行政院國家科學委員會補助專題研究計畫 研究成果報告(完整版)
前列腺素 E2 調控巨噬球細胞在子宮內膜異位症成因之研究 (第三年)
計畫類別: 個別型計畫
計畫編號:NSC
97-2314-B-006-020-MY3
執行期間:2008
年8
月01
日至2011
年07
月31
日計畫主持人:吳孟興
成果報告類型(依經費核定清單規定繳交):完整報告
本成果報告包括以下應繳交之附件:
出席國際學術會議心得報告:
2011 年第 27 屆歐洲生殖醫學會(ESHRE)年會國科會計畫出國報告書
處理方式:二年後可公開查詢
執行單位:成功大學醫學院婦產學科
中 華 民 國 一百 年 十 月 二十 日
行政院國家科學委員會專題研究 研究成果報告(完整版)
前列腺素 E2 調控巨噬球細胞在子宮內膜異位症成因之研究 (第三年)
計畫類別:個別型計畫
計畫編號:NSC 97-2314-B-006-020-MY 執行期限:2008年8 月01 日至2011 年07月 31日
主持人:吳孟興 成功大學醫學院婦產學科 成果報告類型:完整報告
處理方式:二年後可公開查詢 執行單位:成功大學醫學院婦產學科
中 華 民 國 一百 年 十 月 二十 日
一、中英文摘要
子宮內膜異位症是造成婦女腹部疼 痛及經痛一個常見的疾病,也因此會影 響婦女的生活品質。子宮內膜異位症是 影響在生殖年齡 2-10% 的婦女,儘管發 生頻率高但是造成子宮內膜異位症發展 的真正原因目前仍然不清楚,其中一個 關鍵性因素是局部免疫系統的改變或功 能不良,研究報告指出腹膜腔內巨噬球 與子宮內膜異位症嚴重程度有高度地相 關性,從子宮內膜異位症病人分離的腹 膜腔巨噬球傾向有強烈地細胞動力素製 造能力但是其吞噬能力不足。有許多其 他的因子例如類固醇、前列腺素、及血 管新生的因子也都與子宮內膜異位症的 病理機制有關。其中前列腺素 E2 是腹膜 液中重要的因子,而前列腺素 G/H 合成 酶(COX)-2 是主要控制其生成的酵素,
我們發現子宮內膜異位症患者的腹膜腔 巨噬球細胞表現 COX-2 的能力明顯增 加,而且與腹膜腔巨噬球細胞的活化與 腹腔液中前列腺素的增加有關。腹膜腔 前列腺素的增加繼而使倒流入腹腔內經
血的子宮內膜組織本身得以產生荷爾蒙 並異位穩定附著在骨盆腔及持續的生 長。腹膜腔巨噬球細胞的功能及形態上 在子宮內膜異位症患者是異常的,而子 宮內膜異位症的異位子宮內膜組織正是 製造 COX-2 和前列腺素 E2 的來源,推測 有可能腹腔中局部內分泌環境影響尤其 是腹腔液中巨噬球細胞的功能變化,支 撐異位子宮內膜的生長而存活下來,並 且使得子宮內膜異位症更加惡化。因此 這個 3 年計畫的目的是想要去探討前列 腺素 E2 對腹膜腔內巨噬球的表現是經 由下游的那些訊息傳遞產生影響腹膜腔 內巨噬球的吞噬能力,我們發現腹膜腔 內巨噬球表面與其吞噬能力有相關的因 子例如 CD36 以及 annexin II 在腹膜腔 內巨噬球的下降變化。子宮內膜異位症 患 者 的 腹 膜 腔 內 巨 噬 球 CD36 以 及 annexin II 的表現受到前列腺素 E2 的 刺激後下降,而且與腹膜腔內巨噬球的 吞噬功能有關。前列腺素 E2 如何對腹腔 液中巨噬球細胞形狀以及功能產生變化 或影響可能是經由 EP2 和 EP4 接受器延 續 PKA 訊息傳遞發揮作用。這個研究可
以讓我們發現前列腺素 E2 除了在子宮 內膜或異位的組織有異常表現外,也會 影響腹膜腔內巨噬球,並了解前列腺素 E2 如何影響腹膜腔內巨噬球形狀與吞噬 功能的機制,其次了解前列腺素 E2 的訊 息傳遞與腹膜腔內巨噬球上各種細胞表 面因子互動關係,藉此瞭解病理機轉進 而有利於找出適合治療子宮內膜異位症 的方式。
關鍵詞:子宮內膜異位症,腹膜腔內巨 噬球,吞噬功能,前列腺素 E2。
Endometriosis is a mysterious disease which affects as many as 10-15% of all women of reproductive age. Despite the high frequency with which the disease is encountered, the pathophysiology of the disease still remains unclear. One theory for the development of endometriosis suggests an aberrant immune system as a causative factor. Macrophage hyperactivation in patients with endometriosis plays a crucial role in the pathogenesis of this disease. They increase the ability of inflammatory cytokine production, but decrease phagocytotic capability. Coupling with the increased resistance to macrophage-mediated cytolysis, ectopic endometrial cells can survive within the peritoneal cavity of patients with endometriosis. However, the exact role of peritoneal macrophages in the formation of endometriosis remains to be clarified. PGE2 is the major factor in the peritoneal fluid. In this study, we report here that the phagocytic ability of macrophages is reduced in peritoneal macrophages isolated from women with endometriosis. In-depth investigation revealed that the level of CD36, a class B scavenger receptor, in peritoneal macrophages derived from women with endometriosis was lower than that in normal macrophages. Blockage of CD36 function by neutralized antibody or knocking down CD36 using siRNA impaired the phagocytic ability of normal macrophages. In contrast, forced expression of CD36 in macrophages
isolated from women with endometriosis restored phagocytic ability. Herein, we further report that prostaglandin (PG) E2, via the EP2 receptor-dependent signaling pathway, inhibits the expression of CD36 in peritoneal macrophages, resulting in reduced phagocytic ability.
PGE2-mediated inhibition of macrophage phagocytic capability was restored by ectopic expression of CD36. Treatment with PGE2 inhibited CD36-dependent phagocytosis of peritoneal macrophages and increased the number and size of endometriotic lesions in mice. In contrast, blockade of PGE2 production by cyclooxygenase inhibitors enhanced the phagocytic ability of peritoneal macrophages and reduced endometriotic lesion formation. Similarly, decreased annexin II mRNA expression was noted in peritoneal macrophages of women with endometriosis. The expression of annexin II in macrophage was inhibited by PGE2, and the inhibitory effect was mediated via EP2/EP4 receptor-dependent signaling pathway. This decreased annexin II level in endometriotic peritoneal macrophage was reversed on treatment with selective COX inhibitors. The results of this 3-year proposed project should provide new insights of peritoneal macrophages for better understanding of the formation of endometriosis. Taken together, we identified that the scavenger receptor CD36 and annexin II is reduced in the peritoneal macrophages of women with endometriosis, which leads to a decrease of the phagocytic ability of macrophages.
Our findings reveal a potential mechanism of immune dysfunction during endometriosis development and may contribute to the design of an effective prevention/treatment regimen.
Key Words: endometriosis , peritoneal macrophages , phagocytosis , CD36 , annexin II, prostaglandin E2
二、緣由與目的
Endometriosis is a common gynecological disorder, characterized by the presence of endometrial tissue outside of the uterine cavity, with a complex, multifactorial aetiology that causes chronic pelvic pain, dysmenorrhea, and even infertility. The prevalence of this disease is about 10-15%
among women of reproductive age. The underlying pathophysiological mechanism is still enigmatic. Although retrograde menstruation has been suggested to be the crucial constituent in the development of endometriosis (Sampson, 1927), factors allowing the implantation and propagation of endometriotic lesions are largely unclear.
Aberrant production of steroids by ectopic
endometriotic lesions and alteration/dysfunction of the immune system
may lead to the development of endometriosis.
During the development of endometriosis, immune cells are recruited into the peritoneal cavity. Among these immune cells, macrophages are the dominant cell type in the peritoneal cavity and are involved in phagocytosis and inflammation, especially in cleaning the retrograded endometrial debris (Haney et al, 1981, Dunselman et al, 1988).
Nevertheless, the role of macrophage in the pelvic cavity, where it is recruited due to sterile inflammation triggered by discharge of endometrial debris, is like a double-edged sword. On one hand, macrophage may involve in direct removal of endometrial cells discharged into pelvic cavity. On the other, secretion of cytokines or growth promoting factors by this cell type may facilitate the proliferation or implantation of endometrial cells, thus results in formation of endometriosis. It has been suggested that peritoneal macrophages isolated from patients with endometriosis have phenotypic and functional alterations, which is highly associated with endometriosis (Dmowski et al, 1994, Raiter-Tenenbaum et al, 1998).
Nevertheless, the mechanism of suppressed phagocytotic capability of macrophages in endometriosis is poorly understood.
The scavenger function of macrophage is mediated via, at least, two lines of mechanisms. The first line of mechanism is the secretion and activation of MMPs to
break down the extracellular matrix of foreign entities (Osteen et al., 2003).
Macrophages can secrete MMP-2, 7, 9, and 12, all of which can degrade elastin and have been implicated to play important roles in the pathogeneses of emphysema and aortic aneurysm (Curci et al., 1998; Dhami et al., 2000). MMP-9 has been suggested to involve in cell migration occurring in the various physiological and pathological processes such as tumor cell invasion and chronic inflammation by facilitating the destruction of the type IV collagen-containing basement membrane which separate the epithelial and stromal compartment (McMillan et al., 1996). Beside the ECM degradations, MMP-9 can activate several cytokines such as latent transforming growth factor (TGF)- and pro-tumor necrosis factor (TNF)- to their active forms (Yu and Stamenkovic, 2000; Sternlicht and Werb, 2001b). Thus, MMP-9 not only plays important roles in macrophage’s scavenger activity, it also has significant impacts on the immune system’s defensive mechanism.
Annexin II is a peripheral membrane protein that is widely distributed and binds to negatively charged phospholipids in a Ca2+-dependent manner. Annexin II plays an important role in the pathophysiological action of macrophages during the inflammatory process. Annexin serves as an activator of MMPs by facilitating the interaction between t-PA and plasminogen with subsequent activation of plasmin (Brownstein et al, 2004). Plasmin serves as a physiological activator which, in turn, converts pro-MMP-9 to active MMP-9. Thus, annexin acts as a receptor–effector of MMP through the activation of plasmin. Therefore, it is important to evaluate the expression of annexin in peritoneal macrophages and its relationship with MMP-9 expression in order to characterize the etiology of endometriosis.
The second line of scavenging activity involves expression of scavenger receptors on the macrophages to enhance the uptake and degradation of cell debris (Febbraio et al., 2001; Linton and Fazio, 2001). Macrophages express several scavenger receptors that enable taking up pathogens and apoptotic
cells, phosphatidylserine, and oxidized lipoprotein (Febbraio et al., 2001; Linton and Fazio, 2001). As a class, these proteins tend to recognize polyanionic macromolecules and may have physiological functions in the recognition and clearance of foreign entities.
Based on their multidomain structure, the scavenger receptors have now been classified into eight different classes (Class A, B, C, D, E, F, G and H). The ligand-binding domains of scavenger receptors can recognize a wide range of negatively charged macromolecules, including oxidized low-density lipoproteins, damaged or apoptotic cells, and pathogenic microorganisms. After binding, these ligands can be either internalized by endocytosis or phagocytosis, or remain at the cell surface and mediate adhesion or lipid transfer through carveolae. Under physiological conditions, scavenger receptors serve to scavenge or clean up cellular debris and other related materials, and they play a role in host defense. In pathological states, they mediate the recruitment, activation and transformation of macrophages and other cells which may be related to the development of atherosclerosis and to disorders caused by the accumulation of denatured materials, such as Alzheimer's disease (Yamada et al, 1998). Considering the functional role of scavenger receptors in mediating phagocytosis, it is reasonable to hypothesize that the levels of scavenger receptors in peritoneal macrophages may play an important role in phagocytosis and thus the pathogenesis of endometriosis.
Prostaglandins (PGs) are known, for many decades, to play pivotal roles in many physiological and pathological processes including modulation of immune responses.
In our previous study, peritoneal macrophages of women with endometriosis appear to have acquired the capacity to synthesize PG owing to over-expression of COX-1 and/or COX-2, leading to elevationof PGE2 and PGF2 concentrations in the peritoneal fluid (Wu et al., 2002).
Furthermore, increased amounts of cyclooxygenase-2 have also been reportedin ectopic and eutopic endometria of women with endometriosis (Wu et al, 2005a).
We hypothesize that decreased
phagocytotic capability of peritoneal macrophages in women with endometriosis may be due to decreased expression and activity of MMP and/or reduced level of scavenger receptors on the surface of macrophages. The factors that are capable of inhibiting MMP activity and scavenger receptor expression are likely to be presence in the peritoneal fluid of women with endometriosis. In deed, in our previous study (Wu et al, 2005b), peritoneal macrophage isolated from women with severe stages of endometriosis has decreased concentration and activity of MMP-9 without change of tissue inhibitor of matrix metalloproteinase.
Reduced MMP-9 activity was reproduced when peritoneal macrophages were treated with the dilution of peritoneal fluid derived from patients with endometriosis but not from normal women. Further investigation reveals that PGE2 is the major factor in the peritoneal fluid that inhibits MMP-9 expression and activity. This previous result demonstrated that PGE2-inhibited MMP-9 expression and activity may contribute to the decreased phagocytotic capability of peritoneal macrophage. Nevertheless, whether decreased MMP-9 expression by PGE2 alone accounts for the attenuation of phagocytosis or is (are) there other factor (factors) also contributing to the functional loss of phagocytotic ability of macrophages remain unknown. This proposed study is designed to further investigate the mechanism of reduced phagocytotic capacity of peritoneal macrophages in women with endometriosis.
As elaborated above, annexin II plays an important role in the activation of MMPs.
Whether annexin II takes part in the reduced activity of MMP-9 in peritoneal macrophage is not known. In addition, whether PGE2 can mediate the expression of annexin II thus modulate MMP-9 activity has never been reported. Most importantly, does reduced phagocytotic ability by PGE2 be mediated by inhibiting the expression of the scavenger receptors in macrophage? If so, what are the cellular and molecular mechanisms responsible for PGE2-mediated inhibition of scavenger receptor expression?
三、研究結果與討論
RESULTS
Peritoneal macrophages of women with endometriosis have reduced phagocytic ability
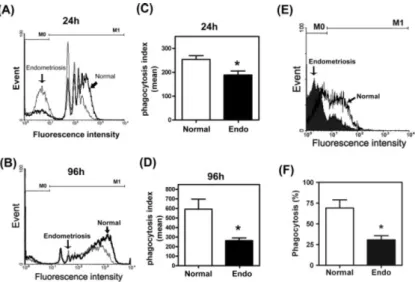
To investigate the role of macrophages in the development of endometriosis, we examined the phagocytic ability of peritoneal macrophages in normal and endometriotic women. The results showed that peritoneal macrophages derived from endometriosis patients had reduced phagocytic ability compared to those derived from normal women (Figure 1A, B). Notably, the number of macrophages with great phagocytic ability was significantly less in the endometriosis group than than in the normal group. The phagocytic index, the mean of ingested beads per macrophage, was also significantly lower in the endometriosis group (Figure 1C, D).
Although the use of synthetic beads is an excellent model to study phagocytosis, it is necessary to demonstrate that a similar mechanism was adopted by macrophages in taking up biological substrates. Therefore, we used FITC-conjugated annexin V-labelled apoptotic Jurkat cells as substrates to repeat the above-described experiments. As expected, peritoneal macrophages isolated from patients with endometriosis had reduced ability in phagocytosing apoptotic cells (Figure 1E, F).
This result was similar to those using FITC-labelled beads as substrates.
Expression of CD36 is downregulated in peritoneal macrophages of women with endometriosis
To evaluate whether reduced phagocytic activity in endometriotic macrophages was due to alteration in cell surface scavenger receptors, mRNAs encoding the SRA and SRB families were evaluated by RT–PCR. The results revealed a marked decrease of CD36 (also known as SR-BIII) in endometriotic macrophages compared to that in normal macrophages (Figure 2A). No significant difference between normal and endometriotic macrophages was found in mRNAs encoding SR-AI, SR-AII, SR-BI and SRBII. Transcript of SR-AIII was not detected in either normal or
endometriotic samples (Figure 2A). Further analysis revealed that phagocytic ability and levels of CD36 were not different between macrophages derived from different stages of endometriosis (data not shown); thus, the data were combined as the ‘endometriosis group’.
Since only CD36 showed a significant decrease in endometriotic macrophages, we thus focused on the role of CD36 in causing the reduction of phagocytic ability of macrophages. First, the expression level of CD36 mRNA in freshly isolated normal and endometriotic macrophages was analysed by real-time RT–PCR. The result showed that the level of CD36 in endometriotic macrophages was significantly lower compared to that of normal macrophages (Figure 2B). As a control, levels of SRBI were also quantified and no difference between normal and endometriotic macrophages was observed (Figure 2C). Since scavenger receptors only function when expressed on the cell surface, we next quantified membrane CD36 using a flow cytometer. The result showed that the membrane CD36 was reduced in peritoneal macrophages freshly isolated from women with endometriosis compared to that in peritoneal macrophages isolated from normal women (Figure 2D, E).
Together, these results suggest that reduced phagocytic ability of peritoneal macrophages derived from women with endometriosis might be due to insufficient amounts of CD36 present in these cells.
Reduced expression of CD36 in peritoneal macrophages of women with endometriosis is not systemic
It is possible that reduced CD36 expression and thus phagocytic ability in peritoneal macrophages isolated from women with endometriosis might be due to systemic defect in immune system. To test such possibility, we isolated mononuclear cells from peripheral blood and peritoneal macrophages from women with and/or without endometriosis and evaluated levels of CD36 in these cells. Concordant with previous results, macrophages isolated from peritoneal macrophages expressed less amounts of CD36 than those isolated from endometriosis-free women (Fig 3). In contrast, monocytes/macrophages derived from peripheral blood of normal and endometriotic
individuals expressed similar level of CD36 (Fig 3) indicating peripheral blood-derived monocytes/macrophages did not have defect in expressing CD36. These results provide evidence to demonstrate that difference in CD36 levels in normal and endometriotic peritoneal macrophages is not systemic.
CD36 is the functional scavenger receptor that controls phagocytic ability of macrophages
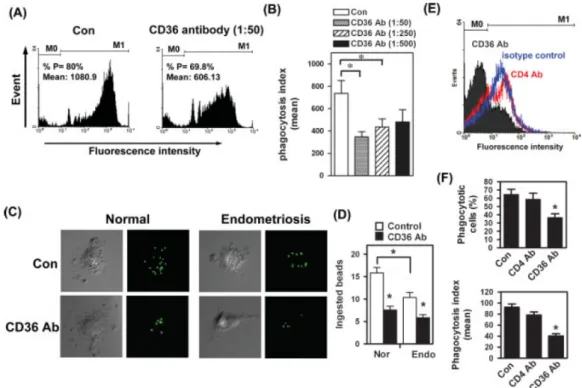
To test whether CD36 indeed contributed to the phagocytic ability of macrophages, peritoneal macrophages isolated from endometriosis-free women were treated with monoclonal antibody to block the function of CD36 and phagocytic ability was determined. Administration of CD36 antibody dose-dependently inhibited phagocytic ability, while mouse IgA (isotypic control) had no effect (Figure 4A, B).
Furthermore, confocal microscopy was used to directly visualize and count the fluorescent beads ingested by macrophages. As shown in Figure 1C, the number of beads ingested by individual macrophages can be quantified by counting the green fluorescent dots. The resultd demonstrated that the number of fluorescent dots was less in macrophages treated with anti-CD36 antibody (Figure 4C, D). Parallel to this experiment, we also treated peritoneal macrophages isolated from women with endometriosis. Concordant with previous observations, the number of fluorescent beads ingested by peritoneal macrophages derived from women with endometriosis was less than that ingested by normal peritoneal macrophages (Figure 4C, D). In addition, treatment of macrophages with anti-CD36 antibody, but not irrelevant antibody (anti CD4 antibody), also resulted in a reduced number of ingested apoptotic Jurkat cells (Figure 4E, F). Taken together, these data demonstrated that CD36 clearly plays an important role in the phagocytic ability of macrophages.
Inhibition of CD36 expression impairs phagocytosis of normal macrophages
To demonstrate that the reduced phagocytic ability of endometriotic
macrophages was a result of down-regulation of CD36, we employed a siRNA knockdown approach to test that CD36 is indispensable for the suppression of phagocytic ability.
SiRNAs against CD36 were transfected into normal macrophages (those with greater levels of CD36 and higher phagocytic ability) and phagocytic activity was determined.
Expression of CD36 mRNA was significantly inhibited by siRNA 1 and 2 but not siRNA 3 or scrambled GC content-matched siRNA (Figure 5A). The expression of OAS-1 was not elicited by any siRNA, demonstrating that the knockdown effect was specific (Figure 5A). In addition, levels of other scavenger receptors were not affected by CD36 siRNA (Figure 5A and data not shown). Confocal microscopy clearly showed the diminishment of membrane CD36 in siCD36-transfected macrophages but not in those transfected with scramble siRNA (Fig 5B). Flow cytometric analysis further demonstrated that levels of plasma membrane CD36 were reduced nearly 80% by siRNA (Figure 5C). Concordantly, phagocytic ability was markedly reduced in CD36-knockdown macrophages but not those transfected with scramble siRNA (Figure 5D, E).
To demonstrate that knocking down CD36 indeed impairs ability to phagocytose apoptotic cells, FITC-conjugated annexin V-labelled apoptotic Jurkat cells were used as substrates. The results showed that knocking down CD36 inhibited the phagocytic ability of macrophages and the number of macrophages with phagocytic ability (Figure 5F, G). The results were consistent with those using FITC-labelled latex beads as substrates (Figure 5D, E), suggesting that CD36 is involved in phagocytosing synthetic beads and apoptotic cells.
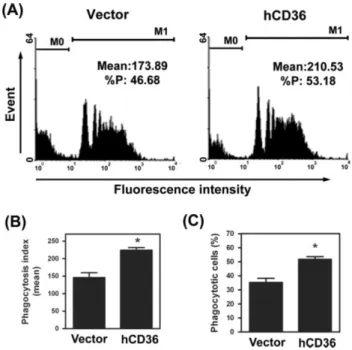
Restoration of CD36 in endometriotic macrophages rescues phagocytic ability
We aimed to test whether the phagocytic ability of peritoneal macrophages derived from women with endometriosis (those with low CD36 levels and poor phagocytic ability) can be restored by ectopic expression of CD36. After trying all currently available
methodologies, microporation yielded the best result with roughly 25-30% of macrophages were successfully transfected with full-length human CD36 cDNA. Force expression of CD36 in endometriotic macrophages enhanced phagocytic ability (Figure 6). The mean number of beads ingested by macrophages and the number of macrophages with phagocytic ability were significantly increased in groups with ectopic expression of CD36 (Figure 6B, C). These data clearly demonstrated that ectopic expression of CD36 is sufficient to rescue the loss of phagocytic ability in low-CD36 endometriotic macrophages.
PGE2-Induced Suppression of Macrophage CD36 Expression and Phagocytic Activity
Toward our goal of addressing mechanistic issues, we aimed to identify local-acting factors that could contribute to the observed reduced expression of CD36 in endometriotic peritoneal macrophages. We screened several previously identified candidate factors (including PGE2) that are elevated in peritoneal fluid of women with endometriosis34 for their potential effects on the mRNA levels of scavenger receptors, and we found that PGE2 potently down-regulated CD36 mRNA level in macrophages (Figure 7A). In contrast, other class A and class B scavenger receptors (SR-AI, SR-AII, SR-BI, and SR-BII) were not affected by PGE2 treatment. Transcripts of SR-AIII were not detected in macrophages (Figure 7A). Next, we used quantitative RT-PCR to test whether PGE2 can inhibit CD36 mRNA expression in both normal and endometriotic peritoneal macrophages. In normal macrophages, the level of CD36 mRNA increased over time in the absence of PGE2, indicating auto-activation of macrophages when cultured on a solid surface (Figure 7B). Treatment with PGE2 acutely and profoundly inhibited CD36 expression in macrophages derived from normal women, as well as endometriosis patients (Figure 7B). Notably, the basal level of CD36 mRNA in endometriotic macrophages was significantly lower than that in normal macrophages, and the level in PGE2-treated normal macrophages was similar
to that in endometriotic macrophages without PGE2 treatment after 24 hours (Figure 7B).
These results implied that reduced expression of CD36 in endometriotic macrophages is likely due to exposure to PGE2 in the peritoneal fluid of women with endometriosis.
To test whether the pattern of CD36 expression we observed is a unique feature of peritoneal macrophages, human U937 monocytic cells were induced by TPA (10 nmol/L) to differentiate into macrophages, and levels of CD36 were determined after PGE2 treatment. Indeed, CD36 expression was observed in TPA-induced U937 cells. Similar to what was observed in human peritoneal macrophages, the levels of CD36 mRNA in TPA-induced U937 cells increased with time in culture (Figure 1C), and treatment with PGE2 significantly decreased the levels of CD36 mRNA and protein (Figure 7, D and E).
These data demonstrated that regulation of CD36 by PGE2 is a common phenomenon of the monocyte/macrophage lineage.
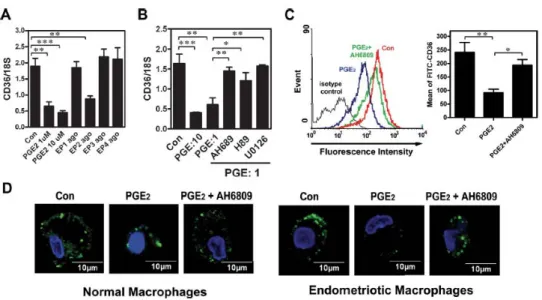
PGE2-Induced Suppression of CD36 Expression Is Mediated by the EP2 Receptor
We next addressed the mechanism of PGE2-induced suppression of CD36 expression. Results from RT-PCR revealed that all four EP receptors were expressed by macrophages, with no substantial difference between normal and endometriotic cells.
Treatment of macrophages with PGE2 (1 or 10 μmol/L) and various EP receptor agonists revealed that PGE2-induced inhibition of CD36 expression was mediated by the EP2 receptor (Figure 8A). These data were supported by the result that treatment with the EP2 receptor antagonist (AH6809) or inhibitors of signaling kinases (protein kinase A and mitogen-activated protein kinase kinase) acting downstream of the EP2 receptor significantly blocked PGE2-induced CD36 down-regulation (Figure 8B). Concordant with the mRNA data, the membrane form of CD36 was markedly decreased in PGE2-treated macrophages, whereas administration of the EP2 receptor antagonist abolished that down-regulation (Figure 8C). These results were confirmed by confocal microscopy,
which clearly showed a reduction in CD36-specific fluorescence intensity in PGE2- treated macrophages but not in macrophages pre-treated with AH6809 (Figure 8D).
Down-Regulation of CD36 by PGE2 Inhibits Phagocytosis
To directly test whether PGE2 treatment of macrophages inhibited their phagocytic function, macrophages isolated from normal women (those with high CD36 levels and a high level of phagocytic ability) were treated with PGE2; indeed, PGE2 significantly decreased both the phagocytic capacity of each cell and the total number of phagocytic cells (Figure 9, A and B). Both effects were reversed by co-treatment with the EP2 receptor antagonist (Figure 9, A and B). These flow cytometry results were confirmed by confocal laser microscopy, which could distinguish phagocytosed beads from potentially adhered beads. Figure 3C shows representative images of macrophages with ingested beads, the mean values of which were 15.0 ± 1.5, 4.7 ± 1.1, and 10.3 ± 1.5 in the control, PGE2-, and PGE2_AH6809-treated groups, respectively (Figure 9D). These results demonstrated that PGE2 suppresses macrophage phagocytosis via an EP2 receptor-dependent pathway.
Ectopic Expression of CD36 Rescues PGE2-Inhibited Phagocytosis
We next used a CD36 overexpression approach to demonstrate that CD36 is indispensable for PGE2-mediated suppression of the phagocytic ability of peritoneal macrophages. Human CD36 cDNA was ectopically expressed in peritoneal macrophages isolated from women with endometriosis (those with low CD36 levels) to restore phagocytic ability. After trying all currently available methodologies, electroporation yielded the best result with roughly 25% of macrophages being successfully transfected with full-length human CD36 cDNA (data not shown). Ectopic expression of CD36 in endometriotic macrophages enhanced phagocytic ability (Figure 10, A and B). More significantly, PGE2-inhibited phagocytic ability was also restored by ectopic expression of CD36
(Figure 10, A and B). The mean number of beads ingested by macrophages was higher in groups with ectopic expression of CD36, whereas the total number of macrophages with phagocytic ability was not significantly increased owing to the small percentage of macrophages that were successfully transfected. Nevertheless, these data clearly demonstrated that ectopic expression of CD36 is sufficient to rescue the loss of phagocytic ability in low-CD36 endometriotic macrophages and in PGE2- treated macrophages.
Expression of annexin A2 was reduced in peritoneal macrophages isolated from endometriosis patients
To determine annexin A2 is responsible for reduced phagocytosis, peritoneal macrophages were isolated from patients with or without endometriosis and the expression of MMP-9 was also examined by real time RT-PCR. Our data revealed that mRNA levels of annexin A2 and MMP-9 in peritoneal macrophages isolated from endometriosis were lower than that from normal macrophage (Figure 11).
PGE2 inhibited expression of annexin A2 mRNA expression
To further determine whether PGE2 can inhibit annexin A2 mRNA expression in peritoneal macrophages, peritoneal macrophages isolated from normal and endometriotic women were treated with 10mM PGE2 for 24 hr and expression of annexin A2 mRNA was examined using quantitative real time RT-PCR (Figure 12). In normal peritoneal macrophages, the level of annexin A2 mRNA increased over time in the absence of PGE2, indicating auto-activation of macrophages when cultured on a solid surface.
Treatment with PGE2 inhibited annexin A2 expression in peritoneal macrophages derived from normal women, as well as endometriosis patients. In addition, the basal level of annexin A2 mRNA in endometriotic macrophages was significantly lower than that in normal macrophages, and the level in PGE2-treated normal macrophages was similar to that in endometriotic macrophages without PGE2 treatment during the different time points of
culture. These results implied that reduced expression of annexin A2 in peritoneal macrophages is likely due to increased PGE2 exposure in the peritoneal fluid of women with endometriosis, indicating that annexin A2 might be involved in PGE2-induced suppression of phagocytosis.
PGE2-induced suppression of annexin A2 expression was mediated by the EP2 and EP4 receptor signaling pathway
We next tried to address the mechanism of PGE2-induced inhibition of annexin A2 expression (Figure 13). Macrophages isolated from normal and endometriotic women were treated with vehicle, PGE2 (1 or 10 μM), or selective receptor agonists for EP1 (ONO-D1-004, 10 μM), EP2 (ONO-AE1-259-01, 10 μM), EP3 (ONO-AE-248, 10 μM), or EP4 (ONO-AE1-329, 10 μM) for 12 and 24 hr.
Annexin A2 mRNA expression detected by real-time quantitative RT-PCR. Annexin A2 mRNA was markedly decreased in PGE2-treated macrophages from normal and endometriotic women, and treatment with the EP2 and EP4 receptor agonists found the similar down-regulation. It showed that decreased PGE2-induced annexin A2 expression was mediated by the EP2 and EP4 receptors.
DISCUSSIONS
Endometriosis is the most severe gynaecological disease, apart from malignant neoplasia, that affects >10% of women of reproductive age. Although Sampson's retrograde hypothesis provides some theoretical explanation of its aetiology, immune dysfunction has been thought to play important roles in endometriosis development.
Nevertheless, systematic evaluation of the phagocytic capability of macrophages in normal and endometriotic samples has never been done, neither has the underlying mechanism been characterized. In this study, we showed that peritoneal macrophages in women with endometriosis had reduced phagocytic ability. In-depth evaluation of
scavenger receptors revealed that the expression of CD36 and annexin II in peritoneal macrophages of women with endometriosis was significantly reduced.
Down-regulation of CD36 and annexin II led to the impairment of phagocytic ability of macrophages, while forced expression of CD36 in endometriotic macrophages restored their phagocytic ability. Our findings help to explain the long-standing notion that the peritoneal macrophages of endometriosis patients are less active in phagocytosis compared to that of normal patients and provide the molecular basis of the immune dysfunction that occurs during the development of endometriosis.
Endometriosis is a chronic inflammation that recruits many immune cells, especially macrophages, to the peritoneum.
Our previous study demonstrated that the number of macrophages with higher inflammatory agent-producing ability was increased in the peritoneal fluid of patients with endometriosis. Infiltrated macrophages supposedly function as scavengers, which help to remove peritoneal debris. However, due to some unknown mechanism, peritoneal macrophages may fail to complete their job, which allows the implantation and growth of endometrial tissues in the peritoneal cavity.
Thus, dysfunction of peritoneal macrophages is a severe impairment of the defence system and may be an important factor leading to the development of endometriosis. Unfortunately, due to the extreme difficulty of collecting peritoneal macrophages from patients, little progress was made in elucidating the underlying mechanisms of immune dysfunction during the development of endometriosis for many decades.
In this study, we report that the phagocytic ability of endometriotic peritoneal macrophages was impaired. In addition, peritoneal macrophages isolated from patients with endometriosis also demonstrated lack of capacity to be activated when cultured in a Petri dish. It is well known that a unique property of macrophages is that they will be activated simply by attaching to a solid surface. Thus, culturing macrophages in Petri dish is the simplest way to evaluate their
function. Indeed, the phagocytic ability of normal macrophages increased with increasing incubation time. In contrast, the phagocytic ability of endometriotic macrophages was not increased, suggesting that an intrinsic property might be impaired.
After systematic evaluation, we found that mRNA and protein of CD36 and annexin II were decreased in endometriotic macrophages compared to those in normal macrophages. These data imply that CD36 and annexin II may be a critical factor leading to the impairment of phagocytic ability.
In this study, we demonstrated that the level of CD36 and annexin II is directly associated with endometriosis. Expression of CD36 and annexin II was markedly reduced in peritoneal macrophages derived from endometriosis patients, which showed a significant impairment of phagocytic ability.
By using neutralizing antibody and siRNA knock-down approaches, we demonstrated that CD36 is indeed the critical factor that controls the phagocytic ability of peritoneal macrophages. Normal macrophages can be converted to endometriotic-like macrophages by simply inhibiting CD36 expression or blocking the function of CD36. On the other hand, ectopic expression of CD36 was sufficient to restore the phagocytic ability of endometriotic macrophages. These data demonstrate that CD36 and annexin II are necessary and sufficient for the phagocytic capacity of peritoneal macrophages and further suggest that CD36 and annexin II may be an important factor in the pathogenesis of endometriosis.
In our present study, we demonstrated that PGE2 is a potent locally acting factor that inhibits the expression of CD36 and annexin II in peritoneal macrophages. Administration of PGE2-inhibited CD36 expression in both primary-culture peritoneal macrophages and TPA-induced monocytic U937cells, indicating that this effect is likely to be a common phenomenon. More importantly, we demonstrated that ectopic expression of CD36 in peritoneal macrophages derived from women with endometriosis not only increased basal phagocytic activity but also restored PGE2-inhibited phagocytic ability. The
signaling that occurs downstream of PGE2 is very complex, as actions of PGE2 can be mediated by one or more of the four EP receptors. In this study, we demonstrated that inhibition of CD36 and annexin II expression by PGE2 was mediated by the EP2 receptor.
PGE2, via binding to the EP2 receptor, can activate protein kinase A and the mitogen-activated protein kinase pathway.44 Indeed, administration of the protein kinase A inhibitor, H89, or mitogen-activated protein kinase kinase inhibitor, U0126, significantly
reversed PGE2-induced CD36 down-regulation. In contrast, treatment of
macrophages with an EP2 receptor agonist was sufficient to inhibit CD36 expression.
Suppression of CD36 by PGE2 probably occurs at the transcriptional level because our preliminary data revealed that PGE2 can inhibit CD36 promoter activity (Chuang, unpublished data). We are currently trying to elucidate the molecular mechanism responsible for inhibition of CD36 and annexin II gene expression by PGE2.
Nevertheless, our current study shows for the first time that EP2 receptor- dependent PGE2 signaling is an important factor for impairing macrophage function in endometriosis.
Taking together our current and previous studies, it is clear that the functions of peritoneal macrophages in endometriotic patients were markedly impaired. Previously, we showed that the expression and enzyme activity of MMP-9 was inhibited in endometriotic macrophages. In this study, we further demonstrated that levels of the scavenger receptor CD36 and annexin II were also decreased in the peritoneal macrophages of women with endometriosis. Reduced expression of MMP-9, annexin II and CD36 indicates loss of both the first- and second-line defence mechanisms of macrophages. This may provide a reasonable explanation for macrophage dysfunction during the development and/or progression of endometriosis, although whether this represents the cause or effect of endometriosis can not be addressed by the current study. In addition, since the reduction of MMP-9, annexin II and CD36 expression is not due to a systemic defect of the immune system, it is particularly interesting to
identify the local factor (or factors) that suppresses the function of peritoneal macrophages. Our current findings provide novel information to point out a new avenue for studying the aetiology of endometriosis by directly targeting the phagocytic ability of macrophages. Further investigation on identifying the factors inhibiting annexin II and CD36 expression during the early stage of endometriosis development should be able to answer these questions and may shed light on developing new therapeutic regimens.
四、計畫成果自評
本研究內容是與原計畫相符程度及達成預 期目標情況類似,目前國內尚無很多針對子 宮內膜異位症患者的腹膜腔內巨噬球免疫 相關之研究分析,這項研究目的除了腹膜腔 內巨噬球與子宮內膜異位症的研究外,我們 希望了解這些免疫變化應該是局部而非全 身的相關性,試行找出適當的預後因子加以 分析,減少往後後遺症發生的機會。我們所 得成果於各研討會發表,例如生殖醫學年會 等,並將其成果寫成論文投稿於在學術學會 以及期刊發表,目前部分成果已經刊登於 Journal of Pathology 以及 The American Journal of Pathology, 另外準備投稿 Human Reproduction,希望對於臨床醫學上的診察 及治療具參考價值。
五、圖表
Figure 1. Reduced phagocytic ability in peritoneal macrophages isolated from patients with endometriosis. (A, B) Representative figure of phagocytic ability of peritoneal macrophages isolated from normal and
endometriotic patients measured by flow cytometry at 24 or 96 h after plating on a Petri dish. M0, no beads ingested; M1, at least one bead ingested. (C, D) Mean phagocytic ability of normal and endometriotic macrophages at 24 and 96 h after attachment. Data are means ± SD of six and four independent experiments, using different batches of macrophages from normal women and women with endometriosis, respectively. (E) Representative figure of phagocytic ability of peritoneal macrophages isolated from normal and endometriotic patients measured by flow cytometry at 96 h after plating, using FITC-conjugated annexin V-stained apoptotic Jurkat T cells as substrates. (F) Means ± SD of four independent experiments using different batches of
macrophages isolated from normal women and women with endometriosis, respectively. Significant difference at p < 0.05 by Student’s t-test
Figure 2. Expression of CD36 was downregulated in peritoneal macrophages isolated from women with endometriosis. (A) Expression of mRNA encoding several scavenger receptors in macrophages freshly isolated from normal and endometriotic patients. The experiment was repeated at least five times with peritoneal
macrophages isolated from different individuals and the results were similar. (B, C) Expression of CD36 (B) and SR-BI (C) mRNA in freshly isolated normal and endometriotic macrophages quantified by real time RT–PCR.
Amounts of CD36 and SR-BI transcripts were normalized to those of 18S ribosomal RNA due to subject variations. Data are means ± SD of 11 and 10 independent experiments, using different batches of macrophages from normal women and women with endometriosis, respectively. (D) Representative figure shows membrane
CD36 on freshly isolated normal and endometriotic macrophages detected by flow cytometry. (E) Means and standard deviation (SD) of three and five independent experiments, using different batches of macrophages obtained from normal women and women with endometriosis, respectively. Significant difference at p < 0.05 by Student’s t-test.
Figure 3. Reduced expression of CD36 in endometriotic peritoneal macrophages was not a systemic defect of the immune system. (A) Representative picture of RT–PCR results showed levels of CD36 mRNA, SR-BI mRNA and 18S ribosomal RNA in macrophages derived from peripheral blood (PB) and peritoneal fluid (PF) from normal (N) and endometriotic (E) women. NC, negative control without the addition of reverse
transcriptase. (B) Mean and SD of CD36 transcripts in macrophages derived from peripheral blood (PB) and peritoneal fluid (PF) of women without (N) or with (En) endometriosis. Significant difference at p < 0.05
Figure 4. CD36 is the functional scavenger receptor to maintain phagocytic ability. (A) Macrophages were treated with anti-CD36 antibody or mouse IgA (Con) as described and fluorescence intensities were determined.
M0, no beads ingested; M1, at least one bead ingested; %P, percentage of cells that ingested at least one bead.
(B) Means and SD of M1 obtained from four experiments using different batches of cells. (C) Representative
confocal microscopic images showed phagocytic macrophages isolated from normal and endometriotic patients.
Macrophages were treated with mouse IgA (Con) or CD36 antibody and then incubated with FITC-conjugated beads for 1 h. (D) Means and SD from five normal (Nor) and four endometriotic (Endo) individuals. At least 10 cells per individual were counted. (E) Macrophages were treated with mouse IgA (isotype control), anti-CD36 antibody (specific antibody), or anti-CD4 antibody (irrelevant antibody) as described. FITC-conjugated annexin V-stained apoptotic Jurkat T cells were used as substrates for phagocytosis. M0, no Jurkat cells ingested; M1, Jurkat T cells ingested. (F) Means and SD of percentage of phagocytic cells (upper panel) and phagocytic ability (lower panel) from four experiments, using different batches of normal peritoneal macrophages. *Significant difference at p < 0.05.
Figure 5. Inhibition of CD36 in normal macrophages blocks phagocytosis. (A) A representative RT-PCR picture shows the efficiency and specificity of siRNA. This experiment was repeated four times and the results were similar. (B) Representative confocal microscopic images showed macrophages transfected with scramble or siRNA against CD36. CD36 was stained as described in Materials and methods. N, nucleus. (C) Left panel, representative flow cytometric figure showing the fluorescence intensity of macrophages transfected with scramble or siRNA against CD36. Right panel, means and SD of three independent experiments using different batches of macrophages obtained from normal women. (D) Macrophages were transfected with scramble or siRNA against CD36 for 24 h and phagocytic ability was determined by flow cytometry. M0, no beads ingested;
M1, at least one bead ingested. (E) Means and SD of phagocytic ability (upper panel) and percentage of phagocytic cells (lower panel) from four normal macrophages with CD36 knockdown experiments. (F) Macrophages were transfected with scramble or siRNA against CD36 for 24 h and phagocytic ability was determined by flow cytometry, using FITC-conjugated annexin V-stained apoptotic Jurkat T cells as substrates.
M0, no Jurkat cells ingested; M1, Jurkat T cells ingested. (G) Means and SD of phagocytic ability (left panel) and percentage of phagocytic cells (right panel) from four experiments using different batches of normal peritoneal macrophages. *Significant difference at p < 0.05
Figure 6. Ectopic expression of CD36 restores phagocytic ability of endometriotic macrophages. (A) Macrophages were transfected with vector or human CD36 cDNA (hCD36) for 24 h. Phagocytic ability was determined by flow cytometry. M0, no beads ingested; M1, at least one bead ingested. (B, C) Means and SD of phagocytic ability and percentage of phagocytic cells from four endometriotic macrophages over-expressed with or without CD36. *Significant difference at p < 0.05.
Figure 7. PGE2 inhibits CD36 expression in normal and endometriotic macrophages. A: Macrophages from normal women were treated with 10 _M PGE2 or vehicle for 24 hr, and expression of scavenger receptors was determined by RT-PCR. This experiment was done four times using different batches of macrophages, and the results were similar. B: Normal (N) and endometriotic (E) macrophages were cultured for different periods as
indicated in the absence (control) or presence of PGE2, and levels of CD36 were quantified by real-time quantitative RT-PCR. Data represent the mean and standard deviation (SD) of four independent experiments using different batches of cells and were analyzed by ANOVA with repeated measurement. C: U937 monocytic cells were treated with 10 nM TPA for various periods as indicated, and CD36 mRNA expression was
quantified by real-time quantitative RT-PCR. Data represent the mean and SD of three independent experiments.
D, E: U937 cells were incubated with TPA for 48 hr and then treated with PGE2 (10 μM) for 24 hr or 48 hr.
Expression of CD36 mRNA (D) and protein (E) was determined (β-actin served as a loading control). The arrowhead indicates the glycosylated membrane form of CD36. Molecular size markers
(kDa) are shown to the left. *P <0.05, **P<0.01.
Figure 8. Inhibition of CD36 by PGE2 is mediated by the EP2 receptor signaling pathway. A: Macrophages isolated from normal women were treated with vehicle, PGE2 (1μM), or selective agonists for EP1
(ONO-D1-004, 10μM), EP2 (ONO-AE1-259-01, 10μM), EP3 (ONO-AE-248, 10μM), or EP4 (ONO-AE1-329, 10μM) for 12 hr. CD36 mRNA expression was detected by real-time quantitative RT-PCR. B: Macrophages isolated from normal women were treated with PGE2 (1 or 10 μM) or PGE2 plus inhibitors as indicated for 12 hr, and CD36 mRNA expression was determined by real-time quantitative RT-PCR. C: Macrophages isolated from normal women were treated with vehicle (Con), PGE2, or PGE2_AH6809 (10μM) for 96 hr, and CD36 expression was assessed by staining with anti-CD36 antibody or isotype-matched FITC-IgA (isotype control) followed by quantification via flow cytometry. Representative flow cytometric data are shown in the left panel.
The right panel shows the mean and SD of four independent experiments using different batches of peritoneal macrophages. D: Representative confocal images show CD36 staining in macrophages isolated from normal women treated with vehicle (Con), PGE2, or PGE2_AH6809 as described in (C). This experiment was done four times using different batches of cells. *P < 0.05, **P < 0.01, ***P < 0.001.
Figure 9. PGE2 inhibits phagocytosis via the EP2 receptor–dependent signaling pathway. A: Macrophages isolated from normal women were treated with vehicle (Con), anti-CD36, PGE2 (1 μM), or PGE2 plus EP2 antagonist (AH6809, 10 μM) for 24 hr, and fluorescence intensity was determined by flow cytometry. M0: no bead was ingested; M1: with at least one bead ingested. B: Mean and SD of M1 obtained from four experiments using different batches of cells. C: Representative confocal microscopic images of phagocytic macrophages isolated from normal patients. Macrophages were treated with vehicle (Con), PGE2, or PGE2_AH6809 and then incubated with FITC-conjugated beads for 1 h. D: Mean and SD of the number of beads phagocytosed by macrophages isolated from normal women after treatment with vehicle (Con), PGE2, or PGE2_AH6809. Four independent experiments using different batches of macrophages were conducted, and at least 20 cells per experiment were counted. *P< 0.05, **P<0.01, ***P < 0.001.
Figure 10. Ectopic expression of CD36 rescues PGE2-inhibited phagocytosis. A: Macrophages isolated from women with endometriosis were transfected with vector (as a control) or vector containing human CD36 cDNA (hCD36) for 24 hr. Cells were then treated without or with PGE2, and phagocytic ability was determined by flow cytometry. M0: no bead was ingested; M1: with at least one bead ingested. B: Mean and SD of phagocytic ability (upper panel) and percent of phagocytic cells (lower panel) from four independent experiments using
different batches of macrophages. *P <0.05.
Figure 11.
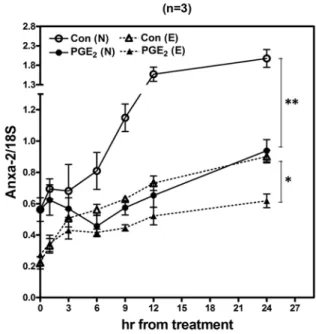
Figure 12. Normal (N) and endometriotic (E) macrophages were cultured for different periods as indicated in the absence (control) or presence of PGE2, and levels of Anxa-2 were quantified by real-time quantitative RT-PCR. Data represent the mean and standard deviation (SD) of three independent experiments using different batches of cells and were analyzed by ANOVA with repeated measurement.
Figure 13. Inhibition of Anxa-2 by PGE2 is mediated by the EP2 and EP4 receptor signaling pathway. Macrophages isolated from normal or endometriosis women were treated with vehicle, PGE2 (1 μM), or selective agonists for EP1 (ONO-D1-004, 10μM), EP2
(ONO-AE1-259-01, 10 μM), EP3 (ONO-AE-248, 10 μM), or EP4 (ONO-AE1-329, 10 μM) for 12 hr. Anxa-2 mRNA expression was detected by real-time quantitative RT-PCR.