The Role of Apurinic/apyrimidic endonuclease DNA-repair gene in Endometriosis
Chin-Mu Hsu1,*, Wen-Shin Chang1,2,*, Jeng-Jong Hwang3,*, Ju-Yu Wang4, Yun-Lin Hsiao2, Chia-Wen Tsai1, Juhn-Cherng Liu2, Tsung-Ho Ying5 and Da-Tian Bau1,2
1Terry Fox Cancer Research Laboratory, China Medical University Hospital, Taichung, Taiwan;
2Graduate Institute of Clinical Medical Science, China Medical University, Taichung, Taiwan;
3Department of Biomedical Imaging and Radiological Sciences, National Yang-Ming University, Taipei, Taiwan;
4Basic Medical Science, Department of Nursing, Hung-Kuang University, Taichung, Taiwan;
5Department of Obstetrics and Gynecology, Chung Shan Medical University Hospital, Taichung, Taiwan.
Running head: APEX1 IN ENDOMETRIOSIS
* These authors contributed equally to this work
*Correspondence to: Da-Tian Bau, Terry Fox Cancer Research Lab, China Medical
University Hospital, 2 Yuh-Der Road, Taichung, 404 Taiwan, Tel: +886 422052121 Ext 1523, Fax: +886 422052121 Ext 1543, E-mail: [email protected]/
Abstract
Background: The altered cellular repair capacity plays a critical role in genomic instability and carcinogenesis. We aim at evaluation the contribution of the polymorphic variant in apurinic/apyriminidic endonuclease (APEX1) gene to its mRNA and protein levels and the risk of endometriosis. Materials and methods: In the current case-control study, 153 endometriosis patients and 636 non-endometriosis controls were recruited. APEX1 Asp148Glu (rs1130409) genotyping was conducted by polymerase chain reaction-restriction fragment length polymorphism.
At the same time, twenty eight endometriosis tissue samples with different genotypes were examined of the expression levels of APEX1 mRNA and protein by quantitative reverse transcription polymerase chain reaction and Western blotting, respectively. Results: Compared with wild- type TT genotype, TG and GG genotypes of APEX1 Asp148Glu had a risk of endometriosis of 0.93- and 0.87-fold. The results from in vivo transcriptional (RNA) and translational (protein) level analysis revealed that the APEX1 mRNA and protein were of similar levels among the endometriosis tissues of people carrying TT, TG, or GG genotypes. There was no joint effect of APEX1 Asp148Glu genotype with menarche,
pregnancy, smoking or alcohol drinking lifestyles on endometriosis risk.
The evidence suggested that the APEX1 Asp148Glu genotype correlates well with its mRNA and protein expression among endometriosis patients, and may not serve as a sensitive marker for prediction of endometriosis risk in Taiwan.
Key words: APEX1, DNA repair, endometriosis, genotype, polymorphism.
Introduction
All over the world, about 10% of females at reproductive ages suffer from endometriosis, which is an hormone-dependent inflammatory benign gynaecological disease defined as that functional endometrial tissue is unusually present outside the uterus [Cramer, D.W., Wilson, E., Stillman, R.J., Berger, M.J., Belisle, S., Schiff, I., Albrecht, B., Gibson, M., Stadel, B.V., Schoenbaum, S.C., 1986. The relation of endometriosis to menstrual characteristics, smoking, and exercise. JAMA 255, 1904–
1908; Giudice, L.C., Kao, L.C., 2004. Endometriosis. Lancet 364, 1789–
1799]. In clinical, the main manifestations are dysmenorrhea, chronic pelvic pain, intercourse and infertility [Bulun, S.E., 2009. Endometriosis.
N. Engl. J. Med. 360, 268–279]. Although the mechanisms underlying endometriosis is still unrevealed, mounting evidence showed that endometriosis is a multi-factor, multi-step disease affected by inflammation, hormonal regulation, genetic and environmental interactions [Falconer, H., D‘Hooghe, T., Fried, G., 2007. Endometriosis and genetic polymorphisms. Obstet. Gynecol. Surv. 62, 616–628; Olive, D.L., Schwartz, L.B., 1993. Endometriosis. N. Engl. J. Med. 328, 1759–
1769; Rizner, T.L., 2009. Estrogen metabolism and action in
endometriosis. Mol. Cell. Endocrinol. 307, 8–18].
Among the cancer hallmarks, the genomic instability may also be closely related to the etiology of endometriosis and several polymorphic biomarkers on DNA repair genes was found [Monteiro MS. Association among XRCC1, XRCC3, and BLHX gene polymorphisms and chromosome instability in lymphocytes from patients with endometriosis and ovarian cancer. Genet Mol Res 13: 636-48, 2014; Gallegos-Arreola, MP. The TP53 16-bp duplication polymorphism is enriched in endometriosis patients. Gynecol Obstet Invest 73: 118-23, 2012; Bau, DT. Polymorphism of XRCC1 codon arg 399 Gln is associated with higher susceptibility to endometriosis. Chin J Physiol 50: 326-9, 2007;
Hsieh YY. XRCC4 codon 247*A and XRCC4 promoter -1394*T related genotypes but not XRCC4 intron 3 gene polymorphism are associated with higher susceptibility for endometriosis. Mol Reprod Dev 75: 946-51, 2008]. The DNA repair capacity of a cell is vital to the integrity of its genome and thus to its regular functions and that of the organism
[Sugimura, T., Kumimoto, H., Tohnai, I., Fukui, T., Matsuo, K., Tsurusako, S.,
Mitsudo, K., Ueda, M., Tajima, K. and Ishizaki, K. Gene-environment interaction
involved in oral carcinogenesis: molecular epidemiological study for metabolic and
DNA repair gene polymorphisms. J. Oral Pathol. Med. 35:11-18, 2006]. Therefore, any subtle mutations or polymorphisms in the DNA repair genes which contribute to the loss of its function are thought to play a critical role in carcinogenesis [Miller, K.L., Karagas, M.R., Kraft, P., Hunter, D.J., Catalano, P.J., Byler, S.H. and Nelson, H.H. XPA, haplotypes, and risk of basal and squamous cell carcinoma. Carcinogenesis 27:1670-1675, 2006;
Bau DT. Cancer res 2004; Bau DT. Carcinogenesis 2007]. Since endometriosis also has the deregulated proliferation property similar to cancers, it is reasonable that genetic variants of the DNA repair genes might also involve in the initiation and development of endometriosis.
Among the known DNA repair pathways, base excision repair (BER) pathway is in charge of removing the DNA adducts induced by mainly oxidation and alkylation and thus protecting cells against the cytotoxic effects at the first line [Fleck, O. and Nielsen, O. DNA repair. J. Cell Sci.
117:515-517, 2004; Hoeijmakers, J.H. Genome maintenance mechanisms for preventing cancer. Nature 411:366-374, 2001]. Immediately after the DNA insults, DNA glycosylases recognize and excise the altered bases of purine or pyrimidine and leave some abasic sites. Consequently, apurinic/apyrimidinic endonucleases incise the DNA 5’ to the abasic
sites. Then the continued processes undergo to short-patch (when the gap is only one nucleotide) or long-patch (when the gap is two or more nucleotides) subpathways of BER system [Krokan, H.E., Nilsen, H., Skorpen, F., Otterlei, M. and Slupphaug, G. Base excision repair of DNA in mammalian cells. FEBS Lett. 476:73-77, 2000]. From the above background we know that the virtue human apurinic/apyrimidinic endonucleases, APEX1 (also known as APE, APE1, APEX, HAP1, and Ref-1) is uniquely responsible to the repair of apurinic/apyrimidinic sites at altered DNA produced either spontaneously hydrolyzing the 5’- phosphodiester bone of the apurinic/apyrimidinic site or after enzymatic removal of damaged bases. In addition to its apurinic/apyrimidinic endonuclease activity, APEX1 can also act as a 3’-phosphodiesterase, initiating repair of DNA strands breaks with 3’-blocking damage, which are produced either directly by reactive oxygen species or indirectly through the enzymatic removal of damaged bases [Fortini, P., Pascucci, B., Parlanti, E., D'Errico, M., Simonelli, V. and Dogliotti, E. The base excision repair: mechanisms and its relevance for cancer susceptibility.
Biochimie 85:1053-1071, 2003; Izumi, T., Hazra, T.K., Boldogh, I., Tomkinson, A.E., Park, M.S., Ikeda, S. and Mitra, S. Requirement for
human AP endonuclease 1 for repair of 3'-blocking damage at DNA single-strand breaks induced by reactive oxygen species. Carcinogenesis 21:1329-1334, 2000]. Furthermore, APEX1 also conducts a reduction- oxidation activators for several transcription factors reported to be important in carcinogenesis, such as activator protein (Fos/Jun), hypoxia- inducible factor 1, cAMP-responsive element binding protein, and p53 [Evans, A.R., Limp-Foster, M. and Kelley, M.R. Going APE over ref-1.
Mutat. Res. 461:83-108, 2000; Tell, G., Damante, G., Caldwell, D. and Kelley, M.R. The intracellular localization of APE1/Ref-1: more than a passive phenomenon? Antioxid. Redox Signal. 7:367-384, 2005;
Jayaraman, L., Murthy, K.G., Zhu, C., Curran, T., Xanthoudakis, S. and Prives, C. Identification of redox/repair protein Ref-1 as a potent activator of p53. Genes Dev. 11:558-570, 1997]. In knockout mice model, APEX1 deficiency-induced embryonic lethality strengthened the importance of APEX1 in maintaining the genomic integrity and viability of cells and whole organisms [Burdak-Rothkamm, S., Rube, C.E., Nguyen, T.P., Ludwig, D., Feldmann, K., Wiegel, T. and Rube, C. Radiosensitivity of tumor cell lines after pretreatment with the EGFR tyrosine kinase inhibitor ZD1839 (Iressa). Strahlenther. Onkol. 181:197-204, 2005].
It is been shown that genetic polymorphisms in DNA repair genes have conferred predisposition to many types of cancer, and as mentioned above there were a few studies investigated the association between the genotypes of some DNA repair genes and the endometriosis risk, such as
XRCC1, XRCC3, BLHX, TP53, and XRCC4 [Monteiro MS. Association among XRCC1, XRCC3, and BLHX gene polymorphisms and chromosome instability in lymphocytes from patients with endometriosis and ovarian cancer. Genet Mol Res 13: 636-48, 2014; Gallegos-Arreola, MP. The TP53 16-bp duplication polymorphism is enriched in endometriosis patients. Gynecol Obstet Invest 73: 118-23, 2012; Bau, DT.
Polymorphism of XRCC1 codon arg 399 Gln is associated with higher susceptibility to endometriosis. Chin J Physiol 50: 326-9, 2007; Hsieh YY. XRCC4 codon 247*A and XRCC4 promoter -1394*T related genotypes but not XRCC4 intron 3 gene polymorphism are associated with higher susceptibility for endometriosis. Mol Reprod Dev 75: 946-51, 2008]. However, no study had yet confirmed the association between the polymorphisms of APEX1, which is a central gene in human BER, and the risk of endometriosis. In recent years, several epidemiological studies have examined the contribution of APEX1 genotypes to several types of
cancers, including bladder [Gangwar, R., Ahirwar, D., Mandhani, A. and Mittal, R.D. Influence of XPD and APE1 DNA repair gene polymorphism on bladder cancer susceptibility in north India. Urology 73:675-680, 2009], lung [Li, Z., Guan, W., Li, M.X., Zhong, Z.Y., Qian, C.Y., Yang, X.Q., Liao, L., Li, Z.P. and Wang, D. Genetic polymorphism of DNA base-excision repair genes (APE1, OGG1 and XRCC1) and their correlation with risk of lung cancer in a Chinese population. Arch. Med.
Res. 42:226-234, 2011; Lo, Y.L., Jou, Y.S., Hsiao, C.F., Chang, G.C., Tsai, Y.H., Su, W.C., Chen, K.Y., Chen, Y.M., Huang, M.S., Hu, C.Y., Chen, C.J. and Hsiung, C.A. A polymorphism in the APE1 gene promoter is associated with lung cancer risk. Cancer Epidemiol. Biomarkers Prev.
18:223-229, 2009; Chen WC. The contribution of DNA apurinic/apyrimidinic endonuclease genotype and smoking habit to Taiwan lung cancer risk. Anticancer Res 33: 2775-8, 2013], gastric [Gu, D., Wang, M., Wang, S., Zhang, Z. and Chen, J. The DNA repair gene APE1 T1349G polymorphism and risk of gastric cancer in a Chinese population. PLoS One 6:e28971, 2011], prostate [Kuasne H. Base excision repair genes XRCC1 and APEX1 and the risk for prostate cancer. Mol Biol Rep 38: 1585-91, 2011], colorectal [Kasahara M.
Association of MUTYH Gln324His and APEX1 Asp148Glu with colorectal cancer and smoking in a Japanese population. J Exp Clin Cancer Res 27: 49, 2008], head and neck cancer [Mahjabeen, I. Genetic and expressional variations of APEX1 are associated with increased risk of head and neck cancer. Mutagenesis 28: 213-8, 2013]. We assumed that the genotypes together with phenotypes of APEX1 may also contribute to the personal endometriosis risk determination. Thus, the present case- control study is aiming at revealing the relationship between APEX1 genotype/phenotype and risk for endometriosis in a moderate Taiwan female population. We have the following strategies: 1) as for APEX1 genotype, we chose the most common polymorphism in literature, APEX1 Asp148Glu (rs1130409), to investigate the association between APEX1 genetic polymorphism with endometriosis risk. 2) as for APEX1 phenotype, the mRNA and protein expression levels of various APEX1 genotypes in vivo were examined by reversed transcript PCR and Western blotting assays, respectively. To the best of our knowledge, this is the first study to evaluate the contribution of APEX1 Asp148Glu genotype and its phenotype to endometriosis susceptibility.
Materials and Methods
Study Population
One hundred and fifty three patients diagnosed with endometriosis were recruited at the outpatient clinics of general surgery during 2000- 2010 at Chung Shan Medical University Hospital in Taiwan. The endometriosis patients were diagnosed by laparoscopy and classified according to the American Society for Reproductive Medicine and all the patients were confirmed histologically of the disease. Among the endometriosis patients, the disease was found to be minimal/mild (stages I and II) in 62 cases (40.5%) and moderate/severe (stages III and IV) in 91 cases (59.5%). Patients with pathological confirmation or clinical suspicion of leiomyoma, adenomyosis, or invasive carcinoma of the uterine cervix or ovary were excluded from this study. No patient had received hormone therapy during the preceding 12 months. The mean age of the endometriosis patients was 40.3 ± 4.9 years, and 55 of them (35.9%) did not have a child or full pregnancy. The basal FSH level was 7.2 ± 1.4 IU/l. The non-endometriosis statuses were confirmed after detail ultrasonography. All operations were performed by experienced surgeon Dr. Yin and his colleagues. According to the revised American Fertility Society (AFS) classification, 32 (20.9%) had minimal or mild
endometriosis (stage I–II) and 121 (79.1%) had moderate or severe endometriosis (stage III–IV). All women accepted to provide their peripheral blood sampling for genotype analyses with their informed consents. The experiment was approved by the Ethical Committee and Institutional Review Board of the Chung Shan Medical University Hospital. Six hundred and thirty six non-endometriosis healthy volunteers as the controls were selected by matching for their age and habits after initial random sampling from the Health Examination Cohort of China Medical University Hospital. The exclusion criteria of the control group included previous malignancy, metastasized cancer from other or unknown origin, or any familial disease. Both groups completed a well- informed questionnaire which included the individual smoking and drinking habits. Smokers were defined as daily or almost daily smokers, who had smoked at least five packs of cigarettes in their lifetime.
Smokers were asked for the age of initiation, whether they were currently smoking or had already quit, and if so, when they had quit, and on average, how many cigarettes they smoked or had smoked daily. The non-drinkers included those with social drinking behavior of less than 200 ml per week and less than twice per month.
Genotyping Protocol
The total genomic DNA of each participant was extracted from the leucocytes of peripheral blood using a QIAamp Blood Mini Kit (Qiagen, Taipei, Taiwan) as previously published [Chen WC. AR 33: 2775-8, 2013]. The primers used for APEX1 Asp148Glu (rs1130409) were: forward 5’-
CCAGCTGAACTTCAGGAGCT-3’, and reverse 5’-
CTCGGCCTGCATTAGGTACA-3’. The following cycling conditions were performed: started with one cycle at 94oC for 5 min; 35 cycles of 94oC for 30 s, 55oC for 30 s, and 72oC for 30 s; and a final extension at 72oC for 10 min. The PCR resultant 350 bp PCR product was mixed with 2 U MnlI and the enzyme digestion was carried out for 2 h at 37 oC. The G form PCR products could be further digested while the T form could not. Two fragments 252 and 98 bp were present if the product was digestible C form. Then, 10 l of product was loaded into a 3% agarose gel electrophoresis and recognized under UV light with ethidium bromide staining.
APEX1 mRNA Expression Pattern
To evaluate the correlation between the APEX1 mRNA expression and APEX1 genotype, 28 surgically removed tissue samples adjacent to endometriosis with different genotypes were subjected to extraction of the total RNA using Trizol Reagent (Invitrogen, Carlsbad, CA, USA).
The total RNA was measured by real-time quantitative RT-PCR using FTC-3000 real-time quantitative PCR instrument series (Funglyn Biotech Inc., Canada). GAPDH was used as an internal quantitative control. The primers used for amplification of APEX1 mRNA were 5’- GCCCACTCAAAGTTTCTTAC-3’ as forward one and 5’- TGTGCCACATTGAGGTCTCC-3’ as reverse one, and the primers for GAPDH were 5’-GAAATCCCATCACCATCTTCCAGG-3’ as forward one and 5’-GAGCCCCAGCCTTCTCCATG-3’ as reverse one. Fold changes were normalized by the levels of GAPDH expression, and each assay was performed in at least triplicate.
Western Blotting Analysis
The endometriosis or control specimens were homogenized in RIPA lysis buffer (Upstate Inc., Lake Placid, NY, USA), the homogenates were
then centrifuged at 10,000 g for 30 min at 4°C, and the supernatants were used for Western blotting. Samples were denatured by heating at 95°C for 10 min, then separated on a 10% SDS-PAGE gel, and transferred to a nitrocellulose membrane (Bio-Rad). The membrane was blocked with 5% non-fat milk and incubated overnight at 4°C with anti-APEX1 antibody (1:1000; Santa Cruz Biotechnology), then with the corresponding horseradish peroxidase-conjugated goat anti-mouse IgG secondary antibody (Chemicon, Temecula, CA) for 1 h at room temperature. After reaction with ECL solution (Amersham, Arlington Heights, IL, USA), a bound antibody was visualized using a chemiluminescence imaging system (Syngene, Cambridge, UK). Finally, the blots were incubated at 56°C for 18 min in stripping buffer (0.0626 M Tris-HCl, pH 6.7, 2% SDS, 0.1 M mercaptoethanol) and re-probed with a monoclonal mouse anti-β-actin antibody (Sigma) as the loading control.
The optical density of each specific band was measured using a computer-assisted imaging analysis system (Gene Tools Match software;
Syngene, Cambridge, UK).
Statistical Analysis
To ensure that the controls used were representative of the general population and to exclude the possibility of genotyping error, the deviation of the genotype frequencies of APEX1 single nucleotide polymorphisms in the control subjects from those expected under the Hardy-Weinberg equilibrium was assessed using the goodness-of-fit test.
Chi-square test was used to compare the distribution of the APEX1 genotypes between cases and controls. The associations between the APEX1 polymorphisms and endometriosis risk were estimated by computing odds ratios (ORs) and their 95% confidence intervals (CIs) from logistic regression analysis with the adjustment for possible confounders. The expression levels of mRNA and protein were examined by unpaired Student’s t-test. Those p < 0.05 were considered statistically significant, and all statistical tests were two-sided.
Results
Basic Comparisons of the Cases and Controls
The characteristics of the control and endometriosis subjects are presented in Table I. No differences between the case and control group
was found on age, menarche age, smoking or alcohol drinking status (p>0.05). Noticeably, there were less percentages of females having full pregnancy among cases than in the controls (64.1% versus 75.9%), and the significance (p = 0.0041) suggested that full pregnancy may be one of the protective factor for endometriosis.
Association of APEX1 Genotypes with Endometriosis Risk
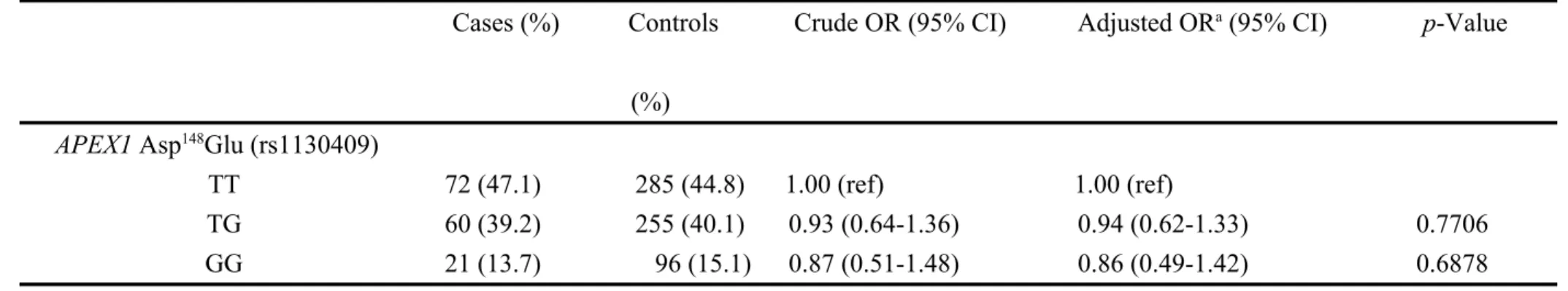
The genotypic distributions of the APEX1 Asp148Glu polymorphism in endometriosis cases and controls are presented in Table II. The crude ORs for the people carrying TG and GG genotypes were 0.93 (95% CI = 0.64-1.36) and 0.87 (95% CI = 0.51-1.48) respectively, compared to those carrying TT wild-type genotype. After the adjustment of the confounding factors (age, full pregnancy, smoking and alcohol drinking status) the ORs for the people carrying TG and GG genotypes became 0.94 (95% CI = 0.62-1.33) and 0.86 (95% CI = 0.49-1.42) respectively, compared to those carrying TT wild-type genotype. The p for trend was not significant (p = 0.8532). In the dominant (TG plus GG versus TT) or recessive (GG versus TT plus TG) analyzing models, the association between APEX1 Asp148Glu polymorphism with endometriosis risk was
not statistically significant either. (Table II). To sum up, these results indicated that individuals carrying variant G allele at APEX1 Asp148Glu may not have a higher risk of endometriosis.
Correlation of the APEX1 Asp148Glu Genotype with the Expression Levels of the APEX1 mRNA and Protein
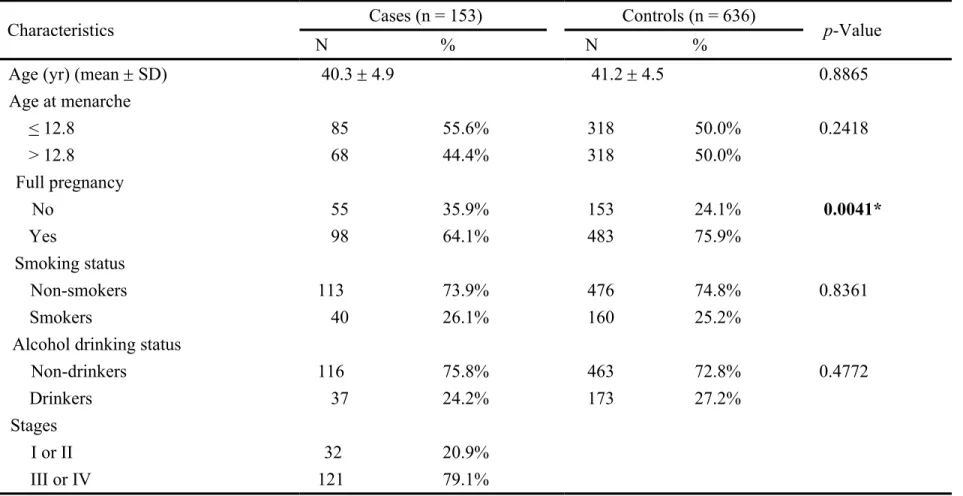
We have collected 28 surgically removed endometriosis tissue samples and their frequencies of the TT, TG, and GG genotypes of the APEX1 Asp148Glu were 13, 11, and 4, respectively. The effects of these three genotypes on the transcriptional expression of mRNA levels and translational expression of protein levels were evaluated by quantitative RT-PCR and Western blotting, respectively (Fig. 1). There was no obvious difference found among the mRNA or protein levels of various genotypes (Fig. 1).
Interaction of APEX1 Genotypes with Pregnancy, Menarche, Smoking and Alcohol Drinking Status
It is reasonable that the genetic variation of a low-penetrate gene, such as APEX1 in this paper, may not significantly contribute to the susceptibility
of endometriosis as much as the environmental factors, such as the individual smoking lifestyle to smoking-related cancers. The environmental factors for endometriosis among Taiwan females are not clear. Therefore, we are interested in analyzing the interaction of genotype of APEX1 Asp148Glu with some factors including pregnancy, menarche, smoking and alcohol drinking status. As show in Table III, the frequencies of various APEX1 Asp148Glu genotypes were not significantly different between endometriosis and non-endometriosis control groups among those who have early menarche (< 12.8 years old) or late menarche (> 12.8 years old) (p=0.7673 and 0.3030, respectively) (Table III). As show in Table IV, the frequencies of various APEX1 Asp148Glu genotypes were not significantly different between endometriosis and non-endometriosis control groups among non-smokers or smokers (p=0.4694 and 0.4094, respectively) (Table IV). The frequencies of various APEX1 Asp148Glu genotypes were not significantly different between endometriosis and non-endometriosis control groups among those who with and without full pregnancy, and alcohol drinkers or non- drinkers (data not shown).
Discussion
Endometriosis, which has the same features of malignant tumors, is one of the serious gynecological diseases among females. The DNA repair systems play as gatekeepers for genome integrity and genomic instability of cancer cells is a common event in cancer initiation and progression. In current study, the association of APEX1 genotype and endometriosis risk was firstly investigated in Taiwan, where the prevalence of endometriosis was 2.7% during 1998 to 2008 [Fang RC.
Evid Based Complement Alternat Med 591391, 2012]. The APEX1 Asp148Glu is the most common APEX1 polymorphism studied that result in amino acid alteration [Hung, R.J., Hall, J., Brennan, P. and Boffetta, P.
Genetic polymorphisms in the base excision repair pathway and cancer risk: a HuGE review. Am. J. Epidemiol. 162:925-942, 2005]. In previous literature, the contributions of APEX1 Asp148Glu genotype to the risk of other types of cancer are very controversial. For instance, the association of APEX1 Asp148Glu genotype with lung cancer risk is most well studied, but the findings are not conclusive yet [Chen W. Tumour Biol 35: 3597- 603, 2014; Pan H. PLOS ONE 8: e69018; 2013; Chen WC. AR 33: 2775-
8, 2013; Ito, H., Matsuo, K., Hamajima, N., Mitsudomi, T., Sugiura, T., Saito, T., Yasue, T., Lee, K.M., Kang, D., Yoo, K.Y., Sato, S., Ueda, R.
and Tajima, K. Gene-environment interactions between the smoking habit and polymorphisms in the DNA repair genes, APE1 Asp148Glu and XRCC1 Arg399Gln, in Japanese lung cancer risk. Carcinogenesis 25:1395-1401, 2004; Kiyohara, C., Takayama, K. and Nakanishi, Y.
Association of genetic polymorphisms in the base excision repair pathway with lung cancer risk: a meta-analysis. Lung Cancer 54:267-283, 2006]. Up to now, there is no report investigating the association of APEX1 genotype with endometriosis risk.
There is less percentage of women with full pregnancy experience in the endometriosis group (64.1%) than the control groups (75.9%) (Table I). The results correlate with the concept that endometriosis is suspected to contribute to infertility of women [Holoch KJ, Lessey BA:
Endometriosis and infertility. Clin Obstet Gynecol 2010, 53:429-438].
From the results of APEX1 Asp148Glu genotyping, we found that individuals carrying variant TG or GG genotypes were not of higher risk of endometriosis compared with those carrying wild-type TT genotype (Table II). We have also investigated the correlation of APEX1 Asp148Glu
genotype from the angles of transcriptional (mRNA) and translational (protein) expression levels of APEX1 with tissues collected from endometriosis patients. The results showed that the endometrial tissues from people of TT, TG or GG APEX1 Asp148Glu genotypes were of similar level of APEX1 mRNA and protein levels (Figure 1). It is not surprising that the amino-acid substitution variants on APEX1 Asp148Glu lead to similar consequent levels of mRNA and protein of itself. Further efforts could be made on 1) the genotyping of polymorphisms in the promoter region, such as the promoter -141T/G (rs1760944) [Li Z.
Genetic polymorphism of DNA base-excision repair genes (APE1, OGG1 and XRCC1) and their correlation with risk of lung cancer in a Chinese population. Arch. Med. Res. 42:226-234, 2011]; 2) the functional analysis of overall base excision repair capacity in the primarily cultured cells from tissues with different genotypes. Compared with the wild-type mice, the APEX1 heterozygous mice were reported to have accelerated DNA damage adducts accumulated in mitochondrial DNA and spontaneous mutagenesis [Vogel KS. Mol Repord Dev 78: 906-19]; 3) the enlarging of sample size is always helpful for enhancing the analyzing power of overall or stratified comparisons.
There is no denying that endometriosis is multi-step and multi- factorial disease and factors such as hormone exposure, inflammation, familial predisposition, growth factors, diet, altered immune system, environmental factors and oxidative stress status may contribute to the initiation and progression of endometriosis and its possible transformation into endometrial cancer. However, largely of the mechanisms are unknown. As for endometriosis and endometrial cancer, early menarche, low parity, late menopause, infertility, organochlorinated persistent pollutants have been implicated to be risky factors of them [Verit FF. Asian Pac J Cancer Prev 14: 5589-97, 2013; Itoh H. Sci Total Environ 408: 37-42, 2009; Porpora MG. Environ Health Perspect 117:
1070-5, 2009]. However, the collection of precise records and investigating direct and indirect relationship of them are both time- and effort-consuming and the associations of them with endometriosis are not well established. Thus, in Tables III and IV, we attempted to evaluate the joint effect of the polymorphism of APEX1 genes stratified by menarche age and smoking status. We found that APEX1 Asp148Glu genotypes had no obvious preferential distribution among each subgroup (Tables III and IV), suggesting that APEX1 Asp148Glu genotype had selective adding the
risk of endometriosis to either those females with early or late menarche, or to smokers or non-smokers. As for full pregnancy experience and alcohol drinking, the effect is not obvious either (data not shown). As mentioned above, the limited sample size may restrict the reliability and feasibility of stratification and interaction analyses.
In this study no significant association of APEX1 genotype was found. However, we could not ignore the contribution of APEX1 in the etiopathology of endometriosis. It is possibly that some other polymorphic sites, such as the promoter -141T/G abovementioned may related to an alteration in the expression level of mRNA, protein, and consequently its function, which lead to an enhanced risk of endometriosis. To make the predictive, preventive, personalized and participatory medicine and therapy for endometriosis feasible and to lower the speed and the prevalence of several types of cancer such as endometriosis in developed countries, the environmental factors which may cause lots of oxidative damage related to BER, such as the shoddy oil used in cosmic and in daily diet, smoking and alcohol drinking, should be restrictly or adaptively prohibited among all of the subjects in our society.
In conclusion, the current study has provided evidence from DNA, RNA and protein levels showing that APEX1 Asp148Glu was not associated with endometriosis in Taiwan. Further understanding of the role of DNA repair genes such as APEX1 in this paper, with genotypic together with phenotypic evidence is in urgent need to reveal the etiology of endometriosis.
Acknowledgements
This study was supported by research grants from Terry Fox Cancer Research Foundation and Taiwan Ministry of Health and Welfare Clinical Trial and Research Center of Excellence (MOHW103-TDU-B- 212-113002) at China Medical University Hospital. The assistance from Tsai-Ping Ho in clinical sample and data collection, and that from Hong- Xue Ji, Chieh-Lun Hsiao, Chia-En Miao, Lin-Lin Hou in genotyping, RT- PCR and Western blot were highly appreciated by the authors. The authors declared no conflict of interest.
Figure legend
Fig. 1 Analysis of APEX1 mRNA and protein expression levels. A,
Quantitative RT-PCR for APEX1 from the endometriosis tissue samples of TT, TG and GG genotypes was performed and GAPDH was used as an internal quantitative control. Fold changes were normalized by the levels of GAPDH expression, and each assay was done in at least triplicate. B, The Western blotting analysis was performed from the endometriosis tissue samples of TT, TG and GG genotypes and quantitated for comparison. -tubulin was used as an internal standard.
References 1.
2.
Table I. Distributions of selected characteristics among endometriosis cases and control subjects
Characteristics Cases (n = 153) Controls (n = 636)
p-Value
N % N %
Age (yr) (mean SD) 40.3 4.9 41.2 4.5 0.8865
Age at menarche
< 12.8 85 55.6% 318 50.0% 0.2418
> 12.8 68 44.4% 318 50.0%
Full pregnancy
No 55 35.9% 153 24.1% 0.0041*
Yes 98 64.1% 483 75.9%
Smoking status
Non-smokers 113 73.9% 476 74.8% 0.8361
Smokers 40 26.1% 160 25.2%
Alcohol drinking status
Non-drinkers 116 75.8% 463 72.8% 0.4772
Drinkers 37 24.2% 173 27.2%
Stages
I or II 32 20.9%
III or IV 121 79.1%
* Statistically significant
Table II. Distributions of genotypic frequencies and their association with risk of endometriosis
Cases (%) Controls
(%)
Crude OR (95% CI) Adjusted ORa (95% CI) p-Value
APEX1 Asp148Glu (rs1130409)
TT 72 (47.1) 285 (44.8) 1.00 (ref) 1.00 (ref)
TG 60 (39.2) 255 (40.1) 0.93 (0.64-1.36) 0.94 (0.62-1.33) 0.7706
GG 21 (13.7) 96 (15.1) 0.87 (0.51-1.48) 0.86 (0.49-1.42) 0.6878
a Adjusted by age, pregnancy, smoking and alcohol drinking status
Table III. Association between APEX1 genotype and risk of endometriosis stratified by menarche age
APEX1 genotype Early menarche Late menarche
Cases/controls aOR (95% CI) a Cases/controls aOR (95% CI) a
Asp148Glu (rs1130409)
TT 38/147 1.00 (ref) 34/138 1.00 (ref)
TG 32/125 0.98 (0.61-1.65) 28/130 0.88 (0.51-1.58)
GG 15/46 1.25 (0.62-2.48) 6/50 0.49 (0.22-1.19)
p for trend 0.7673 0.3030
Abbreviations: aOR, adjusted odds ratio.
a Data were adjusted for pregnancy, smoking and alcohol drinking status.
Table IV. Association between APEX1 genotype and risk of endometriosis stratified by personal smoking habits
APEX1 genotype Non-smokers Smokers
Cases/controls aOR (95% CI) a Cases/controls aOR (95% CI) a
Asp148Glu (rs1130409)
TT 53/200 1.00 (ref) 19/85 1.00 (ref)
TG 48/197 0.94 (0.61-1.40) 12/58 0.94 (0.43-2.01)
GG 14/79 0.69 (0.38-1.15) 7/17 1.80 (0.68-4.89)
p for trend 0.4694 0.4094
Abbreviations: aOR, adjusted odds ratio.
a Data were adjusted for age, pregnancy, and alcohol drinking status.
(A)
Relative expression level (fold of TT genotype) 0.6 0.8 1.0 1.2 1.4
Genotype
TT TG GG
A
(B)
Relative expression level (fold of TT genotype) 0.6 0.8 1.0 1.2 1.4
Genotype
TT TG GG
-tubulin APEX1
TT TG GG