嘉南藥理科技大學專題研究計畫成果報告
計畫編號: CN9621
計畫名稱:CpG motifs 誘發魚類免疫系統產生細胞素之分析研究
執行期間:96 年 1 月 1 日至 96 年 12 月 31 日
□整合型計畫 □個別型計畫
計畫總主持人: 計畫主持人:沈國愉
子計畫主持人:
中華民國97 年 3 月 31 日
CpG motifs 誘發魚類免疫系統產生細胞素之分析研究 生科系 沈國愉
中文摘要
未甲基化CpG ODNs可活化哺乳類動物免疫系統,也已經在哺乳類動物做過 非常多的試驗。而目前也已經有研究指示CpG ODNs可以活化魚類免疫系統。在 魚類種CpG ODNs可以誘導抗病毒cytokines產生,提高非特異性毒殺細胞產生、
活化一些免疫相關基因,如:IFN / , IL-1, IL-12,及間接誘導細胞遷移與提高 血清中溶菌酶產生。本實驗選用在南台灣養殖業常見的魚種金目鱸魚(Lates
Calcarifer) 做為材料。CpG ODNs可以誘導鳟魚頭腎細胞增生及促使頭腎細胞產
生細胞增生因子(PIFs),進而促使細胞增生。當B細胞移除後,以CpG ODNs刺激 鳟魚頭腎細胞仍會促使細胞增生。這結果顯示,PIFs非由B細胞產生其作用標的 亦非B細胞。因此在本研究中將試圖尋找PIFs之來源及作用標的細胞?此外,也 希望可以純化及鑑別出PIFs為何?本研究結果顯示經由CpG ODNs刺激後,吞噬 性細胞會產生蛋白質PIFs,誘導淋巴細胞中某一族群的增生。當吞噬性細胞移除 後,PIFs並不能促使細胞增生,因此吞噬性細胞在此反應中也同時扮演如同是 accessory cells角色。
關鍵字:CpG ODNs、金目鱸魚、頭腎、細胞增生因子、淋巴細胞、吞噬性細胞
Abstract
The effects of unmethylated CpG oligodexynucleotides (ODNs) on the mammalian immune system are relatively well studied. CpG ODNs have been shown to induce a variety of different immune responses in fish including IFN / induction along with antiviral activity, macrophage activation, cytokine production and activation of other immune-related genes and cell proliferation.
CpG ODNs can indirectly inducing cellular migration in vitro and enhancing serum lysozyme activity in vivo. Sea bass (Lates Calcarifer), a common economical aquaculture species in south Taiwan, was chosen as the objective in this study.
Previous studies showed that CpG ODNs can induce trout head kidney cells to produce proliferation inducing factors (PIFs), and then result in cell proliferation.
However, B cells depletion did not affect the responses of trout head kidney cells to CpG ODNs stimulation. These results indicate that neither the source of the proliferation-inducing factors or the proliferating responder cells are B cells. The aims of our study are to find the source of PIFs and their target cells. Furthermore, we would try to purify and identify of the proliferation-inducing factors. Our results showed PIFs were proteins secreted by CpG ODNs stimulated phagocytes, and PIFs induced a subpopulation of lymphocyte to proliferate. Besides, phagocytes were supposed to act as accessory cells in this response.
Keywords: CpG ODNs, Sea bass (Lates Calcarifer), head kidney, proliferation inducing factors (PIFs), lymphocyte, phagocytes
一、 緒論 1-1 CpG motifs
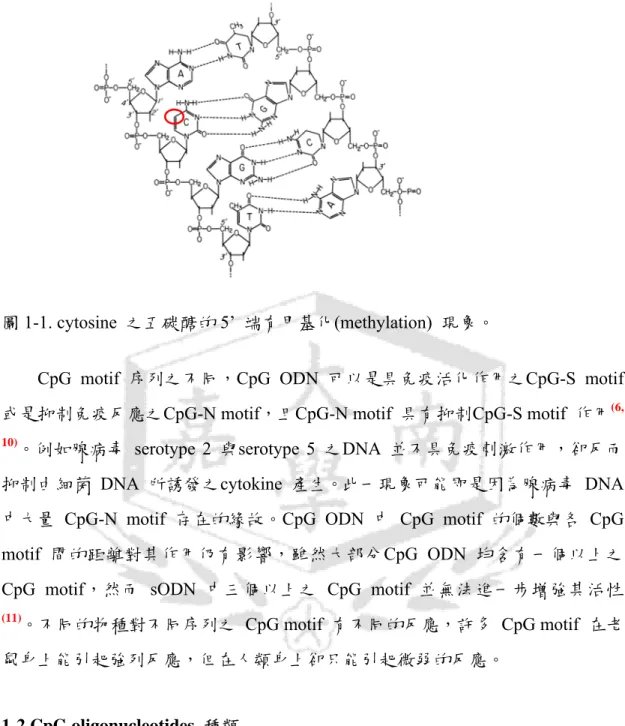
1984 年Tokunaga 與其研究團隊發現 bacille Calmette-Guérin (BCG) 細胞粹 取物中之DNA 具有抗腫瘤之功能,能夠活化 NK cell 並刺激脾臟細胞釋出 interferon(1, 2)。在鼠類身上進一步的研究發現,只有在細菌的 DNA 能誘導促使 老鼠B 細胞分裂,而哺乳類的DNA 則沒有具備此功能。DNA 對免疫細統的刺 激作用與其序列有關(3) 。在1992 年時發現NK cell 能夠被一單股之合成寡核苷 酸(synthetic oligodeoxynucleotides; sODN) 所活化,此一單股合成寡核苷酸具有 由六個核苷酸 GACGTC 所構成之迴文結構(4) 。具活化 NK cell 功能之序列迴 文結構尚有AACGTT與AGCGCT等,然而其餘迴文結構如 ACCGGT 則無法活 化 NK cell。曾經研究人員認為 DNA 對 B細胞及NK cell 之活化作用可能與其 結構有關,然而 Krieg 分析數百個不同序列之 sODN 試圖找出具有免疫活化功 能 之 sODN 其 結 構 特 徵 。 他 發 現 所 有 具 免 疫 活 化 功 能 之 sODN 皆 具 有 cytosine-guanine (胞嘧啶-鳥糞嘌呤)之雙核苷酸序列,而其活化免疫系統能力之 強 弱 則 與cytosine-guanine 雙 核 苷 酸 序 列 兩 旁 之 核 苷 酸 序 列 有 關(5, 6)。 Cytosine-guanine 雙核苷酸 5’ 端含兩個purin,3’ 端含兩個pyrimidine 具有最強 之活化鼠類B 細胞的能力,而稱為CpG motif。在細菌及無脊椎動物的DNA 與 脊索動物的DNA 有許多不同。首先CpG 雙核苷酸序列在細菌及無脊椎動物 DNA 中出現的頻率遠高於脊椎動物,此外脊索動物 DNA 中 cytosine 之五碳 醣的5’ 端有甲基化(methylation) 之現象(7) (圖 1-1)。在脊椎動物大約 60%之CpG motif 均有 methylation 之現象,細菌及無脊椎動物 DNA 則無此現象(7, 8),此一 特點可能是免疫系統用以區分細菌 DNA 與自身DNA 之依據。在進一步的研究 中發現,將具有免疫活化功能 sODN 上之cytosine 甲基化後,此一sODN 即失 去其對B 細胞之刺激活化能力(9)。同樣的,細菌之 DNA 在經由 CpG methylase 處理過後,即失去免疫活化功能。脊索動物免疫系統將 unmethylated CpG motif 視為一危險訊號(danger signal) 之推論得到初步的證實。
圖1-1. cytosine 之五碳醣的 5’ 端有甲基化(methylation) 現象。
CpG motif 序列之不同,CpG ODN 可以是具免疫活化作用之CpG-S motif 或是抑制免疫反應之CpG-N motif,且CpG-N motif 具有抑制CpG-S motif 作用(6,
10)。例如腺病毒 serotype 2 與serotype 5 之DNA 並不具免疫刺激作用,卻反而 抑制由細菌 DNA 所誘發之cytokine 產生。此一現象可能即是因為腺病毒 DNA 中大量 CpG-N motif 存在的緣故。CpG ODN 中 CpG motif 的個數與各 CpG motif 間的距離對其作用仍有影響,雖然大部分CpG ODN 均含有一個以上之 CpG motif,然而 sODN 中三個以上之 CpG motif 並無法進一步增強其活性
(11)。不同的物種對不同序列之 CpG motif 有不同的反應,許多 CpG motif 在老 鼠身上能引起強列反應,但在人類身上卻只能引起微弱的反應。
1-2 CpG oligonucleotides 種類
含有 CpG motif 之 sODN (CpG ODN) 因其 backbone 或序列之不同而引 起 截 然 不 同 的 免 疫 反 應 。CpG ODNs 依 其 所 誘 發 之 反 應 不 同 可 分 為 CpG-A(D-class) 與 CpG-B(K-class) 兩 型 。 CpG-A ODN 具 有 磷 酸 二 酯 (phosphodiester backbone , PO form) (圖1-3),在CpG-A ODN的3’端具有poly G motifs具有最佳活性。對於 NK cell 之活化與增加 IFN-α/β 產生具有較強之能力
(12)。 對 於B 細 胞 比 較 沒 有 活 化 能 力(13)。CpG-B ODN 具 硫 代 磷 酸 (phosphorothioate backbone, PS form)(圖1-3),在 CpG-B ODN 的5’ 端則沒有包 含有 poly G motifs(13)。則對 B 細胞具極強的刺激活化能力,及可以促使
cytokine 分泌,但對 NK cell 活化能力則較弱(9, 14)。
在2003年有研究人員結合 CpG-A 與 CpG-B ODN 之特性,合成兼具活化 B 細胞與刺激樹突狀細胞產生 IFN-α/β 之 sODN 稱之為 CpG-C (15)。
圖1-2. Phosphodiester 與 phosphorothioate backbone 骨架結構。
1-3 CpG motifs 對哺乳類免疫系統之作用
自從 CpG motif 被發現具有免疫活化功能後,自1996 年之後有大量的文獻 述及 CpG motif 對哺乳類免疫系統之作用。CpG ODNs 能夠活化巨噬細胞產生 IL-12,IL-12 則進一步促進 NK cell 產生 INF- (16)。CpG ODNs 能促進 B 細 胞產生 IL-6、IL-10 與 IgM (17, 18)。除了cytokine 與 Ig 外,CpG motifs 亦促進 B 細胞表面 Fc receptor 、MHC-II 及協同刺激分子(costimulatory molecules) 如 CD80、CD86、CD54 表現量之增加。協同刺激分子(costimulatory molecules)是 抗原呈現細胞上的一個分子,可以傳遞訊息給互相作用的淋巴細胞。因為 CpG ODNs 能提高樹突狀細胞 (dendrictic cell, DC) 等抗原呈現細胞 (antigen
presentingcell) 上MHC-II、CD80(B7-1)、CD86(B7-2) 之表現,因此可以用來當 作佐劑促進 Th1 與 cytotoxic T cell 之反應(19, 20)。樹突狀細胞經 CpG ODNs 的 刺激後亦產生大量的IL-2、IL-6、TNF- 。然而 cytokine 的產生可因 sODN 序 列之改變而變化。
CpG ODNs 對 T 細胞的協同刺激作用十顯得分複雜,雖然 CpG ODNs 不 能直接活化 T 細胞,但CpG ODNs 能夠在缺乏 APC 的情況下作為協同刺激分 子,誘使receptor-triggered T 細胞分裂(19) 當純的 T 細胞族群以anti-CD3 抗體經 由 T cell receptor 進行刺激活化作用後 CpG ODNs 能促使 T 細胞分裂而不需 要 IL-2 的產生。CpG motifs 對哺乳類動物免疫細統的作用簡單綜合以圖1-3表 示。
β β
圖1-3. CpG motif 對哺乳類免疫系統作用簡圖(11)。
ADCC: 抗體依賴型細胞毒素(Antibody-dependent cellular cytotoxicity) 1-4 CpG motifs 之作用機制
CpG ODN 與 LPS 均 為 具 免 疫 刺 激 活 性 的 物 質 , 兩 者 皆 由 Toll-like receptor(TLR) 做為受器,但由於 CpG ODN 與LPS 經由不同的訊息傳遞模式影 響免疫系統,因此細胞的反應模式有所不同。目前 TLR-9 被認為是CpG ODN 的 接受體 (receptor)(21)。然而與其餘 TLR family 不同的是,TLR-9 與CpG ODN 結 合之後並不能立即將此一訊息傳遞至細胞核中誘導基因的表現。sODN 與細胞表 receptor 之結合為sequence-independent,DNA 藉此一非專一性的結合後經由 endocytosis 進入細胞內。此一 endosome 經過 endosomal maturation 的過程之 後,方能起始訊息之傳遞(22)。此外 CpG ODN 對APC 與 B 細胞之刺激作用可
以藉由加入 endosomal acidification inhibitors (例如chloroquine 或monesin) 將其 完全抑制,但 LPS 之作用則不受影響(23, 24)。CpG ODN 所誘發之訊息傳遞起始 於 Toll/IL-1 receptor homology domain (TIR domain) 與 adaptor protein MyD88 之複合體(圖1-5),接著經由 MyD88 之death domain 與IL-1 receptorassociated kinases (IRAK) 之 作 用 經 另 一 adaptor protein tumor necrosis factor receptor-associated factor 6 (TRAF6) 將訊息傳遞下去,進而影響一些轉錄調節因 (transcription factors) NF-κB 來調節基因的表現(25)。此外以 CpG ODN 刺激 B 細胞和巨噬細胞之後,經由 stress-activated protein kinase (SAPKs) pathway、c-Jun NH2 terminal kinase (JNK)、p38 與 activatingtranscription factor-2 (ATF-2) 之活 化,亦促進另一轉錄調節因子activating protein-1 (AP-1)表現(23, 26, 27)。而近期來 Toll-like receptor (TLR) 9 基因從日本比目魚(Paralichthys olivaceus) 被複製 (clone)出來(28)。
圖1-4. CpG motif 經由TLR-9 訊息傳遞路徑(29)。
1-5 CpG oligonucleotides 對魚類免疫系統之影響 1-5-1 魚類免疫系統概述
硬骨魚綱 (Ostrichthyes) 包括許多高等硬骨魚 (teleosts) 在魚類中是最多
的,硬骨魚也包括大多數的食用魚,在養殖漁業上近來也發現許多感染病原菌案 例。因此緣故,大多數實驗室開始研究有關魚類免疫系統。
水生動物中,無脊椎動物僅具非特異性免疫系統(各含體液及細胞性免疫),
但缺乏特異性免疫的抗原-抗體反應,如非特異性的吞噬細胞(phagocyte) 之吞 噬作用及發炎反應能力。低等脊椎動物只有淋巴細胞 (lymphocyte),但其分化和 分工能力仍不足。魚類屬於脊椎動物,除了無顎魚類外大多已有胸腺和淋巴細 胞,也有細胞免疫 (cellular immunity, cell-mediated immunity) 反應。硬骨魚類具 有特異性及非特異性免疫系統(各含體液及細胞免疫),已有類似哺乳動物可產生 抗體之後天免疫細胞及機制,其在免疫系統功能上尚屬完整,但反應上則和哺乳 動物有所差異(30)。
雖然目前在多種高等脊椎動物體內所知的重要免疫器官如胸腺、脾臟在魚類 體內亦有發現,且具備相同功能,但就免疫系統的結構與形態而言仍存在著明顯 的差異,但目前為止尚未發現魚類有完整的淋巴結 (lymph nod) 存在(31, 32),取而 代之的是特化的腎臟前端----頭腎(head kidney, HK)來扮演初級淋巴器官(33, 34, 35, 36)。在硬骨魚中頭腎組織是造血細胞和免疫反應主要器官。它扮演角色如同哺乳 類動物中骨髓,是造血器官以及幹細胞主要來源。此外,頭腎組織在非特異性免 疫及清除損壞細胞上扮演重要角色,在硬骨魚中頭腎(HK)組織也是主要抗體產 生的器官。另外魚類亦具有淋巴細胞、巨嗜細胞 (macrophage)、吞噬細胞、漿細 胞 (plasma cell) 等抗體形成的必須物質,在脾臟與胸線的組織切片觀察中也發 現皮質(cortex) 與髓質(medulla) 的分界並不明顯,顯現並無十分完整的特化(37)。
魚類在經由各種抗原刺激後,會產生特異的細胞性和體液性免疫反應,並產 生記憶細胞,所以下次再遇到同一抗原時,就能迅速、大量的產生免疫反應而消 滅它,此為二次免疫反應(38)。當魚以抗原浸泡後,抗原成分由鰓中之巨嗜細胞 所 攝 取 , 經 血 液 系 統 送 達 脾 臟 、 腎 臟 及 心 外 膜 中 之 黑 色 素 巨 嗜 細 胞 中 心 (melanomacrophage centers),有些小魚則會到達胸腺,這些免疫器官中的淋巴球 (細胞)隨後暴露於抗原而開始產生細胞性(cell-mediated, T細胞)或抗體(體液)性 (antibody-mediated; humoral, B細胞)免疫反應(39)。
目前在魚類的免疫相關器官,其確切的免疫作用機制截至目前為止尚未完全 明瞭,部分在高等動物所具備之相似器官,在魚類體內亦常被推測具有類似之功 能,目前已知的魚類數個重要免疫器官有胸腺(40, 41)、腎臟(42)、脾臟(43) 、肝臟、
胃腸道、黏膜組織(44)等。
另一方面除了免疫器官之不同外,後天免疫 (acquired immunity) 藉由特定 之免疫細胞如B 淋巴球、T 淋巴球針對特定外來物進行消滅,其中 B 淋巴球會 產生免疫球蛋白 (immunoglobulin, Ig) 又稱為抗體 (antibody),可分為細胞膜上 的抗原結合蛋白 (附著型,membrane form),或活化之漿細胞(plasma cell) 分泌 至循環系統中的蛋白產物 (分泌型,secretory form)。存在於血液循環系統中之抗 體,是生物體抵禦外來病源菌入侵、保護個體安全的重要成員之一。高等哺乳動 物的免疫蛋白可區分為五種不同形態,分別是IgG、IgA、IgM、IgE、IgD,其彼 此之間的大小、電荷、胺基酸組成及免疫功能均不盡相同。相較於高等哺乳動物,
位居脊椎動物演化最原始地位的硬骨魚類,其免疫球蛋白相關研究一直遲到 1970 年代才逐漸被重視,至目前為止已有數種硬骨魚類與軟骨魚類如彩虹鱒 魚、大西洋鮭魚、鱈魚、鯊魚等(45, 46)的免疫球蛋白被純化、分析。
1-5-2 魚類 cytokines
Cytokines 是免疫系統中重要調節者並且是基因發現者,在魚類中可能可以 應用在疫苗發展及養殖魚業上對魚類免疫性刺激(47),在一些魚類中發現之 cytokines 功能與在哺乳動物中其實是相似的,在魚類中,最早一開始是在河豚 (Fugu rubripes 和 Tetraodon nigroviridis)及斑馬魚(Donio rerio)的基因序列中鑑定 出少數cytokines。之後也陸續在一些硬骨魚類中鑑定出來,例如:
Cytokines Fish type 參考文獻
Rainbow trout (Oncorhynchus
mykiss)
(48)
Common carp (Cyprinus carpio)
(49)
Pufferfish (Fugu rubripes and Tetraodon nigroviridis)
(50) IL-1
Smooth dogfish (51)
Medaka (Japanese rice fish)
(52)
IL-6 (抗發炎) Pufferfish (Fugu rubripes and Tetraodon nigroviridis)
(53)
Rainbow trout (Oncorhynchus
mykiss)
(54) IL-11 (IL-6 family)
pufferfish (Fugu rubripes and Tetraodon nigroviridis)
(54)
IL-2 (TH1 類型)
Pufferfish (Fugu rubripes and Tetraodon nigroviridis)
(55, 56)
Pufferfish (Fugu rubripes and Tetraodon nigroviridis)
(55, 56) IL-15 (TH1
纇型)
zebrafish (Danio rerio)
(55, 56)
IL-12 (TH1 類型)
Pufferfish
(Fugu rubripes and Tetraodon nigroviridis)
(57)
Rainbow trout (Oncorhynchus mykiss)
(58) IL-18 (TH1
類型)
Medaka
(Oryzias latipes)
(59)
Pufferfish
(Fugu rubripes and
Tetraodon nigroviridis) (53) IL-10 (TH1
類型)
Common carp
(Cyprinus carpio) (60)
Zebrafish (Danio rerio)
(61)
Rainbow trout (Oncorhynchus Mykiss)
(62)
Pufferfish
(Fugu rubripes and
Tetraodon nigroviridis) (53) IL-19/20 (IL-10
family)
Zebrafish (Danio rerio)
(53)
Pufferfish
(Fugu rubripes and
Tetraodon nigroviridis) (47) IL-22/26 (IL-10
family)
Zebrafish (Danio rerio)
(47)
Zebrafish
(Danio rerio)
(63, 64, 65, 66, 67)
Atlantic salmon
(Salmo salar)
(63, 64, 65, 66, 67) interferon 基因
Pufferfish
(Fugu rubripes and Tetraodon nigroviridis)
(63, 64, 65, 66, 67)
Japanese flounder (Paralicthys olivaceus)
(68)
Rainbow trout (Oncorhynchus mykiss)
(69, 70)
Gilthead seabream (Sparus auratus)
(71)
Common carp (Cyprinus carpio)
(72, 73) TNF-
Catfish (70)
(Ictalurus punctatus) Pufferfish
(Fugu rubripes and
Tetraodon nigroviridis) (74)
Zebrafish (Danio rerio)
(74)
Rainbow trout (Oncorhynchus Mykiss)
(75)
Common carp Cyprinus carpio
(76) CXC chemokine
(由 IL-8 所誘導)
Catfish (77)
Common carp (Cyprinus carpio)
(78)
Japanese flounder (Paralicthys olivaceus)
(79, 80)
Cichlids (81, 82, 83, 84) CC chemokine
Catshark (81, 82, 83, 84)
但是在TH2 類型cytokines (IL-4, IL-5, IL-13),IL-3, IL-7, IL-9 還沒有從魚類 身上被複製 (clone) 出來(47)。從這些研究顯示,在魚類cytokines的類型 (type) 及 多樣性 (diversification) 與哺乳動物是非常相似的(47)。
1-5-3 CpG ODNs 對魚類免疫系統影響
有關 CpG motif 對魚類免疫系統作用之研究,由於缺乏細胞表面的 marker 與抗體以區分不同之細胞與觀察細胞表面分子的變化,目前均著重於 CpG motif 對魚類巨噬細胞之活化作用與促進特異性抗體之產生。1999 年 Kanellos 以含有 兩個 5’-AACGTT-3’ CpG motif 之質體 pcDNA3做為佐劑,與 -galactosidase protein ( -gal) 注射老鼠與金魚,與質體中不含 CpG motif 之對照組相較,以 含有 CpG motif 之質體作為佐劑能夠明顯提昇 anti- -gal 抗體之產生量(85)。此 為文獻上最早有關 CpG motif 對魚類免疫系統作用之研究。
近幾年的研究發現 CpG motif 能促進鳟魚(Oncorhynchus mykiss)巨噬細胞 IFN-like cytokine 之產生與提高 IL-1 mRNA 表現量(86),亦可增進鳟魚 (Oncorhynchus mykiss)血球中nonspecific cytotoxic cells (NCC) cytotoxicity 之進 行(87);在鳟魚(Oncorhynchus mykiss) 中也發現在胞外可間接誘導細胞外遷移及在 胞內提高血清中溶菌酶 (lysozyme) 活化(88)。此外 CpG ODN 與質體 DNA 能促 進大西洋鮭魚 (Salmo salar L.)白血球產生具抗病毒作用之分子,這些分子對 chinooksalmon 胚胎細胞株 CHSE-214 有明顯的保護作用,可增強其抵禦 pancreatic necrosis virus 感染之能力(89)。在鯉魚 (Cyprinus carpio),可提高白血球 增生(90),CpG ODN 可促進頭腎中細胞吞噬作用的進行、提高IL-1 、CXC 與 CC-chemokine mRNA 之表現(90)。巨噬細胞可被 CpG ODN 活化,提昇
superoxide anion、hydrogen peroxide、acid phosphatase 在細胞中之含量(91)。
1-6 研究目標
為配合台灣漁業現況,本實驗選擇南台灣常見的養殖魚種之一尖吻鱸(Lates calcarifer) 為實驗材料,除材料取得容易外,有關鱸魚免疫之相關文獻發表量豐 富具參考價值,日後之研究成果亦希望能實際應用於產業界。
金目鱸魚(圖 1-1) 學名為Lates calcarifer, Bloch,俗名(中)為扁紅目鱸、沙鱸、
尖吻鱸、盲鱸,英文名Giant seaperch,屬於鱸亞目鋸蓋魚科尖吻鱸屬,分佈於熱 帶及亞熱帶海域,其體背及各鰭呈褐色,腹部和體側均較淡,瞳孔有紅色光輝,
可作為其最主要之辦別依據(92)。
圖1-5. 金目鱸魚(Lates calcarifer) (93)
依照之前的研究成果顯示,CpG ODN (oligodeoxynucleotide) 能刺激鳟魚 (Oncorhynchus mykiss) 白 血 球 ( 取 自 脾 臟 與 頭 腎 ) 產 生 細 胞 分 裂 促 進 因 子 (proliferation-inducing factors; PIFs),促進脾臟與頭腎細胞之分裂。然而若將B細
胞 (sIgM+) 去除後再以CpG ODN刺激依然能夠促進脾臟與頭腎細胞分裂,所以 即使將B細胞(sIgM+) 去除後,依然可觀察到極旺盛之細胞分裂現象。由此結果 推測PIFs 並非由B 細胞所產生,且PIFs之target cell 也不是B細胞,此與在哺乳 類動物上所觀察到之現象極為不同(94)。
本研究將比較去除與未去除巨噬細胞之頭腎細胞對 CpG ODN 與細菌 DNA之反應強弱就可知巨噬細胞在 proliferation response 中是否扮演 accessary cell 角色。再分別收取經 CpG ODN 與細菌DNA 處理後之去除與未去除巨噬細 胞之脾臟與頭腎細胞培養液上層液,加入未經 CpG ODN刺激之細胞懸浮液中,
結果之交叉分析即可知 PIFs 是否由巨噬細胞所產生以及 PIFs 之標的細胞是 否為 sIgM- 細胞?
PIFs 之來源可能是魚類樹突狀細胞或巨噬細胞等抗原呈現細胞。極有可能 是由巨噬細胞/單核球此一細胞族群所產生,然而目前並沒有直接證據。所以嚐 試以新的血球分離技術 (panning) 將巨噬細胞去除,比較巨噬細胞去除前後頭腎 細胞對 CpG ODNs 刺激反應之強弱與蛋白質電泳結果比對,就可迅速判斷出那 些蛋白質是由巨噬細胞所產生。
二、 材料與方法 2-1 材料
2-1-1 儀器設備
蛋白質電泳槽 Tanon VE-180-10000
半乾式轉印系統 MSD-20
電源供應器 MAJOR SCIENCE MP-2000P
迴轉式震盪器 YIH DER TS-500
分光光度計 JASCO V630
流式細胞儀 Becton, Dickinson and Company Fascan 低溫旋臂式離心機 Eppendorf,Centrifuge 5810R
自動酵素免疫分析儀 Thermo MULTISKAN ASCENT
細胞培養箱 YIH DER LE-509
pH meter DENVER-UB-10
倒立式顯微鏡 OLYMPUS CKX41
蛋白質電泳槽 Tanon VE-180-10000
半乾式轉印系統 MSD-20
2-1-2 試劑
購買公司 藥品名稱 藥品編號
Sodium dodecyl sulfate (SDS) Catalog 161-0300
Glycine Catalog 161-0305
Tris base Catalog 161-0716
Ammonium persulfate (APS) Catalog161-0700
Pre-stained protein standard Catalog 161-0318 BIO-RAD
40% Acrylamide solution (acrylamide : bis-acrylamide =29:1)
Catalog 161-0146
Tetramethylethylenediamine (TEMED) Catalog 1107320110
Tween 20 Catalog 655204
Acetic acid Catalog 6411730 Merck
Dimethyl sulfoxide (DMSO) Catalog 317275
Vybrant Apoptosis Assay Kit V13241 Invitrogen
Ruby protein gel stain S12001
胎牛血清,FBS 2XZ26140079
Gibco
Leibovitz’s L-15 medium 2XZ11415064 Bio Basic Inc Primer 8 O.D. PAGE, 2-phosphonthiat PO21-2P
Urea SI-U6504 Histopaque-1077 SI-10771
Trypan blue SI-T8154
Glycerol G8173 Methanol FLU-02581 Protease inhibitor P8340
phenol : chloroform : isoamyl alcohol 25:21:1
SI-C0549 Sigma
Chloroform FLU-02487
2-1-3 動物來源
金目鱸魚 (Lates Calacrifer)
實驗用金目鱸魚(Lates Calcarifer) 向高雄縣永安鄉養殖漁業生產區發展協會購 買,長度約為30-33 cm、重量約為 500-700g,在實驗前一天購買回後置於實驗 室水槽中,水槽中會使用魚塭養殖池的水並給予打氣。
圖2-1. 金目鱸魚 (Lates Calacrifer)。
圖2-2. 取其頭腎組織圖 (箭頭為頭腎組織位置)。
2-1-4 反應試劑
Deoxyribonucleic acid (DNA) 購自 Sigma。購買包裝容量是 5mg,取 7ml TE buffer 把 DNA 溶解開來直到呈現透明 狀,分裝置於-20℃下備用。
Lipopolysaccharide (LPS)
Escherichia coli 026:B6 之 LPS 購自 Sigma。 以 PBS buffer 稀釋至 0.1 mg/ml 後 保存於-20℃備用。
Histopaque-1077
Histopaque-1077 (Sigma) 是 一 種 含 聚 蔗 糖 (polysucrose) 和 泛 影 鈉 (sodium diatrizoate)的溶液,密度是 1.077g/ml。主要用於分離血液中的單核球細胞,儲存 在4℃。
5-(and -6)-carboxyfluorescein diacetate, succinimidyl ester (5(6)-CFDA, SE)
購自Invitrogen。是一種螢光物質,對細胞沒有毒性,可進入細胞內與蛋白質結 合並隨著細胞分裂而等量分配給子代細胞;已分裂細胞螢光強度下降。利用CFSE 的特性可以研究細胞增生能力。以DMSO (Merck) 溶解 CFSE 至 10 mM stock solution,分裝後置於-20℃備用(避光)。
Concanavalin A
購自Sigma。以 PBS buffer 調整濃度至 5 µg/ml 後保存於-20℃備用。
Polystyrene para-magnetic microparticles
具有磁性的聚苯乙烯磁珠購自Polysciiences Inc。直徑約 1~2 µm。
Synthetic CpG containing phosphorothioate oligodeoxyribonucleotides (CpG ODN) 購自Bio Basic Inc。其核苷酸序列如下表,CpG ODN 溶於 PBS 中後於 4℃下保 存。
CpG sequence
GCATAACGAGGAGCT
GpC sequence (reverse control of CpG) GCATAAGCAGGAGCT
2-1-5 緩衝溶液
PBS (phosphate buffer saline, 1L):
取NaCl (Merck ) 8g、Na2HPO4 (Merck ) 2.8g、KH2PO4 (Merck ) 0.2g、KCl (Merck ) 0.2g 加水至 1000 ml,調整pH至 7.0。
TE buffer (10×,1L) :
取Tris base (Sigma) 1.241g、EDTA (Sigma) 0.3g,加水至 1000ml,調整 pH 至 8.0。
電泳緩衝液 (electrophoresis buffer, 10×) :
Tris base (BIO-RAD) 30.3g、Glycine (BIO-RAD) 144.0g、SDS (BIO-RAD) 10g 加 水至 800 ml,pH 調至 8.3 後,加水至 1000 ml。
2-2 方法
2-2-1 DNA 純化
取900µl DNA solution 加入 100µl 10% SDS (Sigma),把 sample solution 置 於 65℃下加熱 30 分鐘。使用 1ml 針筒 (Terumo) 將溶液注入透析卡匣中 (Pierce),放置在裝有 1000 ml TE buffer 的燒杯於 4℃下進行透析 16 小時。在微 量離心管中裝入 DNA solution 加入等量 phenol : chloroform : isoamyl alcohol 25:21:1 (Sigma)。以 vortex 振盪混勻 1 分鐘後溶液呈現白色,以 700g 離心 5 分
鐘。將上層液移到新的微量離心管中同樣再加入等量phenol : chloroform : isoamyl alcohol 25:21:1 (Sigma),重複前述步驟直到白色消失。最後將上清液加入等量 chloroform 以 vortex 混勻 10 秒鐘,將上清液抽出加入 1/10 (v/v) 3 M ammonium acetate。在冰上加入 2 倍體積之 100% 冰冷酒精後置於冰上 20 分鐘。在 4℃下以 7000g 離心 10 分鐘,最後會得到 DNA pellet。將上清液移除後以 75%酒精清洗 2~3 次,進行抽乾約 30 分鐘,得到 DNA pellet 後溶解於 TE buffer 後,以分光光 度計偵測DNA 濃度。
DNA 濃度=O.D.260 nm × 50ng/m l × 稀釋倍數
2-2-2 鱸魚淋巴細胞之分離
從高雄縣永安鄉鱸魚養殖場直接購買新鮮的鱸魚,將取出鱸魚頭腎組織放到 含有10 ml L-15 培養液 (Gibco) 的離心管中,並馬上移到無菌操作台準備接下 來的實驗流程。
首先將頭腎組織置於70 m 細胞過濾器 (BD) 中,以滅菌後之 PBS buffer 沖洗數次,把70 m 細胞過濾器移到含 20 ml L-15 培養液 (Gibco) 的培養皿 中,然後以1 ml 針筒 (Terumo) 末端擠壓輕輕磨碎組織塊。分別將 10 ml 細胞懸 浮液置於15ml 離心管中,下層以 pasteur pitpet 注入 3 ml Histopaque 1.077 (Sigma) 後,於4℃下以 300g 離心 30 分鐘。離心後,以 pasteur pitpet 小心吸取位於培養 液與 Histopaque 液面間之白色懸浮細胞層(白血球) 到 15ml 離心管內,以 PBS buffer 清洗 3-5 次。再以含有 10% FCS (Gibco) 的 L-15 培養液 (Gibco) 10 ml 使 細胞懸浮。
2-2-3 細胞計數
取10 l 細胞懸浮液到微量離心管中,加入 10 l trypan blue (sigma) 混合。
取 10 l 細胞懸浮液(含trypan blue)到血球計數器中,用顯微鏡觀察計算活細胞 數量。以血球計數器計算細胞密度 (cell density) 後,以含有 10% FCS (Gibco) 的 L-15 培養液 (Gibco) 將細胞密度稀釋至 1×106 cells/ml於 20℃不含CO2培養。
2-2-4 鱸魚白血球細胞增生分析
將 頭 腎 細 胞 培 養 在 96 孔組織培養皿中,每一孔的細胞濃度是 1×106 cells/ml,每一孔加入 200 l 細胞懸浮液,進行三重複實驗,觀察鱸魚的白血球 對LPS, 10µg/ml,CpG ODNs, 5 µg/ml ;或GpC ODNs, 5 µg/ml,的反應反應,細胞
培養在20℃ 。培養 72 小時後加入Quick Cell Proliferation Assay Kit (BioVision),
每一孔加入20µl WST-1。WST-1 是一種類似於MTT的化合物,在電子耦合試劑 存在的情況下,可以被線粒體內的一些脫氫酶還原生成橙黃色的formazan(圖 2-2),細胞增殖越多越快,則顏色越深。WST-1 是MTT 的一種升級替代產品,
和MTT 或其它MTT 類似產品如XTT、MTS 等相比有明顯的優點。首先,MTT 被線粒體內的一些脫氫酶還原生成的formazan 不是水溶性的,需要有特定的溶 解液來溶解;而WST-1 和XTT、MTS 產生的formazan 都是水溶性的,可以省 去後續的溶解步驟。其次,WST-1 產生的formazan 比XTT 和MTS 產生的 formazan更易溶解。再次,WST-1 比XTT 和MTS 更加穩定,使實驗結果更加 穩定。而formazan 可使特定波長下的吸光值增加,之後每 24 小時測一次吸光值 OD450 (A450)。
圖2-3 .WST-1 檢測原理圖(EC=electron coupling reagent,即電子耦合試劑)。
2-2-5 鱸魚淋巴細胞存活率測試
使用Vybrant Apoptosis Assay Kit。Vybrant Apoptosis Assay Kit 可提供一個快 速且便利方式來測試細胞apoptosis。套組中包含有重組annexin V 結合到一個最 佳最明亮螢光染劑,Alexa Fluor® 488 染劑上,提供最大的靈敏度。此外,還包 含有一個紅色螢光 propidium iodide (PI) 核酸染劑。propidium iodide : 為一種特 殊的染劑,能嵌入雙鏈DNA 及 RNA 的鹼基對 (bases pair) 中,但其本身無法 通過細胞膜。加入二種染劑後,apoptotic 細胞會顯示綠色螢光,走向死亡細胞 會呈顯為紅色及綠色螢光,活的細胞會顯示很微量或沒有螢光。
從頭腎組織分離得到的白血球細胞懸浮在含有10% FCS 的L-15 培養液中,
細胞濃度 1×107cells/ml培養在 24 孔組織培養盤,每一孔加入 2ml細胞懸浮液。
細胞加入CpG ODNs, 5 g/ml 、GpC ODNs, 5 g/ml 刺激、及對照組共同培養在 20℃下,不含CO2.,培養72 小時。收集細胞懸浮液以 300g離心 10 分鐘得到cells pelleted。以 500µl PBS buffer清洗 2 次,加入 100 l annexin -binding buffer(1x)、
5µl Alexa Fluor 488 annexin V以及 1 l PI working solution,在室溫下培養 15 分 鐘,最後再加入400 l annexin -binding buffer (染色全程保持在冰上)。各別進行 流式細胞儀分析。
2-2-6 經由玻璃培養皿移除貼附性細胞
利用巨噬細胞與樹突狀細胞對培養容器表面吸附性,將巨噬細胞與B細胞 (sIgM+)、T細胞(sIgM-) 分開。從頭腎組織分離得到的白血球細胞懸浮在含有 10%
FCS 的L-15 培養液中,先取約 1ml 到folw 用管子中,將 20 ml 細胞懸浮液(1×107 cells/ml)置於玻璃培養皿中,以 20℃不含CO2培養 30 分鐘。30 分鐘後輕輕搖晃 培養皿,取出細胞懸浮液至另一個新的玻璃培養皿中繼續30 分鐘。30 分鐘後同 樣輕輕搖晃培養皿,先取約1ml細胞液到到flow用管子中後,剩餘懸浮液再次培 養在另一個新的玻璃培養皿中30 分鐘。30 分鐘後取出懸浮液,總共收集 3 管細 胞液以流式細胞儀分析。
2-2-7 經由磁珠分離吞噬性細胞
利用吞噬性細胞對外來物質之吞噬性,將巨噬細胞與B細胞 (sIgM+)、T細胞 (sIgM-) 分開。先用PBS buffer清洗磁珠數次,加入細胞(1×106 cells/ml) 與磁珠 1:30 比例混合均勻充分接觸,置於 20℃下靜置 3 小時,每 30 分鐘震盪細胞與磁 珠,使之混合均勻。3 小時後使用磁鐵貼附於管壁上,慢慢將會吞噬磁珠之吞噬 細胞吸附到管壁上,再將其他剩下不會被吸附至管壁上細胞收集到另一個新的微 量離心管中。接著就將這二種細胞分別以流式細胞儀分析。
2-2-8 收集上清液 (Conditioned medium) 2-2-8-1 收集頭腎細胞上清液
從頭腎組織分離得到的白血球細胞懸浮在含有10% FCS 的L-15 培養液中,
細胞濃度1×107 cells/ml培養在 24 孔組織培養盤,每一孔加入 2ml細胞懸浮液。
細胞加入CpG ODNs, 5 µg/ml、GpC ODNs, 5 µg/ml刺激、及對照組共同培養在
20℃下,經過 48 小時各別收集細胞,離心 300g 離心 10 分鐘,移除上清液,再 次加入新鮮培養液,使細胞濃度同樣調整在1×107 cells/ml,在新的 24 孔組織培 養盤中每一孔加入2ml細胞懸浮液。培養在 20℃下,經過 24 小時後收集細胞上 清液,分裝,凍於-20℃下備用。
2-2-8-2 上清液之濃縮
從頭腎組織分離得到的白血球細胞懸浮在L-15 培養液中,細胞濃度 1×107 cells/ml 培養在 24 孔組織培養盤,每一孔加入 2ml 細胞懸浮液。細胞加入CpG ODNs, 5 µg/ml、GpC ODNs, 5 µg/ml刺激、及對照組共同培養在 20℃下,經過 72 小時的培養後收集細胞上清液。首先於4℃下以 300g離心 10 分鐘,移除細胞後 取出上清液至 10k蛋白質濃縮過濾離心管(Millipore) 再次於 4℃下以 4000g離心 30 分鐘,分裝後放於-20℃冷凍保存。
2-2-9 Carboxyfluorescein diacetate, succinimidyl ester (CFDA, SE) 染色
CFSE (Invitrogen) 粉末溶解在DMSO(Sigma)中以製備 10mM CFSE stock solution保存在-20℃並且避免光線照射。將剛分離得到的白血球細胞懸浮在 10ml 含有10mM CFSE的PBS中靜置培養 15 分鐘,加入冰冷的 10ml PBS停止反應,
300g離心 5 分鐘得到cells pelleted再重新懸浮在L-15 培養液(含有 10% FCS),把 細胞培養在24 孔培養盤,每一孔加入 1ml細胞懸浮液。細胞加入LPS, 10 g/ml 、 CpG ODNs, 5 g/ml 或GpC ODNs, 5 g/ml 刺激,及對照組共同培養在20℃下,
不含CO2。培養24、48、72 小時後以流式細胞儀分析。
圖2-4. CFSE 染色作用機制。
2-2-10 T cells 鑑別
從CFSE 染色,流式細胞儀分析結果,得知在淋巴細胞族群中有一群細胞 是不進行分裂,另一群細胞會隨著培養時間而分裂。為此我們想知道會進行分裂
細胞族群為何,所以使用Concanavalin A。Concanavalin A 可快速使 T 細胞族群 分裂,因此當加入Concanavalin A 後進行流行細胞儀分析就可得知會進行細胞 分裂的族群是否為T 細胞。
從頭腎組織分離得到的白血球細胞懸浮在含有10% FCS 的L-15 培養液中,
細胞濃度1×107 cells/ml 培養在 24 孔組織培養盤,每一孔加入 2ml細胞懸浮液。
細胞加入Concanavalin A 5 g /ml (Sigma) 刺激、及對照組共同培養在 20℃下,
不含CO2.。各別培養24、48、72 小時後,分別收集三次細胞以流式細胞儀分析。
2-2-11 上清液活性測試
將頭腎細胞培養在96 孔組織培養皿中,每一孔的細胞濃度是 1×106
cells/ml。將經由 (方法 2-2-7-1) 收集上清液放置於 95-100℃中加熱約 10 分鐘,
使蛋白質變性。每一孔再加入100 l 由 (方法 2-2-7-1) 收起來的細胞上清液,進 行三重複實驗。細胞培養在20℃。培養 72 小時後加入Quick Cell Proliferation Assay Kit (BioVision),每一孔加入 20µl WST-1,之後每 24 小時測一次吸光值 OD450 (A450)。
2-2-12 第二維電泳-十二烷硫酸鈉-聚丙烯醯胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDS-PAGE)
SDS-PAGE 的分離原理是利用 SDS 與蛋白質作用使蛋白質帶電荷數趨於 一致,因而使其移動速率完全由蛋白質分子量的大小決定。為了使蛋白質能完全 伸張,常會添加打斷雙硫鍵的還原劑-如 -硫氫乙醇 (β-mercapethanol, -ME) 或1,4-dithiothreitol 等。SDS 凝膠中有許多的孔洞,而蛋白質在強力電場的驅 動下會在凝膠中的孔洞泳動,大分子量的蛋白質移動速度比較慢,小分子量的蛋 白質則相反,所以蛋白質會因分子量大小不同而在凝膠上分離開來,如此一來可 將等電點相同或接受的蛋白質做進一步的分離。
先將電泳裝置裝好,再依表2-2上的配方,配製兩種不同濃度PAGE:8% 及 10% SDS-PAGE。先將 resolution gel 注入,小心注入 stacking gel 且應避免氣 泡產生,再裝上齒梳待其凝固後。待 stacking gel 凝固後,抽掉齒梳成形器,注 入1倍 running buffer 於電泳槽中。取CpG ODN 與細菌 DNA 處理後頭腎細胞 培養液之濃縮液中之蛋白質先與2倍 sample buffer (表2-1)混合均勻,於95℃下加
熱5分鐘;置於冰上5分鐘,將樣品注入上層凝膠的凹槽中。接著蓋上電泳槽蓋子 (樣品應從負極至正極),設定電泳條件為150 V 跑約 70分鐘。將蛋白質依分子 量大小不同作分離;接著以硝酸銀套組 (GE Healthcare) 進行染色染出膠片上之 蛋白質,與對照組相較找出新出現或表現量提高之蛋白質點。
表2-1. Sample buffer 1.25 ml 0.5 M Tris-HCl, pH 6.8
2.0 ml 10% (w/v) SDS 2.5 ml Glycerol
0.2 ml 0.5% Bromophenol blue (BPB), trace 3.55 ml Distilled water
9.5 ml Total volume
使用時取50µl β-Mercaptoethanol ( -ME)加入 950 l sample buffer
表3-2. SDS-PAGE (95)
Separating Gel 5% Stacking Gel Reagent
8% 10%
ddH2O 6.9 ml 5.9 ml 2.1 ml 30% Aceylamide mix 4.0 ml 5.0 ml 0.5 ml 1.5 M Tris (pH8.8) 3.8 ml 3.8 ml - 0.5 M Tris (pH6.8) - - 0.38 ml 10% SDS 0.15 ml 0.15 ml 0.03 ml 10% Ammonium persulfate 0.15 ml 0.15 ml 0.03 ml TEMED 0.009 ml 0.006 ml 0.003 ml
總體積 15ml 15ml 3ml
2-2-13 硝酸銀染色 (Silver stain)
銀染是一種靈敏度極高的蛋白質染色法 (偵測極限為每色帶1-10ng)。主要的 原理為蛋白質上的羧基 (-COO-) 會與銀銨錯合離子結合,而銀離子在酸性環 境下會被還原成金屬銀,所以可以使凝膠上的蛋白質呈現黑褐色,藉此我們就可 以在膠片上看到蛋白質的存在與分佈情況。
每一片膠片所需的溶液體積為50 ml,所有步驟都要在迴轉式震盪器上面搖 晃(表3-3)。電泳後將 SDS-膠片取出浸泡在 fixation solution 中30分鐘。倒掉後 加入sensitizing solution 浸泡30分鐘,接著以 ddH2O 清洗三次各5分鐘。隨後以 silver reaction 浸泡20分鐘,之後以 ddH2O 快速洗滌二次各1分鐘。再倒入 developing solution 浸泡5分鐘等待反應,當膠片呈現清晰的顏色時加入stopping solution 浸泡10分鐘以停止反應,以 ddH2O 清洗三次各5分鐘。
表2-3. 硝酸銀染色 (Silver stain)之配方 Fixation solution 95% Methanol
Acetic acid glacial Sensitizing solution 95% Methanol
Glutardialdehyde (25% w/v) Sodium thiosulphate (5% w/v) Sodium acetate (17g)
Silver reaction Silver nitrate solution (2.5% w/v) Formaldehyde (37% w/v)
Developing solution Sodium carbonate (6.25g) Formaldehyde (37% w/v) Stopping solution EDTA-Na.2H2O (3.65g) Preserving solution 95% Methanol
Glycerol (87% w/v)
三、 結果 3-1 鱸魚頭腎細胞最適生長條件
首先要找出金目鱸魚頭腎細胞最適合生長細胞濃度及培養條件。因此依據
(94)這篇研究來對鱸魚生長條件做適當調整。 一開始先選擇三種不同細胞數 : 1×106cells/ml、5×105cells/ml 、2.5×105cells/ml;以及二種不同培養時間,分別為 48 小時及 72 小時。
以三種免疫刺激因子,LPS、合成之CpG ODN及合成之GpC ODN來刺激不 同細胞數之頭腎細胞48 小時後以WST-1 偵測在第 24、48、72 小時吸光值 450nm 下細胞增生情況。當細胞數為 1×106cells/ml,以LPS刺激細胞時與控制組比較並 沒有明顯差異;而以合成之CpG ODN刺激細胞時與控制組比較有明顯細胞增生 情形 (p < 0.05, p < 0.01);而對照組GpC ODN則無任何反應如圖3-1(a)。而 當細胞數為5×105cells/ml,以LPS、合成之CpG ODN及對照組GpC ODN刺激細 胞時與控制組比較都沒有明顯的任何反應如圖3-1(b)。當細胞數2.5×105cells/ml,
以LPS、合成之CpG ODN及對照組GpC ODN刺激細胞時與控制組比較都沒有明 顯的任何反應如圖3-1(c)。
以三種免疫刺激因子,LPS、合成之CpG ODN及合成之GpC ODN來刺激不 同細胞數之頭腎細胞72 小時後以WST-1 偵測在第 24、48、72 小時吸光值 450nm 下細胞增生情況。當細胞數 1×106cells/ml,可明顯得知以LPS刺激細胞時與控制 組比較有明顯細胞增生情形 (p < 0.01);以合成之CpG ODN刺激細胞時與控制 組比較也有明顯細胞增生情形 (p < 0.05, p < 0.01);而對照組GpC ODN則無 任何反應如圖3-2(a)。而當細胞數5×105cells/ml,當以LPS刺激細胞時與控制組 比較並沒有明顯差異;而以合成之CpG ODN刺激細胞時與控制組比較有明顯細 胞增生情形 (p < 0.05, p < 0.01);而對照組GpC ODN則無任何反應如圖 3-2(b)。細胞數2.5×105cells/ml,當以LPS、合成之CpG ODN及對照組GpC ODN 刺激細胞時與控制組比較都沒有明顯的任何反應如圖3-2(c)。
總結在三種不同細胞數 : 1×106cells/ml、5×105cells/ml、2.5×105cells/ml,以 細胞數1×106cells/ml不管是培養在 48 或 72 小時下以CpG ODN刺激都可以成功 促使細胞增生。而培養在48 小時及 72 小時,以細胞數 1×106cells/ml刺激都可以
促使細胞增生,但培養在72 小時下細胞增生效果為較好。因此往後實驗都選擇
將頭腎細胞以細胞數1×106cells/ml培養在 72 小時的條件下來測試細胞增生情況。
3-2 培養在不同培養液時對頭腎細胞增生效果
在實驗過程中發現當在頭腎細胞培養液中加入10% 胎牛血清時,所收集得 到上清液進行蛋白質電泳分析會造成結果產生一些由胎牛血清中所產生的蛋白 質而影響實驗上觀察。因此試著在頭腎細胞培養液中不添加入胎牛血清,直接以 L-15 培養液在培養頭腎細胞,測試頭腎細胞是否依然可以進行細胞增生。當頭 腎細胞培養在含有胎牛血清培養液時以合成之CpG ODN 刺激 72 小時後,結果 可以成功刺激頭腎細胞分裂,而對照組GpC ODN 則沒有反應,與控制組比較也 有顯著差異 (*p<0.05),如圖3-3(a)所示。而當頭腎細胞培養在不含有胎牛血清 培養液時以合成之CpG ODN 刺激 72 小時後,結果顯示與控制組比較,雖然效
果沒有比有添加胎牛血清佳,但依舊可以成功刺激頭腎細胞分裂,而對照組GpC
ODN 則沒有反應,圖3-3(b)所示。
由此結果得知,就算在細胞培養液中沒有添加胎牛血清依照可以使頭腎細胞 進行細胞分裂。因此之後收集要進行蛋白質電泳分析時之上清液,頭腎細胞培養 液就不添加胎牛血清。
3-3 鱸魚頭腎細胞增生分析
當所有適合培養鱸魚頭腎細胞條件建立完成後,就可進行測試CpG ODN 是 否能促使頭腎細胞增生分析。以三種免疫刺激因子,LPS、合成之 CpG ODN 及 合成之GpC ODN 來刺激頭腎細胞,72 小時後以 WST-1 偵測在吸光值 450nm 下細胞增生情況。結果如圖3-4所示。
由圖中可明顯得知當以LPS 及合成之 CpG ODN 刺激細胞時與控制組比較 有明顯細胞增生情形 (p < 0.05);而對照組 GpC ODN 則無任何反應。
由此結果可得知以合成之CpG ODN 可成功的刺激頭腎細胞增生。
3-4 使用 Vybrant Apoptosis Assay Kit 染色法來鑑別出存活、apoptotic 及死亡細胞 族群。
為了得知頭腎細胞與CpG ODN 共同培養 24、48、72 小時後,細胞是否為 存活狀態或是已經走向apoptosis,所以使用Vybrant Apoptosis Assay Kit 來測試 細胞生長狀況,結果如圖3-5、6、7、8、9、10所示。
由圖中可得知當頭腎細胞與CpG ODN 共同培養不管是在 24、48 或 72 小時後,
細胞存活率依舊良好。在控制組中存活細胞大約有85%,走向apoptosis 大約有 10.45%,而死亡細胞大約只有 4.29%;在以合成之 CpG ODN 刺激過後頭腎細胞 中存活細胞大約87%、走向apoptosis 大約有 8.24%,而死亡細胞大約只有 4.2%。
3-5 流式細胞儀分析來鑑別
以用流式細胞儀分析金目鱸魚細胞組成其結果如圖3-11(a) 所示。R1 為淋巴 球細胞包括有T 和B 淋巴細胞(96) 佔59.31% ; R2 為neutrophils, macrophages 及 monocytes (97, 98) 佔全部細胞9.89%,呈現典型之細胞亞群組成。
之前實驗已經證實了合成之CpG ODN 能夠成功的刺激頭腎細胞分裂,而作 為對照組的 GpC ODN 則無任何反應(圖 3-4)。所以進一步探討此細胞分裂現是 否為CpG ODN 刺激巨嗜細胞所產生的 cytokines,而進一步去促使 T 細胞分裂 所造成呢?因此嘗試以二種方法來測試是否可以順利去除吞噬性細胞。
3-5-1 使用玻璃培養皿
根據研究(94) 發現使用玻璃材質培養皿較適合讓巨嗜細胞貼附上去,因此也
試著使用玻璃培養皿來測試是否可以有效的去除巨嗜細胞,結果如圖3-11所示。
將頭腎細胞培養在玻璃培養皿中每間隔30 分鐘更換一次玻璃培養皿,總共
更換3 次。90 分鐘後收集在玻璃培養皿上頭腎細胞液及沖洗上貼附在培養皿上 細胞液,經由流式細胞分析結果得知R2 之 macrophage、neutrophils、及 monocytes 族群由原本9.89% 降至為 8.59%,而貼附在玻璃培養皿上之 R2 族群有 14.79%。
結果顯示大約會有1/3 的 R2 細胞亞群為巨嗜細胞,而使用玻璃培養皿也的確可 以成功將巨噬細胞移除。
3-5-2 使用吸附性磁珠
使用1:30 比例將頭腎細胞與磁珠混合均勻,使其充分接觸後將具吞噬性細 胞去除,結果如圖3-12所示。頭腎細胞與磁珠作用3 小時後,由流式細胞分析 結果得知R2 之 macrophage、neutrophils 及 monocytes 族群由原本的 31.64% 降 為22.94%。結果顯示大約有 1/4 到 1/3 的 R2 細胞亞群為吞噬性細胞,而使用磁 珠可以成功將其移除。
綜合以上二種測試方法,結果都可以順利將巨嗜細胞移除。而往後實驗為了 方便性所以採用使用磁珠來去除巨嗜細胞。
3-6 將巨噬細胞移除後對頭腎細胞增生影響
確定了使用磁珠確實可以去除巨噬細胞後,將頭腎細胞中巨噬細胞移除後以 CpG ODN 刺激頭腎細胞,測試是否依然會有細胞增生情況。
以合成之CpG ODN 及合成之 GpC ODN 來刺激移除巨噬細胞之頭腎細 胞,72 小時後以 WST-1 偵測在吸光值 450nm 下細胞增生情況。結果如圖3-13 所示。
由圖中可明顯得知當以合成之CpG ODN 及對照組 GpC ODN 刺激移除巨 噬細胞之頭腎細胞時,與控制組比較並無任何明顯增生效果。
由此結果可得知當將巨噬細胞移除後以合成之CpG ODN 刺激細胞時並無 任何反應,因此可以推測當以CpG ODN 刺激頭腎細胞增生時是需要有巨噬細胞 存在。
3-7 收集 CpG ODNs 刺激過後上清液對頭腎細胞增生影響
CpG ODNs 並不是直接去誘導促使細胞增生,而是促使細胞產生細胞促進 因子 (PIFs) 作用到標的細胞上而使細胞增生。所以需要收集由 CpG ODNs 刺激 頭腎細胞後所釋出的cytokines 來測試到底細胞促進因子是作用到哪個細胞身上 呢?分別進行三組實驗來找出是由哪個細胞所產生cytokines?及作用到哪個細 胞上。
第一組:將頭腎細胞以合成之CpG ODN 及合成之 GpC ODN 刺激頭腎細 胞24 小時後以 L-15 培養液清洗 3 次將其剩餘 ODN 移除後,再次把頭腎細胞培 養在24 孔盤中 48 小時後收集其上清液。由 CpG ODN 及 GpC ODN 刺激過後之 上清液與頭腎細胞共同培養72 小時後以 WST-1 偵測在吸光值 450nm 下細胞增 生情況。結果如圖3-14所示。
由圖中可明顯得知當以合成之CpG ODN 刺激過後之上清液與控制組之上 清液比較可明顯得知以CpG ODN 刺激過後之上清液可明顯促使細胞增生( p < 0.05);而對照組 GpC ODN 刺激過後之上清液則無任何反應。
由此結果可得知CpG ODN 刺激過後之上清液可成功的促使頭腎細胞增生。
第二組:將頭腎細胞以合成之CpG ODN 及合成之 GpC ODN 刺激頭腎細胞 24 小時後以 L-15 培養液清洗 3 次將其剩餘 ODN 移除後,再次把頭腎細胞培養 在24 孔盤中 48 小時後收集其上清液。由 CpG ODN 及 GpC ODN 刺激過後之上 清液與將巨噬細胞移除後之頭腎細胞共同培養72 小時後以 WST-1 偵測在吸光值 450nm 下細胞增生情況。結果如圖3-15所示。
由圖中可明顯得知不管當以合成之CpG ODN 或對照組 GpC ODN 刺激過後 之上清液與控制組之上清液比較都無任何反應。由此結果可得知當將巨嗜細胞移 除後以CpG ODN 刺激過後之上清液仍無法促使細胞增生。
第三組:將已把巨噬細胞移除之頭腎細胞以合成之CpG ODN 及 GpC ODN 刺激頭腎細胞24 小時後以 L-15 培養液清洗 3 次將其剩餘 ODN 移除後,再次把 頭腎細胞培養在24 孔盤中 48 小時後收集其上清液。由 CpG ODN 及 GpC ODN 刺激過後之上清液與頭腎細胞共同培養72 小時後以 WST-1 偵測在吸光值 450nm 下細胞增生情況。結果如圖3-16所示。
由圖中可明顯得知不管當以合成之CpG ODN 或對照組 GpC ODN 刺激過後 之上清液與控制組之上清液比較都無任何反應。由此結果可得知當將巨噬細胞移 除後以CpG ODN 刺激所收集得到上清液仍無法促使頭腎細胞增生。
由以上三組實驗可得到三個小結論。
以CpG ODN 刺激所收集得到上清液 + 頭腎細胞 → 細胞增生
以CpG ODN 刺激巨嗜細胞移除所收集得到上清液 + 頭腎細胞 →無反應 以CpG ODN 刺激所收集得到上清液 + 巨嗜細胞移除 →無反應
因此可以得知促進細胞分裂因子是由具吞噬性細胞 (phagocytes) 所分泌,
而吞噬性細胞 (phagocytes)可能扮演著 accessory cells 角色去促使 T 細胞增生。
3-8 使用 CFSE 染色法來鑑別增生之頭腎細胞族群
由之前實驗得知當以CpG ODN 刺激頭腎細胞時會促使細胞增生。為此想要 尋找出CpG ODN 主要是誘使哪個細胞族群增生呢?所以使用 CFSE 染色法來鑑 別出增生細胞族群。結果如圖3-17所示。
當CFSE 染色後以 CpG ODN 刺激結果由圖3-17(a)、(b)可明確得到二個細胞 族群Lymphocytes (R1)及 Macrophages (R2)。
當CFSE 染色後以 CpG ODN 刺激 24 小時結果由圖3-17(c)、(d)可明確得到 在Lymphocytes 中分裂成 R1 及 R3 二個族群;而在 Macrophages (R2)中並沒有 進行細胞分裂。
當CFSE 染色後以 CpG ODN 刺激 48 小時結果由圖3-17(e)、(f)可明確得知 在Lymphocytes 中也同樣分裂成 R1 及 R3 二個族群;而在 Macrophages (R2) 中 並沒有進行細胞分裂。
由以上三個結論得知當CFSE 染色後以CpG ODN 刺激 24~48 小時,在 Lymphocytes 族群中會進行細胞分裂,但究竟進行細胞分裂者是T細胞還是B 細 胞目前不能給予肯定答案,但由(94) 報告中指出B 細胞不是扮演著產生cytokines 角色也不是接受cytokines 刺激而促使細胞增生角色,可以發現在Lymphocytes族 群中有一群細胞會進行細胞分裂。而在Macrophages族群中不管是從一開始以 CpG ODN刺激或刺激 48 小時結果來看Macrophages族群都不會進行細胞分裂,
這個結果與(94) 在鳟魚頭腎細胞上觀察得到結果大不相同,如圖3-18所示。
3-9 使用 Concanavalin A 來刺激頭腎細胞測試分裂族群是否為 T 細胞
從CFSE 染色結果(圖3-17)得知在淋巴細胞族群中有一群細胞是不進行分 裂,另一群細胞會隨著培養時間而分裂。為此想知道會進行分裂細胞族群為何,
所以使用Concanavalin A。Concanavalin A 可快速使 T 細胞族群分裂,因此當 CFSE 染色後以 Concanavalin A 刺激結果由圖3-19所示。
由圖中僅可得知Lymphocytes 族群中的確是有一族群細胞是會進行細胞分裂,
可以無法從這方法來準確判定會進行細胞分裂族群是T 細胞。
3-10 以蛋白質變性方式來測試收集上清液中是否為蛋白質
為了確認收集得到上清液是否真為蛋白質,所以將其上清液以加熱方式讓蛋 白質變性後,再次去刺激頭腎細胞測試是否可讓細胞增生。
將頭腎細胞以細胞刺激因子LPS、合成之 CpG ODN 及合成之 GpC ODN 刺 激頭腎細胞24 小時後以 L-15 培養液清洗 3 次,再次把頭腎細胞培養在 24 孔盤 中48 小時後收集其上清液,收集的上清液以 95℃加熱使其蛋白質變性。加熱變 性後蛋白質再與頭腎細胞共同培養72 小後以 WST-1 偵測在吸光值 450nm 下細 胞增生情況。結果如圖3-20所示。
由圖中可明顯得知當以LPS、合成之 CpG ODN 及對照組 GpC ODN 所收集 得到之上清液刺激頭腎細胞時,與控制組比較並無任何明顯增生效果。 因此可 以確定所收集得到之上清液是為蛋白質。
3-11 以 SDS-PAGE 來分析金目鱸魚頭腎細胞上清液
以合成CpG ODNs、細菌 DNA 刺激頭腎細胞培養 72 小時離心移除細胞後 取出上清液至10k 蛋白質濃縮過濾離心管再次離心,得到上清液之濃縮液進行蛋 白質電泳分析。設定電泳條件為150 V 跑約 70 分鐘。將蛋白質依分子量大小不 同作分離;接著以硝酸銀套組進行染色染出膠片上之蛋白質。結果如圖 3-21 所 示。
由圖中可觀察得知: Lane A 與 Lane C 比較,可明顯發現 Lane C 比 Lane A 多 出三組不同的bead,根據 marker 比對大概可以推測可能是 IL-1、TNF- 或 IL-12。
四、 討論
在宿主防禦 (host defence) 機制演化的過程中,自軟骨魚類始出現適應性免疫系 統 (adaptive immunity),因此科學家可藉由各種疫苗之開發,幫助養殖戶降低因 魚病所造成的損失。然而對魚類免疫系統之研究,始終受限於對魚類免疫系基礎 知識之不足與相關試劑之缺乏而進展有限,對於魚病之控制依舊停留在隔離病源 與施用抗生素等傳統方法。近年來由於人類基因體計劃的完成與基因體學的進 展,河豚與斑馬魚之基因體已先後完成定序,對於魚類免疫學的進展提供相當的 助力,魚類CD4、CD8、TCR、MHC-II 等免疫相關分子的基因亦相繼被定序出 來。然而在市場上依然買不到可供辨識B 細胞與 T 細胞之單株抗體,因此本研 究選用南台灣常見的養殖魚種之一尖吻鱸 (Lates calcarifer) 為實驗材料,轉而依 白血球之吸附性與功能之不同,意圖自鱸魚頭腎細胞中分離出具吞噬作用的細胞
如嗜中性球與巨噬細胞。
本研究結果顯示經由 CpG ODNs 刺激吞噬性細胞後會分泌出促使細胞增生 因子,而實驗也証實這些促使細胞增生因子為蛋白質,同時也證實了促使細胞增 生因子可以誘導在淋巴細胞中某一族群的增生。此外,吞噬性細胞在此反應中也 同時扮演如同是accessory cells 角色。
而由結果圖3-21 可以推斷使用 CpG ODN 可以刺激頭腎細胞產生一些促使 細胞分裂因子,如IL-1、IL-12 或是 TNF- 。
參考文獻
1. Tokunaga T, Yamamoto H, Shimada S, Abe H, Fukuda T, Fujisawa Y.
Antitumor-activity of deoxyribonucleic-acid fraction from Mycobacterium-bovis BCG 1 isolation, physicochemical characterization, and antitumor-activity. J Nat Cancer Inst. 1984; 72: 955-962.
2. Tokunaga T, Yamamoto S, Namba K. A synthetic single-stranded DNA, poly (dG,dC), induces interferon- / and – , augments natural killer activity, and suppresses tumor growth. Jpn J Cancer Res. 1998; 79:682-686.
3. Messina J, Gilkeson G, Pisetsky D. Stimulation of in vitro murine lymphocyte proliferation by bacterial DNA. J Immunol. 1991; 147:1759-1764.
4. Yamamoto S, Yamamoto T, Kataoka T, Kuramoto E, Yano O, Tokunaga T.
Unique palindromic sequences in syntheticoligonucleotides are required to induce IFN and augment IFN-mediated natural killer activity. J Immunol. 1992;
148:4072-1976.
5. Krieg A, Yi A, Matson S, Waldschmidt T, Bishop G, Teasdale R. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995; 374:546-549.
6. Krieg A. Mechanisms and applications of immune stimulatory CpG oligodeoxynucleotides. Biochim Biophys Acta. 1999; 1489:107-116.
7. Bird A. CpG islands as gene markers in the vertebrate nucleus. Trends Genet.
1987; 3:342-347.
8. Karlin S, Doerfler W, Cardon L. Why is CpG in the genomes of virtually all small eukaryotic viruses but not in those of large eukaryotic viruses? J Virol. 1994;
68:2889-2897.
9. Krieg A, Yi A, Matson S, Waldschmidt T, Bishop G, Teasdale R. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995; 374:546-549.
10. Krieg A, Wu T, Weeratna R, Efler S, L L-H, Yang L. Sequence motifs in adenoviral DNA block immune activation by stimulatory CpG motifs. Proc Natl Acad Sci USA. 1998; 95:12631-12636.
11. Krieg A, Wagner H. Causing a commotion in the blood: immunotherapy progresses from bacteria to bacterial DNA. Immunol Today. 2000; 21:521-526.
12. Krieg A. Now I know my CpGs. Trend Immunol. 2001;9:249-252.
13. Verthelyi D, Ishii K, Takeshita F, Klinman D. Human peripheral blood cells differentially recognize and respond to two distinct CpG motifs. J Immunol. 2001;
166:2372-2377.
14. Ballas Z, Rasmussen W, Krieg A. Induction of NK activity in murine and human cells by CpG motifs in oligodexynucleotides and bacterial DNA. J Immunol. 1996;
157:1840-1845.
15. Hartmann G, Krieg A. Mechanism and function of a newly identified CpG DNA motif in human primary B cells. J Immunol. 2000; 164:944-952.
16. Chace J, Hooker N, Mildenstein K, Krieg A, Cowdery J. Bacterial DNA-induced NK cell IFN- production is dependent on macrophage secretion of IL-12. Clin Immunol Immunopathol. 1997; 84:185-193.
17. Yi A, Chace J, Cowdery J, Krieg A. IFN-g promotes IL-6 and Ig M secretion in response to CpG motifs in bacterial DNA and oligodeoxynucleotides. J Immunol.
1996; 156:558-564.
18. Redford T, Yi A, Ward C, Krieg A. Cyclosporin A enhances IL-12 production by CpG motifs in bacterial DNA and synthetic oligodxynucleotides. J Immunol. 1998;
161:3930-3935.
19. Lipford G, Bauer M, Blank C, Reiter R, Wagner H, Weeg K. CpG-containing synthetic oligonucleotides promote B and cytotoxic T cell reponses to protein antigen: a new class of vaccine adjuvants. Eur J Immunol. 1997; 27:2340-2344.
20. Sparwasser T, Koch E, Vabulas R, Heeg K, Lipford G, Ellwart J. Bacterial DNA and immunostimulatory CpG oligonucleotides trigger maturation and activation of murine dendritic cells. Eur J Immunol. 1998; 28:2045-2054.
21. Hemmi H, Takeuchi O, Kawai T, et al. A Troll-like receptor recognizes bacterial DNA Nature. 2000; 408:29-43.
22. Krieg A. CpG motifs in bacterial DNA and their immune effects. Annu Rev Immunol. 2002; 20:709-760.
23. Yi A, Chang M, Peckham D, Krieg A, Ashman R. CpG oligodeoxyribonucleotides rescue mature spleen B cells from spontaneous apoptosis and promote cell cycle entry. J Immunol. 1998; 160:5898-5906.
24. Macfarlane D, Manzl L. Antagonism of immunostimulatory CpG-oligodeoxynucleotides by quinacrine, chloroquine, and structurally related compounds. J Immunol. 1998; 160:1122-1131.
25. Hans H, Ramunas MV, Osamu T, Katsuaki H, Shizuo A, Hermann W. Immune Cell Activation by Bacterial CpG-DNA through Myeloid Differentiation Marker 88 and Tumor Necrosis Factor Receptor–Associated Factor (TRAF)6 J Exp Med.
2000;192:595-600.
26. Hans H, Harald M, Thomas M, et al. CpG-DNA-sprific activation of antigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation. EMBO J. 1998;
21:6230-6240.
27. Hartmann G, Krieg A. Mechanism and function of a newly identified CpG DNA motif in human primary B cells. J Immunol. 2000; 164:944-952.
28. Tomokazu T, Hidehiro K, Ikuo H, Makoto E, Tatsuo Saito T, Takashi A.
Molecular cloning and characterization of Toll-like receptor 9 in Japanese flounder, Paralichthys olivaceus. Mol Immunol. 2007; 44:1845-1853.
29. Ken J I, Shizuo A. Innate immune recognition of, and regulation by, DNA. Trends Immunol. 2006;27(11):525-532.
30. Anderson DP. Immunostimulants, adjuvants, and vaccine carriers in fish:
Applications to aquaculture. Annual Review of Fish Diseases. 1992; 2:281-307.
31. Hans H, Harald M, Thomas M, et al. CpG-DNA-sprific activation of antigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation. EMBO J. 1998;
21:6230-6240.
32. Secombes C, Clements K, Ashton I, Rowley A. The effect of eicosanoids on rainbow trout, Oncorhynchus mykiss, leucocyte proliferation. Vet Immunol Immunopathol. 1994;42(3-4):367-378.
33. Hansen J. Characterization of rainbow trout terminal deoxynucleotidyl transferase structure and expression. TdT and RAG1 co-expression define the trout primary lymphoid tissues. Immunogenetics. 1997; 46(5):367-375.
34. Hansen J, Kaattari S. The recombination activation gene 1 (RAG1) of rainbow trout (Oncorhynchus mykiss): cloning, expression, and phylogenetic analysis.
Immunogenetics. 1995; 42 (3):188-195.
35. Willett C, Cherry J, Steiner L. Characterization and expression of the recombination activating genes (rag1 and rag2) of zebrafish. Immunogenetics.
1997; 45(6):394-404.
36. Willett C, Cherry J, Steiner L. Early hematopoiesis and developing lymphoid organs in the zebrafish. Dev Dyn. 1999; 214(4):323-336.
37. Perrot V, Moiseeva E, Gozes Y, Chan S, Ingleton P, Funkenstein B. Ontogeny of the insulin-like growth factor system (IGF-I, IGF-II, and IGF-1R) in gilthead
seabream (Sparus aurata): Expression and cellular localization. Gen Comp Endocrinol. 1999; 116(3):445-460.
38. 林式修。魚類免疫與疫苗。水產專訊。2003; 3:14-18。
39. Ellis AE. Meeting the requirements for delayed release of oral vaccines for fish. J.
Appl. Ichthyol. 1998; 14(3-4 ):149-152.
40. Fishelson L. Cytological and morphological ontogenesis and involution of the thymus in cichlid fishes (Cichlidae, Teleostei). Journal of Morphology. 1995;
223:175-190.
41. Willett C, Zapata A, Hopkins N, Steiner L. Expression of zebrafish rag genes during early development identifies the thymus. Dev Biol. 1997; 182(2):331-341.
42. Fange R. Lymphoid organs in sturgeons (Acipenseridae). Vet Immunol Immunopathol. 1986; 12:153-161.
43. Secombes C, Manning M. Comparative studies on the immune system of fishes and amphibians. J Fish Dis. 1980; 3(5):399-412.
44. Dalmo R, Ingebrigtsen K, Bogwald J. Non-specific defence mechanisms in fish, with particular reference to the reticuloendothelial system (RES). J Fish Dis. 1997;
20(4): 241-273.
45. Navarro V, Quesada J. Immunochemical characterization of monoclonal antibodied to gilthead seabreami (Sparus aurate) immunoglobulin. Fish Shellfish Immunol. 1993; 3:167-177.
46. Pilstrom L, Petersson A. Isolation and partial characterization of immunoglobulin from cod (Gadus morhua L.). Dev Comp Immunol. 1991; 15 143-152.
47. Ram S, Masahiro S. Genomics of fish cytokines. Comp Biochem Physiol.
2006:89-101.
48. Zou J, Cunningham C, Secombes C. The rainbow trout Oncorhynchus mykiss interleukin-1 beta gene has a differ organization to mammals and undergoes incomplete splicing. Eur J Biochem. 1999; 259:901-908.
49. Pelegrin P, Garcia-Castillo J, Mulero V, Meseguer J. Interleukin- 1beta isolated from a marine fish reveals up-regulated expression in macrophages following activation with lipopolysaccharide and lymphokines. Cytokine. 2001; 16:67-72.
50. Scapigliati G, Buonocore F, Bird S, et al. Phylogeny of cytokines: molecular cloning and expression analysis of sea bass Dicentrarchus labrax interleukin-1beta.
Fish Shellfish Immunol. 2001; 11:711-726.