid embryos and the euploid embryos that established successful clinical pregnancy were ranked and stratified into respective centiles and represented as box plots.
A sub-analysis was also performed for transferred euploid embryos of different development rates (Day 5 and Day 6 blastocysts). The results from the above analysis are summarize in Table 1.
When all the transferred euploid embryos were analyzed together, the inter-quartile range (IQR) of mtDNA score for embryos that resulted in clinical pregnancy (Q1 = 0.000443845, Q3: 0.000850386; range = 0.000406541) corresponded approximately to the range of mtDNA score between the median and the 1st decile for all transferred euploid embryos (1st decile = 0.0003513795, median = 0.0008071195; range = 0.00045574).
Similar trends were also observed in the sub-analysis for embryo of different development rate. For transferred Day 5 euploid embryo, the inter-quartile range (IQR) of mtDNA score for embryos that resulted in clinical pregnancy (Q1 = 0.000468681, Q3: 0.00095547;
range = 0.000486789) corresponded approximately to the range of mtDNA score between the median and the 1st decile for all transferred Day 5 euploid embryos (1st decile = 0.00038259, median = 0.0008634155; range = 0.00038259). For transferred Day 6 euploid embryo, the inter-quartile range (IQR) of mtDNA score for embryos that resulted in clinical pregnancy (Q1 = 0.000379062, Q3 = 0.000729541; range = 0.000350479) corresponded approximately to the range of mtDNA score between the median and the 1st decile for all transferred Day 6 euploid embryos (1st decile = 0.000313369, median = 0.0006550315; range = 0.0003416625). Theresults from the above analysis are outlined in Figure 6.
研究討論
The results of the present study collaborated, and at the same time contradicted numerous findings from the past studies on mitochondria and IVF outcomes. First of all, our data provided another robust evidence, that aneuploid embryos contained significantly greater quantity of mtDNA compared to eupoid embryos. Secondly, when considering the embryo development rate, day 5 blastocysts had a significantly higher mtDNA quantity than day 6 blastocysts, independent of embryo ploidy. Thirdly, when factoring the effects of maternal age, no differences in the mtDNA quantity was noted when analyzing blastocysts pooled from subjects aged < 40 and >40 years-old, irrespective of embryo ploidy and development rate. The comparison of embryo euploidy rates, clinical pregnancy rates, and mtDNA score of euploid embryos that resulted in clinical pregnancy between day 5 blastocysts and day 6 blastocysts also showed significant difference favoring day 5 blastocysts. Lastly, the calculated optimal range of mtDNA scores that predicted clinical pregnancy of euploid blastocysts were the values between median and the 1st decile of all the transferred euploid embryos, respectively. The same upper and lower cutoff range were also observed when blastocysts of different development rate were analyzed separatety.
第一節 粒線體於卵子及胚胎發育時期所扮演之角色
The role of mitochondria in oogenesis and embryogenesis
In order to truly appreciate the complex dynamics of mitochondria biogenesis during embryonic development, one must also consider their roles in oogenesis. At early phase of female embryogenesis, the mtDNA content of primordial germ cells (PGCs) first undergoes a marked reduction in quantity, followed by gradual increase during ensuing
stages of oogenesis, peaking at late folliculogenesis when the oocyte reached metaphase II (MII) stage [24]. Following fertilization, due to absence or minimal levels of mtDNA replications factors such as POLGA, POLGB and TFAM, the mtDNA copy number in mammalian embryos either remained constant [25] or reduced [26-27] throughout the cleavage stage up until morula stage, which roughly coincided with the commencement of embryonic genome activation (EGA) / maternal to zygote transition (MZT) [28].
Consequently, the number of mtDNA copies in each blastomere would progressively dwindle after each cell division, limiting individual blastomere’s capacity to generate ATP through OXPHOS, as the organelles become more reliant on anaerobic respiration as energy source [29-30]. Morphology analysis at this stage revealed immature spherical mitochondria with few cristae and low membrane potentials [31]. When the embryo reached the blastocyst stage, the mtDNA replication factors would once again be upregulated, leading to reactivation of mtDNA replication [25-27]. Morphology wise, mature mitochondria at this time would appear as elongated organelles containing swollen cristae, with increased mitochondrial membrane potential and oxygen consumption that facilitate OXPHOS activity and ATP production [31]. This process of initial reduction and subsequent amplification of mtDNA at the end of folliculogenesis constitute a genetic bottleneck effect that allows the preferential passage of only a fraction of mtDNA content to oocytes [32]. It is probable that the founding mitochondria were selected based on their mtDNA integrity and OXOPHOS functions, thus ensuring only the healthiest mitochondria would be preserved, and at the same time minimizing the transmission of abnormal mitochondria genome.
第二節 於染色體套數異常或品質不佳胚胎中 粒線體 DNA 含量增加之原因與意義
The significance of increased mtDNA quantity in embryo with aneuploidy or reduced viability
The vital roles of mitochondria in oocyte and embryo development have been long recognized [33-34]. Therefore, one may wonder, if mitochondria have such important functions, how can increased mitochondria quantity in a cell be detrimental? The possible explanations for such paradoxical relationship may be viewed in context of suboptimal mitochondria quality per se, or the overall dysfunction of embryo development.
In the first scenario, when the embryo contained excessive number of functionally deficient mitochondria, a compensatory mechanism of mitochondria pool expansion would be activated as an effort to restore normal ATP production. However, if most of the newly replicated mitochondria still harbor the same defect, the defective energy state of the embryo would still be unrectified thus affecting embryo viability. Indeed, the above theory merits consideration since mtDNA is highly prone to mutation due to its proximity to the DNA-damaging reactive oxygen species (ROS) generated by the electron transport chain, as well as the absence of protective histone that shields DNA against hydroxyl radical-induced DNA strand breaks [35]. Therefore, the mutation rate of mtDNA has been estimated to be almost 25 times higher than that of nuclear DNA [36]. Tzeng et al. have reported decreased expressions of mitochondrial oxidative phosphorylation genes ND2, CO I, CO II, ATPase 6, CO III, ND3, ND6 and Cyt b in both unfertilized human oocytes and arrested embryos [37]. In addition, the same group also reported increased frequency of mitochondrial gene deletions in unfertilized human oocytes and arrested embryos, with the 4977-bp deletion being the most common [38]. The evidence of compensatory increment of mtDNA content in response to mutation has also been observed in the
oocytes and embryos from patients of myopathy, encephalopathy, lactic acidosis and stroke-like syndrome (MELAS) [39]. In spite of the above findings, a definitive link between mutation of the mitochondrial genome and the increased mtDNA levels associated with reduced implantation has not been demonstrated.
Another plausible theory considers the quality of embryo as a whole, rather than the functional status of mitochondria alone. In the “quiet embryo hypothesis” by Leese et al., the authors proposed that embryos with increased metabolic activity are in fact suffering from certain extrinsic stress (i.e. culture condition) or intrinsic structural defects (i.e., aneuploidy) [40]. Thus greater energy expenditures are required as an effort to restore proper embryonic development, which resulted in compensatory mitochondria replication and a net gain of mtDNA. Since blastocyst stage is the period when significant mtDNA synthesis takes place, which also coincide with the timing of embryo biopsy for PGS, therefore this might explain the phenomenon of embryos with elevated mtDNA levels detected at this stage of development. In theory, the excessive amount of ATP generated could lead to increased free radical production, causing damage to DNA and other organelles, eventually impairing embryo viability [41-42]
第三節 粒線體異質性與試管嬰兒成功率之關係
The role of mitochondrial heteorplasmy in relation to IVF outcomes
The extent of heteroplasmy, defined as the presence of more than one mtDNA variant in the mitochondria of an individual, was often used to determine the quality of mitochondria gene pool. It has been estimated that each person on average carry one heteroplasmy, with one in eight carry disease-associated heteroplasmic variant [43]. Even though mitochondrial heteroplasmy has been extensively implicated in numerous human disease states [44-45] and cancer progression [45-46], its association with the viability of
embryos derived from ART is less clear. In the study by Fraguli et al., varying degrees of heteroplasmy were observed in all of the embryo biopsy samples, irrespective of embryo mtDNA levels and transfer outcomes [10]. On the other hand, Lledo et al. reported a significant positive association between the degree of hetroplasmy and mtDNA copy number [14]. Though a lower trend of implantation rate and ongoing pregnancy rate, along with a higher miscarriage rate were observed for embryos that carried at least one heteroplasmic variant, the differences did not reach statistical significance. In essence, since each cell can contain up to thousands of mitochondria molecules, and each mitochondrion also has multiple copies of mtDNA, therefore a threshold of hetroplasmy, or “mutational load” must be exceed for the phenotypic manifestation of the defect to be evident [47]. For human embryos, it has been estimated that a heteroplasmic level up to 18% could still bear 95% or greater chance of having unaffected offsprings [48]. Base on the above observations, we opted not to incorporate the degree of heteroplasmy in our present study.
第四節 “越少越好?” 粒線體 DNA 含量多寡與胚胎發育速度之關係
“The less the better?” The significance of increased mtDNA quantity and embryo developmental rate
An interesting discovery was made when comparing the mtDNA score from blastocysts with different development rate. The day 5 blastocysts consistently demonstrated a higher mtDNA score than the day 6 blastocysts, even when euploid and aneuploid embryos were considered separately. Similar finding was observed in the study by Lledo et al., and the authors proposed a “dilution effect” as the explanation for such phenomenon [14]. The theory is based on the observation from a mouse model study that found the quantity of mtDNA remained relatively constant between fertilization and the early post-
implantation stage, and no additional mtDNA replication would occur during that period of time. Therefore, with increased rounds of cell divisions in an embryo, the “same”
amount of mtDNA would be distributed among growing number of embryonic cells, and thus by day 6 of development each cell would contain fewer copies of mtDNA than at day 5. The above theory is questionable since no comparison between the transfer outcomes of day 5 and day 6 euploid embryos was performed. Such distinction is crucial, since in order for the above-mentioned “dilution effects” to be valid, the authors would be making the assumption that day 5 and day 6 blastocysts possessed similar developmental competency, therefore it did not matter when biopsy was performed as the mere difference would be just the number of cell divisions.
We hereby propose a “energy threshold” theory for the discrepancy. In our study, all of the blastocysts that reached expanded stage of development (Gardner’s grading ≥ 3CC) on day 5 were biopsied on the same day, while the ones that had a slower growth rate were either biopsied on day 6 when the same criteria were met or not biopsied. The slower blastocyst formation rate could be viewed as inferior developmental competency due to lower number of mitochondria organelles (thus mtDNA) presented, making embryos less energetically favorable to sustain normal blastocyst formation. This theory is backed by the results of the present study that demonstrated significantly higher embryo euploid rate and clinical pregnancy rate for day 5 blastocysts than for day 6 blastocysts. Other studies also collaborated our theory, that embryos that developed into mature blastocysts on day 5 almost always had higher euploidy rate [49-51], implantation rate [52-53], and live birth rate [53-56] when compared day 6 blastocysts, with or without PGS. Therefore, according to the above findings, the lower quantity of mtDNA in an embryo does not always imply increased competency, rather the mtDNA quantity above a certain threshold must also be maintained in order for optimal embryo development.
第五節 粒線體 DNA 量與母體年齡無顯著關聯之原因
The absence of maternal age effect on mtDNA quantity – Why?
In line with previous reports, the present study did not find a significant correlation between maternal age and mtDNA score, regardless of ploidy status. The possible rationale for this observation could be explained by the bias of our sample selection process. Since all of the study’s samples were from embryos that developed successfully either on day 5 or day 6 post fertilization into matured blastocysts, in essence all the embryos which exhibited slower growth rate, poor cellular components, and growth arrest were excluded from the final analysis. Therefore, only the mtDNA content from the embryo that demonstrated adequate degree of developmental competence were analyzed, thus diluting the adverse effects of advancing maternal age. Indeed, similar phenomenon has been observed in past studies with PGS, whereas the clinical pregnancy rate and live birth rate after the transfer of euploid embryos did not demonstrated significant differences across all age groups, even for women of advanced age (>40 year-old) [57- 59]. These results are in accordance with the ultimate goal of embryo selection, which is to minimize the transfer of developmentally compromised embryos.
第五章 研究限制
Study Limitations
The main limitation of the present study was its retrospective single-center design. In addition, the mtDNA was evaluated exclusively in quantitative manner, while the degree heteroplasmy, as well as other important mitochondrial function such as oxygen consumption rate (OCR) and ATP synthesis capacity, were not evaluated. The method mitochondrial quantification used in the present study also have several drawbacks. First of all, as stated previously, the number of trophectoderm cells obtained during biopsy cannot be controlled. Therefore, the quantity of mitochondria DNA must always be presented in a relative manner instead of the actual amount. In theory, the most ideal study design would be the analysis of individual trophectoderm separately. That way, the actual amount of mitochondrial DNA presented would be specifically for a single trophectoderm cell. However, such strategy faces numerous technical challenges under the original study setting. Presently there is no known effective method for separating the individual cell in a given trophectoderm biopsy sample without compromising the integrity the cellular contents. Secondly, with single cell analysis, each cell must undergo its own round of WGA, which would result in variable degree of ADO and preferential amplification among cells within a given sample, making the NGS results more susceptible to technical biases. Thirdly, the overall cost of PGS procedures would be significantly increased with single cell analysis, thus inflicting more financial burden upon patients. For example, with a five cell biopsy sample, a total of five rounds of WGA would need be performed, and more rounds of multiplexing would also be required for processing increased number of WGA samples. It is also technically difficult to infer the number of cells analyzed from the NGS data, since the sample has already undergone WGA. Furthermore, whether a single trophectoderm biopsy can adequately represent the chromosomal and
mitochondrial composition of an entire embryo still merits careful consideration. Lastly, all the evaluated embryos were derived from stimulated IVF cycles from a very specific population. Therefore, whether these findings are translatable in general population warrant further studies.
第六章 研究結論
Conclusions
The results of the present study have proven that the mtDNA quantity alone is a useful indicator that accurately predicts the implantation potential of euploid embryos. With the prevalence of high throughput NGS platforms used in PGS, scientists now are allowed to examine in depth nuclear genome as well as mitochondria genome simultaneously.
Considering the context of our study is to investigate the additional markers of embryo selection for IVF, the task can be considered fait accompli. The results from the present study provided an additional piece of puzzle to the complex yet important roles of mitochondria in embryonic development. Specifically, our data demonstrated that lower mtDNA quantity in an embryo does not always imply increased embryo viability, rather mtDNA above a certain threshold must be maintained in order for optimal embryo development. Further prospective, randomized trials that encompass the multi-faceted mitochondria biology are require to fully elucidate the complex relationship between mitochondria and human reproductions.
參考文獻
References
1. Findikli N, Kahraman S, Saglam Y, et al. Embryo aneuploidy screening for repeated implantation failure and unexplained recurrent miscarriage. Reprod Biomed Online 2006;13(1):38–46.
2. Munné S, Lee A, Rosenwaks Z, Grifo J, Cohen J. Diagnosis of major chromosome aneuploidies in human preimplantation embryos. Hum Reprod 1993;8:2185–2191.
3. Franasiak JM and Scott RT: Contribution of immunology to implantation failure of euploid embryos. Fertil Steril 2017;107:1279–1283.
4. Simon A, Laufer N. Assessment and treatment of repeated implantation failure (RIF).
J Assist Reprod Genet 2012;29(11):1227–39.
5. Dumollard R, Carroll J, Duchen MR, Campbell K, Swann K. Mitochondrial function and redox state in mammalian embryos. Semin Cell Dev Biol 2009;20:346–353.
6. Circu ML, Aw TY. Reactive oxygen species, cellular redox systems, and apoptosis.
Free Radic Biol Med 2010;48:749–762.
7. Giorgi C, Agnoletto C, Bononi A, Bonora M, De Marchi E, Marchi S, Missiroli S, Patergnani S, Poletti F, Rimessi A et al. Mitochondrial calciumhomeostasis as potential target for mitochondrial medicine. Mitochondrion 2012;12:77–85.
8. Ravichandran K, McCaffrey C, Grifo J, Morales A, Perloe M, Munne S, Wells D, Fragouli E. Mitochondrial DNA quantification as a tool for embryo viability assessment: retrospective analysis of data from single euploid blastocysts transfers.
Hum Reprod 2017;32:1282–1292.
9. Diez-Juan, A., Rubio, C., Marin, C., Martinez, S., Al-Asmar, N., Riboldi, M., Díaz- Gimeno, P., Valbuena, D., and Simón, C. Mitochondrial DNA content as a viability score in human euploid embryos: less is better. Fertil Steril 2015; 104: 534–541.
10. Fragouli, E., Spath, K., Alfarawati, S., Kaper, F., Craig, A., Michel, C.-E., Kokocinski, F., Cohen, J., Munne, S., and Wells, D. Altered levels of mitochondrial DNA are associated with female age, aneuploidy, and provide an independent measure of embryonic implantation potential. PLoS Genet 2015;11:e1005241.
11. Fragouli, E.; McCaffrey, C.; Ravichandran, K.; Spath, K.; Grifo, J.A.; Munné, S.;
Wells, D. Clinical implications of mitochondrial DNA quantification on pregnancy outcomes: A blinded prospective non-selection study. Hum Reprod 2017;32:2340–
2347.
12. Victor, AR., Brake, A.J., Tyndall, J.C., Griffin, D.K., Zouves, C.G., Barnes, F.L., and Viotti, M. Accurate quantitation of mitochondrial DNA reveals uniform levels in human blastocysts irrespective of ploidy, age, or implantation potential. Fertil Steril 2017; 107: 34–42.
13. Treff, NR., Zhan, Y., Tao, X., Olcha, M., Han, M., Rajchel, J., Morrison, L., Morin, S.J., and Scott, R.T. Levels of trophectoderm mitochondrial DNA do not predict the reproductive potential of sibling embryos. Hum Reprod 2017; 32:954–962.
14. Lledo B, Ortiz JA, Morales R, García-Hernández E, Ten J, Bernabeu A, Llácer J, Bernabeu R. Comprehensive mitochondrial DNA analysis and IVF outcome. Hum Reprod Open 2018; 2018(4): hoy023.
15. Harton GL, Wells D. Current and Future Preimplantation Genetic Screening, Screening the Single Euploid Embryo: Molecular Genetics in Reproductive Medicine, 2015 Springer (pg.183-184).
16. SurePlex Summary Protocol–Reference Guide. (2018) [PDF file]. Retrieved from https://support.illumina.com/content/dam/illumina-support/documents/
documentation/chemistry_documentation/veriseq-pgs/sureplex-protocol-reference- guide-15053626-01.pdf
17. Deleye L, De Coninck D, Christodoulou C, Sante T, Dheedene A, Heindryckx B, et al. Whole genome amplification with SurePlex results in better copy number alteration detection using sequencing data compared to the MALBAC method. Sci Rep 2015;5:11711.
18. Wells D, Kaur K, Grifo J, Glassner M, Taylor JC, et al. Clinical utilisation of a rapid low-pass whole genome sequencing technique for the diagnosis of aneuploidy in human embryos prior to implantation. J Med Genet 2014;51:553–562.
19. Kung A, Munné S, Bankowski B, Coates A, Wells D. Validation of next-generation sequencing for comprehensive chromosome screening of embryos. Reprod Biomed Online 2015;31:760–769.
20. VeriSeq PGS Library Prep–Reference Guide (2014) [PDF file]. Retrieved from https://support.illumina.com/content/dam/illumina-support/documents/
documentation/chemistry_documentation/veriseq-pgs/veriseq-pgs-library-prep- reference-guide-15052877-03.pdf
21. Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009;25:1754–1760.
22. BlueFuse Multi v4.4 Software Guide (2017) [PDF File]. https://support.illumina.
com/content/dam/illumina-support/documents/documentation/ software_
documentation/bluefuse-multi-software/bluefuse-multi-v4-4-software-guide- 15053620-03.pdf
23. Anderson S , Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger Fet al. Sequence and organization of the human mitochondrial genome. Nature 1981;290:457–465.
24. May-Panloup P, Boucret L, Chao de la Barca JM, Desquiret-Dumas V, Ferré- L'Hotellier V, Morinière C, Descamps P, Procaccio V, Reynier P. Ovarian ageing: the role of mitochondria in oocytes and follicles. Hum Reprod Update 2016;22:725–743.
25. Thundathil J, Filion F, Smith LC. Molecular control of mitochondrial function in preimplantation mouse embryos. Mol Reprod Dev 2005;71:405–413.
26. Spikings EC, Alderson J, St John JC. Regulated mitochondrial DNA replication during oocyte maturation is essential for successful porcine embryonic development.
Biol Reprod 2007;76:327–335.
27. May-Panloup P, Vignon X, Chretien MF, Heyman Y, Tamassia M, Malthiery Y, Reynier P. Increase of mitochondrial DNA content and transcripts in early bovine embryogenesis associated with upregulation of mtTFA and NRF1 transcription factors. Reprod Biol Endocrinol 2005;3:65.
28. Niakan, K. K., Han, J., Pedersen, R. A., Simon, C. & Pera, R. A. R. Human pre- implantation embryo development. Development 2012;139:829–841.
29. Leese HJ. Metabolic control during preimplantation mammalian development. Hum Reprod Update 1995;1:63–72.
30. Van Blerkom J, Davis P, Mathwig V, Alexander S. Domains of high-polarized and low-polarized mitochondria may occur in mouse and human oocytes and early embryos. Hum Reprod 2002;17:393–406.
31. Wilding M, Dale B, Marino M, di Matteo L, Alviggi C, Pisaturo ML, Lombardi L, De Placido G. Mitochondrial aggregation patterns and activity in human oocytes and preimplantation embryos. Hum Reprod 2001;16:909–917.
32. Brown WM. Polymorphism in mitochondrial DNA of humans as revealed by restriction endonuclease analysis. Proc Natl Acad Sci USA 1980;77:3605–9.
33. Van Blerkom J: Mitochondria in human oogenesis and preimplantation
embryogenesis: engines of metabolism, ionic regulation and developmental competence. Reproduction 2004;128:269-280.
34. Dumollard R, Duchen M, Carroll J. The role of mitochondrial function in the oocyte and embryo. Curr Top Dev Biol 2007;77:21–49.
35. Alexeyev M., Shokolenko I., Wilson G., et al. The maintenance of mitochondrial DNA integrity—critical analysis and update, Cold Spring Harb Perspect Biol 2013;5:a012641.
36. Lynch M, Koskella B, Schaack S. Mutation pressure and the evolution of organelle genomic architecture. Science 2006;311:1727–1730.
37. Hsieh RH, Au HK, Yeh TS, Chang SJ, Cheng YF, Tzeng CR. Decreased expression of mitochondrial genes in human unfertilized oocytes and arrested embryos. Fertil Steril 2004;81:1912–918.
38. Hsieh RH, Tsai NM, Au HK, Chang SJ, Wei YH, and Tzeng CR. Multiple rearrangements of mitochondrial DNA in unfertilized human oocytes. Fertil Steril 2002;77:1012–1017.
39. Monnot S, Samuels DC, Hesters L, Frydman N, Gigarel N, Burlet P, Kerbrat V, Lamazou F, Frydman R, Benachi A et al. Mutation dependance of the mitochondrial DNA copy number in the first stages of human embryogenesis. Hum Mol Genet 2013;22:1867–1872.
40. Leese HJ. Quiet please, do not disturb: a hypothesis of embryo metabolism and viability. Bioessays 2002;24:845–9
.
41. Liu H, Trimarchi J, Keefe D. Involvement of mitochondria in oxidative stress induced cell death in mouse zygotes. Biol Reprod 2000;62:1745–53.
42. Van Blerkom J, Davis P, Lee J. ATP content of human oocytes and developmental potential and outcome after in vitro fertilization and embryo transfer. Hum Reprod 1995;10:415–24.
43. Rebolledo-Jaramillo B, Su MS, Stoler N, McElhoe JA, Dickins B, Blankenberg D, Korneliussen TS, Chiaromonte F, Nielsen R, Holland MM, Paul IM, Nekrutenko A, Makova KD. Maternal age effect and severe germ-line bottleneck in the inheritance of human mitochondrial DNA. Proc Natl Acad Sci USA 2014;111(43):15474-9.
44. Kenney MC, Chwa M, Atilano SR, Falatoonzadeh P, Ramirez C, Malik D, Tarek M, Cáceres-del-Carpio J, Nesburn AB, Boyer DS, et al. Inherited mitochondrial DNA variants can affect complement, inflammation and apoptosis pathways: Insights into mitochondrial-nuclear interactions. Hum Mol Genet 2014;23:3537–3551.
45. Kenney MC, Chwa M, Atilano SR, Falatoonzadeh P, Ramirez C, Malik D, Tarek M, Del Carpio JC, Nesburn AB, Boyer DS, et al. Molecular and bioenergetic differences between cells with African versus European inherited mitochondrial DNA haplogroups: Implications for population susceptibility to diseases. Biochim Biophys Acta 2014;1842:208–219.
46. Atilano SR, Malik D, Chwa M, et al. Mitochondrial DNA variants can mediate methylation status of inflammation, angiogenesis and signaling genes. Hum Mol Genet 2015;24:4491–4503.
47. Rossignol R, Faustin B, Rocher C, Malgat M, Mazat JP, Letellier T. Mitochondrial threshold effects. Biochem J 2003;15:751–762.
48. Hellebrekers DM, Wolfe R, Hendrickx AT, de Coo IF, de Die CE, Geraedts JP, Chinnery PF, Smeets HJ. PGD and heteroplasmic mitochondrial DNA point mutations: a systematic review estimating thevchance of healthy offspring. Hum Reprod Update 2012;18:341–349.
49. Taylor TH, Patrick JL, Gitlin SA, Wilson JM, Crain JL, Griffin DK. Comparison of aneuploidy, pregnancy and live birth rates between day 5 and day 6 blastocysts.
Reprod BioMed Online 2014;29:305–10.
50. Piccolomini MM, Nicolielo M, Bonetti TC, Motta EL, Serafini PC, Alegretti JR.
Does slow embryo development predict a high aneuploidy rate on trophectoderm biopsy? Reprod Biomed Online 2016;33:398-403.
51. Kaing A, Kroener LL, Tassin R, Li M, Liu L, Buyalos R, Hubert G, Shamonki M.
Earlier day of blastocyst development is predictive of embryonic euploidy across all ages: essential data for physician decision-making and counseling patients. J Assist Reprod Genet 2018;35:119–125.
52. Shapiro BS, Richter KS, Harris DC, Daneshmand ST: A comparison of day 5 and day 6 blastocyst transfers. Fertil Steril 2001;75:1126-1130.
53. Liebermann J, Mathews J, Machlin J, Baranyi L, Elzy I, Brohammer R, Nani J, Marut E. Euploid single frozen day 5 versus day 6: can we expect equal outcome? Fertil Steril 2017;108:e284.
54. Irani M, O'Neill C, Palermo GD, Xu K, Zhang C, Qin X, Zhan Q, Clarke RN, Ye Z, Zaninovic N, Rosenwaks Z. Blastocyst development rate influences implantation and live birth rates of similarly graded euploid blastocysts. Fertil Steril 2018; 110:95- 102.
55. Ferreux L, Bourdon M, Sallem A, Santulli P, Barraud-Lange V, Le Foll N, et al. Live birth rate following frozen-thawed blastocyst transfer is higher with blastocysts expanded on Day 5 than on Day 6. Hum Reprod 2018;33:390–8.
56. Haas J, Meriano J, Laskin C, Bentov Y, Barzilay E, Casper RF, Cadesky K. Clinical pregnancy rate following frozen embryo transfer is higher with blastocysts vitrified on day 5 than on day 6. J Assist Reprod Genet 2016;33:1553–1557.
57. Barash O, Willman SP, Ivani KA, Wachs DS, Rabara FB, Huen N, Weckstein LN.
Single embryo transfer as an imperative choice for patients over 38 years old in autologous IVF PGS cycles. Reprod Biomed Online 2018;36:e29-e30.
58. Forman EJ, Tao X, Ferry KM, Taylor D, Treff NR, Scott RT Jr. Single embryo transfer with comprehensive chromosome screening results in improved ongoing pregnancy rates and decreased miscarriage rates. Hum Reprod 2012; 27:1217–22.
59. Simon AL, Kiehl M, Fischer E, et al. Pregnancy outcomes from more than 1,800 in vitro fertilization cycles with the use of 24-chromosome single-nucleotide polymorphism-based preimplantation genetic testing for aneuploidy. Fertil Steril 2018;110:113–121.
圖 1. 胚胎染色體套數與粒線體 DNA 含量之關係
Figure 1. The relationship between mtDNA score to embryo poidy, with consideration of difference in embryo development rate.
圖 2. 胚胎發育速度與粒線體 DNA 含量之關係
Figure 2. The relationship between mtDNA score to developemnt rate, with consideration of difference in embryo ploidy
.
圖 3. 母體年齡與粒線體 DNA 含量之關係
Figure 3. The relationship between mtDNA score to maternal age, with consideration of difference in embryo ploidy and development rate.
圖 4. 第五天和第六天囊胚之染色體正常比例以及植入後臨床懷孕率之比較 Figure 4. The comparison of embryo euploidy rates and clinical pregnancy rates between Day 5 blastocysts and Day 6 blastocysts.
圖 5. 植入後有成功達成臨床懷孕的囊胚與植入後未能達成臨床懷孕的囊胚中粒腺 體含量之比較.
Figure 5. The comparison of mtDNA Score between transferred the euploid embryos that resulted in clinical pregnancy and the euploid embryos that failed to establish clinical pregnancy.
圖 6. 粒線體 DNA 的含量用於預測胚胎之懷孕率
Figure 6. The optimal upper & lower threshold mtDNA Score that predict clinical pregnancy for euploid embryos
表 1. 植入後胚胎的粒腺體 DNA 含量之個別百分位比
Table 1. The individual centiles of mtDNA Score for transferred euploid embryos.