IINNTTRROODDUUCCTTIIOONN
Dietary fatty acids are known to affect plasma cholesterol concentrations and to play important roles in the development of atherosclerotic disease [1]. One of the major
factors associated with atherosclerotic disease is diet-induced elevations of plasma LDL concentrations. Dietary cholesterol causes an increase in LDL-C, but this effect may be modified by the type and amount of fatty acids in the diet. Saturated fatty acids (SFA) also increase LDL-C while polyunsaturated fatty acids (PUFA) decrease it [2,3]. A monounsaturated fatty acid (MUFA) diet was reported to reduce plasma total cholesterol and LDL-C levels without decreasing
O
Obbjjeeccttiivveess.. Monounsaturated fatty acid (MUFA)-rich diets were once considered an alternative therapy to prevent coronary heart disease. However, MUFA-rich diets were found to increase liver total cholesterol levels in animal experiments. The purpose of this study was to examine whether the addition of dietary cholesterol alters the effects of monounsaturated fatty acid (MUFA)-rich diets on lipid metabolism.
M
Meetthhooddss.. Diets containing 0%, 0.25%, 0.5%, 0.75%, and 1% (w/w) cholesterol were added to MUFA- rich meals and fed to rats for 21 days. The concentrations of plasma cholesterol, low-density lipoprotein-cholesterol (LDL-C), high-density lipoprotein-cholesterol (HDL-C), and triacylglycerol (TAG) were determined by enzymatic precipitation methods. The concentrations of hepatic cholesterol, TAG, and hepatic lipase (HL) were also determined.
R
Reessuullttss.. Plasma total cholesterol and LDL-C concentrations increased but the concentrations of HDL-C decreased in rats which were fed either a 0.75% or a 1% cholesterol diet. The LDL-C/HDL-C ratio significantly increased from 0.58 to 1.91 as the dietary cholesterol level increased from 0%
to 1%. The cholesterol-fed rats showed significant increases in hepatic cholesterol and TAG levels.
The addition of 0.25% (w/w) cholesterol resulted in increased hepatic cholesterol (2.9 fold) and TAG (72%) levels compared with a cholesterol-free diet. There was, however, no significant difference in HL activity between cholesterol-fed rats and rats which were fed a cholesterol-free diet.
C
Coonncclluussiioonnss.. Extra dietary cholesterol altered plasma and hepatic lipid concentrations in rats which were fed a MUFA-rich diet. ( Mid Taiwan J Med 2003;8:207-13)
K
Keeyy wwoorrddss
cholesterol, hepatic lipase, monounsaturated fatty acid, triacylglycerol
Received : August 5, 2003. Revised : September 4, 2003.
Accepted : September 5, 2003.
Address reprint requests to : Nai Wen Chang, Department of Biochemistry, China Medical University, 91 Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C.
Supplementation of Cholesterol Modulates the Effect of a Monounsaturated Fatty
Acid-Rich Diet on Plasma and Hepatic Lipids in Rats
Nai Wen Chang, Fei Na Chen1, Chen Teng Wu2
Department of Biochemistry, 1Department of Social Medicine, China Medical University, Taichung; 2Department
of Surgery, Feng Yuan Hospital, Department of Health, Executive Yuan, Taiwan, R.O.C.
the HDL-C induced by a PUFA diet in humans [4]. However, most rodent animal models do not seem to exhibit a hypocholesterolemic response to a MUFA diet [5-12]. In fact, a MUFA diet appears to increase the plasma and liver total cholesterol levels in rats [5-8], rabbits [12], guinea pigs [10,11], and hamsters [9].
Hepatic lipase (HL), which hydrolyzes both phospholipid (PL) and triacylglycerol (TAG) [13], is synthesized and secreted by liver parenchymal cells in humans. It functions as a phospholipase A1 and hydrolyzes fatty acids at positions 1 and 3 of TAG. HL was also shown in vitro to hydrolyze TAG in both very low density lipoprotein (VLDL) and chylomicron remnants [13]. In two of our previous studies, we found that an increase in plasma VLDL-C and VLDL-TAG levels was related to an increase in MUFA content in the diet; this high MUFA diet, therefore, resulted in greater accumulation of liver cholesterol in rats [6,7]. However, whether dietary supplementation of cholesterol alters the effects of MUFA-rich diets on lipid metabolism and HL activity in rats remains unclear.
The National Cholesterol Education Program (NCEP) considers MUFA-rich diets to be an alternative therapy to prevent coronary heart disease (CHD) [14]. MUFA was found,
however, to increase liver total cholesterol levels in animal experiments [5-12]. We, therefore, examined whether dietary cholesterol modifies the effects of MUFA-rich diets on plasma and hepatic cholesterol concentrations. The activities of hepatic lipase were also examined.
M
MAATTEERRIIAALLSS AANNDD MMEETTHHOODDSS Animals and Diets
Male Wistar rats were obtained from the Laboratory Animal Center, College of Medicine, National Taiwan University. They were housed individually in stainless steel, wire-bottomed cages, fed a MUFA-rich diet and given water ad libitum for 21 days. The MUFA-rich semisynthetic diet contained (w/w): 4% salt mixture (AIN-76, ICN), 1% vitamin mixture (AIN-76, ICN), 3% methyl cellulose, 20% casein, 20% fat, and 52% corn starch. The fat mixtures which contained 87.7% olive oil and 12.3%
soybean oil had a PUFA/SFA ratio of 1.0 and a (PUFA+MUFA)/SFA ratio of 5.7. The profiles of the fatty acids in the oil mixture (soybean oil and olive oil) were analyzed by high-performance liquid chromatography (HPLC) [15] to verify the correct PUFA/SFA and (PUFA+MUFA)/SFA ratios (Table 1). Thirty male Wistar rats 180 5 g were equally divided into five groups. Food consumption by each rat and their individual body weights were measured every three days. All rats were fed the same basic, semisynthetic powder diet supplemented with 0.0%, 0.25%, 0.5%, 0.75%, or 1% (w/w) cholesterol. Each diet supplied 15% of total energy as protein, 45% as carbohydrate, and 40% as fat.
Sample Collection and Analysis
Fasting blood samples were taken after decapitation on day 21. The plasma (1 mg EDTA/mL) was stored at 4 C before analysis.
Each liver was excised, weighed and frozen until analysis. Plasma total cholesterol, LDL-C, HDL- C, and TAG levels were determined by enzymatic precipitation methods (Boehringer Mannheim Co.).
The liver lipids were extracted by a chloroform-methanol 2:1 (v/v) mixture according
Fatty acids MUFA-rich diet
(g/100 g fatty acid) 14:0
16:0 18:0 Subtotal
16:1 18:1 Subtotal
18:2 18:3 Subtotal PUFA/SFA ratio
(PUFA+MUFA)/SFA ratio
Table 1. Fatty acid composition in MUFA-rich diet
0.2 11.2 3.5 14.9 0.6 69.7 70.3 13.4 1.4 14.8 1.0 5.7
Content of MUFA-rich diet (g/kg): 24.6 g soybean oil and 175.4 g olive oil. PUFA = polyunsaturated fatty acids; MUFA = monounsaturated fatty acids; SFA=
saturated fatty acids.
to Folch et al [16]. Liver total cholesterol was determined by the Abell [17] method. Liver TAG was determined by the Soloni [18] method.
Assay of Hepatic Lipase
Each liver was homogenized in 10 volumes of Kreb-Ringer-HEPES buffer (pH 7.4).
Each homogenate was centrifuged for 15 min at 6000 g at 4 C. The fat-free pellets were resuspended in 0.1 M TRIS-HCl (pH 9.0) containing 0.015 U heparin/L, and centrifuged for 15 min at 6000 g [19]. Hepatic lipase in the supernatants was determined as described previously [20]. Total assay volume was 0.1 mL containing 0.23 µmol tri[1-14C]oleoylgylcerol, 0.4 mg of gum arabic, and 0.5 mg of bovine serum albumin in 0.2 M Tris-HCl (pH 8.8) containing 0.75 M NaCl. The assay was started by adding 10 µL of enzyme fraction and incubated for 30 min at 37 C. The reaction was stopped by adding 1.6 mL of chloroform-heptane-methanol (5.0:4.0:5.6, by vol) followed by 0.5 mL of 0.5 M NaOH.
The samples were then centrifuged for 30 min at 3000 g. A 0.6 mL aliquot of the upper phase was counted by a Backman liquid scintillation counter (LS-250). One unit of enzyme activity represented the release of 1 µmol oleic acid per min at 37 C.
Statistical Analysis
Data were analyzed by one-way ANOVA, and by multiple comparison tests whenever a statistically significant difference was shown by ANOVA. A p value < 0.05 was considered
significant. All data are presented as mean SD.
R
REESSUULLTTSS
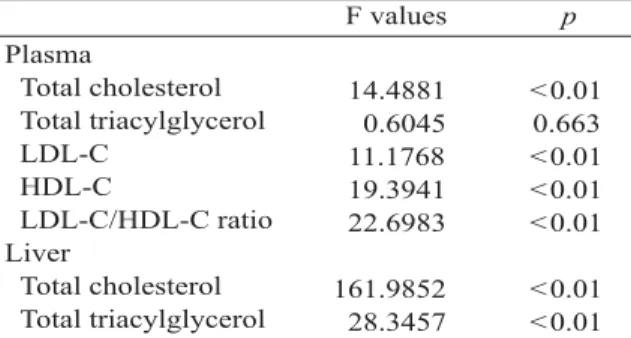
There were no significant differences in body weight gain or feeding efficiency (weight gain/100g diet) among the groups of rats (data not shown). Dietary cholesterol significantly raised plasma total cholesterol and LDL-C levels, and lowered HDL-C (p < 0.01, Table 2). However, there was no significant effect on plasma TAG levels. The addition of 0.75% or 1% cholesterol to the diet significantly increased plasma total cholesterol (p < 0.01, Table 3) compared with the control diet (0% cholesterol). The increase in total cholesterol was found in the LDL fraction (p <
0.05, Table 3), whereas cholesterol in the HDL fraction significantly decreased (p < 0.05, Table 3) in rats fed either a 0.75% or 1% cholesterol diet. The LDL-C/HDL-C ratio also significantly increased from 0.58 to 1.91 as the dietary cholesterol content increased from 0% to 1% (p <
0.05, Table 3).
The addition of cholesterol to MUFA significantly increased both liver cholesterol and TAG levels (Table 2, p < 0.01). The addition of 0.25% (w/w) cholesterol resulted in an increase in hepatic cholesterol (2.9 folds) and TAG (72%) levels compared with the 0% cholesterol diet (Table 4). Moreover, the hepatic cholesterol and TAG levels significantly increased from 21.6 2.3 and 48.9 9.0 µmol/g liver in diet with 0%
cholesterol supplement to 192.9 7.8 µmol/g liver (7.9 folds) and 120.8 12.3 µmol/g liver (1.5 folds) in diet with 1% cholesterol supplement (p < 0.05, Table 4), respectively.
In addition, there was no significant difference in hepatic lipase activity in cholesterol- fed rats (p > 0.05, Table 5).
D
DIISSCCUUSSSSIIOONN
The main goal of the present study was to examine the modulation of cholesterol supplementation on the effects that monounsaturated fatty acid (MUFA)-rich diets have on lipid metabolism. We found that plasma total cholesterol levels in rats fed a MUFA-rich F values
Plasma
Total cholesterol Total triacylglycerol LDL-C
HDL-C
LDL-C/HDL-C ratio Liver
Total cholesterol Total triacylglycerol
Table 2. Effect of dietary cholesterol on plasma and hepatic lipids
14.4881 0.6045 11.1768 19.3941 22.6983
161.9852 28.3457
p
0.01
0.01 0.01 0.01
0.01 0.01 0.663
<
<
<
<
<
<
LDL-C = low density lipoprotein-cholesterol; HDL-C = high density lipoprotein-cholesterol.
diet increased after the addition of 0.75% or 1%
cholesterol, while they remained unchanged in rats fed 0.25% or 0.5% extra cholesterol diets.
However, the LDL-C/HDL-C ratio significantly increased proportionally with cholesterol in diets.
The ratio of LDL-C/HDL-C was considered an index of the atherogenicity of lipoproteins; ie, the higher the ratio, the more atherogenic the lipoprotein profile was supposed to be. The plasma total cholesterol concentration was not significantly different in rats fed 0.25% or 0.5%
cholesterol. An increase in LDL-C and a decrease in HDL-C offset the effect of 0.25% or 0.5%
cholesterol diets on plasma total cholesterol concentrations. Furthermore, MUFA-rich diets were reported not to alter [21,22], to decrease [23], or to increase [4,24] plasma HDL-C concentrations compared with PUFA- or carbohydrate- rich diets. Thus, the effect of diets enriched in MUFA on HDL-C is still controversial. In this study, we found that supplementation of cholesterol reduced the
plasma HDL-C in rats fed high-MUFA diet.
In the presence of 0.25% cholesterol, there was a 2.9 fold increase in hepatic cholesterol and a 72% increase in TAG levels. When 1%
cholesterol was added, there was a 7.9 fold increase in hepatic TAG and a 1.5 fold increase in cholesterol level. The increase in TAG synthesis may result in increased VLDL secretion. In order to stabilize the hydrophobic TAG core, there is an increase in the secretion of VLDL-C [25]. The increase in VLDL secretion may also contribute to the increase in intermediate density lipoprotein (IDL) and LDL, which are associated with cholesterol feeding [25]. Moreover, the delivery of cholesterol to the liver resulted in decreased receptor-mediated uptake of LDL [26]. These effects of dietary cholesterol might result in an increase in plasma LDL-C/HDL-C ratio and an accumulation of hepatic cholesterol and TAG.
Most VLDLs are degraded to IDLs and then to LDLs. This step-wise catabolism of chylomicrons and VLDL might be mediated by Total cholesterol
( mmol/L) Cholesterol
(%)
1.94 2.08 2.44 2.64 3.16
0.21a 0.11a 0.31a,b 0.48b,c 0.32c,d
LDL-C ( mmol/L) 0.55 0.65 0.72 0.88 1.11
0.05a 0.10a,b 0.06a,b 0.20b,c 0.27c,d
HDL-C ( mmol/L) 0.95 0.57 0.65 0.46 0.58
0.12a 0.10b,c 0.06b 0.09c 0.13b,c
LDL-C/HDL-C (ratio) 0.58 1.14 1.11 1.91 1.91
0.05a 0.16a,b 0.08a,b 0.40b,c 0.43b,d Table 3. Effect of dietary cholesterol and MUFA-rich diet on plasma total cholesterol, LDL-C and HDL-C
Values are mean SD. Within each column, values with different superscript letters were significantly different at p < 0.05 by multiple comparison tests.
0 0.25 0.50 0.75 1.00
Cholesterol (µmol/g) Cholesterol
(%)
21.6 84.8 144.3 179.4 192.9
2.3a 18.5b 15.6c 16.9d 7.8d
Triacylglycerol (µmol/g) 48.9 84.1 132.1 127.4 120.8
9.0a 18.1b 9.9c 26.0c 12.3c Table 4. Effect of dietary cholesterol and MUFA-rich diet on hepatic cholesterol and triacylglycerol
Values are mean SD. Within each column, values with different superscript letters were significantly different at p < 0.05 by multiple comparison tests.
0 0.25 0.50 0.75 1.00
Cholesterol (%) Hepatic lipase (U/g) 9.08
9.83 9.17 12.25 11.09
3.51a 2.71a 1.67a 1.64a 1.93a Table 5. Hepatic lipase activities of cholesterol-fed rats
Hepatic lipase: 1U = 1 µmol oleic acid/min. Values are mean SD. Within the column, values with the same superscript letters were not significantly different at p <
0.05 by multiple comparison tests.
0 0.25 0.50 0.75 1.00
lipoprotein lipase and HL [13]. In the present study, however, we found that the increase in plasma lipid levels in rats which were fed cholesterol and MUFA-rich diets was not mediated by HL. Oleic acid was reported to have no effect on the release of HL, whereas VLDL secretion increased [27]. In addition, Okuno et al [28] suggested that the expression of lipoprotein lipase is not significantly affected in epididymal fat tissue of rats fed perilla oil, beef tallow or olive oil.
In summary, this study suggests that supplementation of cholesterol modifies the effect of MUFA-rich diets on plasma cholesterol and LDL-C concentrations. The existence of cholesterol enhances the effects of high MUFA diet on accumulation of hepatic cholesterol and TAG.
A
ACCKKNNOOWWLLEEDDGGMMEENNTT
This work was supported by grants (CMC90-M-017 and CMC-91-M-15) from the China Medical University, Taiwan, R.O.C.
R
REEFFEERREENNCCEESS
1. Hu FB, Manson J, Willett WC. Types of dietary fat and risk of coronary heart disease: a critical review.
[Review] J Am Coll Nutr 2001;20:5-19.
2. Schonfeld G, Patsch W, Rudel LL, et al. Effects of dietary cholesterol and fatty acids on plasma lipoproteins. J Clin Invest 1982;69:1072-80.
3. Woollett LA, Spady DK, Dietschy JM. Saturated and unsaturated fatty acids independently regulate low density lipoprotein receptor activity and production rate. J Lipid Res 1992;33:77-88.
4. Grundy SM. Comparison of monounsaturated fatty acids and carbohydrates for lowering plasma cholesterol. N Engl J Med 1986;314:745-8.
5. Beynen AC. Serum and liver cholesterol in rats fed cholesterol free or high-cholesterol diets differing in type and amount of fat. Nutr Rep Int 1987;35:1327-32.
6. Chang NW, Huang PC. Effects of the ratio of polyunsaturated and monounsaturated fatty acid to saturated fatty acid on rats plasma and liver lipid concentrations. Lipids 1998;33: 481-7.
7. Chang NW, Huang PC. Comparative effects of polyunsaturated-to saturated fatty acid ratio versus
polyunsaturated- and monounsaturated fatty acids to saturated fatty acid ratio on lipid metabolism in rats.
Atherosclerosis 1999;142:185-91.
8. Kris-Etherton PM, Ho CY, Fosmire MA. The effect of dietary fat saturation on plasma and hepatic lipoproteins in the rat. J Nutr 1984;114:1675-82.
9. Dietschy JM. Dietary fatty acids and the regulation of plasma low density lipoprotein cholesterol concentrations. [Review] J Nutr 1998;128(2 Suppl):S444-8.
10. Fernandez ML, McNamar DJ. Dietary fat-mediated changes in hepatic apoprotein B/E receptor in the guinea pig: effect of polyunsaturated, monounsaturated, and saturated fat. Metabolism 1989;38:1094-102.
11. Fernandez ML, McNamara DJ. Dietary fat saturation and chain length modulate guinea pig hepatic cholesterol metabolism. J Nutr 1994;124:331-9.
12. Beynen AC, Visser JJ, Schouten JA, et al. Cholesterol metabolism in rabbits fed diets containing either corn oil or olive oil. Nutr Rep Int 1987;35:111-6.
13. Musliner TA, Herbert PN, Kingston MJ. Lipoprotein substrates of lipoprotein lipase and hepatic triacylglycerol lipase from human post-heparin plasma. Biochim Biophys Acta 1979;575:277-88.
14. Executive Summary of the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2001;285:2486-97.
15. Miwa H, Yamamoto M. Improved method of determination of biologically important C10:0-C22:6 fatty acids as their 2-nitrophenylhydrazides by reversed-phase high-performance liquid chromatography. J Chromatogr 1986;351:275-82.
16. Folch J, Lees M, Sloane-Stanley GH. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 1957;226:497-509.
17. Abell LL, Brodie BB, Kendall FE. A simplified method for the estimation of total cholesterol in serum and demonstration of its specificity. J Biol Chem 1951;195:357- 67.
18. Soloni FG. Simplified manual micromethod for determination of serum triglycerides. Clin Chem 1971;17:529-34.
19. Haug A, Hostmark AT, Spydevold O. Plasma lipoprotein distribution, faecal cholesterol excretion, and activities of lipoprotein lipase, hepatic lipase, and lecithin: cholesterol acyltransferase in rats fed diets
rich in sucrose or sunflower oil. Acta Physiol Scand 1985;125:609-17.
20. B o b e r g J , A u g u s t i n J , B a g i n s ky ML, et al.
Quantitative determination of hepatic and lipoprotein lipase activities from human postheparin plasma. J Lipid Res 1977;18:544-7.
21. Dreon DM, Vranizan KM, Krauss RM, et al. The effects of polyunsaturated fat vs monounsaturated fat on plasma lipoproteins. JAMA 1990;263:2462-6.
22. Khosla P, Hayes KC. Comparision between the effects of dietary saturated (16:0), monounsaturated (18:1), and polyunsaturated (18:2) fatty acids on plasma lipoprotein metabolism in cebus and rhesus monkeys fed cholesterol-free diets. Am J Clin Nutr 1992;55:
51-62.
23. Wahrburg U, Martin H, Sandkamp M, et al.
Comparative effects of a recommended lipid-lowering diet vs a diet rich in monounsaturated fatty acids on serum lipids profiles in healthy young adults. Am J
Clin Nutr 1992;56:678-83.
24. McNamara DJ. Dietary fatty acids, lipoproteins, and cardiovascular disease. [Review] Adv Food Nutr Res 1992;36:253-351.
25. Fungwe TV, Cagen L, Wilcox HG, et al. Regulation of hepatic secretion of very low density lipoprotein by dietary cholesterol. J Lipid Res 1992;33:179-91.
26.Spady DK, Dietschy JM. Dietary saturated triacylglycerols suppress hepatic low density lipoprotein receptor activity in the hamster. Proc Natl Acad Sci USA 1985;82:4526-30.
27. Rustan AC, Nossen JO, Blomhoff JP, et al. Release of hepatic lipase and very low density lipoprotein by cultured rat hepatocytes. Int J Biochem 1986;18:
909-16.
28.Okuno M, Kajiwara K, Imai S, et al. Perilla oil prevents the excessive growth of visceral adipose tissue in rats by down-regulating adipocyte differentiation. J Nutr 1997;127:1752-7.
1 2
1 2
0% 0.25% 0.5% 0.75%
1% 21 - (LDL-C) -
(HDL-C)
(hepatic lipase)
0.75% 1% 21
- -
0% 1% LDL-C/HDL-C 0.58 1.91
0.25% 0%
2.9 72%
2003;8:207-13
404 91
2003 8 5 2003 9 4
2003 9 5