國立臺灣大學生物資源暨農學院園藝暨景觀學系 碩士論文
Department of Horticulture and Landscape Architecture College of Bioresources and Agriculture
National Taiwan University Master Thesis
鎳與尿素對水耕‘桃園一號’蕹菜生長、要素及 硝酸鹽含量之影響
Effects of Nickel and Urea on Growth, Nutrient and Nitrate Content of Hydroponic-cultivated Water Spinach
(Ipomoea aquatica cv. Taoyuan No. 1)
謝博中 Po-Chung Hsieh
指導教授:陳右人 博士 Advisor: Iou-Zen Chen, Ph.D.
中華民國 109 年 2 月
Feb, 2020
致謝
本論文得以完成,首先感謝業師陳右人教授自大學時期以來的悉心指導,奠 定了頗為前瞻的方向,使本題目於大學專題課程時即有初步成果;且在後續的試驗 設計規畫方面,也讓我有一定程度的發揮空間,因此進一步擴展而成本論文的落 實。陳老師不僅於百忙之中對拙著內容的不斷斧正,常常在本研究遇到困難時指點 迷津與協助鼓勵;其身教言教更讓我在日常生活、為人處事以及研究能力上獲益良 多。
試驗進行的過程當中,也得到許多人的幫忙:特別是來交流的鍾鳳林老師,
除了給予寶貴的試驗規畫思維,也提示要注意蕹菜材料的特性;還在我出國的時候 為水耕箱補水,使開頭的試驗得以順利完成。另得感謝在多次的研究室meeting 中,
阮素芬老師及林書妍老師於試驗各階段報告的指正以及建議。還有在初期階段,我 對植體營養分析不甚熟悉,幸蒙美玲姐以及懷慶學長的耐心教導,以及茶業改良場 研究人員戴佳如、李建漢的協助操作分析,遂得以一一克服難關。
再者,還得感謝研究室黃翔及俊陵學長的愛護,同窗十方三年來常交換經驗 資訊,一起從錯誤中學習、解決問題以及過程的互相鼓勵。並有 Billy、亭瑋共同 處理擔任課程助教的大小事,珮淳以及瑞娟、雙君的鼎力協助,還有曾玲蓉阿姨的 照顧。由於本研究室成員總是甘苦與共,共同克服了許多難題,也一起經歷了很多 充實快樂的時光;歷歷在目的回憶不僅充滿感激,也覺得有參與的榮譽感,因為我 是其中的一份子。
最後,在碩士路途的尾聲,感謝口試委員李金龍教授、林慧玲教授等五位老 師,撥冗精密審查論文,惠賜寶貴修改意見。另外,還要感謝父母的養育之恩,因 為家人無微不至的照顧與精神上的支持,始終是我的堅強後盾,使我能專心致志於 學業上的精進。
謹以此論文研究成果獻給所有關懷我的師長、父母、家人以及朋友,更期望能 將所有感恩之心,化做日後貢獻所學的動力。
摘要
尿素雖為農業主要施用之氮肥,但因尿素用於水耕時氮同化效率較差,鮮少 作為水耕氮源,此可能與配方未考慮鎳添加有關。作物對尿素同化過程需經脲酶 (urease)水解成銨態氮後得以利用。鎳為脲酶之中心離子,故鎳可能改善作物對養 液尿素的同化效率。以尿素取代養液部分硝酸氮源,可降低生產蔬菜之硝酸鹽含 量,避免其過量攝入之風險。本研究特別以鎳之添加,重新評估含尿素蔬菜水耕養 液配方之開發,期望運用此配方生產低硝酸鹽蔬菜。‘桃園一號’蕹菜(Ipomoea aquatica)以湛水式水耕系統進行栽培,養液參考 Hoagland 配方。總氮維持不變,
以尿素取代部分硝酸鹽並輔以鎳之添加,探究含尿素水耕配方對蕹菜之生長、要素 含量以及硝酸鹽含量之影響。本研究共分為四個試驗,試驗一為7 mM 尿素態氮搭 配0 至 85.3 μM 共六鎳變級之鎳毒害試驗。水耕液過量鎳(> 8.6 μM)抑制蕹菜生長,
產生新葉黃化徵狀,蕹菜葉片鐵含量較低,顯示過量鎳除本身造成毒害,可能阻礙 蕹菜對鐵之吸收,並占據以鐵為中心之酵素,造成類似缺鐵之徵狀。試驗二則改以 0 至 4 μM 共五個鎳變級與兩個尿素變級(0, 8 mM)以求得適宜鎳添加範圍。0.5、1 μM 鎳之添加,可改善含尿素水耕蕹菜的生長,且尿素的存在,使蕹菜有較高根部 鎳累積。試驗三則以0 至 10 mM 共七個尿素變級及適量鎳之添加(0、1 μM),來探 討尿素取代硝酸態氮的程度對蕹菜之反應。高尿素濃度下,蕹菜仍可正常生長。適 量鎳添加可提升6、8 mM 尿素組之生長,且蕹菜地上部硝酸鹽含量較無尿素對照 下降21%。高尿素下蕹菜鈣、鐵及鋅含量較低,但錳含量則較高。其中蕹菜根部鐵 含量於高尿素下,降低幅度較大,此應為蕹菜應用於含尿素養液與傳統養液栽培之 間的差異。於相同適量鎳下(1 μM),蕹菜根部鎳含量與養液尿素濃度成正相關,與 試驗二之結果呼應。試驗四則以三個尿素變級(2、4、10 mM)與鎳添加(0、1 μM)來 探討蕹菜,吸收尿素態氮、銨態氮及硝酸態氮之情形。鎳添加可提升蕹菜對養液尿 素態氮之吸收速率,並降低銨態氮之吸收速率,鎳添加則僅降低第一週蕹菜對硝酸 態氮之吸收速率。蕹菜於高尿素養液下仍能正常生長,且所生產的蕹菜硝酸鹽含量 顯著較低,表示含尿素水耕配方具一定應用潛力,蕹菜對鎳需求與尿素之存在有 關,並適量鎳可改善6、8 mM 尿素組下之生長,增進蕹菜對養液中尿素的吸收,
顯示含尿素水耕配方之開發,鎳為配方中扮演不可或缺之必要關鍵成分。
關鍵字: 尿素、蕹菜、水耕氮源、脲酶、尿素同化
Abstract
Urea is the most widely used and low-cost nitrogen fertilizer in agriculture.
However, it is seldom used as a nitrogen source in hydroponic culture due to its poor nitrogen use efficiency. These results could be related to the lose sight of Ni supplement.
Plants utilized urea via urease to hydrolysis it into ammonia for further assimilation process. Ni is of importance for activation of urease and hence for promoting plants to utilize urea in urea-based nutrients. Partially replacing nitrate with urea in hydroponic culture could reduce nitrate content in vegetable crops and reduce the risk of excessive nitrate intake. Therefore, the objective of this study was to develop urea-containing nutrient formula especially in view of Ni addition. We hope this nutrient formula could be applied on low-nitrate vegetable crops production. In order to investigate the effect of Ni on urea-based solution on plant growth, nitrate and nutrient content. Water spinach (Ipomoea aquatic cv. Taoyuan No. 1) were grown in deep flow technic system (DFT) with nutrient based on Hoagland solution. NO3-N were partially replaced with urea-N, while the total-N were the same. The research was divided into four experiments. In the 1st experiment, we used six Ni concentration (0-85.3 μM) with 7 mM urea-N to observe the Ni toxic effect on water spinach. Excess Ni (> 8.6 μM) inhibited plant growth. Young leaves had chlorosis or necrosis symptoms and Fe content in leaf were aslo decreased. Ni might affect Fe uptake and occupy Fe-related enzyme, and cause symptoms that similar
to Fe deficiency. Then in the 2nd experiment, five Ni concentration (0-4 μM) with two urea-N level (0, 8 mM) were used to find the suitable Ni dosage in nutrients. With appropriate Ni addition (0.5, 1 μM), plant growth were improved and the Ni content in roots were higher in the presence of urea. In the 3rd experiment, seven urea levels (0-10 mM) combined with suitable Ni dosage (0, 1 μM ) to evaluate the degree of substitution urea for nitrate on plants response. Water spinach could growth well under high urea-N dosage (10 mM). Ni addition could increase water spinach growth under 6, 8 mM urea- N and drcreased nitrate content 21% compared to no urea control. The content of Ca in whole plant, Fe and Zn in root decreased under high urea dosage, while Mn in roots increased under high urea dosage. Among these nutrients, root Fe content significantly declined as urea dosage increased, which showed the distinct difference compared to traditional no-urea formula. Under the same 1 μM Ni addition, root Ni content in plants were positively correlated to urea concentration in nutrients. These trend were in agreement with the result of the 2nd experiment. The 4th experiment was to monitor the absorption of urea-N, NH4-N and NO3-N from nutrients. Nutrients contained three levels of urea-N (2, 4, 10 mM) and two levels of Ni (0, 1 μM). Ni addition significantly increased water spinach urea-N uptake rate and decreased NH4-N uptake rate. While Ni addition only decreased NO3-N uptake rate in the first week under Ni addition. Water spinach could grow well and effectively reduce nitrate content in shoots under high urea-N
dosage. These advantages provided some potential of applying this urea-based formula on water spinach. Appropriate addition of Ni promoted water spinach urea absorption and improve growth in 6, 8 mM urea-N. The demand of Ni for water spinach was also higher in the existence of urea. These evidences demonstrated the essentiality of Ni on designing a urea-based hydroponic culture system.
Key word: urea, water spinach, hydroponic nitrogen source, urease, urea assimilation.
目錄
前言 ...1
前人研究 ...3
一、尿素氮肥於農業上之重要性 ...3
二、植物對尿素吸收及同化 ...4
三、鎳對以尿素栽培的作物之重要性 ...5
四、植物對鎳與鐵鋅吸收之影響 ...7
五、鎳毒害對作物外觀生長及要素含量之影響 ...8
六、其他氮源對尿素吸收之影響 ...10
七、尿素取代硝酸鹽配方之開發與低硝酸鹽蔬菜之生產 ...11
八、以熱帶作物蕹菜評估含尿素及鎳之水耕養液配方可行性 ...13
材料與方法 ...15
一、植物材料、水耕系統及養液配方 ...15
二、試驗方法與調查項目 ...15
三、分析方法 ...17
四、統計分析 ...22
結果 ...23
一、鎳毒害對‘桃園一號’蕹菜生長與要素含量之影響(試驗一) ...23
二、適宜鎳對含尿素水耕‘桃園一號’蕹菜生長及要素含量之影響 (試驗二) ...25
三、尿素濃度與鎳添加對水耕‘桃園一號’蕹菜生長及要素含量之影響 (試驗三) .29 四、尿素濃度與適量鎳對水耕‘桃園一號’蕹菜氮源與巨量要素吸收之影響 (試驗 四) ...31
討論 ...34
一、蕹菜鎳毒害病徵與鎳累積 ...34
二、適量鎳添加之必要性 ...36
三、含尿素及鎳養液配方對蕹菜要素含量之影響 ...37
四、含尿素養液配方評估與建立 ...38
結論 ...41
參考文獻 ...43
圖 ...53
表 ...64
附錄 ... 85
圖目錄
圖1、養液 85.3 μM Ni 濃度處理對蕹菜鎳毒害徵狀 ...53
圖2、鎳毒害對蕹菜生長及葉片外觀之影響 ...54
圖3、養液鎳濃度與水耕蕹菜葉片鎳含量之相關性 ...55
圖4、養液鎳與尿素對水耕蕹菜根部鎳含量相關性 ...56
圖5、養液尿素與鎳對水耕蕹菜株高之影響 ...57
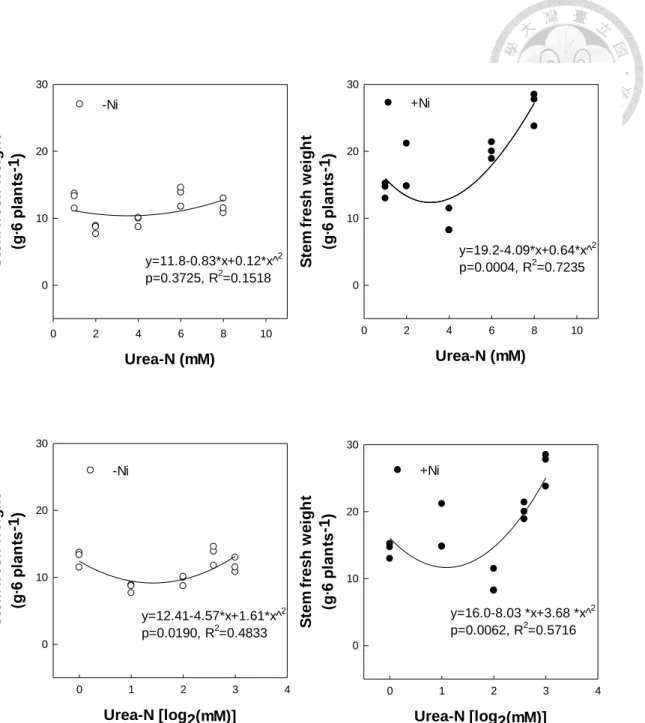
圖6、養液尿素鎳添加對水耕蕹菜莖部鮮重之影響 ...58
圖7、養液尿素與鎳處理對水耕蕹菜莖部鮮重迴歸分析 ...59
圖8、養液尿素與鎳對水耕蕹菜地上部硝酸鹽含量之影響 ...60
圖9、養液尿素與鎳對水耕蕹菜根部鎳累積之影響 ...61
圖10、尿素與鎳添加養液配方對水耕蕹菜株高之影響 ...62
圖11、尿素與鎳添加對養液校正前 pH 值之影響 ...63
表目錄
表1、鎳對以尿素佔 46.6%氮源之水耕蕹菜葉片葉綠素濃度、葉長及莖徑之影響
...64
表2、鎳對以尿素佔 46.6%氮源之水耕蕹菜株高之影響 ...65
表3、鎳對以尿素佔 46.6%氮源之水耕蕹菜株高、莖徑及乾鮮重之影響(26 天) .66 表4、鎳對以尿素佔 46.6%氮源水耕蕹菜根莖葉乾鮮重之影響 ...67
表5、鎳對以尿素佔 46.6%氮源之水耕蕹菜葉片要素含量之影響(26 天) ...68
表6、養液中鎳濃度與尿素對水耕蕹菜葉綠素濃度與生長之影響 ...69
表7、養液中鎳濃度與尿素濃度對水耕蕹菜乾鮮重單因子分析 ...70
表8、養液中鎳濃度與尿素對水耕蕹菜乾鮮重之複因子分析 ...71
表 9、養液中鎳濃度與尿素對水耕蕹菜單株要素含量之影響 ...72
表 10、養液中鎳與尿素濃度對蕹菜葉片要素含量之影響 ...73
表 11、養液中鎳與尿素濃度對蕹菜莖部要素含量之影響 ...74
表 12、養液中鎳與尿素濃度對蕹菜根部要素含量之影響 ...75
表13、養液尿素濃度與鎳添加對水耕蕹菜葉綠素濃度、株高、莖徑以及根長之影 響 ...76
表14、養液尿素濃度與鎳添加對蕹菜乾鮮重之影響 ...77
表15、養液尿素濃度與鎳添加對蕹菜單株要素含量之影響 ...78
表16、養液尿素濃度與鎳添加對蕹菜葉片要素含量之影響 ...79
表17、養液尿素濃度與鎳添加對蕹菜莖要素含量之影響 ...80
表18、養液尿素濃度與鎳添加對蕹菜根要素含量之影響 ...81
表19、尿素與鎳對水耕蕹菜乾鮮重之影響 ...82
表20、尿素與鎳對水耕蕹菜氮肥吸收量之影響 ...83
表21、尿素與鎳對水耕蕹菜巨量要素吸收量之影響 ...84
前言
尿 素 是 目 前 最 廣 為 使 用 的 氮 肥 , 占 整 體 全 世 界 氮 肥 消 耗 量 的 一 半 (http:/faostat.org)。尿素價格相對便宜、易溶於水且方便施用,含氮量高(46.6%),較 硝酸態氮肥便於運輸且較無爆炸風險(Glibert et al., 2006)。尿素施於土壤被視為緩 效型有機態氮肥,施於土壤後需經土壤微生物脲酶(urease)作用分解成銨態氮,才 得以被植物及微生物利用;但是尿素施於土壤被水解成銨態氮過程中,會使土壤 pH 值升高,而銨態氮於此高 pH 值的鹼性環境下,就容易轉變回氨氣揮發逸失 (ammonia volatilization) (Ernst and Massey, 1960)。此氨氮揮發逸失之現象,若在只 將尿素灑施於表土、未混拌入土壤中,或者將尿素施用於鹼性土壤等狀況下會更加 劇,其流失率可達3 成以上,造成施用上的浪費損失(Silva et al., 2017)。
尿素不只是應用於土施氮肥,也可用於水耕養液,作為傳統配方中,硝酸態氮 以及銨態氮之外的第三種氮肥來源。養液氮源以尿素取代硝酸鹽的使用,可以降低 水耕蔬菜之硝酸鹽含量(Khan et al., 1999),此為含尿素水耕養液之優點。由於蔬菜
於栽培上,常有硝酸鹽的累積問題,且蔬菜占飲食硝酸鹽攝入的 50%至 70%
(Habermeyer et al., 2015);因此降低蔬菜硝酸鹽含量,即可解決大部分硝酸鹽之攝 取風險。歐盟根據季節、品種與栽培方法,曾訂定不同蔬菜硝酸鹽含量之上限規定,
其中針對嬰兒有特別低的硝酸鹽含量上限(EC No 466/2001)。
降低蔬菜硝酸鹽的累積問題,除了選擇低硝酸鹽累積品種、改善栽培環境之 外,也可以從改變氮肥種類組成來達成(Anjana et al., 2007)。若以尿素取代養液硝 酸鹽的使用,應該具有生產低硝酸鹽水耕蔬菜之潛力,加上尿素用於養液配方養液 pH 值調整容易,可將 pH 值保持於 5.5-6.0 之間,與施用於土壤方式相比較,就較 無氨氣逸失問題。
相當多研究將尿素作為水耕氮源,但大多成果認為尿素並非理想水耕氮源,造 成尿素鮮少用於水耕養液。於水耕環境以尿素作為氮肥,與硝酸態氮及銨態氮相
比,番茄對尿素吸收及同化效率較低,植株生長也不佳(Ikeda and Tan, 1998);玉米 與小麥同樣以硝酸銨有較高的氮肥施用效率(nitrogen use efficency),而尿素則較低 (Merigout et al., 2008)。其原因應該是植物對尿素代謝路徑受阻。由於尿素經由水 耕養液吸收至植物體內,需以存在於細胞質的脲酶(urease)將尿素分解成銨態氮及 二氧化碳;銨態氮進一步被植物所利用,同化為胺基酸以供植物所需(Witte, 2011)。
因此,脲酶的存在與否及活性,即為尿素利用效率之關鍵。
鎳為植物第十七個必要元素,目前唯一之機能是作為脲酶的中心離子,可將精 胺酸(arginine)代謝過程所產生的內生尿素順利水解成銨態氮,進而形成胺基酸 (Eskew et al., 1983)。所以在含尿素的水耕養液系統加入鎳,可改善含尿素水耕栽培 作物之生長(Gerendas et al., 1998; Khan et al., 1999)。
鎳可改善植物對尿素代謝之功能研究雖得到肯定,但由於大家已受限於早先 尿素作為水耕氮源效率不彰的說法,所以現今大多仍沿用舊有未含鎳與尿素之養 液配方;且作物種子內所含的鎳足以提供未含尿素配方所需,最終造成鎳添加的必 要性被遺忘。本研究以鎳之添加,重新評估尿素作為水耕氮源之可能性,並期望藉 此研究能再次彰顯鎳於水耕配方中的必要性,並藉由適當控制鎳添加之濃度,使所 栽培之蔬菜鎳累積含量既符合食品安全之要求,又可免除硝酸鹽過量累積於蔬菜 對人體健康之風險。
本研究使用蕹菜作為試驗材料,養液配方以尿素取代部分硝酸鹽,並加入適當 濃度的鎳,在總氮濃度不變的方式下來評估對蕹菜生長、必要元素及硝酸鹽含量之 影響。期望求得水耕環境最適蕹菜的尿素及鎳濃度,以及鎳添加於此系統所扮演的 角色,並透過結果可作為實際應用策略上之參考。
前人研究 一、尿素氮肥於農業上之重要性
尿素化學式為(NH2)2CO,分子量 60.0。產業上尿素的製程是以哈伯法製氨 (Haber-Bosch process)取得液氨,通入二氧化碳即得尿素;液氨進一步可生產硫酸 銨及硝酸態氮肥。其中,硝酸態肥料則主要以奧士華-哈伯法(Haber-Ostwald)製成,
製程上較尿素成本高(Barnum, 2003)。尿素為農業用氮肥中含氮量最高者(46.6%),
在施用相同氮素下,尿素所需重量較其他氮肥少。於肥料儲藏及運輸上尿素也較方 便安全,硝酸態氮肥如硝酸銨則有儲藏環境不良產生塵爆之風險(Glibert et al., 2006)。尿素極易溶於水,在 20℃下溶解度為 108 g∙100 mL-1,除了施用於土壤,也 方便配製成液肥以及應用於葉面施肥。以上特點使尿素成為現今之主流氮肥。
尿素被視為緩效型有機態氮肥,施於土壤後需經土壤微生物脲酶(urease)分解 成銨態氮後得以被植物及微生物利用,然而於土壤分解產生的銨態氮則易於轉變 成氨氣揮發逸失(ammonia volatilization);尿素施於土壤後,經土壤微生物脲酶的作 用,水解成兩分子的氨及一分子之二氧化碳,而氨氣則與土壤溶液中氫離子作用轉 換成銨離子,此步驟會造成土壤pH 值的升高,而銨態氮於鹼性土壤環境則易行逆 反應轉變回氨氣揮發損失(Jones, 2013)。溫度、土壤含水量、土壤 pH 值、施用深 度,皆影響到土施尿素轉變成氨氮佚失之程度;環境溫度越高,適中的土壤含水量、
土壤pH 值越高,以及尿素施用深度越淺,皆加速尿素於土壤中水解,並轉變成氨 氣逸失(Ernst and Massey, 1960)。施用尿素後充足的降雨,則可將尿素淋溶至土壤 深層,降低氨氣揮發(Black et al., 1987)。土施尿素於在鹼性土壤或施用於表土後,
如無灌溉淋溶入土壤深處,易被分解成氨氣逸失,可高達 3 成以上(Silva et al., 2017)。市售尿素氮肥以添加脲酶抑制劑(N-(n-butyl) thiophosphoric triamide;NBPT),
來延緩尿素施於土壤之揮發氨氮(AGROTAIN®)。脲酶抑制劑因與尿素結構類似,
是 以 占 據 脲 酶 與 尿 素 反 應 部 位 來 抑 制 脲 酶 活 性 , 屬 競 爭 型 抑 制(competitive inhibition) (Mazzei et al., 2017)。脲酶抑制劑 NBPT 與尿素混合於土壤可顯著降低
由尿素造成的氨氮逸失;但脲酶抑制劑也可被作物吸收體內,影響作物對尿素的吸 收與同化,對作物生長有不良影響(Cruchaga et al., 2011; Krogmeier et al., 1989)。
二、植物對尿素吸收及同化
尿素分子量小且為非電解質,早期認為尿素是以擴散方式通過植物細胞膜進 出細胞(Gallucci et al., 1971)。Wilson 等(1988)以14碳標定尿素搭配ATPase 抑制劑,
來探討綠藻對尿素吸收,發現跨膜氫離子濃度梯度的存在,可促進尿素吸收,顯示 植物可利用主動運輸方式調控尿素之吸收。植物對尿素吸收與外界尿素濃度有關,
依照外界尿素濃度的高低可分為高親和性運輸系統(high affinity transport system)及 低親和性運輸系統(low affinity transport system)兩種不同途徑進入細胞。
阿拉伯芥中的AtDUR3 被認為是專一性高的尿素主動運輸蛋白,在 ATPase 參 與造成細胞膜內外氫離子濃度梯度差下,產生質子動力勢 (proton motive force),
而後尿素與氫離子得以透過運輸蛋白協同運輸(cotransport)進入植物細胞,Km值為 3-4 μ M (Liu et al., 2003a)。阿拉伯芥於缺氮環境下,於根部表皮細胞大量表現 AtDUR3 運輸蛋白以獲取外界低濃度尿素,AtDUR3 運輸蛋白也表現於根部木質部 及地上部,將尿素從根部經維管束運輸至地上部(Kojima et al., 2007)。當外界尿素 濃度較高時,尿素的吸收則為LAT 路徑;尿素除了經由運輸蛋白行主動運輸,也 可 經 水 通 道 蛋 白 (aquaporins) 、 TIP (tonoplast intrinsic proteins) 及 PIP (plasmamembrane intrinsic proteins)等通道蛋白,行低親和性之運輸,將外源尿素吸 收至根部細胞或將細胞質內的尿素運輸至液胞中儲存(Liu et al., 2003b)。
尿素除了可從外界環境獲得,尿素本身也為植物次級氮素代謝—鳥胺酸循環 (ornithine cycle)的中間產物;鳥胺酸循環中,精胺酸(argnine)於粒線體基質中因精 胺酸分解酶(arginase)作用而產生鳥胺酸以及內生尿素,內生尿素則運輸至細胞質,
並由存在於胞質的脲酶(urease)分解成銨態氮以及二氧化碳(附圖 1)。而銨態氮則進 一步透過於胞質及葉綠體中的麩醯胺酸合成酶(glutamine synthetase)合成麩醯胺酸 (glutamine),麩醯胺酸可作為主要運移用氨基酸,或藉由一系列轉氨酶,將有機酸
加上氨基轉變成天門冬醯胺(asparagine)運移,並隨後可同化為不同的胺基酸,作為 植物合成所需蛋白質之原料(附圖 2)。內生尿素及脲酶廣泛存在於植物各部位之細 胞中,鳥胺酸循環更是生物體不可或缺的代謝途徑,外源尿素同樣是以此路徑代謝 尿素,其中存在於胞質的脲酶則扮演關鍵角色(Merigout et al., 2008)。
尿素可於根部進行同化,以5 mM 尿素栽培之大豆根部銨態氮含量為地上部 銨態氮的10 倍左右,添加 NBPT 脲酶抑制劑抑制大豆根部及地上部脲酶活性,但 銨態氮含量只有根部受抑制,地上部則無,顯示大豆應主要於根部而非地上部同化 尿素(Cruchaga et al., 2011)。尿素也可隨蒸散流運輸至地上部進行同化,番茄幼苗 期吸收之尿素可於24 小時內迅速運移到地上部葉片進行同化,占總吸收 94%;尿
素同化能力也與生長階段有關,番茄生殖生長期尿素於 24 小時內葉片同化能力
(30%)較幼苗期(6%)高(Tan et al., 2000)。
尿素施用過多易造成累積毒害,葉尖及葉緣焦枯為尿素累積毒害常見徵狀,
尿素混合脲酶抑制劑施用會使作物葉緣產生壞疽情形,壞疽組織具高含量尿素,顯 示此壞疽為尿素累積所造成的毒害病徵(Krogmeier et al., 1989)。欲降低尿素於栽培 中毒害之情形,可從尿素於植物氮同化過程窺知,脲酶為代謝尿素之關鍵酵素,如 提升脲酶活性,應可改善以尿素氮肥栽培作物之生長。
三、鎳對以尿素栽培的作物之重要性
尿素被植物吸收後同化成胺基酸之過程,需要脲酶的參與,將其分解為氨及 二氧化碳。Dixon 等(1975)將白刀豆(Canavalia ensiformis)之脲酶純化後進行吸收光 譜分析,推測脲酶內含有鎳離子。Brown 等(1987)指出經缺鎳培養三個世代之大麥 種子,在無鎳養液下發芽受抑制,適當鎳可活化脲酶,種子儲存之精胺酸於發芽時 可順利同化成銨態氮供發芽所需;脲酶為精胺酸代謝過程必要酵素,如無鎳活化脲 酶,精胺酸代謝產生的內生尿素無法順利同化而使大豆發芽受阻。大豆種子主要以 精胺酸的形式將氮儲存於種子,約佔種子氮含量的17.3% (van Etten et al., 1967)。
大豆種子發芽時添加鎳,可增加25%的產量(Kutman et al., 2014)。大豆於無鎳 2.5%
尿素養液產生葉尖焦枯徵狀為尿素代謝受阻之故,鎳可活化脲酶之功能使得鎳被 認為是植物第17 個必需元素(Eskew et al., 1983)。
鎳 除 了 活 化 脲 酶 , 也 可 活 化 與 植 物 共 生 根 瘤 菌 之 鎳 鐵 氫 化 酶([NiFe]
hydrogenase),鎳添加不只增進大豆脲酶活性,也提高其共生固氮菌(Rhizobium japonicum 122 DES)氫化酶之活性(Klucas et al., 1983)。根瘤菌固氮過程中,每一分
子N2透過固氮酶(Nitrogenase),加上 8 個氫離子及 8 個電子,產生 2 分子 NH3以 及1 分子 H2,鎳鐵氫化酶可將固氮後多產生氫氣加以回收,並獲得能量,加速根 瘤菌固氮效率(Phillips, 1980)。
適量鎳之添加可活化脲酶,在水耕的狀態下,可改善以給予尿素之作物的生 長。養液添加0.5 μM 鎳可提升水稻脲酶活性,在有外源尿素(6 mM)下,鎳可提高 稻米的地上部及地下部之乾鮮重,不含尿素之硝酸銨組則無此改善效果(Gerendas et al., 1998)。尿素占總氮供給量 20%之狀況下,0.05 mg∙L-1鎳添加,使水耕菠菜有 較佳乾鮮重及葉長(Khan et al., 1999)。水耕萵苣於三種尿素濃度(5、10、20 mM)配 合鎳添加皆提升脲酶活性,使尿素被吸收後得以順利代謝,改善尿素組葉片鮮重及 葉片總氮含量;而硝酸銨組添加鎳,較無提升總氮含量之效果(Khoshgoftarmanesh et al., 2011)。
傳統養液配方因發展時期較早,如 Hoagland 養液配方於二戰後即提出 (Hoagland and Arnon, 1950),故配方中微量元素部分並未考慮到鎳添加。而隨後各 種修正配方,如Steiner 配方加強除硼外之微量元素濃度,主要改變為巨量元素之 組成,以及鐵螯合劑之種類(Steiner, 1961)。鎳於 1980 年代以後才被加入必要元素 的行列(Brown et al., 1987),時間晚於傳統養液配方建立時期,所以未考慮到鎳添 加,養液組成也只有銨態氮及硝酸態氮兩種。而由於鎳需求很低,水體及植體所含 的鎳,已能滿足代謝內生尿素所需,故水耕作物絕大多數均能正常生長。近年來水 耕蔬菜硝酸鹽含量備受重視,本研究欲以尿素取代硝酸鹽使用,以降低食用風險,
因此鎳之添加可能是成功之關鍵因子。
四、植物對鎳與鐵鋅吸收之影響
鐵、鋅、與鎳於週期表上同為過渡金屬,因電子組態牽涉d 軌域,從原子半徑 可得知Fe2+ (82 pm),、Zn2+ (83 pm)、Ni2+ (78 pm)三者十分相近(Emsley, 1991),根 部吸收上可能有共用運輸蛋白之情形。金屬離子進入根部後,雖可經由共質體運輸 (symplast)、質外體運輸(apoplast)以及跨膜運輸(transmembrane)的方式,逐漸往根部 維管束組織運送,但以質外體運輸為主,因卡氏帶(Casparian strip)限制金屬離子只 以共質體運輸方式通過,因此質外體運輸要轉為共質體運輸,並需要經過跨膜運輸 (Kim and Guerinot, 2007)。跨膜運輸需經細胞膜上的運輸蛋白,在金屬離子的吸收 調控上扮演重要角色。
ZIP family (ZRT、IRT-like Protein)為與鐵及鋅運輸蛋白相似的運輸蛋白家族。
蛋白質結構預測顯示ZIP family 在膜上有 8 個跨膜結構域(transmembrane domain),
其中第 3 及第 4 跨膜結構域之間的變異區胺基酸序列,與該運輸蛋白之專一性有 關(Guerinot, 2000)。ZIP family 中的 IRT1 鐵運輸蛋白存在於阿拉伯芥根部之皮層細 胞及內皮細胞,IRT1 所屬基因於缺鐵時,表現於根部外圍組織以及根毛。IRT1 鐵 運輸蛋白的功能為在缺鐵環境下,將鐵主動運輸至細胞內的鐵運輸蛋白;然而IRT1 的專一性不高,在缺鐵的環境下,如果環境有其他過渡金屬離子如鎳、鋅、錳及鈷,
也會於細胞內有相對高的累積量,表示這些金屬離子也可經IRT1 主動運輸進入細 胞,造成最終累積的情形(Vert et al., 2002)。鎳二價離子同樣也可經由 IRT1 進入根 部細胞;以(dimethylglyoxime;DMG)藥劑染色,在缺鐵與添加鎳之處理,阿拉伯 芥野生型的根部有大量鎳累積,可高達200 ppm,約為鐵充足下的 4 倍累積量,而 IRT1 突變株則無此現象(Nishida et al., 2011)。鎳與鐵之吸收因鐵運輸蛋白專一性不 高,可能共用鐵運輸蛋白進出細胞,而使兩元素在吸收上會互相影響。
鎳可於阿拉伯芥根部細胞累積大量濃度,與液泡運輸蛋白有關。AtIREG2 運 輸蛋白mRNA 同樣於缺鐵環境表現於根部,並透過綠螢光蛋白報導基因 GFP 標記 顯示AtIREG2 位於阿拉伯芥根部細胞液胞上。AtIREG2 過量表現株可在高鎳環境 (200 μM)下保有較高鮮重,過量表現株相較於野生株的鎳累積也較高,而野生型則 無法在此高濃度鎳下良好生長;於缺鐵下添加鎳,過量表現株較野生株生長良好,
而AtIREG2 突變株則根部及地上部生長皆受抑制。AtIREG2 為位於液胞上可以調 節細胞質鎳濃度的動態平衡,進而與鎳的耐受性有關(Schaaf et al., 2006)。專一性
不高的鐵運輸蛋白,搭配AtIREG2,可將缺鐵時過量吸收的鎳,從細胞質主動運輸 至液胞儲存,進而減低細胞質內過高鎳濃度的傷害,並造成鎳會於缺鐵下累積於根 部細胞的結果。
鎳可經由 IRT1 鐵運輸蛋白進入細胞,但 IRT1 突變株仍有鎳累積,應有獨立 於IRT1 的運輸蛋白參與鎳之運輸。ZIP1、ZIP2 與 ZIP3 與鋅有關運輸蛋白與 IRT1 結構相近(Grotz et al., 1998),且皆同屬 ZIP family 成員,這些運輸蛋白也可能為鎳 吸收之管道。阿拉伯芥於缺鋅環境下,與缺鐵狀況相似,同樣於根部有累積鎳的現 象,同位素 63Ni 添加於缺鋅的阿拉伯芥植株,配合缺鋅養液處理,於短時間有明 顯較高同位素63Ni 累積量;IRT1 突變株於缺鋅下,雖然相較於野生型於缺鋅下有 較低之鎳累積,但仍相較於鋅充足的處理有較高鎳累積,代表有獨立於IRT1 之運 輸蛋白會於缺鋅下,將鎳主動運輸至根部細胞內,造成鎳累積之結果(Nishida et al., 2015)。鎳可不只經鐵運輸蛋白進出細胞,也可能有其他 ZIP family 運輸蛋白參與,
進而影響到其他原子半徑相似之元素吸收。
五、鎳毒害對作物外觀生長及要素含量之影響
鎳為地殼前20 豐富之元素,土壤鎳含量從 10-1000 mg∙Kg-1不等,其中以蛇紋 岩化育之土壤含有較高鎳含量(Gasser and Dahlgren, 1994)。一般土壤環境較不會有 鎳缺乏問題。鎳添加於養液常見方法為添加七結晶水硫酸鎳,提供植物吸收有效性 佳的二價鎳。鎳添加不可造成作物過度鎳累積,以免造成食品安全疑慮。鎳為對人 體有害之重金屬,我國法規《飲用水水質標準》(民國106 年 1 月修訂)訂定飲用 水所含的鎳最大限值為0.2 mg·L-1。美國環境保護署(U.S. EPA)建議,飲用水鎳濃 度應不超過0.1 mg·L-1,每日容許攝取量(Tolerable Daily Intake;TDI)為 0.02 mg·
Kg-1·day-1,每人每日攝入低於此量,則終身無受毒害之虞,以60 公斤重成人為例,
每日鎳可容許攝取量為1.2 mg。
鎳以二價離子形式被根吸收,除了如生長於南非蛇紋岩土壤上Berkheya coddii 鎳超累積植物可將大量鎳蓄積於地上部之外(Robinson et al., 1997),一般作物如玉 米、黑麥(Yang et al., 1997)、大豆(Dalton et al., 1985)、萵苣及番茄(Poulik, 1999)等,
多將鎳累積於根。運移進維管束的鎳可被有機酸或胺基酸螯合,促進鎳於植物體內
的運輸(Cataldo et al., 1988)。少量鎳可進入維管束而輸送至地上部,根與地上部之 鎳濃度可相差近70 倍;組胺酸(histidine)可與鎳形成 1:1 錯合物,在水耕液搭配鎳 施用增加小麥鎳根共質體運輸之比例(Symplastic Ni /Apoplastic Ni),並使小麥木質 部鎳含量增加,促進鎳從根傳輸到地上部(Dalir and Khoshgoftarmanesh, 2014)。鎳 經維管束傳輸到地上部後,會累積於新生部位,大豆在含鎳栽培下,新葉比老葉的 濃度高6 至 12 倍(Kutman et al., 2014) 。番茄將鎳吸收至地上部,有 67.0-77.3%分 佈至植株頂端(Poulik, 1999)。如欲食用含鎳蔬菜,應去除根部以減少攝取在根部累 積的鎳。
一般作物利用將鎳限制於根部,減少運移至地上部的方式,來忍受鎳含量較 高之環境(Seregin and Kozhevnikova, 2006),當環境鎳含量高於一定閾值,地上部鎳 含量即開始上升,大豆地上部鎳含量於養液鎳超過 50 μM 後迅速增加(Cataldo et al., 1978),而一般作物地上部鎳含量高於 20-50 mg·Kg-1D.W.門檻濃度,即開始產 生鎳毒害徵狀(Davis et al., 1978; Macnicol and Beckett, 1985)。
鎳如同其他重金屬具有和酵素蛋白質上巰基結合能力(SH-group binding)可抑 制許多酵素活性,且可干擾電子傳遞鏈,產生氧化逆境以及自由基,對細胞造成傷 害(Toppi and Gabbrielli, 1999)。200 μM過量鎳處理之小麥超氧歧化酶SOD及過氧化 氫酶CAT活性升高,過氧化物酶POD及穀胱甘肽轉移酶GST等過氧化物清除酵素活 性下降,使植株處於氧化逆境,進而抑制地上部生長以及造成葉綠素含量下降 (Gajewska et al., 2006)。鎳離子也可能取代其他酵素中金屬離子之位置,特別是與 鎳離子大小相似之金屬離子,導致該酵素功能喪失。鎳可取代葉綠素中的鎂離子,
以及Rubisco酵素之鎂輔因子,影響光合作用量以及速率(Yusuf et al., 2011)。芥菜於 100 μM鎳下根長受抑制33% (Alam et al., 2007),大麥於0.1 mM鎳處理14天也產生 葉片黃化壞疽現象(Rahman et al., 2005)。鎳毒害所造成的葉片黃化壞疽徵狀即可能 為鎳取代光合作用相關酵素中金屬輔因子所造成,而使得此徵狀與缺鐵、缺鋅、缺 鎂類似。
六、其他氮源對尿素吸收之影響
尿素、銨態氮、硝酸態氮為三種常用肥料氮源,然而其同化路徑並不相同。
尿素同化路徑與銨態氮較類似,尿素經細胞質脲酶轉換成銨態氮後,則後應與銨態 氮同化路徑相似,可直接合成胺基酸;而硝酸態氮則需先經兩階段還原成胺態氮,
分別經硝酸還原酶(nitrate reductase)以及亞硝酸還原酶(nitrite reductase),而亞硝酸 還原酶則存在於色素體(plastid),因此硝酸鹽同化過程必須經色素體參與,才能將 硝酸態氮順利還原成銨態氮,並隨後透過位於色素體之 GS/GOGAT(glutamine synthetase/glutamine oxoglutarate aminotransferase)進一步合成胺基酸(Merigout et al., 2008; Witte, 2011)。
尿素不適合單獨作為養液氮源,如同傳統養液氮源所包含的銨態氮及硝酸態 氮。單獨使用尿素之水耕番茄生長較差,而以尿素配合硝酸態氮或銨態氮,可提高 番茄乾重及總氮含量(Ikeda and Tan, 1998)。水耕液同時含有硝酸態氮及尿素,可使 小麥幼苗對尿素吸收速率提升,而銨態氮則無提升尿素吸收速率;硝酸態氮及尿素 共存組小麥地上部有較高銨態氮含量而有較低尿素含量,水耕養液硝酸鹽的存在,
可以提升小麥幼苗對尿素之吸收,且尿素較快同化成銨態氮之速率(Garnica et al., 2009)。
Zanin 等(2015a)指出,硝酸鹽及尿素共存處理相對於其個別之處理,玉米幼 苗根部側根有較好發育;以 Real-time RT-PCR 觀察氮同化相關基因之表達,在硝 酸鹽及尿素共存處理下,細胞質之GS 基因 ZmGln1‐5 以及色素體 GS2 基因 ZmGln2 表達量提高,而與硝酸鹽同化相關基因如硝酸還原酶基因ZmNR 及亞硝酸還原酶 基因ZmNiR 也較單獨硝酸鹽處理組上升,顯示尿素與硝酸鹽共存處理具較佳氮同 化能力,存在於細胞質及色素體的兩種與氮素同化相關酵素皆上調,可增進尿素同 化效率。
色素體 GS 酵素可將光合產物與亞硝酸還原後產生的銨離子反應產生麩醯胺
酸(glutamine),經 GOGAT 產生麩胺酸(glutamate),麩胺酸則可運移至細胞質,透
過存在於細胞質的GS,作為其反應之受質,將尿素同化過程產生的銨離子整合產 生麩醯胺酸,而最終可同時提高尿素與硝酸鹽同化效率(Zanin et al., 2015b)。
七、尿素取代硝酸鹽配方之開發與低硝酸鹽蔬菜之生產
人體攝入硝酸鹽來源主要有肉製品、水及蔬菜,其中蔬菜占飲食硝酸鹽攝入 的85% (Güneş et al., 1994)。水耕蔬菜常有硝酸鹽含量過高問題,極易超過世界衛 生組織所訂定之每日攝取安全容許量(0-3.7 mg/Kg body weight/day)。嬰兒攝食超過 1-8.3 mg NaNO3 易產生高鐵血紅蛋白血症(methemoglobinaemia),或稱為藍嬰症 (blue baby syndrome)。
硝酸鹽在一定含量下本身沒有毒性,其為害主要是轉為亞硝酸鹽而表現;大
約 25%攝入之硝酸鹽會經唾腺再分泌回口腔,而經口腔細菌還原成亞硝酸鹽,約
6-7%攝入的硝酸鹽經此循環轉換成亞硝酸鹽被攝入 (Habermeyer et al., 2015)。亞 硝酸鹽與亞硝基前驅物作用後,可在人體中產生內生性氮-亞硝基化合 (N-nitroso compounds),經動物實驗證明為強烈致癌物。歐盟根據不同季節、品種、栽培方法 訂定不同蔬菜硝酸鹽含量之上限,其中針對嬰兒有特別低之硝酸鹽含量上限(附表 1)。
降低蔬菜硝酸鹽含量,即可解決大部分硝酸鹽之攝取風險。降低蔬菜硝酸鹽 的累積問題除了選擇低硝酸鹽累積品種、改善栽培環境,也可以從改變氮肥種類組 成來達成(Anjana and Iqbal, 2007)。在菠菜上,尖葉菠菜較圓葉菠菜有較高之硝酸 鹽累積量(Maynard et al., 1976)。800 µmol∙m−2∙s−1光照環境下,可有效降低硝酸鹽 含量(Proietti et al., 2004)。以尿素取代養液 20%硝酸鹽可降低‘Zaobao’ 及 ‘Lvbao’
芥藍之硝酸鹽含量(Zhu et al., 2018),以尿素取代養液 20%硝酸鹽分別降低‘Berlo’
及‘Kirsten’ 萵苣 14%及 11%硝酸鹽含量(Günes et al., 1994)。以 50%以及 20%尿素 取代硝酸鹽使用可分別降低菠菜41%以及 26% 硝酸鹽含量(Khan et al., 1999)。顯 示水耕蔬菜以尿素取代硝酸鹽使用,可有效降低蔬菜硝酸鹽含量,並且取代比例越 高,蔬菜硝酸鹽含量越低。
養液以尿素取代硝酸鹽,也影響作物生長以及必要元素之含量,尿素取代比 例因每種作物對尿素耐受性程度不一而不同。Luo 等(1993)以蕹菜、萵苣以及青梗 白菜分別在全尿素、尿素及硝酸鹽各半,以及全硝酸鹽三種處理下,來評估尿素對 此三種蔬菜之耐受性以及應用潛力;結果顯示蕹菜於此三種氮源處理皆有良好生 長,而萵苣以及青梗白菜則於尿素及硝酸鹽各半處理即可觀察到植株發育不良,且 有些許根系褐化,全尿素組則有根系黑化壞疽死亡之徵狀,三種蔬菜之尿素耐受性 為蕹菜>青梗白菜>萵苣。於 20%尿素取代硝酸鹽相較於 50%尿素組菠菜有較高乾 重,並且有較高鈣含量,其他要素含量則無顯著差異(Khan et al., 1999)。
養液以尿素取代硝酸鹽對作物之影響除了尿素本身,配方中其他要素組成也 會改變。第一為硝酸鹽濃度降低,雖有生產低硝酸鹽蔬菜之優點,但硝酸鹽濃度降 低,可能使作物有缺氮之情形。養液銨態氮濃度保持不變,硝銨比也因硝酸鹽濃度 減少而下降。
硝銨比與養液 pH 值之變化有密切關係,作物在吸收硝酸態氮以及銨態氮,
為了保持植物體內電中性,必須進行離子交換來平衡體內陰陽離子(Kirkby, 1968)。
硝酸態氮常伴隨陽離子(cotransport)一起被根部細胞吸收,使養液 pH 值上升,而銨 態氮吸收過程則是會將根部細胞內的氫離子置換至養液(antiport),使養液 pH 值降 低,因此養液配方硝銨比可決定於栽培中pH 值變動之方向(Bernardo et al., 1984)。
氮源吸收速率也因作物而有差異。以硝酸態氮及銨態氮相等養液處理,草莓 及菠菜對銨態氮吸收速率較高,養液pH 值下降快,而萵苣及豌豆則是對硝酸態氮 吸收速率較高,養液pH 值較無變動(Ikeda and Osawa, 1981)。尿素於土壤水解成銨 態氮的過程,會使土壤 pH 值升高,因土壤富含微生物之脲酶之故(Cabrera et al., 1991)。
尿素用於養液環境,尿素可被作物根部直接吸收,或者是於養液水解成銨態 氮而吸收,因養液仍不可避免有微生物參與尿素的水解。環境溫度越高,尿素於養 液水解速度越快,尿素於養液中,依然可能先經水解成銨態氮,再被作物吸收。
養液 pH 值的變動與土施尿素可能略有不同,單獨以尿素作為水耕氮源栽培 三種蔬菜,養液pH 皆下降(Luo et al., 1993)。尿素為非電解質,但養液其他要素則 為陽離子或陰離子,養液中陰陽離子總當量是相等的。本研究是探討以尿素取代硝 酸鹽陰離子的使用,硝酸鹽陰離子減少,為了兼顧養液陰陽離子當量相等,養液配 製設計中,可直接減少硝酸鉀用量,或者如欲保持鉀濃度不變,需以硫酸鉀或者以 氯化鉀來補償養液中的陰離子,本試驗以硫酸鉀取代硝酸鉀來保持鉀濃度之一致,
而養液中硫酸根的加入可能也會影響作物對要素之吸收。
八、以熱帶作物蕹菜評估含尿素及鎳之水耕養液配方可行性
蕹菜(Ipomoea aquatic Forsk)為旋花科(Convolvulaceae)半水生蔓性蔬菜。原產於 中國中南部,於旱地及水田皆能有良好生長,蕹菜莖部中空而可利於半水生環境之 生長,故又俗稱空心菜、甕菜。台灣蕹菜種植面積約為2000 公頃,年產量約 3500 公噸,其中雲林縣為蕹菜生產大縣,占約一半(106 年農業統計年報)。除以旱地種 植,於宜蘭縣礁溪鄉,臺中縣大里市、霧峰鄉,南投縣名間鄉、竹山鎮等地區則是 以水田生產蕹菜。蕹菜可耐35-40℃高溫,為夏季蔬菜供應的重要來源,蕹菜與其 同一科之甘藷葉營養價值類似,因為病蟲害少,不僅農民喜愛栽培,也深受消費者 歡迎 (謝,2009)。
蕹菜栽培期短,夏季播種至採收約15-26 天,生產穩定,可紓解夏季蔬菜短缺 問題以及於夏季颱風侵襲後迅速供應市場(陳,2001)。目前最主要品種為行政院農 業委員會桃園區農業改良場,於民國84 年從竹葉種選出之桃園 1 號;植株屬直立 性,葉色濃綠、葉柄短、節間較短、葉數多、莖部刺瘤少等特徵;本品種為生長快 速,全年約可採收6~8 次,扦插再生能力強,以及具有播種後以宿根方式採收二次 後,其葉片仍可保持狹長等優良特性(范等,1996)。
沈與許 (1991)以蕹菜作為目標作物,研究熱帶性作物適合之培養液組成,蕹菜 為好鉀作物,養液鉀比鈣為 7:3 養液 pH 及 EC 值變化較穩定,養液 EC 值以 2.0 mS/cm 有較高產量;硝酸態氮與銨態氮之比例於夏季為 6:4 為宜,而秋冬季由於光
照不足則以4:6 為宜。蕹菜相對於萵苣以及青梗白菜有較高尿素耐受性(Luo et al., 1993),華南農大蕹菜專用配方也有尿素之添加(2.5 mM)。
本研究探討鎳與尿素對‘桃園一號’蕹菜生長及要素含量之影響,特別以鎳添 加來評估含尿素水耕養液應用之可行性。使用湛水式水耕系統進行栽培,配方養液 總氮濃度不變,以尿素取代硝酸鹽進行試驗。蕹菜應具有應用含尿素水耕養液之潛 力,並在養液配方設計需考慮較高鉀添加。期望藉由含尿素水耕養液的開發,除了 尿素取代硝酸鹽氮肥可降低氮肥成本,可能還具有低硝酸鹽累積之優點。
材料與方法 一、植物材料、水耕系統及養液配方
植物材料為‘桃園一號’蕹菜實生苗,種子購自農友種苗,栽培環境為臺灣大學 園藝系具破風網之簡易設施。水耕系統為湛水式水耕(deep flow technique;DFT),
植株以泡棉固定於保麗龍板穴上,由其下容器提供養液,並以空氣加壓管線與氣泡 石提供養液足夠氧氣。蕹菜種子經次氯酸鈉消毒並用自來水洗淨,浸種一日後,播 種於洗淨珍珠石介質,待發芽子葉完全展開後,取出幼苗洗淨根部,上板定植,以 幼苗用要素減半養液配方使幼苗適應水耕環境(附表 2),約半週後才更換為不同的 養液試驗配方。養液容積於試驗一及試驗四為9 升,試驗二為 14 升,試驗三為 12 升,使用自來水來配製及補充養液。行株距為4×4 公分,每箱初始約種植 40 株,。
並於試驗取樣而逐漸減少,最終加大至5.6×5.6 公分。養液 pH 值每半週校正,維 持於6.0 左右,每週更新。養液配方參考 Hoagland 與 Arnon (1950),微量元素於各 試驗除鎳之外皆一致,各試驗之鎳變級是以直接於養液添加 1 mM 七水硫酸鎳 ( NiSO4·7H2O, Nickel(II) sulfate heptahydrate, Sigma, EP grade) 母液達成。尿素變級 則是以尿素取代原傳統養液中之硝酸鹽,並保持總氮濃度不變(附表 2)。
二、試驗方法與調查項目
試驗一 鎳毒害對水耕‘桃園一號’蕹菜生長與要素含量之影響
本試驗為6 個鎳變級之單因子試驗,6 個鎳變級為 0, 0.85, 1.7, 8.5, 17.0, 85.3 μM,即養液鎳濃度為 0, 0.05, 0.1, 0.5, 1, 5 mg∙L-1。養液總氮濃度為15 mM,NH4- N :Urea-N:NO3-N=1:7:7,其餘巨量要素濃度 P、K、Ca、Mg 分別為 2, 6, 4, 2 mM (附表 2)。
自2016 年 8 月 19 日起至 9 月 14 日結束,共持續 26 天,每處理標記 8 株測量 株高、葉綠素SPAD 值、莖徑及葉長,視為 8 重複。而乾鮮重則於 26 日結束時取 樣,將植株根莖葉分開後,每三株合併視為1 重複測量乾鮮重,共 3 重複。
試驗二 ‘桃園一號’蕹菜在含尿素水耕液栽培下之鎳適宜濃度
本試驗為5 個鎳變級搭配 2 個尿素變級之複因子試驗,5 個鎳變級為 0, 0.5, 1, 2, 4 μM。尿素變級為 0 及 8 mM,即 0%與 50%。養液總氮濃度為 16 mM,其餘巨 量要素濃度P、K、Ca、Mg 分別為 2, 8, 3, 1 mM (附表 2)。
自2017 年 8 月 16 日起至 8 月 30 日結束,共持續 14 天,每處理標記 10 株,
於試驗第7 日第 14 日測量株高、莖徑、根長及葉綠素 SPAD 值,視為 10 重複。而 乾鮮重則將植株根、莖、葉分開後,每5 株合併視為 1 重複,測量乾鮮重,共 4 重 複,並將烘乾後的樣品進行要素分析。
試驗三 尿素濃度與適量鎳添加對水耕‘桃園一號’蕹菜生長及要素含量之影響 本試驗為 7 個尿素鎳變級搭配 2 個鎳變級之複因子試驗,7 個尿素態氮變級 為1, 2, 4, 6, 8, 10 mM (以下尿素態氮皆簡稱為尿素),鎳變級為(0, 1 μM)。養液總氮 濃度為14 mM,其餘巨量要素濃度 P、K、Ca、Mg 分別為 1, 8, 3, 1 mM (附表 2)。
自2018 年 3 月 20 日起 4 月 10 日結束,共持續 21 天,每處理標記 10 株測量 株高、莖徑,視為10 重複。而乾鮮重則將植株根、莖、葉分開後,每 6 株合併視 為1 重複,測量乾鮮重,共 3 重複,並將烘乾後的樣品進行要素分析。
試驗四 尿素濃度與鎳添加對水耕‘桃園一號’蕹菜氮源與巨量要素吸收之影響 本試驗為3 個尿素鎳變級搭配 2 個鎳變級之複因子試驗,試驗配方與試驗三 相同,將7 個尿素變級縮減為 3 個尿素變級(2, 4, 10 mM),鎳變級為(0, 1 μM)。養 液總氮濃度為14 mM,其餘巨量要素濃度 P、K、Ca、Mg 分別為 1, 8, 3, 1, mM (附 表2)。
自2018 年 10 月 21 日起至 11 月 12 日結束,共持續 22 天,每處理標記 10 株 測量株高,視為10 重複。而乾鮮重則將植株根、莖、葉分開後,每 6 株合併視為
1 重複,測量乾鮮重,共 3 重複。每週於更換養液前後獲取水樣,而蕹菜對尿素態 氮、銨態氮、硝酸態氮與巨量要素吸收速率,是以養液每週各要素濃度差,再除以 水耕箱上所種植蕹菜株數所得。
三、分析方法 (一)、葉綠素含量
為量測蕹菜新葉黃化之現象,以葉綠素計(SPAD-502,Konica Minolta)測量葉 綠素相對含量,測量頂端第一片完全展開葉之葉中段之左右兩側,並將兩側數據平 均而得到該觀察值。
(二)、養液 pH 值及 EC 值
以手持型 pH 儀(Janco,6010M)以及手持型 EC 儀(Janco,3010M)測量養液 pH 及 EC 值。
(三)、植體樣品乾燥與磨粉
參考Cataldo 等(1975)之樣品前處理。
植體各部位經二次水洗淨後以紙巾拭乾,以 70℃烘乾 48 小時或至恆重,再 經磨粉機均質後至可通過 40 mesh 篩網之顆粒大小,均質樣品置於塑膠樣品盒於 乾燥環境保存,待後續進行元素與硝酸鹽含量分析。於分析稱重前,置於乾燥皿中 一日以去除吸溼水分。
(四)、植體硝酸鹽樣品製備
參考Miner and Queiruga (1995)植體硝酸鹽熱水萃取法。
精秤0.4 g 樣品於塑膠離心管中,以 80℃熱水加至約 40 毫升後,以 80℃水 浴30 分鐘,水浴結束後趁熱通過 ADVENTEC 1 號濾紙,以熱水潤洗濾紙三次,
待溶液冷卻後以去離子水定量至50 mL,即完成為植體硝酸鹽樣品製備。
(五)、植體礦物元素樣品製備(乾灰化法)
參考Clegg 等(1981)針對微量元素分析之乾灰化法。
精秤0.2 g 磨粉樣品,製於陶瓷坩鍋中,以灰化爐加熱至 450 度℃灰化,至樣 品為灰白色為止。降溫後取出坩鍋,加入5 mL 1.2 N HCl (Merck,GR grade),混 合均勻後通過ADVENTEC 1 號濾紙,以熱水潤洗濾紙三次後,待溶液冷卻後以去 離子水定量至50 mL,即完成為礦物元素樣品製備。此灰化法製備樣品溶液可於後 續以鉬藍法測磷,以及用來測量鉀、鈣、鎂、鐵、錳、鋅、鎳等要素含量。
(六)、磷:
參考King (1932)之鉬藍法。
1.反應藥劑配製:
(1) P-B solution (Acid molybdate stock solution): 37.63 g (NH4)Mo7O24·4H2O (Merck,GR grade)加熱溶於 250 mL 去離子水,冷卻至室溫後加入 750 mL 37.5% HCl (Merck, GR grade)並定量至 1000 mL,加入 H3BO3 (Kanto, EP grade) 直至飽和沉澱。溶液存放於褐色血清瓶,冷藏4℃保存。
(2) P-C powder (Drying reducing agent): 1.25 g 1-amino-2-naphthol-4-sulfonic acid + 2.5 g Na2SO3 (Sodium sulfite) (Merck,GR grade) + 73.125 g Na2S2O5
(Sodium pyrosulfite) (PanReac,ACS grade) 均勻混合,密封並遮光,可存放 6 個月。
(3) P-C solution (Dilute reducing agent): 取 3.2 g P-C powder 加熱溶於 20 mL 水後冷卻至室溫,全程以不透光之錫箔紙包覆,需現配現用。
(4) 標準液配製: 精秤 0.4394 g K2HPO4 (Merck, GR grade),以去離子水定量至 1000 mL,為 1000 mg∙L-1之標準溶液,將標準溶液稀釋至100 mg∙L-1得工作母 液,以工作母液配製1, 3, 5, 7, 9 mg∙L-1之標準液用於檢量線製作。
2.試驗流程:
於玻璃式管中依序加入0.25 mL P-B solution、5 mL 適當稀釋之礦物元素樣品、
0.25 mL P-C solution,同時以 1, 3, 5, 7, 9 mg·L-1之操作標準液製作標準曲線、以及 以5 mL 去離子水製作空白樣本,震盪混合均勻,靜置 40 分鐘反應後,以微量盤
分光光度計(Microplate Spectrophotometer,Epoch 2,BioTek)測定 660 nm 吸光值,
並根據檢量線以內插法求得樣品磷濃度。
(七)、全氮-凱式氮(Kjeldahl nitrogen)
參考(Kirk, 1950)之含硒催化劑凱氏氮法,再以擴散法與反滴定回推樣品之全 氮含量。
1.氮消化液製備:
烘 乾 磨 粉 後 之 樣 品 精 秤 0.2 g , 加 入 等 重 之 催 化 劑 (Selenium mixture, K2SO4:CuSO4:Selenium= 100:10:1) (Merck,GR grade),以秤藥紙完全包覆後置入 50 mL 氮分解管(digestion tube)中,加入 5 mL 濃硫酸(Merck,GR grade),以加熱板加 熱至400℃約 6-8 小時,至氮分解管內液體呈透明無色,冷卻後以去離子水定量至 50 mL,即製成氮消化液,裝入塑膠樣品瓶後於乾燥陰涼環境下保存,並可隨後以 擴散法收集氨氣,並以反滴定求得氮消化液內含的銨態氮含量,而最終回推樣品全 氮含量。
2.擴散法:
(1) 指示劑(bromophenol blue):取 0.2 g bromophenol blue 溶於 100 mL 95% 乙 醇並以1 N NaOH 調整 pH 至 5.0 左右。
(2) 氨吸收劑:混合指示劑之硼酸溶液:以 1000 mL 4% H3BO3 (Kanto, EP grade) 溶液:5 mL 指示劑之比例混合。
(3) 操作流程:
取適當稀釋之氮消化液10 mL 置於 90 mm 玻璃培養皿中,並於培養皿中 央放至含有4 mL 混合指示劑之硼酸溶液之小擴散皿,以分注器加入 10 mL 10 N NaOH (第一化工,工業級) 與氮消化液反應後,立即蓋上玻璃培養皿上蓋並 以膠帶密封,並稍微搖動使液體均勻混合使氨氣可順利擴散至中央小擴散皿 中,常溫下放置48 小時後開蓋取出小擴散皿,以 0.01 N 標準酸 H2SO4 (Merck,
Titrisol®)以反滴定方式滴定至原先指示劑之黃色,紀錄滴定至當量點所需之 容積,每樣品做兩套重複,誤差需小於5%,最後取兩組平均為最終滴定量。
3.氮含量計算
氮含量 (mg·g-1) = (平均滴定量(mL)+0.05)*14*0.01*稀釋倍率*0.96/取樣量(g) 0.05:空白試驗所需毫升數 稀釋倍率:取樣 5 mL 定量至 50 mL
0.96:回收率 96% 0.01:滴定用硫酸當量濃度 14:N 原子量 (八)、尿素(二乙醯一肟呈色法)
參考(Kyllingsbæk, 1975)之二乙醯一肟呈色法(Diacetyl monoxime assay)。
1.反應藥劑配製:
(1) DAM-KNO3: 2.5 g diacetyl monoxime (Sigma,EP grade) 與 2.0 g potassium nitrate 溶於二水,定量至 100 mL。
(2) TSC: 0.25 g thiosemicarbazide (Sigma,EP grade)溶於二次水,定量至 100 mL。
(3) Acid solution: 取 175 mL 37% hydrochloric acid (Merck,GR grade)與 100 mL 85% phosphoric acid (Kanto,EP grade),以二次水定量至 500 mL。
(4) Color reagent: 使用前配製,將 30 mL DAM-KNO3溶液、12 mL TSC 溶液和 450 mL acid solution 均勻混和。
2.標準液配製:
精秤0.3005 g 尿素(Sigma,EP grade),以去離子水定量至 500 mL,為 10 mM 之尿素標準溶液並以標準溶液配製50, 100, 150, 200, 250 μM 之標準液用於檢量線 製作。
3.試驗流程:
以微量塑膠離心管加入 0.2 mL 之適當稀釋樣品或標準品,加入 0.6 mL color reagent,以 80℃乾浴 30 分鐘,冷卻後以微量盤分光光度計測定 527 nm 吸光值,
並根據檢量線以內插法求得樣品尿素濃度。
(九)、硝酸鹽(水楊酸呈色法)
參考Cataldo 等(1975)之水楊酸呈色法。
1.反應藥劑配製:
(1) Salicylic acid-H2SO4 5% (w/v): 5 g Salicylic acid (Merck GR grade)溶於 100 mL 濃硫酸,以棕色血清瓶於室溫下可存放一週。
(2) 10N NaOH: 400.0 g NaOH(第一化工,工業級)以二次水定量至 1 L。
2.標準液配製:
精秤0.5055 g KNO3 (Nihon Shiyaku,EP grade),以去離子水定量至 50 mL,為 100 mM 之標準溶液,並以標準溶液配製 1, 3, 5, 7, 9 mM 標準液用於檢量線製作。
3.試驗流程:
於平底試管中加入適當稀釋之樣品溶或標準液 0.2 mL,再加入 0.8 mL 5%
Salicylic acid-H2SO4,混合均勻,於室溫下靜置反應20 分鐘,加入 5 mL 二次水,
再加入5 mL10N NaOH,混合均勻,水浴冷卻至室溫後,以微量盤分光光度計測定 410 nm 吸光值,並根據檢量線以內插法求得樣品硝酸態氮濃度。
(十)、銨態氮(納氏比色法)
參考Jeong 等(2013)改良之納氏比色法(modified Nesslerization)。
1.反應藥劑配製:
(1) 檸檬酸鈉-酒石酸鉀鈉溶液(MS)
將300 g KNaC4H4O6 (Hanawa,EP grade)與 250 g Na3C6H5O7 (Nacalai,EP grade)溶於 900 mL 二次水,加入 1.35 g PVA,定量至 1 L,以 H2SO4與NaOH 校正pH 至 7.0。
(2) 納氏反應劑 (Nessler's reagent):購自 MERCK 公司。
2.標準液配製:
精秤 0.4002 g NH4NO3 (Merck,EP grade),以去離子水定量至 500 mL,為 10 mM 之 stock solution。並以標準溶液配製 25, 50, 100, 200, 250 μM 標準液用於檢量 線製作。
(3) 試驗流程:
以塑膠微量離心管加入適當稀釋樣品溶液樣品或標準溶液 0.75 mL,再加入 0.25 mL Nessler's reagent,混合均勻,於室溫下靜置反應 20 分鐘,微量盤分光光度 計測定425 nm 吸光值,並根據檢量線以內插法求得樣品銨態氮濃度。
(十一)其他礦物元素(K, Ca, Mg, Fe, Mn, Zn, Ni)之測定:
試驗一之蕹菜葉片樣品委由行政院農委會茶業改良場茶作技術課土壤肥力實 驗室,以感應偶合電漿原子吸收光譜儀(ICP-OES,PerkinElmer,Optima 2100 DV) 測量鉀、鈣、鎂、鐵、錳、鋅、鎳等元素,其餘試驗二至試驗四的礦物元素分析則 以火焰式原子吸收光譜測鉀、鈣、鎂、鐵、錳、鋅、鎳等元素。
1.標準液配製:
使用MERCK 之 1000 mg∙L-1 之鉀、鈣、鎂、鐵、錳、鋅、鎳標準液,稀釋 為100 mg∙L-1的儲備溶液,再將其稀釋為操作標準液以製作標準曲線。
2.原子吸收光譜儀分析:
以火焰式原子吸收光譜儀 (atomic absorption spectrophotometer,Shimadzu,AA- 6300 ) (後續討論簡稱為 AA)及鹵素燈管 (Hamamatsu Photonics K. K., Japan),測定 鉀、鈣、鎂、鐵、錳、鋅、鎳,空氣分壓5 Kg∙cm-2,乙炔分壓1 Kg∙cm-2。
四、統計分析
試驗數據利用 CoStat 6.4 (CoHort Software, Monterey, CA, USA)進行變方分析 (analysis of variance, ANOVA),以最小顯著差異 (least significant difference test, LSD) 比較5%差異顯著性。以 SigmaPlot 14.0 (Systat Software, San Jose, CA, USA) 進行 圖表繪製及迴歸分析。
結果
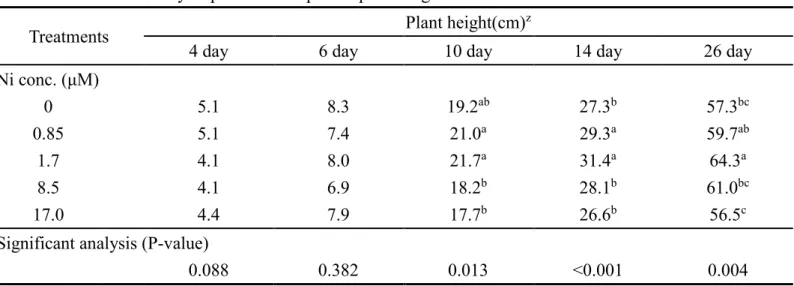
一、鎳毒害對‘桃園一號’蕹菜生長與要素含量之影響(試驗一)
本試驗為求得於蕹菜於水耕環境中,對鎳之適宜與毒害濃度範圍,因此試驗 一之鎳濃度範圍較廣,共有6 個鎳濃度變級(0 至 85.3 μM),養液氮源以尿素取代 46.6%氮源(以下簡稱半尿素)來判斷於半尿素環境中,概略之鎳毒害與鎳適宜濃度,
作為後續試驗參考依據,並探討鎳於蕹菜吸收後累積於地上部之情形。
(一)、蕹菜於高鎳環境產生之毒害病徵
試驗從定植上板開始至結束共持續26 天。蕹菜經 85.3 μM 最高鎳濃度處理第 4 天後,子葉以上新生部位開始黃化;於第 5 天莖部倒伏並產生水浸狀斑塊,且新 葉可觀察到捲曲皺縮、部分焦枯之徵狀;第6 天子葉以上新生部位則完全焦枯。蕹 菜子葉以上部位對 85.3 μM 鎳濃度處理反應迅速,於 6 天內即可使蕹菜地上部焦 枯,而至實驗後期,地上部因完全嚴重影響蕹菜生長,而無後續新生長(圖 1)。
相較於子葉以上部位,根部以及宿存子葉對高鎳環境反應較緩慢。處理第 6
天雖地上部完全焦枯,但宿存子葉以及根部仍無明顯病徵。處理第8 天至第 10 天,
根部支根開始壞死並脫落。宿存子葉從試驗開始至14 天外觀則幾無變化(圖 1)。
鎳濃度次高之兩處理(8.5 μM、17.0 μM)則於試驗中期,植株株高較低鎳處理 低(圖 2A)。8.5 μM 鎳濃度處理使蕹菜新葉黃化,且新葉缺乏蠟質,與未加鎳對照 相比除葉片體積較小,表面也較無光澤(圖 2B)。17.0 μM 除了新葉黃化,少部分新 葉葉緣則有壞疽及皺縮之徵狀(圖 2B)。8.5 μM 及 17.0 μM 鎳濃度處理,新葉葉綠 素SPAD 值於第 7 天及第 10 天較其他低鎳組別低;但此新葉黃化之徵狀於試驗後 期則減輕,新葉SPAD 值在所有鎳處理皆無顯著差異(表 1)。
(二)、植株生長
鎳濃度處理間之株高差異於試驗前期(第 4 天及第 6 天)不顯著,於試驗中期 之後各組間之株高則開始有差異(表 2)。處理第 10 天之蕹菜株高與莖徑,於 8.5 μM 及17.0 μM 鎳濃度處理下顯著較細,但葉長於各組間仍無差異;第 14 天蕹菜株高、
莖徑以及葉長,於8.5 μM 及 17.0 μM 鎳濃度下較低。8.5 μM 及 17.0 μM 對蕹菜而 言,應為偏高的鎳添加濃度(表 1, 表 2)。
0 μM、0.85 μM、1.7μM 三個處理間之生長至 14 天後開始有差異。0.85 及 1.7 μM 鎳較無鎳之對照,植株有較大的株高以及莖徑(表 1, 表 2)。至試驗末期第 26 天 之蕹菜已具有基部新生之側枝。綜觀株高、主幹莖徑、側枝莖徑及全株乾鮮重,0.85 μM 及 1.7 μM 鎳處理者為較適宜之鎳添加範圍(表 3)。
較高劑量之8.5 μM 及 17.0 μM 鎳濃度處理使蕹菜的根莖葉乾鮮重皆下降,於 此高鎳環境下,抑制全株各部位蕹菜之生長(表 4)。較低劑量之 0.85 μM 及 1.7 μM 鎳處理為水耕環境中較適宜的添加劑量,蕹菜葉之乾鮮重與未含鎳對照相比無顯 著差異,然而莖及根部鮮重於此適宜劑量則較無鎳對照高(表 4)。
(三)、要素含量及鎳累積
蕹菜葉片磷含量於適量鎳0.85 μM 及 1.7 μM 處理較高,可能與此鎳濃度處理 下有較良好生長有關;蕹菜葉片鐵含量則於1.7 μM 以上之鎳濃度處理較低(表 5)。
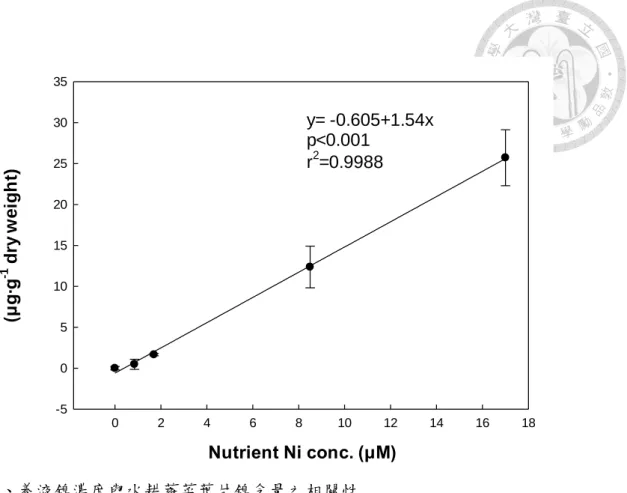
鎳也可經根部吸收,而於地上部葉片檢測到鎳之含量;蕹菜養液鎳濃度越高,則蕹 菜葉片累積之鎳含量也越高,養液鎳濃度與蕹菜葉片鎳含量,以線性迴歸分析結果 R2 值為 0.9988,p 值<0.001,顯示為極顯著正相關(圖 3)。鎳的每日容許攝取量 (Tolerable Daily Intake;TDI)為 0.02 mg∙Kg-1∙day-1,以60 公斤重成人為例,每日鎳 可容許攝取量為1.2 mg。17.0 μM 鎳濃度處理之蕹菜葉片鎳含量為 25.9 μg∙g-1DW,
以蕹菜乾物重百分比約為10%換算,每人每日需食用約 460 g 鮮重之蕹菜才會超過 鎳的每日容許攝取量,因此以17.0 μM 鎳濃度所栽培蕹菜而言,鎳含量稍高,故具 食用風險。
以整體生長及葉片鎳累積作評估,8.5 μM 及 17.0 μM 為過量鎳濃度範圍,此 過量鎳抑制半尿素水耕蕹菜之生長,早期有新葉黃化之現象,使葉片葉綠素SPAD 值較低,雖於後期恢復,但蕹菜前期因鎳所造成的徵狀,導致最終整體產量下降。
蕹菜不適合於此鎳濃度範圍下生長,而葉片鎳累積之含量也稍高,以鎳累積角度來
判斷也不建議食用。
0.85 μM 及 1.7 μM 應為較適當的鎳添加範圍,可相較於無鎳對照提高採收期 蕹菜的株高以及莖與根之乾鮮重。葉片則只有微量鎳含量(0.5 及 1.6 μg∙g-1),可符 合食用上低鎳含量之要求。
二、適宜鎳對含尿素水耕‘桃園一號’蕹菜生長及要素含量之影響 (試驗二)
試驗一結果顯示8.5 μM 以上為鎳過量之範圍,因此試驗二排除 8.5 μM 以上 之鎳添加濃度,進一步在0 至 4 μM 之間,共 5 個鎳濃度變級範圍,探究最適之區 間。氮源部分以兩個尿素變級0, 8 mM,也就是 0%、50%的尿素比例(以下簡稱無 尿素組與尿素組),比較尿素之存在,與適量鎳添加範圍,對所栽培的蕹菜生長與 要素含量的差異。
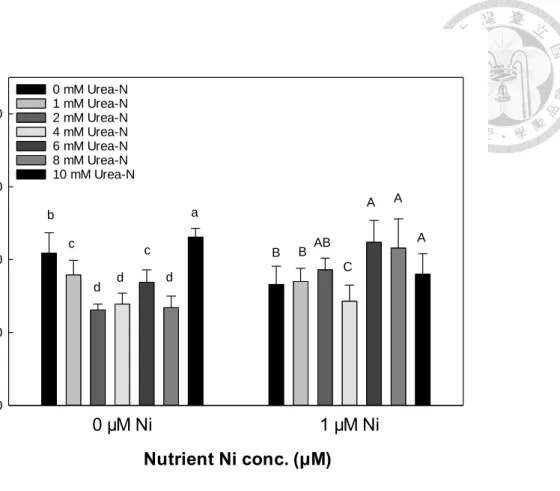
(一)、外觀與生長
生長前期(第 7 天)以及生長後期(第 14 天),蕹菜株高以 0.5 μM、1 μM 兩組鎳 處理較其他鎳處理組高,尿素變級間則在株高無顯著影響(表 6)。蕹菜新葉葉綠素 SPAD 值僅於生長前期有差異,新葉葉綠素 SPAD 值尿素組較無尿素組低,而鎳處 理中2 μM 及 4 μM 鎳濃度下新葉葉綠素 SPAD 值相較其他鎳濃度低(表 6)。蕹菜生 長後期之新葉葉綠素SPAD 值則於尿素或鎳變級下皆無顯著差異(表 6)。
生長前期蕹菜之莖徑於鎳變級中以 2 μM 鎳處理最細,而尿素組的莖徑則較 無尿素組細(表 6)。莖徑與新葉 SPAD 值有相似之趨勢,2 μM 鎳可能抑制蕹菜的前 期外觀生長。生長後期(第 14 天)葉綠素濃度及莖徑於各組間皆無顯著差異(表 6)。
生長前期與後期蕹菜根長皆以無尿素組較尿素組較大,而鎳濃度對蕹菜根長影響 較不顯著。
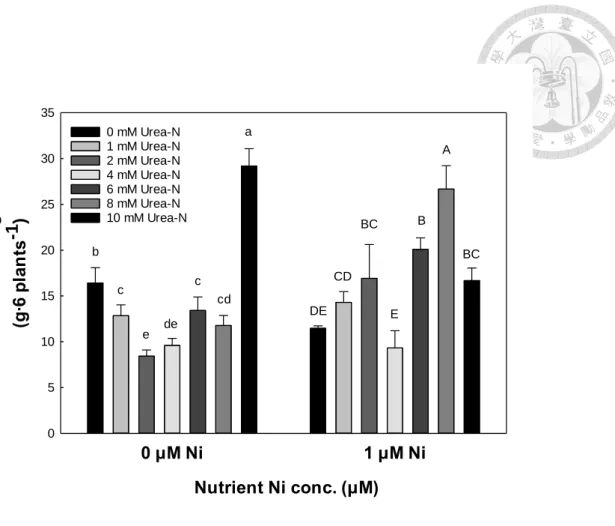
(二)、乾鮮重
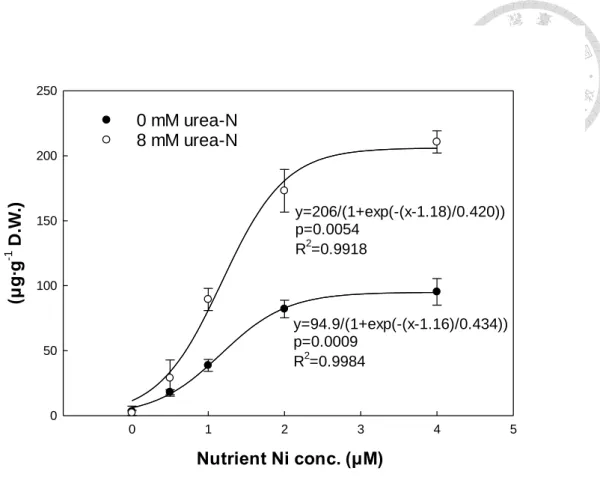
將生長後期(第 14 天)蕹菜各部位乾鮮重以單因子分析顯示,蕹菜葉與莖部之 乾鮮重於0.5、1 μM 鎳處理下較高,且在無尿素組與尿素組皆有相同趨勢(表 7)。
蕹菜根部對鎳處理的影響則依尿素的存在與否有差異。養液0.5、1 μM 鎳處理僅於
尿素組下,蕹菜根部乾鮮重較高,而無尿素對照組添加鎳,則未有提升蕹菜根部乾 鮮重的趨勢(表 7)。
鎳與尿素對蕹菜各部位與地上部經濟產量(葉與莖相加)複因子分析中,乾鮮 重與株高趨勢相近;蕹菜地上部葉與莖之乾鮮重同樣以0.5 μM、1 μM 兩組鎳處理 較其他鎳處理組高,在較高之鎳濃度(2 μM、4 μM)則比 0.5、1 μM 鎳濃度低,而尿 素變級間則無顯著影響 (表 8)。
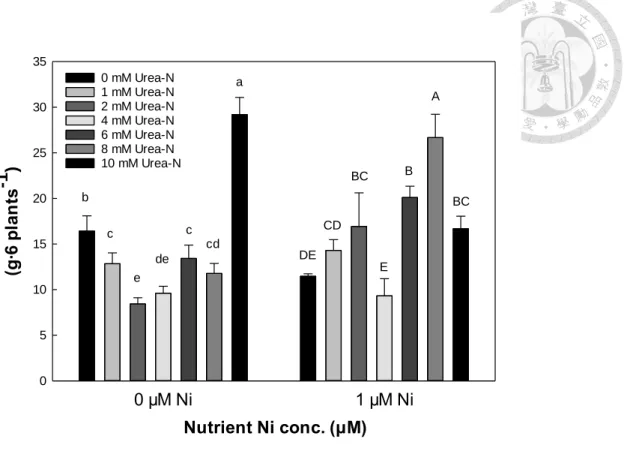
蕹菜地下部根之乾鮮重僅無鎳對照有較低之根長,其他鎳添加濃度之根乾鮮 重皆高於無鎳對照,並鎳濃度添加多寡(0.5 μM 至 4 μM)對蕹菜根之乾鮮重則較無 顯著差異,而根部之乾重於尿素組比無尿素組高(表 8)。蕹菜地上部和地下部對鎳 濃度的反應不同,水耕環境於0 μM 至 4 μM 區段,地上部的生長在此濃度區段,
大致於各部位0.5 μM 及 1 μM 兩鎳處理有較高的乾鮮重(表 8)。
根據養液鎳及尿素對蕹菜乾鮮重的複因子分析可大致推估,0.5 μM 及 1 μM 鎳濃度應為較適宜之鎳添加濃度,而且尿素變級與鎳變級之間無交感顯著(表 8)。
但於單因子分析中,雖於統計上未達5%差異水準,最適鎳濃度可能因尿素存在而
有差別(表 7)。蕹菜莖與葉之鮮重於無尿素組是以 0.5 μM 鎳濃度較高,尿素組則是 以1 μM 鎳濃度較大(表 7)。養液如有尿素可能相較無尿素對照,有較高的最適鎳 添加濃度。
(三)、巨量要素含量
蕹菜氮、磷、鉀、鈣及鎂之單株要素含量(含量乘以單株重量),於適量鎳 0.5 μM、1 μM 相較於無鎳處理高,顯示適量鎳添加可提升蕹菜單株之巨量要素含量(表 9)。葉片鈣含量於無鎳組較高,鎂含量則於 2 μM 鎳組較高,磷與鉀則於各鎳變級 下無顯著差異(表 10)。莖部於 0.5 μM 鎳處理下則有最高之磷、鉀、鈣及鎂含量,
而1 μM 鎳則較 0.5 μM 鎳略低(表 11)。蕹菜根部磷、鉀及鈣要素含量於無鎳對照 較高(表 12)。
無鎳組蕹菜根部巨量要素含量雖較高,但無鎳組之蕹菜根部乾鮮重較低,此
情形可能為生長較緩慢而導致(表 8, 表 12)。1 μM 鎳下,蕹菜葉、莖、根之鈣與鎂 含量皆較其他鎳處理較低;莖之磷與鉀含量以0.5 μM 鎳處理較高,而 1 μM 鎳則 略低(表 10, 表 11, 表 12)。然而,1 μM 鎳下蕹菜磷、鉀、鈣及鎂含量較低或略低 之趨勢,並未於單株要素含量見得(表 9)。單株要素含量為要素含量乘以單株總乾 重而得,因此1 μM 鎳下磷、鉀、鈣及鎂於各部位含量較低,實為快速生長所造成 含量稀釋效應之故。
兩尿素變級間對蕹菜外觀生長與乾鮮重較無顯著差異,但要素含量則有顯著 差異。尿素變級對蕹菜氮、磷、鈣於根莖葉各部位含量影響的趨勢皆相同。蕹菜葉 片與莖部於尿素組相較無尿素組有較高全氮含量,而葉片、莖部及根部磷含量於也 是皆於尿素組較高;鈣含量則相反,是於無尿素組於葉片、莖部及根部皆較高(表 10, 表 11, 表 12)。
尿素變級對鉀與鎂含量影響因蕹菜部位而異。葉於無尿素組有較高鉀含量,
莖與根則是無尿素組有較低鉀含量;根於尿素組有較高鎂含量,莖與葉則是於尿素 組有較低鎂含量(表 10, 表 11, 表 12)。兩尿素變級對單株要素含量之趨勢與莖部 要素含量趨勢接近,因莖占全株乾重比例較大之故(表 9)。
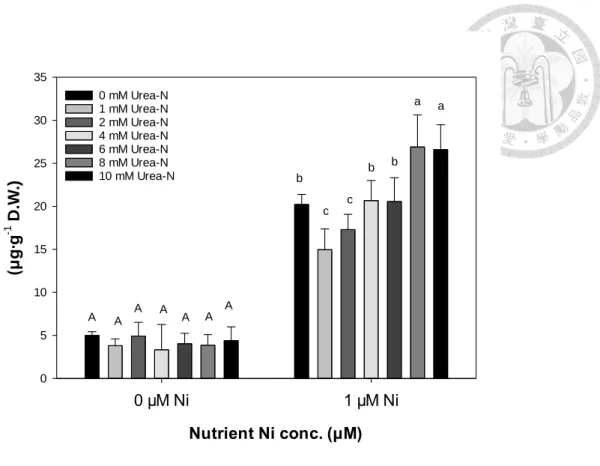
(四)、微量要素含量與鎳累積
鐵、錳、鋅及鎳於蕹菜各部位之含量差異大。鐵、鋅與鎳於蕹菜根部含量最 高,根部鐵含量約是莖及葉含量的10 倍、根部鎳含量也約為葉片鎳含量之 10 倍,
莖部鎳含量則低於AA 之儀器檢測極限(< 5 μg∙g-1DW),而根部鋅含量約為地上部 的2 倍。錳含量非在根部含量最高,而是以葉片含量最高,約是莖部及根部含量的 2 倍(表 10, 表 11, 表 12)。
尿素變級對蕹菜錳與鋅含量於根莖葉趨勢皆一致,皆是尿素組較無尿素組高 其中錳含量的差異幅度較大,莖部高尿素組錳含量約是無尿素組的4 倍(表 11)。鐵 含量僅莖部於尿素組下較高,葉與根部則無顯著差異。
鎳變級也影響蕹菜其他微量元素鐵、錳、鋅之含量。鋅含量於無鎳組下較高,