Disruption of the peroxisomal citrate synthase CshA affects cell growth and
multicellular development in Dictyostelium discoideum
Ying-Chieh Huang, Yi-Hsing Chen, San-Ren Lo, Chia-I Liu, Cheng-Wei Wang, and
Wen-Tsan Chang*
Department of Biochemistry, National Cheng Kung University Medical College, Tainan 701, Taiwan, ROC.
* Corresponding author
Tel: +886-6-2353535 Ext. 5533 Fax: +886-6-2741694
E-mail: [email protected]
Running title: Peroxisomal citrate synthase in Dictyostelium
Summary
Nonmitochondrial citrate synthase catalyzes citrate synthesis in the glyoxylate cycle in
gluconeogenesis. Screening Dictyostelium discoideum mutants generated by insertional
mutagenesis isolated a poor-growing mutant that displayed aberrant developmental
morphology on bacterial lawns. Axenically grown mutants developed normally and
formed mature fruiting bodies on buffered agar. The affected locus encoded a novel
protein (CshA) that was homologous to glyoxysomal citrate synthase. cshA was
expressed maximally during vegetative growth and gradually decreased through
subsequent developmental stages. An in vitro citrate synthase assay revealed that cshA
disruption resulted in a 50 percent reduction of enzyme activity, implicating CshA as an
active citrate synthase. The amino terminus of CshA was found to have an atypical
mitochondrial targeting signal, instead containing a unique nonapeptide sequence
(RINILANHL) that was homologous to the conserved peroxisomal targeting signal 2
(PTS2). CshA protein was shown to be localized in the peroxisomes and the
RINILANHL sequence only efficiently targeted the peroxisomal GFP protein. The
growth defect of cshA- cells was associated with the impairment of phagocytosis and
fluid-phase endocytosis, independent from cytokinesis. Disrupted multicellular
development on bacterial lawns resulted from the abnormal susceptibility to the
results provide new insights into the function of peroxisomal citrate synthase in the cell
Introduction
Nonmitochondrial citrate synthase participates in the glyoxylate cycle, catalyzing the condensation of acetyl-coenzyme A (acetyl-CoA) with oxaloacetate to produce citrate. In many organisms, this metabolic pathway serves as a mechanism for converting fatty acids or acetate to carbohydrates. In contrast, vertebrate animals lack the glyoxylate cycle-specific enzymes and so cannot utilize lipids and acetate for glucose synthesis. The glyoxylate cycle can be carried out in the membrane-bound peroxisome or in the cytosol. In plants, the enzymes are sequestered in a specialized peroxisome called the glyoxysome (Olsen and Harada, 1995). To complete the glyoxylate cycle, glyoxysomes cooperate metabolically with mitochondria to convert succinate to oxaloacetate, which is catalyzed by the enzymes involved in the tricarboxylic acid (TCA) cycle. Thus, there is a continuous interchange of intermediates between these two organelles (Tolbert, 1981; van den Bosch et al., 1992).
Podospora anserine exhibits no visible vegetative defects but impairs meiosis progression
beyond the diffuse stage, a key stage of meiotic prophase (Ruprich-Robert et al., 2002). As a final example, an increase in citrate excretion from citrate synthase transgenic plants improves phosphorus uptake, which promotes the growth and development in phosphorus-limited soil (de la Fuente et al., 1997; Koyama et al., 1999; Koyama et al., 2000; Lopez-Bucio et al., 2000). These results clearly indicate that mitochondrial citrate synthases play a decisive role in the cell growth and development.
stabilized by a basal disc of stalk cells, supporting a mass of spores (Parent and Devreotes, 1996; Aubry and Firtel, 1999).
Peroxisomes are characterized by the presence of hydrogen peroxide (H2O2)-producing
oxidases and catalase, which catalyze the decomposition of H2O2. They are near ubiquitous in
eukaryotes. In plants and some microorganisms such as fungi and yeast, the enzymes of the glyoxylate cycle are sequestered in the peroxisomes or in related glyoxysomes. It has been reported that D. discoideum peroxisomes contain only urate oxidase and catalase, with other key enzymes of the glyoxylate cycle, including H2O2-producing oxidases as well as isocitrate
lyase and malate synthase, being absent (Parish, 1975). Peroxisomal citrate synthase catalyzes the first reaction of the glyoxylate cycle and what is generally assumed to be the rate determining step for the cycle. Despite this key position of the enzyme in cellular metabolism, very few studies have addressed the effects of its deficiency in cell growth and multicellular development.
particular, contains a peroxisomal targeting signal 2 (PTS2) located near the amino terminus. Moreover, we demonstrate that CshA is indeed a peroxisomal protein involved in the cellular biosynthesis of citrate, and that it is required primarily for cell growth and modulation of multicellular development. The growth defect of the cshA- cells is closely coupled to the inability of endocytosis and phagocytosis, and the aberrant developmental morphology is strongly linked to the abnormal susceptibility to the environmental conditions.
Results
Isolation of REMI-induced cell growth mutant WTC127
blot analysis confirmed that they were disruptants of the same locus (data not shown). Both the original and recreated mutants were used in subsequent experiments.
Analysis of the cshA gene
To identify the disrupted gene in the WTC127 mutant, we first sequenced the 3.4 kb flanking region of the rescued genomic DNA. This was then used to clone the full length of coding sequence using RT-PCR from vegetative phase total RNA. The cDNA obtained contained one long open reading frame of 1476 bp that encoded a predicted protein of 492 amino acids with a calculated molecular mass of 55.7 kDa. This protein contained a conserved citrate synthase domain (amino acids 81-449) with the three conserved residues (307H, 346H and 402D) (Figure
2A) that are critical for catalytic activity (Karpusas et al., 1990; Remington, 1992).
Alignment of the high scoring segments obtained from a BLAST search of the non-redundant NCBI database (http://www.ncbi.nlm.nih.gov/BLAST/), using the deduced amino acid sequence of the isolated cDNA coding region revealed that the putative product was homologous to the citrate synthase. The sequence is highly conserved, with amino acid identities of 56 percent to the citrate synthase of Cucurbita maxima (P49299) (Kato et al., 1995), and 57 percent to the putative citrate synthase protein of Arabidopsis thaliana (NP_181807) (data not shown).
search the non-redundant NCBI database, as well as the D. discoideum genomic and cDNA databases (All Dictyostelium BLAST Search; http://dicty.sdsc.edu/). The search indicated that
D. discoideum contained another highly related citrate synthase gltA gene (AAO50962)
(Glockner et al., 2002).
To further confirm that the mutant phenotype was due to loss of the protein encoded by
cshA gene, we reexpressed the complete coding sequence fused in-frame with glutathione-S
transferase (GST) gene under the control of actin 15 promoter and actin 8 terminator in WTC127 cells. The results showed that only expressed CshA-GST constructs restored the ability of WTC127 cells to grow relatively normal (data not shown). The re-expression of the CshA-GST fusion protein was verified by Western blot analysis using anti-GST antibody (Figure 2C).
The expression pattern of cshA during development was determined by extracting RNA at various times during development and hybridizing the RNA with full-length cshA cDNA.
cshA encoded a single transcript of 1.8 kb, which was expressed maximally at the vegetative
growth with expression subsequently decreasing during the following developmental stages (Figure 3). No transcript was found in the WTC127 mutant (data not shown).
Table 1, cshA- cells exhibited about 50 percent of the citrate synthase activity of wild-type cells.
Compartmental localization of the CshA protein
The deduced amino acid sequence of CshA lacks a typical mitochondrial targeting signal at the amino terminus (Douglas et al., 1986; Neupert, 1997) and a typical peroxisomal targeting signal 1 (PTS1) at the carboxyl terminus (Subramani et al., 2000). However, the predicted amino terminus did contain a unique nonapeptide sequence, RINILANHL (amino acids 6-14) (Figure 2A), that was homologous to sequences of microbody enzymes (Figure 2B) such as glyoxysomal citrate synthase of C. maxima (Kato et al., 1995; Kato et al., 1996; Kato et al., 1999) and the putative citrate synthase protein of A. thaliana (NP_181807). The terminus could possibly serve as a conserved peroxisomal targeting signal 2 (PTS2) for the transport of polypeptides into the peroxisomal matrix (Subramani et al., 2000).
4B).
To further elucidate the specific topogenic information of this putative PTS2 sequence, different site-directed mutations were made and analyzed for their effect on peroxisomal targeting. Substitutions of 6RI into DD or 13HL into DE (Figure 4A) were associated with the
subsequent mislocalization of the mutated CshA-GFP fusion proteins into the cytosol (Figure 4B). In addition, GFP protein tagged with this PTS2 signal localized in the peroxisome of D.
discoideum. (Figures 4A and 4B).
Effects in cell growth
To analyze in detail the phenotypes of cshA null mutation, we examined the growth rate of
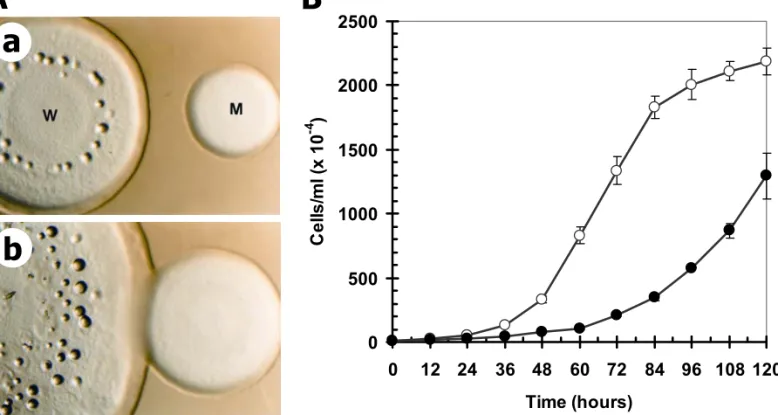
cshA- cells on SM bacterial lawn and in axenic suspension culture. Klebsiella aerogenes is widely used for the efficient cultivation of D. discoideum amoebae on the nutrient SM-agar (Williams and Newell, 1976). When grown on the K. aerogenes lawn, cshA- cells showed a greatly reduced growth rate and formed a thin growth zone. The rate of plaque expansion in the cshA- cells was also slower than the rate of wild-type cells (Figure 5A). This growth defect was compared by measuring the plaque diameter after 7 days of plating. The plaque diameter of cshA- cells was only about 40 percent that of the wild-type cells (data not shown).
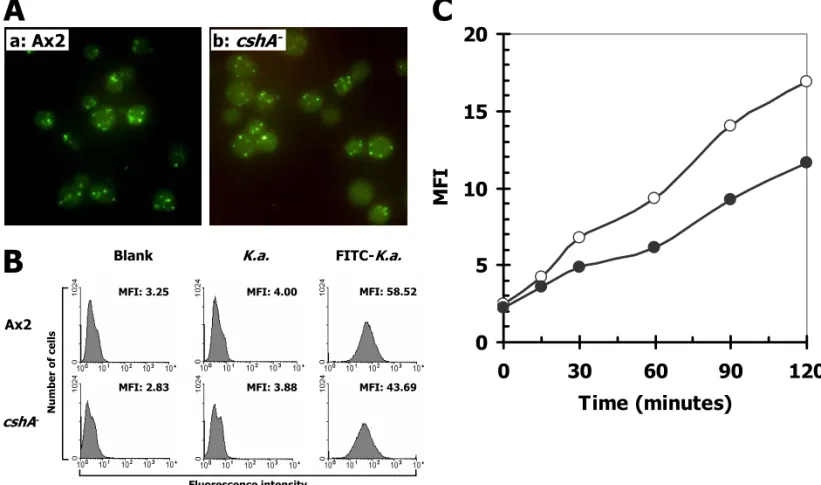
phagocytose live FITC-labeled K. aerogenes bacteria was measured. As shown in Figure 6A, the uptake of K. aerogenes by cshA- and wild-type cells was similar. However, under more sensitive flow cytometric analysis the rate of FITC-labeled K. aerogenes uptake in cshA- cells was observed to be 75 percent that of the wild-type rate (Figure 6B).
When cultured in axenic suspension, cshA- cells grew very slowly in comparison with the wild-type Ax2 cells (Figure 5B). The wild type cells duplicated with a doubling time of approximately 8 hours whereas the cshA- cells failed to duplicate markedly even after 84 hours of cultivation. When viewed under an inverted microscope, cshA- cells kept in shaking axenic culture for three days did not exhibit the huge increase in cell size (data not shown). Staining with 4,6-diamidino-2-phenylindole (DAPI) showed that most of the cshA- cells contained a single nucleus (data not shown).
Effects in multicellular development
When grown on a bacterial lawn, cshA- plaques displayed a relatively small and thin clearing zone (Figures 1 and 5A). In the central area of the plaque, there were a small number of aberrant multicellular structures. Mature fruiting bodies did not form (Figure 1). Interestingly, when cshA- cells were developed axenically on non-nutrient buffered agar, they completed
multicellular development within 24 hours, forming mature spore-containing fruiting bodies (data not shown).
To investigate the differences of multicellular development on the bacterial SM-agar and on the non-nutrient KK2-agar, we plated the wild-type Ax2 and cshA- WTC127 cells on the
non-nutrient KK2-agar in the presence of K. aerogenes bacteria. Under this condition, the
wild-type cells formed mature normal fruiting bodies by 48 hours, while cshA- cells formed aberrant developmental structures after 72 hours of incubation (Figure 7A).
We further examined the development of wild-type and cshA- cells in experiments in which various ratios of the cell types were established on non-nutrient KK2-agar with the K. aerogenes bacteria. The chimeric aggregates were able to form mature fruiting bodies in the
presence of as little as 10 percent of the wild-type cells (Figure 7B), consistent with the suggestion that wild-type cells rescued the developmental defect of cshA- cells.
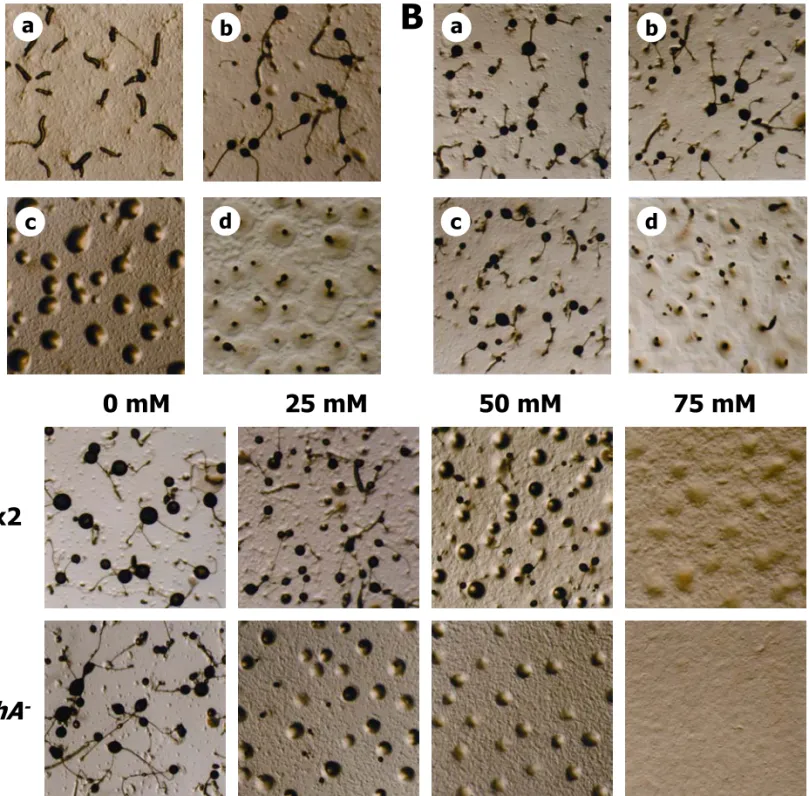
large amounts as a result of protein catabolism. Previous studies have shown that ammonia inhibits aggregation and culmination during D. discoideum development (Davies et al., 1993; Gee et al., 1994). To test whether developmental processes of the cshA- cells were hypersensitive to ammonia, wild-type and cshA- cells were allowed to develop on non-nutrient KK2 agars in the presence of various concentrations of ammonia. The
aggregation of both Ax2 and cshA- cells was inhibited by greater than 75 mM of ammonia (Figure 7C), however, ammonia delayed the onset of cshA- cellular aggregation by six to eight hours relative to that of Ax2 cells (data not shown). Moreover, wild-type cells displayed an ammonia concentration-related decrease in the number of fruiting bodies produced. Low concentrations of ammonia did not prevent wild-type fruiting body formation, but induced a progressive decrease in fruiting body size, while the formation of fruiting bodies was halted in similarly-treated cshA- cells.
Discussion
mononucleated or binucleated when grown under either stationary or suspension conditions. Quantitative analysis of the internalization of FITC-labeled living bacteria and dextran indicated that disruption of the cshA impairs the internalization of particles and fluid. Previous studies have clearly indicated that mutant strains with impairment in fluid-phase endocytosis are correlated with an inability to proliferate in axenic culture (Bacon et al., 1994), while impairment of the phagocytosis system affects cell growth on the bacterial lawn (Cohen et al., 1994). In accordance with these studies, the impaired growth phenotypes of the cshA- cells in the axenic culture and on the bacterial lawn are consistent with the observed defects in endocytosis and phagocytosis.
The cshA- cells displayed aberrant developmental morphology on bacterial lawns, but developed normally and formed mature fruiting bodies on buffered agar. With the addition of
K. aerogenes bacteria on the buffered agar, the mutant cells develop aberrantly as on a
bacterial lawn, but this abnormal development can be overcome by the coexistence of wild-type cells in the chimeric aggregates. These results indicate that the presence of K.
aerogenes interferes with the normal multicellular development of cshA- cells, and that this
assays revealed that cshA- cells possessed approximately 50 percent of the enzyme activity of wild-type cells, indicating that CshA is a functional citrate synthase that takes part in the synthesis of citrate, and that D. discoideum contains additional citrate synthases.
Since citrate synthase occupies such a key position within energy metabolism and is encoded by a single gene in many organisms, disruption of the gene may induce serious damage to the cell survival. We have isolated two distinct forms of citrate synthase, CshA and GltA (S.-R. Lo, unpublished), suggesting that they may have some functional redundancy.
cshA is expressed maximally at the vegetative growth and decreased gradually during the
following developmental stages, suggesting that CshA is required mainly for the vegetative phase of cell growth and the early aggregation stage of multicellular development. Analysis of the novel gltA expression using RT-PCR revealed that the transcript is present throughout the entire development, indicating that gltA gene is constitutively active in D. discoideum (Y.-H. Chen, unpublished). Previous studies have showed that citrate synthase specific activity is present throughout the development (Porter and Wright, 1977).
for directing the protein into peroxisomes. Mutations in this conserved nonapeptide residues cause mislocalization of the CshA-GFP fusion protein throughout the cytosol. In addition, the present results further show that the N-terminal RINILANHL sequence is sufficient to target a leaderless version of GFP to peroxisomes. Moreover, both the CmGCS and AtPCS not only show high homology with CshA but also contain the homology PTS2 sequences near their amino termini (Kato et al., 1995; Kato et al., 1996), suggesting that these citrate synthases may participate in the similar biological function. These results clearly indicate that CshA is localized into the peroxisomes via the conserved PTS2 at its amino terminus.
CshA is the second peroxisomal protein from D. discoideum for which sequence data have been characterized. The multifunctional enzyme MFE1 contains the SKL tripeptide at the carboxyl terminus instead of the RINILANHL nonapeptide located near the amino terminus (Matsuoka et al., 2003). Therefore, it would appear that the process of targeting proteins to peroxisomes is complex, and that more than one type of targeting signals exists. It is perhaps surprising that the novel citrate synthase GltA has neither a typical mitochondrial targeting sequence nor the conserved peroxisomal targeting signals including PTS1 and PTS2. It is conceivable that GltA possesses a cryptic targeting signal or is a cytosolic protein. Based on subcellular localization of the GltA-GFP fusion protein (S.-R. Lo, unpublished), we favor a cytoplasmic location.
matrix. This loss of peroxisomal content was initially taken as evidence for the permeability of the peroxisomal membrane in vivo, but is now known to be an artifact of the isolation (Tabak et al., 1999). Previous studies have shown that isolated peroxisomes from D.
discoideum contain only urate oxidase and catalase; no other H2O2-producing oxidases and
Experimental procedures
Growth and development of Dictyostelium
Dictyostelium discoideum wild-type strain Ax2 was maintained in association with Klebsiella aerogenes on SM-agar plates (Sussman, 1987) and grown in axenic medium (Klein et al.,
1989) at 22oC in the dark. Blasticidin S-resistant (Bsrr) transformants were selected and
grown in axenic medium with 5 µg/ml of blasticidin S (Sutoh, 1993). Neomycin-resistant (Neor) transformants selected in axenic medium supplemented with concentrations of
geneticin (G418) that gradually increased from 5–100 µg/ml, were grown in axenic medium containing 20 µg/ml G418 (Dingermann et al., 1989). For development, exponentially growing cells (3-8 × 106 cells/ml) were harvested and washed three times in KK
2 (16.5 mM
KH2PO4, 3.8 mM K2HPO4, pH6.2), resuspended at 5 × 107 cells/ml and developed on KK2
buffered agar.
REMI mutagenesis and re-disruption
The genomic DNA flanking the integrated vector was isolated from strain WTC127 by digesting the DNA with BclI, re-ligating and selecting for ampicillin-resistant transformants in
Escherichia coli. The resulting plasmid pWTC127/BclI contained 3.4 kb of flanking genomic
DNA. To recreate the mutant phenotype by homologous recombination, the rescued plasmid was linearized with BclI and electroporated into the parental Ax2 cells. Transformants were selected in axenic medium with blasticidin S and screened for transformants with the same phenotype as the original mutant.
Nucleotide and amino acid sequence analysis
Cloned DNAs were sequenced by using the Applied Biosystems BigDye terminator cycle sequence ready reaction mix with template and pertinent primers, and analyzed on a Perkin Elmer-Cetus ABI Prism 377 DNA Sequencer. Nucleotide and amino acid sequences were analyzed using the “BLAST” (http://www.ncbi.nlm.nih.gov/BLAST/), “All Dictyostelium BLAST Search” (http://dicty.sdsc.edu/), or “ExPASy Molecular Biology Server” (http://tw.expasy.org/).
Cellular location of CshA protein
plasmid (Chang et al., 1998). The resulting construct contained the cshA coding sequence fused in-frame with DNA for the green fluorescent protein (GFP-C3) (Crameri et al., 1996) under the control of the actin 15 promoter and actin 8 terminator, and also conferred resistance to G418. The constructs were used for transformation of wild-type Ax2 and WTC127 using electroporation and selection by resistance to G418.
Mutagenesis of the 6RI into DD or 13HL into DE in the CshA protein
The 6RI or 13HL residues in CshA protein were changed to 6DD or 13DE by PCR from the
full-length cshA cDNA with appropriate primers and cloned into pAct15-GFP plasmid. The
primer for 6RI was CshA-PTS2(R6D,I7D):
5’-GAAGATCTATGGCACATATTGATGATGATAATATACTTGCC-3’ and for 13HL was
CshA-PTS2(H13D,L14E):
Southern blot analysis
Genomic DNAs were purified by phenol/chloroform extraction and isolated by ethanol precipitation, then digested with various restriction enzymes, and analyzed by Southern blot as described previously (Chang et al., 1995), using radio-labeled DNA fragment of cshA gene as a probe.
Northern blot analysis
Total cellular RNAs were prepared using Catrimox-14TM Surfactant (Iowa Biotech Corp.,
Iowa City) according to Dahle and Macfarlane (1993) and analyzed on Northern blots as described by Chang et al. (1996), using radio-labeled DNA fragment of cshA gene as a probe.
Complementation by transformation
and selection by resistance to G418.
Western blot analysis
Exponentially growing cells were harvested and washed three times with KK2 buffer by
centrifugation, lysed in SDS-PAGE sample buffer and boiled for 5 minutes, sized on a 10% SDS-PAGE gel, blotted to Immobilon-P membrane (Millipore Corporation, Bedford, MA), and probed with a commercial goat antibody specific to GST (Amersham Pharmacia Biotech., Piscataway, NJ).
Growth rate on bacterial lawns
The growth rate of Dictyostelium amoebae on bacterial lawns was determined as described by Chen and Katz (2000). Axenically growing cells (3-8 × 106 cells/ml) were plated with K.
aerogenes on SM-agar plates (Sussman, 1987). The plates were then incubated at 22oC for 7
days. The diameters of emerged clonal plaques were measured with a vernier caliper.
Phagocytosis assay
fluorescein isothiocyanate (FITC; Sigma Chemical Co., St. Louis, MO). The amount of internalized FITC-labeled K. aerogenes was analyzed by flow cytometry. The flow cytometry measurements were performed at a FACSCaliburTM flow cytometer (Beckton Dickinson,
Franklin Lakes, NJ).
Endocytosis assay
Quantitative assay of fluid-phase uptake was determined by a modification of a previous method (Hacker et al., 1997). FITC-dextran was used as the fluid-phase marker. The amount of internalized FITC-dextran was analyzed using flow cytometry with an excitation wavelength of 470 nm and an emission wavelength of 520 nm.
Analysis of cytokinesis
10 minutes. The suspension was then transferred to a cover slip, incubated at 22oC for one
hour, and fixed for analysis.
Citrate synthase activity assay
Citrate synthase activity in the cell extracts was measured according to the method of Srere (1969) as described previously (Kim et al., 1986), in which the free acetyl-CoA formed during the reaction reacts with 5’,5’-dithiobis-(2-nitrobenzoate) (DTNB) to form a compound that absorbs strongly at 412 nm. Protein concentrations were determined spectrophotometrically at 562 nm using the bicinchoninic acid (BCA) protein assay (Pierce Biotechnology) as described by Smith et al. (1985), with bovine serum albumin as the standard.
Accession Number
The DDBJ/EMBL/GenBank accession number for the cshA gene is AY124377.
Acknowledgements
References
Aubry, L., and Firtel, R.A. (1999) Integration of signaling networks that regulate
Dictyostelium differentiation. Annu Rev Cell Dev Biol 15: 469-517.
Bacon, R.A., Cohen, C.J., Lewin, D.A., and Mellman, I. (1994) Dictyostelium discoideum mutants with temperature-sensitive defects in endocytosis. J Cell Biol 127: 387-399. Cardelli, J. (2001) Phagocytosis and macropinocytosis in Dictyostelium:
phosphoinositide-based processes, biochemically distinct. Traffic 2: 311-320.
Chang, W.-T., Gross, J.D., and Newell, P.C. (1995) Trapping developmental promoters in
Dictyostelium. Plasmid 34: 175-183.
Chang, W.-T., Newell, P.C., and Gross, J.D. (1996) Identification of the cell fate gene stalky in
Dictyostelium. Cell 87: 471-481.
Chang, W.-T., Thomason, P.A., Gross, J.D., and Newell, P.C. (1998) Evidence that the RdeA protein is a component of a multistep phosphorelay modulating rate of development in
Dictyostelium. EMBO J 17: 2809-2816.
Chen, C.F., and Katz, E.R. (2000) Mediation of cell-substratum adhesion by RasG in
Dictyostelium. J Cell Biochem 79: 139-149.
Cohen, C.J., Bacon, R., Clarke, M., Joiner, K., and Mellman, I. (1994) Dictyostelium
discoideum mutants with conditional defects in phagocytosis.J Cell Biol 126: 955-966.
protein by molecular evolution using DNA shuffling. Nature Biotechnol 14: 315-319. Dahle, C.E., and Macfarlane, D.E. (1993) Isolation of RNA from cells in culture using
catrimox-14 cationic surfactant. BioTechniques 15: 1102-1105.
Davies, L., Satre, M., Martin, J.B., and Gross, J.D. (1993) The target of ammonia action in
Dictyostelium. Cell 75: 321-732.
de la Fuente, J.M., Ramírez-Rodríguez, V., Cabrera-Ponce, J.L., and Herrera-Estrella, L. (1997) Aluminum tolerance in transgenic plants by alteration of citrate synthesis. Science
276: 1566-1568.
Dingermann, T., Reindl, N., Werner, H., Hildebrandt, M., Nellen, W., Harwood, A., Williams, J., and Nerke, K. (1989) Optimization and in situ detection of Escherichia coli beta-galactosidase gene expression in Dictyostelium discoideum. Gene 85: 353-362.
Douglas, M.G., McCammon, M., and Vassarotti, A. (1986) Targeting of proteins into mitochondria. Microbiol Rev 50: 166-178.
Gee K., Russell F., and Gross J.D. (1994) Ammonia hypersensitivity of slugger mutants of D.
discoideum. J Cell Sci 107: 701-708.
discoideum. Nature 418: 79-85.
Grossman, A.D. (1995) Genetic networks controlling the initiation of sporulation and the development of genetic competence in Bacillus subtilis. Annu Rev Genet 29: 477-508. Hacker, U., Albrecht, R., and Maniak, M. (1997) Fluid-phase uptake by macropinocytosis in
Dictyostelium. J Cell Sci 110: 105-112.
Ireton, K., Jin, S., Grossman, A.D., and Sonenshein, A.L. (1995) Krebs cycle function is required for activation of the Spo0A transcription factor in Bacillus subtilis. Proc Natl
Acad Sci USA 92: 2845-2849.
Karpusas, M., Branchaud, B., and Remington, S.J. (1990) Proposed mechanism for the condensation reaction of citrate synthase: 1.9-Å structure of the ternary complex with oxaloacetate and carboxymethyl coenzyme A. Biochemistry 29: 2213-2219.
Kato, A., Hayashi, M., Kondo, M., and Nishimura, M. (1996) Targeting and processing of a chimeric protein with the N-terminal presequence of the precursor to glyoxysomal citrate synthase. Plant Cell 8: 1601-1611.
Kato, A., Hayashi, M., Mori, H., and Nishimura, M. (1995) Molecular characterization of a glyoxysomal citrate synthase that is synthesized as a precursor of higher molecular mass in pumpkin. Plant Mol Biol 27: 377-390.
Physiol 40: 586-591.
Kim, K.S., Rosenkrantz, M.S., and Guarente, L. (1986) Saccharomyces cerevisiae contains two functional citrate synthase genes. Mol Cell Biol 6: 1936-1942.
Klein, G., Cotter, D.A., Martin, J.-B. and Satre, M. (1989) Vanadate, an inhibitor of growth, development and endocytosis in Dictyostelium discoideum amoebae. J Cell Sci 94: 127-134.
Koyama, H., Kawamura, A., Kihara, T., Hara, T., Takita, E., and Shibata, D. (2000) Overexpression of mitochondrial citrate synthase in Arabidopsis thaliana improved growth on a phosphorus-limited soil. Plant Cell Physiol 41: 1030-1037.
Koyama, H., Takita, E., Kawamura, A., Hara, T., and Shibata, D. (1999) Over expression of mitochondrial citrate synthase gene improves the growth of carrot cells in Al-phosphate medium. Plant Cell Physiol 40: 482-488.
Kuspa, A., and Loomis, W.F. (1992) Tagging developmental genes in Dictyostelium by restriction enzyme-mediated integration of plasmid DNA. Proc Natl Acad Sci USA 89: 8803-8807.
Lopez-Bucio, J., de La Vega, O.M., Guevara-Garcia, A., and Herrera-Estrella, L. (2000) Enhanced phosphorus uptake in transgenic tobacco plants that overproduce citrate. Nat
Biotechnol 18: 450-453.
Urushihara, H., Tanaka, Y., and Loomis, W.F. (2003) Changing patterns of gene expression in Dictyostelium prestalk cell subtypes recognized by in situ hybridization with genes from microarray analyses. Eukaryot Cell 2: 627-637.
Maniak, M. (2001) Fluid-phase uptake and transit in axenic Dictyostelium cells. Biochim
Biophys Acta 1525: 197-204.
Matsuoka, S., Saito, T., Kuwayama, H., Morita, N., Ochiai, H., and Maeda, M. (2003) MFE1, a member of the peroxisomal hydroxyacyl coenzyme A dehydrogenase family, affects fatty acid metabolism necessary for morphogenesis in Dictyostelium spp. Eukaryot Cell 2: 638-645.
Neupert, W. (1997) Protein import into mitochondria. Annu Rev Biochem 66: 863-917.
Olsen, L.J., and Harada, J.J. (1995) Peroxisomes and their assembly in higher plants. Annu
Rev Plant Physiol 46: 123-146.
Parent, C.A., and Devreotes, P.N. (1996) Molecular genetics of signal transduction in
Dictyostelium. Annu Rev Biochem 65: 411-40.
Parish, R.W. (1975) Mitochondria and peroxisomes from the cellular slime mould
Dictyostelium discoideum. Isolation techniques and urate oxidase association with
peroxisomes. Eur J Biochem 58: 523-531.
Remington, S.J. (1992) Structure and mechanism of citrate synthase. Curr Top Cell Regul 33: 209-229.
Rupper, A., and Cardelli, J. (2001) Regulation of phagocytosis and endo-phagosomal trafficking pathways in Dictyostelium discoideum. Biochim Biophys Acta 1525: 205-216. Ruprich-Robert, G., Zickler, D., Berteaux-Lecellier, V., Vélot, C., and Picard, M. (2002) Lack
of mitochondrial citrate synthase discloses a new meiotic checkpoint in a strict aerobe.
EMBO J 21: 6440-6451.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gartner, F.H., Provenzano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J., and Klenk, D.C. (1985) Measurement of protein using bicinchoninic acid. Anal Biochem 150: 76-85.
Srere, P.A. (1969) Citrate synthase. Methods Enzymol 13: 3-11.
Subramani, S., Koller, A. and Snyder, W.B. (2000) Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem 69: 399-418.
Sussman, M. (1987) Cultivation and synchronous morphogenesis of Dictyostelium under controlled experimental conditions. Methods Cell Biol 28: 9-29.
Sutoh, K. (1993) A transformation vector for Dictyostelium discoideum with a new selectable marker bsr. Plasmid 30: 150-154.
Tolbert, N.E. (1981) Metabolic pathways in peroxisomes and glyoxysomes. Annu Rev
Biochem 50: 133-157.
van den Bosch, H., Schutgens, R.B., Wanders, R.J. and Tager, J.M. (1992) Biochemistry of peroxisomes. Annu Rev Biochem 61: 157-197.
Watts, D.J., and Ashworth, J.M. (1970) Growth of myxameobae of the cellular slime mould Dictyostelium discoideum in axenic culture. Biochem J 119: 171-174.
Wiegand, G., and Remington, S.J. (1986) Citrate synthase: structure, control, and mechanism.
Annu Rev Biophys Chem 15: 97-117.
Figure legends
Fig. 1. Phenotype of the REMI-tagged WTC127 mutant. An overhead view of wild-type
Ax2 (A) and REMI-induced WTC127 mutant (B) growing and developing on SM-agar plates in association with Klebsiella aerogenes.
Fig. 2. Sequence analysis of the cshA gene. (A) Amino acid sequence of the cshA gene
product. The CshA protein is composed of 492 amino acid residues, which contains a conserved citrate synthase domain (italic letters) and possesses a putative peroxisomal targeting signal PTS2 (boxed) with the conserved amino acids (bold letters). The conserved residues identified by Karpusas et al., (1990) as essential for the catalytic activity of citrate synthases are marked with a triangle. (B) Alignment of the putative CshA PTS2 sequence with other PTS2 sequences from the citrate synthase GCS of Cucurbita maxima (P49299) (Kato et al., 1995) and the putative citrate synthase protein PCS of Arabidopsis thaliana (NP_181807). Consensus amino acids in the PTS2 sequences are indicated with bold letters. (C) Western blot analysis of the CshA-GST fusion protein expression. Cells of the indicated strains were probed with anti-GST antibody.
Fig. 3. Expression of the cshA gene during development. Wild-type Ax2 cells were allowed
probed with the 32P labelled full-length cshA cDNA. IG7 that was expressed at a constant level during development was used as a reference. Amoebae were aggregating by six hours, and formed mounds by nine hours, tipped aggregates by 12 hours, slugs by 16 hours, late culmination by 20 hours, and mature fruiting bodies by 24 hours.
Fig. 4. Subcellular localization of the CshA protein. (A) Constructs and (B) fluorescence
images of the green fluorescent protein (GFP) (a) act15-gfp (b)-(e) CshA-GFP fusion proteins expressed under the control of the actin 15 promoter and actin 8 terminator. (b) wild-type CshA (act15-cshA-gfp); (c) site-directed substitution mutants CshA(6DD)
(act15-cshA(6DD)-gfp); (d) CshA(13DE) (act15-cshA(13DE)-gfp); (e)
PTS2(RINILANHL)-GFP fusion protein (act15-PTS2-gfp).
Fig. 5. Growth kinetics of wild-type and cshA- cells on lawns of K. aerogenes and in axenic
suspension cultures. (A) Growth on the bacterial lawn. The plaques of wild type Ax2 (W) and
Fig. 6. Phagocytosis of bacteria and fluid-phase endocytosis by wild-type and cshA- cells.
(A) Fluorescence images and (B) histogram plots of the phagocytosis of FITC-labeled K.
aerogenes by wild-type Ax2 and cshA- WTC127 mutant. Cells were incubated with
FITC-labeled bacteria on buffered agar at 22oC for 40 min. After washing, live cells were
photographed using a fluorescence microscope and analyzed using a fluorescence-activated cell sorter. (C) Time course of the endocytosis of FITC-dextran by wild-type Ax2 (open circles) and the cshA- WTC127 mutant (filled circles). Cells were incubated with 2.0 mg/ml FITC-dextran at 22oC for various time periods. After washing, live cells were analyzed using
a fluorescence-activated cell sorter.
Fig. 7. Effect of the cshA- mutation on multicellular development. (A) Developmental
morphologies of wild-type Ax2 (a, b) and cshA- WTC127 mutant (c, d) on KK
2 buffered agars
in the presence of K. aerogenes, photographed after 36 (a), 48 (b, c), 72 hours (d) of development. (B) Developmental morphologies of chimeric aggregates of wild-type Ax2 and
cshA- WTC127 mutant mixed in various ratios on KK
2 buffered agars in the presence of K.
aerogenes, photographed after 72 hours of development. a, 100% Ax2; b, 25% Ax2 and 75%
Tables
Table 1. Citrate synthase activity in crude extracts of the wild-type Ax2 and cshA- mutant
A
Figure 3
cshA
IG7
wild-type
Figure 4
B
a b c d e b: act15-cshA-gfp c: act15-cshA(6DD)-gfp d: act15-cshA(13DE)-gfp e: act15-PTS2-gfp a: act15-gfp gfp cshA cshA(6DD) cshA(13DE) Actin 15promoter terminatorActin 8
RINILANHL
DDNILANHL
RINILANDE
RINILANHL
Figure 6
A
a: Ax2 b: cshA -MFI: 2.83 MFI: 3.25 Nu mbe r of cells Ax2 Blank MFI: 3.88 MFI: 43.69 cshA-K.a. FITC-K.a.
Table 1
Table 1. Citrate synthase activity in crude extracts
of the wild-type Ax2 and cshA-mutant WTC127 Strain Citrate synthase activity*nM·mg-1·protein-1·min-1
Ax2 WTC127
228 ± 8 111 ± 6