ALLELOPATHIC POTENTIAL OF Acacia confusa AND
RELATED SPECIES IN TAIWAN
CHANG-HUNG CHOU,1,2,* CHIU-YU FU,2 SHIN-YI LI,1
and YU-FA WANG1 1Institute of Botany, Academia Sinica
Taipei, Taiwan 115
2Department of Botany, National Taiwan University
Taipei, Taiwan
(Received February 19, 1998; accepted August 3, 1998)
Abstract—Acacia confusa (an endemic species) and other introduced species, namely A. aulacocarpa, A. auricumiformis, A. cincinnata, A. crassicarpa, A. leptocarpa, A. margium, A. polystachya, and A. torfilis were evaluated for allelopathic potential. Among these, A. confusa is widely distributed on the hills and lowlands of Taiwn and often exhibits a unique pattern of weed exclusion under stands. Four study sites were selected. Field observations and measurements were carried out at sites that exhibited relatively pure stands of A. confusa. Although the diversity of understory species was comparatively higher in Acacia stands than in adjacent weedy sites, the total coverage and biomass of understory plants was significantly lower than in control sites. Aqueous extracts (0.5%, 1%, 2%, 3%, 4%, and 5%) of dry leaves and litter of Acacia confusa and other species collected from various sites and dates were bioassayed using lettuce, alfalfa, and Chinese cabbage to examine their phytotoxicity. Most extracts exhibited phytotoxicity even at a concentration as low as 0.5%. Inhibition of radicle growth of test plants varied with sampling sites and dates. Bioassay with 5% extracts or above produced more than 85% inhibition of test plants regardless of habitat. In addition, surface soils col-lected from the upper 20 cm layer of Acacia stands revealed significant inhi-bition as compared with adjacent grassland control soils. The most inhibitory compounds isolated from the ether fraction of aqueous extracts were identified as: ferulic, vanillic, caffeic, gallic, m-hydroxybenzoic, and m-hydroxyphenyl-acetic acids. Unidentified flavonoids were also found. Chromatographic bioas-says of compounds isolated from both ethyl acetate and water fractions of
*To whom correspondence should be addressed.
2131
methanolic extracts of Acacia leaves also showed significant phytotoxicity but none was found in the fractions of chloroform and hexane, suggesting the phytotoxic compounds present in Acacia plants are water soluble.
Key Words—Acacia confusa, allelopathy, phytotoxicity, phytotoxin, phe-nolics, flavonoids, herbicide, sustainable agriculture.
INTRODUCTION
Acacia confusa Merr, an endemic species, is widely distributed on the hills and
lowland areas of Taiwan and in the subtropical regions of Asia (Kuo et al., 1989). The plant has been extensively used in many ways, including as a feed-stock, for charcoal-making, and as construction material, and it is particularly important for conservation of soil and water in Taiwan. It often exhibits a unique pattern of lower amounts of understory plants under the canopy. Observations and extensive studies in the field since 1986 have concluded that the pattern is not due to physical competition for light, soil moisture, or nutrients and suggests that the phenomenon could be due to a detrimental biochemical effect (allelo-pathy) of the Acacia plant on its understory plants (Kuo et al., 1989; Fu, 1995). Allelopathic research of subtropical vegetation in Taiwan, including
Phyllos-tachys edulis (Chou and Yang, 1982), Leucaena leucocephala (Chou and Kuo,
1986), Delonix regia (Chou and Leu, 1992), and Zelkova formosana (Chou et al., 1989), has been extensive. These plants show allelopathic potential, but have not yet been used as biological control herbicides.
In recent years, biologically active compounds released from plants, such as Azadirachta indica and Agrostemma githago L., have been used as naturally occurring herbicides, fungicides, and nematicides (Parmar and Singh, 1993; Welte and Szabolcs, 1987). From the view point of sustainable agriculture, allelopathic compounds are particularly meaningful as natural agrochemicals, with potential to protect our environment from pollution and maintain better ecosystem balance. Since there is a substantial quantity of Acacia plants in Taiwan, our goal is to use the plants, which possess phytotoxic substances, as natural herbicides. In a series of studies, we focused on the following tasks: (1) evaluating the phytotoxic activity of leaves, litter, pods, and twigs of the Acacia plants; (2) examining how the phytotoxicity of plants collected varied with seasons and habitats; (3) comparing the phytotoxicity among species of Acacia plants; (4) isolating and identifying phytotoxins from various parts of Acacia plants; and (5) confirming the potential use of phytotoxic compounds as natural herbicides in agricultural practice. Some findings of the aforementioned studies are presented in this paper.
CHOU, FU, LI, AND WANG 2132
METHODS AND MATERIALS
Study Sites. Four sites representing different weather types were situated
at: (1) Chaoushi in Ilan County and (2) Linkow in Taipei County, both belonging to the northeast humid zone; (3) Tachia in Taichung County, belonging to the central west summer rain zone; and (4) Fengkang and Liukua in Pingtung County in the southern tip of Taiwan with a long drought season in winter. In addition, a site chosen for comparison was on the slopes of Nankang mountain in the vicinity of the Institute of Botany, Academic Sinica, Taipei. The climatic pat-terns of these sites were described by Fu (1995). In general, precipitation in Taiwan is usually concentrated in the summer season. In particular, frequent typhoons hit Taiwan during the period from August to October. Northeastern Taiwan has a substantial amount of rainfall in winter in addition to summer rain; however, in the south, it is often dry in the autumn and winter.
Field Measurement. In each site, four plots, 10 X 10 m2 each, were selected
in a stand of Acacia confusa and in the vicinity adjacent to A. confusa. The coverage of each species per plot was measured, and each species present was identified.
Sampling. Leaves, litter, pods, and twigs of A. confusa were sampled from
all four sites in June, September, and December 1994 and March 1995. In addition, leaves and litter of nine Acacia species, namely, A. anlacocarpa,
A. auricumiformis, A. cincinnata, A. confusa, A. crassicarpa, A. leptocarpa, A. mangium, A. polystachya, and A. torfilis growing in the experimental farm
of Liukua were also collected in October 1996. Samples were brought back to the laboratory and allowed to air dry at room temperature. The dry matter was ground into powder (60 mesh screen) for water and methanol extraction.
Soils from the upper 20 cm layer of Acacia floor and that of control grass-land area were collected and brought to the laboratory. All samples including soils were collected from the field after a dry period of at least two weeks. They were allowed to air dry before use.
Preparation of Aqueous Extracts. Materials were allowed to dry, then
ground to a powder. A series of aqueous extracts, 0.5%. (0.5 g plant sample + 99.5 ml distilled water), 1%, 2%, 3%, 4%, and 5%, of each sample was prepared by using techniques described by Chou and Muller (1972).
Bioassays. Aqueous extracts of all six concentrations of each sample were
separately biossayed as described by Muller (1966) and Chou (1997). Seeds of radish (Raphanus acanthiformis), lettuce (Lactuca sativa), Chinese cabbage
(Brassica chinensis), and alfalfa (Medicago sativa) were used as test species.
In addition, a chromatographic bioassay was employed to determine biologically active compounds present in fractions of hexane, chloroform, ethyl acetate, and water of a methanolic extract of Acacia confusa leaves. Lettuce seeds were used as test plants (McPherson et al., 1971; Chou, 1997).
Determination of Phytotoxicity of Decomposed Acacia Leaves in Soils. Acacia leaf powder in amounts of 0.5, 1.0, and 2.0 g was separately mixed
with 100 g loamy soil, which was obtained from the experimental farm of Academia Sinica, placed in pots, and allowed to decompose for 2, 4, and 12 days, respectively. After the decomposition time, 15 lettuce seeds were planted in the soil mixture. Seven days later, the number of seeds that germinated and radicle growth of lettuce in millimeters was measured. The experiment was replicated twice.
Identification of Phytotoxic Compounds. Paper strip chromatography of the
ether fraction of aqueous extracts of Acacia leaves was employed as described by Wang et al. (1967), and thin-layer and column chromatography of the ethyl acetate and water fractions of methanolic extract of the plant leaves were employed as described by Mabry et al. (1970). A solvent of 2% acetic acid was used for paper strip descending chromatography to separate and identify phenolic compounds, while two other solvent systems, t-butanol-acetic acid-water (3 : 1 : 1, v/v/v) and 15% HOAc were employed in two-dimensional paper chro-matography to separate flavonoids. Additional solvent systems for thin-layer and column chromatography followed the systems of Mabry et al. (1970). Com-pounds isolated from column chromatography were further purified by affinity column chromatography by using Sephadex LH20. Phenolic compounds from
Acacia were cochromatographed with phenolic standards on TLC (Chou, 1997)
and HPLC (Chou and Leu, 1992). Flavonoids were identified by UV-visible and mass spectrophotometry.
Statistics. Bioassay data were analyzed by the Student's t test or Duncan's
multiple range test (Gomez and Gomez, 1976).
RESULTS
Comparison of Vegetation Composition Under Acacia and Adjacent Grass-land. Based on long-term field observation (more than 10 years), we found that
there is usually bare ground beneath Acacia stands, and the density of the understory plants is low. This phenomenon is particularly pronounced in the drought area with low winter precipitation at Fengkang in Pingtung County in southern Taiwan (Figure 1, top), and on the hillsides along the Chung-Shan Freeway in the Linkow area about 40 miles south of Taipei city (Figure 1, bottom).
Although Acacia confusa is distributed island-wide, and four study sites were chosen for the field experiments, we only present data from two distinctly different sites, namely Linkow (representing northern Taiwan) and Fengkang (representing the south). The floristic composition and coverage of each species per plot between the Acacia stands and adjacent grassland area are given in
FIG. 1. Relatively bare surface under the canopy of Acacia confusa; (top) at the Tachia site and (bottom) from the roadside along Chungshan freeway at Linkow area.
Table 1. The vegetation composition on the A. confusa floor was quite different between the two sites. Similarly, the floristic composition in each grassland area was also different, perhaps due to weather patterns. The Linkow site has a subtropical climate with a high annual rainfall, around 3000 mm, and is humid most of year; the Fengkeng site has a tropical climate with a long dry winter season and a comparatively low amount of annual rainfall, around 2000 mm.
Both sites were formerly abandoned fields. After several years of Acacia plantation, the number of species was relatively higher in the A. confusa stand than in the grassland area. However, the total coverage of species present in these sites was significantly lower on the Acacia floor than in the grassland area. It was more pronounced at the Fengkang site; for example, the relative coverage was only 43.6% on the Acacia floor as compared with 100% in the grassland area. Understory species coverage was reduced 21.4% at the Linkow site and 56.4% at the Fengkang site. Seed germination of understory was not, however, noticeably suppressed. Growth performance of plants in the grassland area was better than that in Acacia. For example, in the grassland control plot at the Fengkang site, the percent coverage of the two dominant plants, Ipomoea
acu-minata and Lantana camara, reached 50% and 70%, respectively; however, in
the Acacia plot, the coverage was nearly zero. Although the phenomenon was not pronounced at the Linkow site, the percent coverage of dominant species in
2136 CHOU, FU, LI, AND WANG
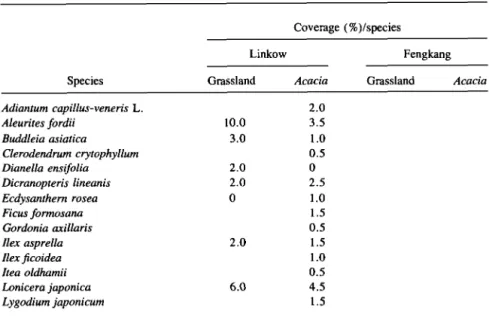
TABLE 1. FLORISTIC COMPOSITION AND COVERAGE OF EACH SPECIES PRESENT IN LINKOW AND FENGKANG
Species Adiantum capillus-veneris L. Aleurites fordii Buddleia asiatica Clerodendrum crytophyllum Dianella ensifolia Dicranopteris lineanis Ecdysanthern rosea Ficus formosana Gordonia axillaris Ilex asprella Ilex ficoidea Itea oldhamii Lonicera japonica Lygodium japonicum Coverage (%)/species Linkow Grassland 10.0 3.0 2.0 2.0 0 2.0 6.0 Acacia 2.0 3.5 1.0 0.5 0 2.5 1.0 1.5 0.5 1.5 1.0 0.5 4.5 1.5 Fengkang Grassland Acacia
TABLE 1. CONTINUED Species Miscanthus floridulus Mollotus paniculatus Passiflora edulis Pleuchea indica Psychstria rubra Randia cochinchinensis Sarcandra glabra Schefflera octophylla Smilax china Thea sinensis Zanthoxylum nitidum Acronychia pedunculata Agave sislana Albizzia julibrissin Breynia afficinalis Clausena excavata Crotalaria pallida Eleusine indica Euphorbia hirta Gelonium aequoreum Geodorum densiflorum Ipomoea acuminata Jasminum hemsleyi Lantana camara Lophantherum gracile Lygodium japonicum Maytenns diversifolia Melothria formosana Passiflora foetida Passiflora subnosa Phyllanthus urinaria Securinega virosa Severinia baxifolia Sida heterophylla Tabernaemontana divaricata Trichanthes kirilowii Tridax procumbens Vitex negundo
Average total coverage (%) % of grassland control Number of species Coverage (%)/species Linkow Grassland 4.0 8.0 4.0 5.0 3.0 2.0 5.0 56.0 100.0 13.0 Acacia 2.0 1.0 2.0 0.5 5.0 0.5 2.5 3.5 1.0 2.5 1.5 44.0 78.6 24.0 Fengkang Grassland 0 0 7.0 0 0 3.0 8.0 5.0 50.0 70.0 5.0 2.0 5.0 3.0 3.0 15.0 176.0 100.0 12.0 Acacia 2.0 8.0 9.0 2.5 5.0 1.0 1.5 2.5 3.0 0.25 15.5 5.0 3.0 0.5 4.0 4.0 5.0 2.5 7.5 76.75 43.6 18.0
the grassland area was relatively high; e.g., Aleurites fordii, Buddleia asiatica,
Miscanthus floridulus, Sarcandra glabra, Smilax china, Thea sinensis, Passi-flora edulis, and Lonicera japonica (Table 1) as compared with that found in
the Acacia confusa stand.
Phytotoxicity of Acacia Leaves Varies with Habitats and Seasons. The
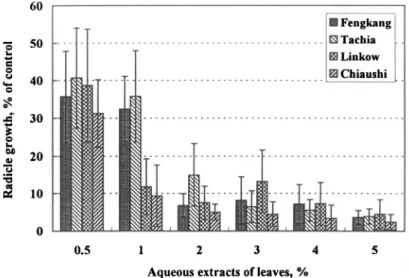
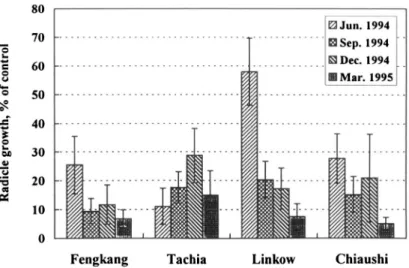
aqueous extracts, 0.5%, 1%, 2%, 3%, 4% and 5%, of A. confusa leaves col-lected in June, September, and December 1994 and March 1995 at the Feng-kang, Tachia, Linkow, and Chiaushi sites were bioassayed by using lettuce seeds as test species. In general, the radicle growth of lettuce was suppressed by the extracts at concentrations as low as 0.5%, and the inhibition increased with increasing concentration. To simplify the presentation of all bioassay data, we have selected only representative results. For example, for 0.5% leaf extracts of samples obtained in May 1995, the inhibition ranged from 60% to 70%, while for 5% extracts the inhibition reached 95% regardless of sampling site (Figure 2). The variation in phytotoxicity was not significantly different between sites when the concentration was above 2%. By using the findings of 2% extracts for comparison among sampling times, the inhibition varied significantly with sampling seasons. Except for samples from Tachia, those obtained in March 1995 revealed higher inhibition (Figure 3), suggesting that phytotoxic metabo-lites accumulated in leaves during the winter and early spring (from November to April in southern Taiwan, which was under a severe drought).
FIG. 2. Effect of aqueous extracts (0.5-5%) of A. confusa leaves on radicle growth of lettuce. The samples were collected from four sites in March 1995.
CHOU, FU, LI, AND WANG 2138
FIG. 3. Effect of 2% aqueous extracts of Acacia confusa leaves on radicle growth of lettuce.
One might question whether the inhibition may be due to an osmotic effect. The osmotic concentration of the 0.5% extracts was below 5 mosmol, while that of 5% extracts ranged from 40 to 50 mosmol (Chou, unpublished data). Chou and Young (1974) indicated that osmotic concentrations of extracts below 30 mosmol would not cause a significant osmotic inhibition. Normally, when the osmotic concentration of extracts exceeds 50 mosmol, the concentration of extract may cause 20-30% inhibition. In a case in which the osmotic concen-tration of extract is 50 mosmol and the inhibition is above 50%, there is at least a 20% inhibition due to a phytotoxic effect. In the present study, even with the 5% extract, the inhibition ranged from 80% to 90%, indicating that significant phytotoxic effects from the Acacia plants occurred. We also conclude that the phytotoxic activity of Acacia plants growing in southern Taiwan was higher than that in the north, suggesting that drought increases phytotoxic activity. Generally, the phytotoxicity of Acacia leaves collected in various seasons was in an ascending order of spring < summer < autumn < winter.
Phytotoxicity of Acacia confusa Twigs Varies with Habitats and Seasons.
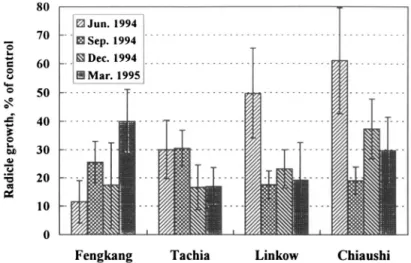
Aqueous extracts of 0.5%, 1%, 2%, 3%, 4%, and 5% Acacia twigs collected from four seasons—summer (June), autumn (September), and winter (Decem-ber) of 1994 and spring (March) of 1995—at the four sites were bioassayed against lettuce. The results, represented by the samples of March 1995, showed that at the 0.5 % aqueous extracts, inhibition of lettuce varied from 25 % to 65 %. The inhibition generally increased with extract concentration and reached
CHOU, FU, LI, AND WANG
FIG. 4. Effect of aqueous extracts, 0.5-5%, of A. confusa twigs on radicle growth of lettuce.
90-95% inhibition for 5% extracts (Figure 4). Using bioassay results from 2% extracts from four seasons and four sites for comparison, the inhibition of Feng-kang samples was generally higher than that of other sites; however, the inhi-bition seems to be variable with sampling times with no general trend (Figure 5).
FIG. 5. Effect of 2% aqueous extracts of A. confusa twigs on radicle growth of lettuce. 2140
In the overall comparison of phytotoxicity between leaves and twigs, we conclude that phytotoxicity is higher in leaves; in addition, in drought areas like Fengkang and Tachia, the phytotoxic metabolites may not be leached out during the winter season, resulting in markedly high inhibition later.
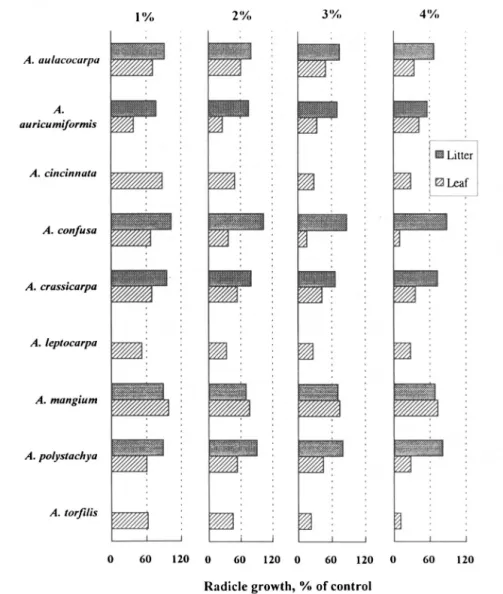
Phytotoxicity Varied with Acacia Species Grown in Liukua. Different
con-FIG. 6. Effect of aqueous leaf and litter extracts (1-4%) of different Acacia species from the Liukua experimental farm on radicle growth of lettuce.
centrations of aqueous extracts (1%, 2%, 3%, and 4%) of leaves (F) and litter (L) of Acacia plants grown in Liukua were bioassayed separately by using germination and radicle growth of seeds of lettuce, Chinese cabbage, and alfalfa. Only the lettuce findings are presented (Figure 6). In general, phytotoxicity in leaves was significantly higher than in litter. For example, comparing the 1% extracts, the inhibition of radicle growth was highest in A. auricumiformis (60%), followed by A. leptocarpa, A. polystachya, A. torfilis, A. crassicarpa, A.
con-fusa, A. aulacocarpa, and was lowest in A. mangium. Patterns of inhibition
shown by the remaining extracts from 2% to 4% were similar to that of the 1% extract; normally, inhibition increased with extract concentration (Figure 6). By comparing the effects of 5 % aqueous extracts of nine Acacia species on radicle growth of Chinese cabbage, alfalfa, and lettuce, we demonstrated that most litter extracts had no significant inhibition (Chou, unpublished data). However, leaf extracts of the aforementioned Acacia species did exhibit inhibition (Figure 7). Among the nine species, phytotoxicity was markedly higher in A. torfilis and A. confusa. However, with the alfalfa bioassay material, the inhibition was much lower in extracts of A. auricumiformis, A. crassicarpa, A. leptocarpa, and A. mangium. We conclude that phytotoxicity of Acacia varies with test species and the inhibition is usually higher in leaves than in litter.
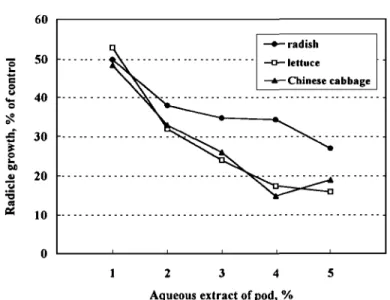
Effect of Aqueous Extract of Seed Pods of A. confusa on Plant Growth.
The aqueous extracts at 1-5% of seed pods were bioassayed against Chinese cabbage, lettuce, and radish. Inhibition ranged from 45% (1% extract) to 65% (5% extract) and was not statistically different between Chinese cabbage and lettuce (Figure 8). Seed pods, thus, possess some phytotoxic activity.
Effect of Aqueous Extracts of Acacia Soils on Plant Growth. Aqueous
extracts of soil samples from the Acacia floor and adjacent open areas as control at the Tachia and Fengkang sites were obtained. The extracts were bioassayed against two test species, radish and lettuce. Results of the bioassay are given in Figure 9A and B. Generally, the growth of test species was affected by soil extracts obtained from both depths of sampled soil regardless of soil habitat. There was no significant difference between depths. In general, the suppression of radicle growth of lettuce reached more than 50% in the Acacia soil, but radish was not affected at all. However, with open ground soils, the radicle growth of lettuce was 40% suppressed by Fengkang soil, but was not affected by Tachia soil (Figure 9). Thus, Acacia soil can possess both stimulatory and inhibitory properties.
Degradation of Phytotoxicity of Decomposed Leaves in Soils. Results are
presented in Figure 10. They show that phytotoxicity was below 40% in the early two- and four-day period of decomposition and lasted for 12 days. The inhibition was not significantly different among treatments that increased the amount of leaves mixed with the soil. The inhibition gradually disappeared CHOU, FU, LI, AND WANG 2142
FIG. 7. Effect of 5% aqueous extracts of leaves of Acacia plants from the Liukua site on radicle growth of three tested species.
during 4-12 days of decomposition even when the amount of leaf powder mixed in the soil was increased. This suggests that allelopathic activity of Acacia plant parts might not persist once the leaves have fallen to the ground. It also suggests that the phytotoxicity of Acacia leaves does not last long when decomposed in soil. This finding supports the low phytotoxicity finding revealed from the bioas-say of Acacia litter (Figure 6).
Phytotoxicity of Compounds in Methanolic Extracts of Acacia Leaves. Four
2144 CHOU, FU, LI, AND WANG
FIG. 8. Effect of aqueous extracts at 1-5% of A. confusa pod on radicle growth of radish, lettuce, and Chinese cabbage.
of Acacia leaves were chromatographed with Whatmann 3MM chromatographic paper. Spots on a two-dimensional paper chromatogram of the four fractions were cut out and bioassayed against lettuce seeds, using chromatographic bioas-say techniques (Chou, 1997). Results of the bioasbioas-say showed that only com-pounds isolated from the fractions of ethyl acetate and water produced significant inhibition of lettuce growth (Figures 11 and 12). For example, among 14 isolated spots from the ethyl acetate fraction, nine revealed an inhibitory effect on lettuce. Spots D, I, M, and N exhibited inhibition from 30% to 40% (Figure 11). On the other hand, nine spots in the water fraction revealed an inhibitory effect, and three spots exhibited a stimulatory effect (Figure 12). In particular, spots F, G, H, J, K, L revealed 25-60% phytotoxicity, and spots G and J were distinguished (Figure 12). The spots were isolated and purified, but the amounts were too small for further assays.
Identity of Phytotoxic Compounds. Since the aforementioned results showed
that potential allelopathic compounds were water-soluble, the aqueous extract was extracted with anhydrous ethyl ether. The ether fraction was chromato-graphed, and phytotoxic phenolics were identified by PC and HPLC. The com-pounds identified are: ferulic, vanillic, gallic, caffeic, m-hydroxybenzoic, and m-hydroxyphenylacetic acids (Table 2). Several unidentified compounds are thought to be flavonoids.
FIG. 9. Effect of aqueous extracts of soils on radicle growth of radish and lettuce. The soils were sampled from two depths of both Acacia and nearby weed dominated soils at two sites: (A) Tachia, (B) Fengkang.
DISCUSSION
As mentioned, field measurements previously revealed that Acacia may suppress the growth of understory plants without much involvement of physical competition for light, soil moisture, and nutrients (Kuo et al., 1989; Fu, 1995). Based on our results, we suggest that allelopathy plays a significant role in
CHOU, FU, LI, AND WANG
FIG. 10. Effect of decomposing Acacia leaves mixed with loam soil on radicle growth of lettuce.
regulating the species diversity, coverage, and dominance of Acacia associated vegetation in the field. This pattern of weed exclusion is not noticeable in some areas where seasonal monsoons take place. The phytotoxicity of fallen Acacia leaves seems to dissipate after heavy rainfall, and potential allelopathic com-pounds are easily degraded when the leaves are incorporated into the soil, as shown in Figure 10. Miles (1979) stated that the existence of phytotoxic sub-stances in a particular plant does not necessarily mean they are released into the environment of another plant in concentrations sufficient to suppress its growth in some aspect. This may explain why leachates of entire Acacia confusa leaves revealed significant phytotoxicity but not in high toxic levels, possibly due to the thick cuticle layer of the leaf; however, aqueous extracts of ground leaves did exhibit a high toxicity of 95% against tested plants. Our evidence shows that phytotoxins present in Acacia leaves are being decomposed within a few days in loamy soil. Allelopathic compounds released into the soil environment could be bound with humic acid or polymerized, thus, rapidly losing their tox-icity (Wang et al., 1978).
One of our major interests in Acacia is to use the compounds present in its plant parts as natural herbicides. Evidence of the role of allelopathy in weed control has been cited in many publications (Chou, 1995; Rice, 1995; Rizvi 2146
FIG. 11. Effect of compounds isolated from paper chromatograms of the ethyl acetate fraction of methanol extracts of A. confusa on the radicle growth of lettuce. Solvent systems are: TBA = r-butyl alcohol-acetic acid-water (3 : 1 : 1, v/v/v), and 15% HOAc = acetic acid-water (15:85, v/v).
and Rizvi, 1992; Waller, 1987). It is agreed that the proper use of allelopathic plants or natural products produced by plants or microorganisms can decrease the use of expensive synthetic herbicides that deteriorate environmental quality. However, they also point out that this goal is difficult to achieve due to the lack of economic incentive. However, there are several examples of successful use in agricultural practice. For example, the neem plant (Azadirachta indica A) has been widely used in India as a fungicide, and nematocide (Parmar and Singh, 1993). A natural product of Agrostemin, obtained from corn cockle of
Agros-CHOU, FU, LI, AND WANG
FIG. 12. Effect of compounds isolated from the water fraction of methanol extracts of A. confusa on radicle growth of lettuce.
temma githago L, has been widely used in eastern European countries to increase
the productivity of many crops; it also suppresses weed growth (Welte and Szabolcs, 1987). In Taiwan, Chou et al. (1989) demonstrated a case study of a grass-forest intercropping system. A grass, Pennisetum clandestinum, was planted on the deforested land of a coniferous tree, Cunninghamia lanceolata. After planting, the grass became predominant and suppressed the growth of many weeds in situ. The bioassay of the aqueous extract of grass revealed significant phytotoxicity; however, the grass was not harmful to the growth of coniferous trees or to the hardwood forest plants. Moreover, the grass could be used as a forage crop for livestock.
From the aforementioned findings, we conclude that Acacia confusa is an allelopathic plant, the leaves of which contain the major phytotoxic agent. In addition, the phytotoxins present are water soluble. Some of the responsible allelopathic compounds have been isolated and some of the phytotoxic phenolics have been identified. The isolation and identification of more compounds, such as flavonoids, are in progress.
Acknowledgments—The authors would like to express their sincere appreciation to Dr. C. C. Yang of the Taiwan Forestry Research Institute for providing plant material of the Acacia species grown in Liukau. This study was supported by grants to C. H. Chou awarded by the Academica Sinica, Taipei and the National Science Council (NSC 86-2311-B001-042, and 87-2311-B001-017) of the Republic of China on Taiwan.
REFERENCES
CHOU, C. H. 1995. Allelopathy and sustainable agriculture, pp. 211-223, in Inderji, K. M. M. Dakshini, and F. A. Einhellig (eds.). Allelopathy: Organisms, Processes and Applications. ACS Series No. 582. American Chemical Society, Washington, DC.
CHOU, C. H. 1997. Methodology for allelopathic research. Proceedings of the 1st International Congress of Allelopathy. September 17-23, 1996. Cadiz, Spain.
CHOU, C. H., and Kuo, Y. L. 1986. Allelopathic research in subtropical vegetation in Taiwan HI. Allelopathic exclusion of understory species by Leucaena lencocephala. J. Chem. Ecol. 12:1431-1448.
CHOU, C. H., and Leu, L. L. 1992. Allelopathic substances and interactions of Delonix regix (Boj. Raf.). J. Chem. Ecol. 18:2285-2303.
CHOU, C. H., and MULLER, C. H. 1972. Allelopathic mechanism of Arctostaphylos glandulosa var. zacaensis. Am. Midl. Nat. 88:324-347.
CHOU, C. H., and YANG, C. M. 1982. Allelopathic research of subtropical vegetation in Taiwan II. Comparative exclusion of understory by Phyllostachys edulis and Cryptomeria japonica. J. Chem. Ecol. 8:1489-1507.
TABLE 2. ALLELOPATHIC SUBSTANCES FOUND IN AQUEOUS EXTRACTS OF LEAVES OF
acacia confusaa Compound Caffeic acid m-Hydroxybenzoic acid m-Hydroxyphenylacetic acid Ferulic acid Vanillic acid Gallic acid PC + + + + + + HPLC _ -+ + + +
aThe compounds were identified by paper chromatography (PC) and high performance liquid
CHOU, C. H., and YOUNG, C. C. 1974. Effects of osmotic concentration and pH on plant growth. Taiwania 19:157-193.
CHOU, C. H., CHANG, C. M., WANG, Y. C., Hsu, F. H., and DEN, W. H. 1989. The selective allelopathic interaction of a pasture-forest inter-cropping in Taiwan II. Interaction between kikuyu grass and three hardwood plants. Plant Soil. 101:207-215.
Fu, C. Y. 1995. The allelopathic potential of Acacia confusa Merr. in Taiwan. MS thesis. National Taiwan University, Taipei, Taiwan, 79 pp.
GOMEZ, K. A., and GOMEZ, A. A. 1976. Statistical Procedures for Agricultural Research with Emphasis on Rice. The International Rice Research Institute, Los Banos, Philippines. Kuo, Y. L., CHIU, C. Y., and CHOU, C. H. 1989. Comparative allelopathic dominance of tropical
vegetation in Hengchun penisula of southern Taiwan, in H. C. Chou and G. R. Waller (eds.). Phytochemical Ecology. Institute of Botany, Academia Sinica Monograph Series No. 9, Taipei. MABRY, T. J., MARKAM, K. R., and THOMAS, M. B. 1970. The Systematic Identification of
Fla-vonoids. Springer-Verlag, New York.
MCPHERSON, J. K., CHOU, C. H., and MULLER, C. H. 1971. Allelopathic constituents of chaparral shrub Adenostome fasciculatum. Phytochemistry 10:2925-2933.
MILES, J. 1979. Vegetation Dynamics. Cambridge University Press, Cambridge, UK, 80 p. MULLER, C. H. 1966. The role of chemical inhibition (allelopathy) in vegetation composition. Bull.
Torrey Bot. Club 93:332-356.
PARMAR, B. S., and SINGH, R. P. 1993. Neem in Agriculture. Indian Agricultural Research Institute, New Delhi, 85 pp.
RICE, E. L. 1995. Biological Control of Weeds and Plant Diseases: Advances in Applied Allelo-pathy. University of Oklahoma Press, Norman.
RIZVI, S. J. H., and RIZVI, V. (eds.). 1992. Allelopathy: Basic and Applied Aspects. Chapman & Hall, London.
WALLER, G. R. (ed.). 1987. Allelochemical: Role in Agriculture and Forestry. ACS Symposium Series 330. American Chemical Society, Washington, DC.
WANG, T. S. C., YANG, T. K., and CHUANG, T. T. 1967. Soil phenolic acids as plant growth inhibitors. Soil Sci. 103:239-246.
WANG, T. S. C., Li., S. W., and FERNG, Y. L. 1978. Catalytic polymerization of phenolic com-pounds by clay mineral. Soil Sci. 126:16-21.
WELTE, E., and SZABOLCS, I. (eds.). 1997. Agrostemin. International Science Center of Fertilizers, Belgrade. 120 pp.
CHOU, FU, LI, AND WANG 2150