Taiwanin C selectively inhibits arecoline-induced oral cancer cell proliferation via ERK1/2 inactivation
Kuan-Ho Lin1#, Marthandam Asokan Shibu2#,Yueh-Hsiung Kuo3#, Yueh-Chiu Chen1, Hsi-Hsien Hsu4, 5, Da-Tian Bau6, Ming-Cheng Chen7, Chuan-Chou Tu8, Vijaya Padma Viswanadha9, Chih-Yang Huang 2, 10, 11, *
1Emergency Department, China Medical University Hospital, Taichung, Taiwan
2Graduate Institute of Basic Medical Science, China Medical University, Taichung, Taiwan.
3Department of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung, Taiwan.
4Division of Colorectal Surgery, Mackay Memorial Hospital, Taipei, Taiwan
5Mackay Medicine, Nursing and Management College, Taipei, Taiwan
6Terry Fox Cancer Research Laboratory, China Medical University Hospital, Taichung, Taiwan
7Division of Colorectal Surgery, Taichung Veterans General Hospital, Taichung, Taiwan
8Division of Chest Medicine, Department of Internal Medicine, Armed Force Taichung General Hospital, Taichung
9Department of Biotechnology, Bharathiar University, Coimbatore, India
10School of Chinese Medicine, China Medical University, Taichung, Taiwan
11Department of Health and Nutrition Biotechnology, Asia University, Taichung, Taiwan.
#These authors contributed equally to this work.
*Correspondence to Chih-Yang Huang Ph.D, Graduate Institute of Basic Medical 1
2 3 4 5
6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
Science, China Medical University, No. 91, Hsueh-Shib Road, Taichung, Taiwan. Tel.:
+886 4 22053366x3313; fax: +886 4 22032295. E-mail: [email protected] Abstract:
Oral cancer is one of the most common cancers in Taiwan and among other Southeast Asian countries due to the betel nut chewing habit that is prevalent in their population . Arecoline, the most abundant alkaloid in betel nut is known to promote abnormal proliferation of epithelial cells by enhancing epidermal growth factor receptor (EGFR) activation and cyclooxygenase-2 (COX2) expression. Taiwanin C, a naturally occurring lignan extracted from Taiwania cryptomerioides, is a potential COX2 inhibitor. The potential and the related mechanism of taiwanin C to inhibit arecoline induced oral cancer cell proliferation are not clearly known yet. The effect of taiwanin C on abnormal proliferation of T28 oral cancer cells and non-tumor N28 oral cells was assessed to determine the tumor specific inhibitory effect of taiwanin C. The modulations in the expression of relevant proteins were determined to understand the mechanism induced by taiwanin C to inhibit T28 cell proliferation. The levels of activated EGFR and COX2 were found to be abnormally high in the T28 oral cancer cells. However, taiwanin C was found to inhibit the activation of EGFR and regulated other related downstream proteins and thereby inhibited the T28 cell proliferation. In conclusion the results indicate that taiwanin C selectively suppresses arecoline induced proliferation of T28 cancer cells via ERK1/2 inactivation.
Key Words: Arecoline; COX2; ERK; Oral Cancer; Taiwanin C 24
25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45
Introduction
Oral cancer is one of the most common cancers reported in Taiwan and based on the mortality rate it ranks sixth in Taiwan (ROC) with similar trends worldwide .
Epidemiological studies have pointed out that betel quid chewing habit in Taiwan and other Southeast Asian countries is one of the most common risk factors in the
development of premalignant oral lesions and oral cancer. According to available reports, areca nut extracts have exhibited substantive mutagenic and genotoxic effects; they have also effectively induced preneoplastic as well as neoplastic lesions in experimental animals . Arecoline in areca nut is a potent carcinogenic substance, long-term exposure of oral mucosa to arecoline prompts activation of epidermal growth factor receptor (EGFR) in epithelial cells and elevates cycloxygenase-2 (COX2) expression and also causes abnormal proliferation of epithelial cells .
Many small molecular compounds isolated from plants have a great potential for medical applications such as in cancer treatment . A number of plant-derived compounds such as taxol (from Taxus brevifolia) and camptothecin derivatives (from Camptotheca
acuminate) have already been approved as drugs for cancer treatment . Taiwania
(Taiwania cryptomerioides hayata), an endemic plant in Taiwan, is a source for many bioactive compounds. In earlier investigations, various sesquiterpenes, lignans, and abietane-type diterpenes, including taiwanin A, savinin, taiwanin C, taiwanin E, helioxanthin, hinokinin, arctigenin, dimethyl matairesinol, acadinol, T-cadinol, and T- muurolol were been isolated and characterized .
Taiwanin C, a podophyllotoxin reported from various sources including Taiwania;
Justicia procumbens and Acanthopanax chiisanensis, although with poor cytotoxic effect 46
47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68
against different human cell lines, was found to significantly inhibit the COX2 activity (Fig. 1) . But the knowledge either on the mechanism of COX2 inhibition by taiwanin C or on whether the COX2 inhibition by taiwanin C could regulate the abnormal oral epithelial cell proliferation are not clearly known yet. In the experiments carried out in T28 oral squamous cancer cells and N28 normal oral squamous cells, taiwanin C was found to inhibit the cancer cell proliferation by promoting COX2 expression but it did not affect the normal cells. This is an ideal property of taiwanin C to be considered as a cancer treatment drug with limited side effects. The enhancement of COX2 was found to be mediated by the activation of extracellular-signal-regulated kinases (ERK).
69 70 71 72 73 74 75 76 77
Material and Methods:
Establishment of Cell lines
An oral squamous cell carcinoma model was created in C57BL/6J Narl male mice by oral administration of 0.5 mg/mL arecoline (Sigma, St. Louis, MO, USA) and 0.2 mg/mL 4- Nitroquinoline-1-oxide (Sigma) for 28 weeks through drinking water. Primary oral squamous carcinoma cell lines (T28) and normal cells (N28) were derived from oral squamous cell carcinoma model and control mice model after 28 weeks of administration. All protocols were reviewed and approved by the Institutional Review Board (IRB), Animal care and use committee of the China Medical University, Taichung, Republic of China (Taiwan), and the study was conducted in accordance with the principles of laboratory animal care . The cells were maintained in Dulbecco’s minimal medium (Sigma) containing 10% charcoal treated FBS (Hyclone, UT, USA), 1%
penicillin (Invitrogen Corp., Carlsbad, CA, USA), L-Glutamine (Sigma) and sodium bicarbonate (Sigma).
MTT (Thiazolyl blue tetrazilium bromide) Assay
The cell viability after taiwanin C treatment was measured using MTT assay (Sigma).
Cells were plated in triplicates in 24-well plates and treated with increasing
concentrations of taiwanin C. After 24 h of incubation, 0.5 mg/ml of MTT was added to each well and incubated for an additional 4 h period. The blue MTT formazan precipitate was then dissolved in 500 μl of dimethyl sulfoxide (DMSO, Sigma). The absorbance at 570 nm was measured on a multi-well plate reader. Cell viability was expressed as a percentage of control. For ERK activation, T28 cells were treated with 0.7 μM phorbol 78
79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99
12- myristate 13- acetatate (ERK activator, Sigma) for 24 h.
Western blot analysis
Cultured cells were lysed with lysis buffer (250 mM sucrose, 50 mMTris-HCl, 5 mM imidazole, 2.5 mM EDTA, 2.5 mM DTT, 0.1% Triton X-100, pH 7.40) and the
concentration of the extracted proteins was measured using the Lowry protein assay . An aliquot of each sample equivalent to 30 μg protein concentration was briefly boiled after addition of the appropriate volume of 5X sample buffer (5 mM EDTA, 162 mM DDT, 5% SDS, 50% glycerol, 0.05% bromophenol blue, 188 mM Tris, pH 8.8). The samples were separated on 10% SDS-polyacrylamide gels (PAGE) and electrophoretically transferred onto nitrocellulose filters using the Bio-Rad electrotransfer system (Bio-Rad Laboratories, Munich, Germany). Equal transfer was verified by ponceau staining of the membranes. The membranes were blotted with 1:1000 dilution of primary antibodies against p27 (Santa Cruz Biotechnology, CA, USA), COX2 (Santa Cruz Biotechnology), Cyclin D1 (Santa Cruz Biotechnology), EGFR (Cell Signaling) p-Tyr1068 EGFR (Abcam), β-actin (Santa Cruz Biotechnology).
Antigen-antibody complexes were visualized with HRP-coupled secondary antibodies (goat anti-mouse and goat anti-rabbit, Santa Cruz Biotechnology).
Statistical Analysis
Each sample was analyzed based on results that were repeated at least three times and Sigma Plot 10.0 software and standard t-test was used to analyze each numeric data. In all cases, differences at P < 0.05 were regarded as statistically significant; ones at P <
0.01 or P < 0.001 were considered higher statistical significances.
100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121
Results:
Taiwanin C inhibits oral cancer cell proliferation.
Normal oral cells-N28 and oral cancer cells-T28 were treated with different concentrations (0, 1, 5, 10, 30 and 60 µM) of taiwanin C and according to the cell viability measured after 24 h by MTT assay, taiwanin C inhibited proliferation of T28 cells in a dose-dependent manner and the IC50 was determined as 60 µM. On the other hand, N28 primary squamous cells were not inhibited by 5 to 30 μM of taiwanin C treatment however, 60 µM of taiwanin C induced cytotoxicity on N28 cells (Fig. 2A and 2B).
Effect of taiwanin C on EGFR signaling pathway and MAPK pathway proteins in oral cancer cells.
The modulation in the expression of MAPK pathway proteins were determined using western blot analysis. Taiwanin C treatment on T28 cells was found to suppress the levels of phosphorylated EGFR (P-EGFR) at different test concentrations (5-60 μM). The suppression in the activation of EGFR was dose dependent as evident from the P-Tyr1068 EGFR levels (Fig. 3). The influence of P-EGFR suppression on the MAPK proteins was determined by analyzing the expression levels of the relevant proteins such as ERK, c- Jun NH2-terminal kinases (JNK), p38 MAP Kinase. Among all the MAPK proteins that were examined, taiwanin C was found to influence only the levels of activated ERK1 and ERK2 and this suppression effect on the phosphorylated ERK1 and ERK2 (P-ERK1/2) was found to be dose dependent in the treatment groups (Fig. 3).
Taiwanin C regulates COX2 by suppressing ERK activation.
122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143
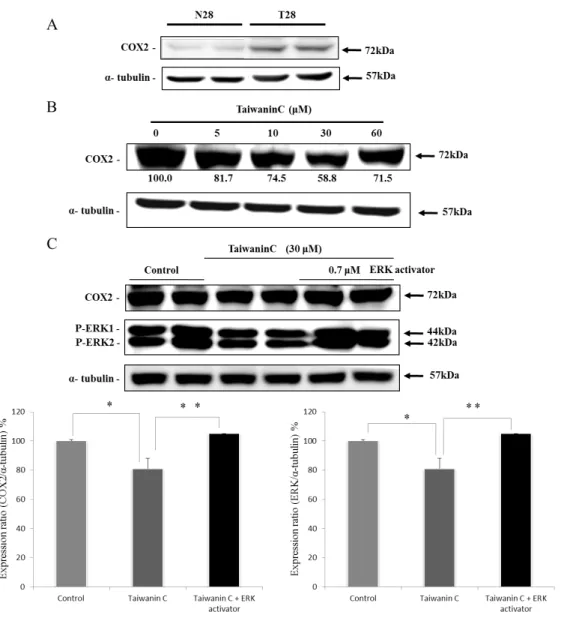
The related downstream protein COX2 was found to express only in T28 oral cancer cells, but not in N28 normal oral squamous cells and when treated with taiwanin C the COX2 levels in T28 cells were suppressed (Fig.3). Moreover, when co-treated with ERK activator and 30 µM taiwanin C, the expression levels of P-ERK1/2, and COX2 proteins were reverted (Fig. 4).
Effect of taiwanin C on cell cycle related proteins.
With the increase in taiwanin C concentration, the level of P27 protein expression increased and the level of cyclin D1 decreased. These modulations would further cause G1 phase arrest and inhibits the proliferation of T28 cells (Fig. 5A). Meanwhile, when T28 cells were co-treated with ERK activator the expression of P27 and the inhibition of cyclin D1were reverted. (Fig. 5).
Taiwanin C inhibits T28 cells via ERK activation
To find the role of ERK in the proliferation of T28 cells and the subsequent inhibition by taiwanin C, the influence of ERK activator in T28 cell proliferation was determined.
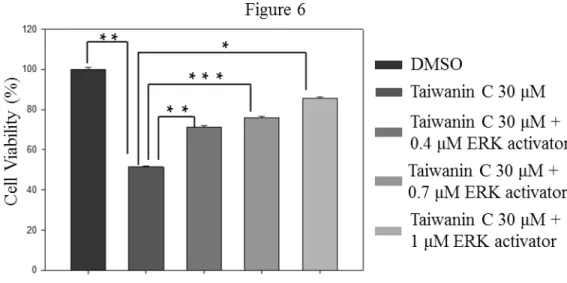
ERK activator co-treatment in T28 cell lines reversed the inhibition effect of taiwanin C in a dose dependent manner (Fig. 6).
144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159
Discussion:
In recent years, anticancer agents and their specific target elements in various
mechanisms of tumorogenesis are been intensively studied. The outcome of these studies would potentially help to improve treatment efficacy and minimize side effects of cancer therapy. EGFR is one of the best studied targets that play an important role in the
pathogenesis of multiple malignancies. EGFR is a growth factor receptor, which activates cell differentiation and hyperplasia. EGFR mediated activation of downstream proteins, such as: Ras, Raf, MAPK and transcription factors are associated with the activation of cell proliferation mechanism . In oral esophageal cancer, EGFR play an important role in oral cancer development as their overexpression and activation leads to the activation of telomerase and changes in the cell morphology in vitro . Further, EGFR mediated COX2 and Prostaglandin E2 (PGE2) expression in the epithelial cells also play an important role in inducing cancer .
The levels of activated EGFR were found to decline in a dose dependent manner when treated with taiwanin C and this modulation can alter the entire EGFR mediated signaling cascade in T28 cells. Extracellular signals from EGFR are communicated to the interior of the cell by protein phosphorylation that is catalyzed by kinases. The deregulation in the kinase activity particularly those that causes constitutively active kinases is a common feature observed in many cancers . MAPKs include the extracellular signal–regulated kinases, ERK1 and ERK2; the JNK enzymes that include JNK 1, JNK 2, and JNK 3; and the four p38 enzymes, p38α, p38β, p38γ, and p38δ. These enzymes are induced by various stimuli including numerous carcinogens and therefore these enzymes are widely considered to be potential markers and targets of cancer . The results show that taiwanin 160
161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182
C effectively suppresses the activation of ERK1 and ERK2 as evident from the decreased levels of their phosphorylated forms. However, taiwanin C did not affect the other MAPKs examined. Therefore the taiwanin C mediated inhibition of T28 cell proliferation may potentially involve ERK activation.
COX2 protein levels in T28 oral cancer cell lines were found to be in a higher concentration compared to the normal levels in the N28 normal oral squamous cells.
COX2 are known to have multiple roles such as in initiation, progression, invasion and metastasis of cancer. Carcinogenic substances such as arecoline formed at higher concentration during areca chewing are known to enhance COX2 and attribute to squamous cancer cell proliferation . PGE2 produced by COX2 transactivates the EGFR by rapid phosphorylation and therefore COX2 is been considered as one of the major targets of cancer therapy . Although, taiwanin C is known to inhibit COX2 expression and oral cancer cell proliferation the mechanism of COX2 inhibition is not clear yet. To find the upstream pathway related with COX2 expression and cancerous proliferation of T28 cells, modulation in the levels of few MAPK family proteins were determined.
The increase in the levels of COX2 expression in T28 cells was gradually suppressed when the cells were treated with increasing taiwanin C concentration and 30 μM of taiwanin C seemed to be the most efficient concentration. Inorder to find whether the suppression of COX2 expression is mediated via ERK pathway, the levels of the P- ERK1/2 with respect to COX2 levels in the T28 cells with and without taiwanin C treatment were determined. ERK activator mediated P-ERK1/2 up-regulation and the taiwanin C mediated down-regulation was found to be accompanied by corresponding and respective increase and decrease in levels of COX2 protein expression as observed 183
184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205
from the Western blot results. Therefore it can be interpreted that increased COX2 expression is in fact mediated by ERK activation.
Further the influence of taiwanin C on the level of G1- S cell cycle phase transition related cyclin D1 and that of cyclin dependent kinase (CDK) inhibitor protein p27 were determined. P27 is a tumor suppressor nuclear protein that inhibits the cyclin D1/CDK complex formation that is crucial during cell cycle . Therefore, expression of p27 has been associated with tumor progression, metastasis and poor prognosis in several cancers.
Taiwanin C enhanced the expression of p27 when treated alone but when co-treated with ERK activator the p27 expression was suppressed to low levels. Conversely, the cyclin D1 levels were suppressed when treated with 30 μM taiwanin C but the levels were reverted when treated in combination with ERK activator. This can be interpreted that the P27 mediated cyclin D1 inhibition by taiwanin C is dependent on ERK1/2 inactivation.
From the obtained experimental results it can be concluded that, taiwanin C inhibits the proliferation of T28 oral squamous cancer cells by regulating the EGFR activation that also involves the inhibitions of ERK1/2 activation and COX2 expression (Fig. 7). The COX2 inhibition further down regulates PGE2 mediated EGFR transactivation. The impact of EGFR activation and P-ERK1/2 down regulation by taiwanin C also increases the cell cycle protein P27, reflecting a recovery from hyperplastic conditions. Moreover, taiwanin C do not show any cytotoxicity at 0 -30 μM concentrations in normal oral squamous cells therefore, taiwanin C administration shows a promising potential to treat oral cancer.
FINANCIAL SUPPORT
This study was supported financially by the Taiwan Department of Health, Clinical Trial 206
207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228
and Research Center of Excellence (DOH102-TD-B-111-004).
229
Reference 230
231
Figure Captions Figure 1. Chemical structure of Taiwanin C.
Figure 2. Influence of taiwanin C on normal (N28) and oral cancer (T28) cellsin vitro. Treatment with taiwanin C reduced the cell viability of the T28 oral cancer cells (A) but 5-30 μM of taiwanin C did not show any cytotoxicity in N28 normal cells (B).
The cell viability was measured by MTT assay after treating with different concentrations of taiwaninC (0, 5, 10, 30, 60 μM). (***p<0.001 **p<0.01 *p<0.05).
Figure 3. Taiwanin C modulated EGFR, MAPK signaling pathway proteins in oral cancer cells. After treatment with taiwanin C in T28 cells the levels of EGFR, P-Tyr1068 EGFR and MAPK proteins were analyzed by Western blot using specific antibodies. The blots were probed with α-tubulin antibody as an internal control. Taiwanin C down regulated the levels of P-Tyr1068 EGFR, P-ERK1 and P-ERK2 as shown in the respective western blots.
Figure 4. Taiwanin C supresses COX2 and ERK activation. Western blot analysis on the influence of taiwanin C in COX2 expression in T28 oral cancer cells (A), in N28 normal oral cells (B) and the influence of ERK activator-taiwanin C co-treatment on COX2 and P-ERK1/2 expression in the T28oral cancer cells (C). (** p <0.01 * p<0.05).
Figure 5. Taiwanin C modulates the cell cycle regulatory proteins P27 in T28 oral cancer cells. (A) T28 cells were treated with taiwanin C for 24 h. Cell lysates were analyzed for the expression of P27 by Western blot using specific antibodies. The blots were reprobed with actin antibody for comparison of protein load.(B) T28 cells were treated with taiwanin C 30 μM and ERK activator 0.7 μM for 24 h. *p< 0.05 represent 232
233
234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253
significant differences when compared with the control group.
Figure 6. Effect of P-ERK1/2 activation on the inhibition effect of taiwanin C on T28 cell proliferation. Cell viability of T28 cells was measured by MTT assay after treatment with 30 μM taiwanin C and co-treatment with different concentrations of ERK activator (0.4, 0.7 and 1 μM).Cell viability was measured after 24 hr by MTT assay. (***p<0.001
**p<0.01 *p<0.05).
Figure 7. Mechanism of taiwanin C mediated Inhibition of T28 oral cancer cell proliferation.
254 255 256 257 258 259 260 261
Figures
Figure 1. Chemical structure of Taiwanin C.
262
263 264
Figure 2. Influence of taiwanin C on normal (N28) and oral cancer (T28) cellsin vitro. Treatment with taiwanin C reduced the cell viability of the T28 oral cancer cells (A) but 5-30 μM of taiwanin C did not show any cytotoxicity in N28 normal cells (B).
The cell viability was measured by MTT assay after treating with different concentrations of taiwaninC (0, 5, 10, 30, 60 μM). (***p<0.001 **p<0.01 *p<0.05).
265 266 267 268 269 270
Figure 3. Taiwanin C modulated EGFR, MAPK signaling pathway proteins in oral cancer cells. After treatment with taiwanin C in T28 cells the levels of EGFR, P-Tyr1068 EGFR and MAPK proteins were analyzed by Western blot using specific antibodies. The blots were probed with α-tubulin antibody as an internal control. Taiwanin C down regulated the levels of P-Tyr1068 EGFR, P-ERK1 and P-ERK2 as shown in the respective western blots.
271 272 273 274 275 276 277
Figure 4. Taiwanin C supresses COX2 and ERK activation. Western blot analysis on the influence of taiwanin C in COX2 expression in T28 oral cancer cells (A), in N28 normal oral cells (B) and the influence of ERK activator-taiwanin C co-treatment on COX2 and P-ERK1/2 expression in the T28oral cancer cells (C). (** p <0.01 * p<0.05).
278 279 280 281 282
Figure 5. Taiwanin C modulates the cell cycle regulatory proteins P27 in T28 oral cancer cells. (A) T28 cells were treated with taiwanin C for 24 h. Cell lysates were analyzed for the expression of P27 by Western blot using specific antibodies. The blots were reprobed with actin antibody for comparison of protein load.(B) T28 cells were treated with taiwanin C 30 μM and ERK activator 0.7 μM for 24 h. *p< 0.05 represent significant differences when compared with the control group.
283 284 285 286 287 288 289
Figure 6. Effect of P-ERK1/2 activation on the inhibition effect of taiwanin C on T28 cell proliferation. Cell viability of T28 cells was measured by MTT assay after treatment with 30 μM taiwanin C and co-treatment with different concentrations of ERK activator (0.4, 0.7 and 1 μM).Cell viability was measured after 24 hr by MTT assay. (***p<0.001
**p<0.01 *p<0.05).
290 291 292 293 294 295
Figure 7. Mechanism of taiwanin C mediated Inhibition of T28 oral cancer cell proliferation.
296 297 298