行政院國家科學委員會專題研究計畫成果報告
近緣種之多食性潛蠅形態進化與系統學研究
Morphological evolution and systematics studies on closely related
polyphagous agromyzid species
計畫編號:
NSC 90-2313-B-002-334執行期限:90 年 8 月 1 日至 91 年 7 月 31 日
主持人:蕭旭峰 國立台灣大學昆蟲學系
一、 中文摘要 傳統分類部份持續整理台灣地區名 錄,蒐集台灣產潛蠅科種類並重新描述及 提供較為詳盡之特徵資料;並研究以相對 扭曲(Relative warp)定序法或聚類分析以潛 蠅翅脈為主作系統學或進化學等相關推 論。本年度主要以分析斑潛蠅害蟲形態鑑 別及潛蠅科系統學關係為主要工作內容; 斑潛蠅(Liriomyza)是臺灣地區近年來新崛 起的蔬菜及花卉重要害蟲,部分種類亦被 認為是經由進口所導致的新侵入害蟲;然 而不論在檢疫與防疫的研究與應用上,種 類的判定與區別一直困擾著研究與檢防疫 工作者。斑潛蠅由於屬內形態同質性太高 且寄主植物範圍廣泛重疊,再加上競爭取 代現象頻繁,因此在國內外種類遭誤判的 情況相當普遍;如此勢必嚴重影響檢防疫 工作之精確性與研究結果的可靠性。本研 究針對臺灣產 6 種主要斑潛蠅害蟲(同時 也是重要的檢防疫對象害蟲),包括:白菜 斑潛蠅(L. brassicae)、蕃茄斑潛蠅(L. bryoniae)、 蔥斑潛蠅(L. chinensis)、南 美斑潛蠅(L. huidobrensis)、蔬菜斑潛蠅(L. sativae)及非洲菊斑潛蠅(L. trifolii)等作 形態區辨之報導;目的在尋求簡易快速的 形態區辨特徵,提供非分類學之相關工作 人員應用參考。除討論雄蟲外生殖器特徵 外,結果亦顯示在以有限的害蟲種類為對 象下,腹部背板斑紋、翅脈形態測量及電 顯局部放大之微毛列等特徵,可以在判定 特定種類上有相當程度的應用價值。 關鍵詞:形態學、形態測量學,系統分類 學,潛蠅科,雙翅目。 AbstractLiriomyza leafminers is a pest group that
causes not only the considerable economic losses but also the serious quarantine problems. Due to the increased volume of international trade of agricultural products and the morphological similarities of the closely related species in this pest group, efficient and accurate pest species recognition on imported or exported crops becomes more and more important; and this is probably the only way to prevent them from outspreading by human-mediated dispersal. Therefore, morphological studies were performed to assist species identification and discrimination of six
Liriomyza pest species of quarantine
importance, which include L. brassicae, L.
bryoniae, L. chinensis, L. huidobrensis, L. sativae, and L. trifolii. Some traditional
morphological characters, such as abdominal color patterns and male genitalia were re-evaluated for their discriminative ability. Besides, the electronmicrography and geometric morphometric methods were introduced to use in separating different species. Illustrative plates of the preceding morphology were collected into mappings for further applications in quarantine inspection; some analyses and evaluations in separating similar species were also discussed in further detail. The results show that the abdominal color patterns can only separate two species from others; nevertheless, the ultra-structure of thoracic microsetae and male genitalia are useful morphological characters to prevent
misidentifications. Thoracic microsetae show their unique arrangement and pattern on each species both in length and in density; meanwhile, the photography of male genitalia, which have larger focal depth, was also proven taxonomically valuable in practice. Although not all pairs between any two species have significant differences in wing shapes, wing morphometric results do reveal that the most variant area of wing shape is located around the cross veins; this suggests that the morphology on this area might be used easily and efficiently in dividing different species.
Keywords: Morphometrics, Systematics,
Agromyzidae, Diptera. 二、緣由與目的 潛蠅科昆蟲原本只是農田的次要害 蟲,但其害蟲地位在過去20 年間有了劇烈 的變化;從80 年代大發生到因人為攜帶所 引發的植物檢疫問題及至目前的入侵、競 爭及取代的種種現象,使得我們對這些具 經濟重要性的昆蟲,其行為、生態甚至分 類、進化的基礎生物學問題有了更大的興 趣。潛蠅科是無瓣類雙翅目中種類數較多 的一個大類群;有趣的是僅有非常少數的 種類在進化過程中衍生出多食的習性,其 中更有多數種類集中在斑潛蠅(Liriomyza) 屬中;這些種類也都是我們所熟知的害 蟲。雖然斑潛蠅的經濟重要性極高,但分 類鑑定的工作卻常常出錯;而這種鑑定錯 誤所造成防治方針偏差的狀況到目前仍是 一大困擾。1980 年代美國加州斑潛蠅大發 生時,分類上的混淆就是一個例子。不過 由實際分類的工作上來看,斑潛蠅在種級 分類上的區辨的確並不容易;由於屬的同 質性高,因此大部份的種類並不容易直接 從外部形態來區分;目前分類學者多半採 用穩定性較高的雄蟲生殖器作為分類依 據。當兩種或兩種以上的雜食性種類同時 並存於一個區域時,競爭、取代現象不斷 發生,而此時正確地判斷出種類,往往是 防治成功與否的第一個關鍵;Parrella 及 Keil (1983) 就曾指出斑潛蠅大發生及防治 的主要原因及盲點首在於學者們對種類鑑 定上的混淆。 三、研究方法與進行步驟 主要標本來源為台大昆蟲系標本收藏 (by Tung-Ching Hsu (T. C. Hsu) and Wen-Jer Wu (W. J. Wu) during early 1980’s and 1990’s),及本研究室近年的標本收藏 ( by Liang-Yih Chou (L. Y. Chou), Yi-Chung Chiu (Y. C. Chiu), Shu-Chen Chang (S. J. Chang), and the author (S. F. Shiao))。除此之外並加入日本(北海道)、 美國(UC Davis, California)及台灣(台大) 等三地之實驗室飼養品系為試驗材料。 雄蟲生殖器以15% KOH 煮沸後解剖 並 鏡 檢 (Leica MZ125, Heerbrugg, Switzerland)後;以 Euparal 封片及以位相差 顯 微 鏡 攝 影(Olympus BH-2, PM-10AD, Tokyo, Japan)。為使外生殖器攝影得到更高 之 景 深 , 以“Auto-montage version 3.04” (Syncroscopy division, Synoptics Ltd., Cambridge, U.K.)影像處理軟體,作不同焦 距平面影像之合成處理。
電子顯微鏡標本處理,採標本自然陰 乾方式後鍍膜 (by Hitachi HUS-5GB High Vacuum Evaporator, Tokyo, Japan);並在 20 KV 下作觀察攝影(by Hitachi S-520 SEM, Tokyo, Japan)。
翅脈測量採用位相差顯微鏡(Olympus BH-2, PM-10AD, Tokyo, Japan)配合數位攝 影機(DMC-1, Polaroid, MA)及電腦影像處 理。翅脈地標點法之分析流程: (1)將各種類翅脈以影像檔方式儲存於電 腦中;影像主要來源包括: (i) 採 集 或 誘 集 標 本 : 將 翅 拔 下 , 以 Euparal 平封於玻片中,利用攝影機及 影像軟體以TIFF 或 BMP 格式儲存。 (ii)文獻繪圖:將繪有翅脈之文獻以掃描 器掃描並配合軟體儲存成TIFF 或 BMP 檔。主要的圖片來源包括: Sasakawa (1961)、Spencer (1973, 1986, 1987)、 Shiao and Wu (1995a, 1996)、蕭(1991)、 蕭及吳(1996)等。
於地標點之選取定位。 (3)地標點的選取及定義 共選取 16 個(地標點作為分析資料;選 取的原則為儘可能選取所有翅脈交點以 完整脈相之資訊;翅脈交點依據地標點分 類定義應可視為第 1 型地標點(Type I landmark)(Bookstein et al., 1985),位置選 取上較無爭議。 (4)數位化地標點座標錄取,採用“TPSDIG version 1.31” (Rohlf, 2001a)軟體,以純文 字檔方式存取座標值。每一標本(影像檔) 重覆3 次錄取座標值,以平均值座標作為 最後之分析資料來源,目的為減低數位化 取點時之人為誤差。 (5)取得地標點後的資料最直接的處理就 是以重疊法(Superimposition)重合各樣本 以觀察及比較各地標點或個體之變異;目 前 的 重 疊 法 主 要 使 用 最 小 平 方 和 法 則 “generalized least-squares fit” (GLS) (Rohlf and Slice, 1990)。
(6)再以薄版曲線及其他相關分析,以更清 楚瞭解變形(Deformation)的情況;主要使 用“相對扭曲”分析(TPSRELW version 1.24) (Rohlf 2001b)及加權矩陣“W matrix” 及仿射因子 (affine components (Rohlf, 1993))之形狀因子以 UPGMA 聚類分析 ( “NTSYSpc version 2.01b” (Rohlf 1997)),表達各樣本的相似程度。
四、結果與討論
Color patterns on abdominal tergites:
Most of the Liriomyza adults can be readily recognized from other agromyzid genera by the appearances of the yellow frons and scutellum, but these color appearances are merely the early concept of this genus and not the consistent character for taxonomic identification (Spencer and Steyskal 1986). However, almost all the
Liriomyza pest species have those kinds of
special color patterns, and that also provided as an obvious external morphological character for preliminary recognition of the members in this genus in fields. In past, color patterns on mesonotum have been proposed to help recognizing some Liriomyza species
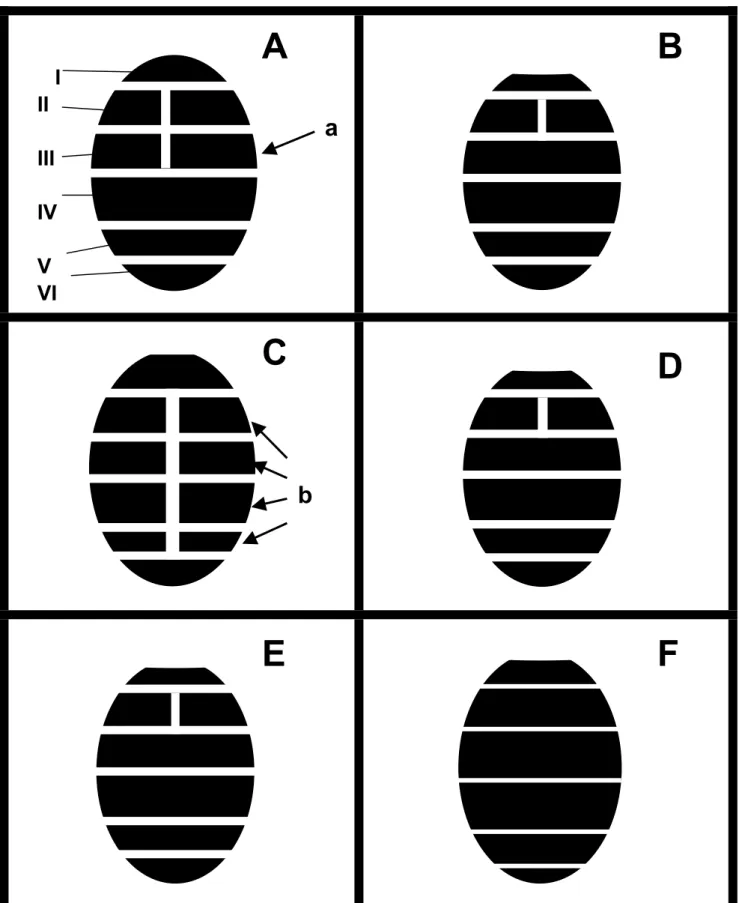
(Spencer & Steyskal 1986). Among these pest species, the abdominal tergites show these color patterns as well, and somehow more characteristic than on frons, mesonotum, and scutellum. Unfortunately, only few literatures have provided such descriptions of the coloration. Sasakawa (1960, 1961) characterized the tergal color patterns by depicting the yellow area of lateral and caudal margins on each tergal segment, but neither drawing nor detailed description has been provided. According to the results here (Fig. 1), the yellow-tinged middle furrows on the central line of abdominal tergites were revealed with much higher value for dividing some species; and three types of color patterns can be concluded in these six species. The principal differences are basically caused by lengthen or shorten of the yellow middle furrows on those six visible abdominal tergites; especially on which or how many tergal segments are divided by the yellow furrow. The most common type, which only has the second visible tergite divided by yellow middle furrow (Fig. 1-B, D, E), including three species of L. brassicae, L.
huidobrensis, and L. sativae. Another two
species of Liriomyza bryoniae and L. trifolii have otherwise longer yellow furrows with two and four divided tergites, respectively (Fig. 1-A, C). The first tergite is not included in this comparison, since it is too narrow to see whether it is divided by the yellow furrow or not, and also there are some variations occur within the same species. On the other hand, L. chinensis is a special species which shows no obvious color pattern and even has the scutellum pale gray but not yellow as the preceding species. Although abdominal color patterns can be provided as an easy and rapid tool for species diagnosis, it still cannot efficiently divide every species, especially when treated with those three species that have the same common type of the color patterns. Generally speaking, L.
trifolii has the most obvious color pattern
which has up to four divided tergites (Fig. 1b), and L. bryoniae has the yellow middle furrow stably appeared on 3rd visible abdominal tergites (Fig. 1a). Meanwhile,
the pattern mapping proposed here is only suitable for applying in male adults, female adults in my examinations presented much ambiguous among species. In addition, the application reality of using these kinds of abdominal color patterns is that they are much easily recognized by using fresh samples; for those dry specimens, the telescoped abdomens are usually shortened and shriveled, so they must be removed from specimens, stretching and smoothly flattened on a slide for better observations.
Thoracic microsetae:
Zehnder et al. (1983) firstly proposed the morphology of thoracic microsetae for diagnostic discrimination of
Liriomyza species by using scanning
electron-microscopy, they concluded that only L. trifolii could be separated from other two species (L. sativae and L.
brassicae) due to its dense covering of
microsetae. They undoubtedly provided an useful direction for exploring new morphological characters by using ultrastructure; and their methods also meet some requirements for rapid diagnosis in quarantine procedures, such as time- and effort-saving in identification for a non-expert. Unfortunately, no detailed and further study was conducted afterward. I herein reexamine those ultra-structural characters of six Liriomyza pest species, and the results show not only the density but also the length and shapes of microsetae are different among them (Fig. 2). Table 3 reveals that L. brassicae has shortest microsetae and L. chinensis has longest ones; and except for L.
bryoniae and L. sativae, thoracic
microsetae have significant differences in length among different species. Meanwhile, the density of microsetae can also be used as a reliable diagnosis character for dividing these six pest species (Table 3). Moreover, the shapes of microsetae themselves are different among species under lager magnification (Fig. 3). This means that the preceding different appearances are largely due to the different morphology of microsetae
themselves; and in some ways, implies that the differences are neither caused by artifact during the procedure of sample preparation nor by parallax from different angles in observation. Therefore, I believe these kinds of microseta mappings have great potential for applying in quarantine inspections in the future.
Wing morphometrics:
Traditional wing characters of Agromyzidae include costa ending points, length comparisons between last section of M3+4 with penultimate, and the
position of inner cross-vein (r-m) on discal cell (Spencer 1973). Sasakawa (1961) has used more other measurements in describing wing venation, such as the length proportion of 2nd to 4th costal sections, length comparison of ultimate section of M1+2
with penultimate, and length comparison of outer cross vein (m-m) with the penultimate section of M1+2. Although
some of the preceding wing characters can partially describe the venation differences, and some proportions can exclude the size effects from shape information; those traditional measurements yet cannot meet the requirements for describing precise and complete information of wing shape and venation. Modern morphometrics uses more direct and intuitive way by superimposing some encoded landmarks’ coordinates to solve this problem (Rohlf and Slice 1990). After rotation, translation, and scaling, one can aligns those landmarks as well as possible, and see the differences among specimens. The results of least-squares superimposition of sixteen landmarks for totally 297 wing specimens in six
Liriomyza species clearly show the
relative variations for each landmark among different samples (Fig. 4). Among the selected sixteen landmarks, landmark #2, #3, and #6 present larger variability which have larger ellipse areas of two standard deviations as shown in Fig. 4. Otherwise, landmarks #14 and
#15 present special directional variability that revealed by the flatter ellipses. This indicates that there have special variations happened on the outer cross vein (landmarks #14 and #15 are on the both ends of outer cross vein); and according to the longer first principal component (PCA) axes and the shorter second PCA axes of one standard deviation, I believe that the landmark variations appeared mostly along the longitudinal vein of M1+2 and M3+4.
Although the landmarks #2, #3, and #6 on wing margin have large variability, the result areas of rounder ellipses of two standard deviations suggest their differences were probably caused by some kinds of random and non-directional changes.
On the other hand, landmark-based geometric morphometrics has recently developed the theories and methods to describe, analyze, and interpret biological shapes and shape variations in more detail (Rohlf and Marcus 1993). Using a special interpolation function of “thin-plate spline” (Bookstein 1989, 1991), we can compute the coordinate transformation between corresponding landmarks on two objects, and reveal the shape differences by D’Acry Thompson’s transformation grids (Thompson 1917). In this study, the thin-plate spline relative warp analysis was computed to summarize the wing variations among species in as few dimensions as possible. For taxonomic studies, the scaling parameter “α” in relative warp analysis was set to 0 as recommended by Rohlf (1993), this becomes a principal components analysis of the covariance matrix of the partial warp scores. Results show the different shape changes were mainly happened on the central area of wing, especially on the location of cross veins (r-m and m-m cross veins) (Fig. 5). And by superimposing the minimum spanning tree onto the scatterplot of first and second relative warp axes; we can clearly see the relative recency among these six species. According to Fig. 5, the minimum
spanning tree suggests L. trifolii, L.
brassicae, and L. sativae have resembling
overall wing shapes, while L. chinensis has relatively different shape that is on the far terminal of this tree. The first and second relative warps account for 57.6 and 30.2% of the total wing variations, respectively.
One-way MANOVA were then performed by using “W matrix” (Rohlf 1993) (the matrix of partial warp scores, the projection of all the specimens onto the partial warps, which can be treated as a decomposition of shape information of non-affine changes for each species) and the affine components to determine if wing shapes varied significantly among different species. The multiple comparison results of both affine and non-affine components show except within L. trifolii, L. brassicae, and L.
sativae (L. trifolii versus L. brassicae, p=
0.43; L. trifolii versus L. sativae, p= 0.55;
L. brassicae versus L. sativae, p= 0.18,
significant unplanned comparisons based on Hotelling’s T2 values), all pairs between two species have significant shape differences.
The thin-plate-spline-based analysis is a precise tool for exploring shape variation; there are at least two advantages, its capacity to discern shape differences and the potential to graphically illustrate shape variation, as pointed out by Adams and Funk (1997). The complete wing shape information can therefore be compared and analyzed in this study, so it worth further development in application of identifying species, especially when using in detecting minor shape differences between closely related species.
Male genitalia:
Male genitalia is probably the most important taxonomic character in Agromyzidae since Nowakowski (1962), and then it was generally adopted as a major character by most of the leading specialists, such as Spencer (1990), Sasakawa (1963), Griffiths (1974), and Tschirnhaus (1971). Nevertheless, most
of these characters are presented by means of drawings, only few photos (Tschirnhaus 1981) can be found in the literature. Genitalia drawings are somewhat difficult to follow, different authors used different ways to express the genital characters, and the correct species identification is always strongly relied on the quality, precision and details of the drawings. Spencer (1992) has mentioned that the Sasakawa’s drawings of genitalia are of the highest quality, and without question the best provided by any specialist on this family. This also implies that he considered the quality of drawings do matter in identifying agromyzid species. However, those genitalia drawings are more or less personalized and somewhat difficult to compare with the real dissected objects, especially for a beginner. For example, some authors use the outline drawings to represent the overall shapes; some prefer dotted drawings to emphasize the different areas or various degree of sclerotization of those hard parts on genitalia. In this study, I carefully dissected the male genitalia and cleaned them using distilled water and alcohol then photographed them. I suppose that using photography of larger focal depth can not only overcome those defects of using traditional photos but also avoid misjudgement by reading from the descriptions or drawings. The terminology used here mainly follows that of Sasakawa (1961) and Spencer (1973).
In the broad sense, male genitalia basically include genital arch (which is composed of epandrium, two surstyli, hypandrium and two cercus), phallus, phallapodeme, and sperm pump. Epandrium is actually the ninth abdominal tergite and frequently with distinctive shape, but it’s hard to conclude those shape differences have any discriminative value on which taxonomical level. Same situation also happened when applying surstylus, hypandrium, cerci, and phallapodeme for taxonomical or identification purpose.
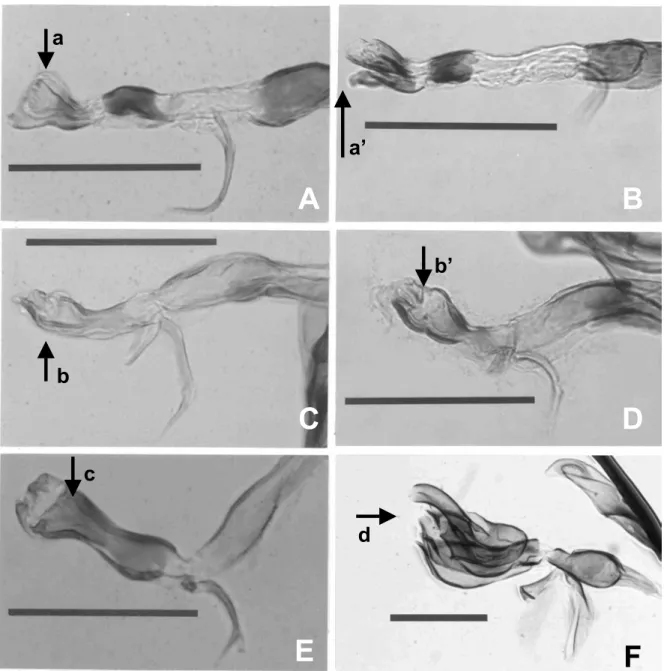
Contrarily, the phallus (or aedeagus) of Agromyizdae in most species is so highly evolved and characteristic that Spencer and Steyskal (1986) have mentioned the illustrations in side, ventral, or both views will suffice to permit immediate identification even in species level. The phallus consists of four sections: basiphallus, mesophallus, hypophallus, and distiphallus. According to the photomicrographic results in Fig.6 and Fig. 7, we can see that the distiphallus has the most conspicuous shape, and this is probably the most useful character in identifying Liriomyza species. But the problem is when we treated on those closely related species, like the species mentioned in this paper, the morphology of distiphallus is still easily confused without careful examination. There are two pairs of species in my studies which have similar shapes in phallus.
Liriomyza bryoniae and L. huidobrensis
is one of the resembling pairs (Fig. 6 A, B and Fig. 7 A, B) which have resembling distiphallus, especially from lateral view (a and a’ in Fig. 6). But carful examination will reveal that the ventral views of distiphallus are actually different, where L. bryoniae has more rounded bowl-shaped distiphallus when compare with L. huidobrensis (a and a’ in Fig. 7). In addition, the mesophallus of these two species have different sclerotization areas (b and b’ in Fig. 7), but they are obscure without properly preparing samples. Another pair of L.
trifolii and L. sativae is much difficult to
be separated, the differences of these two species mainly appear on the distiphallus (Fig. 7 c, c’ and Fig. 6 b, b’). However, the differences between these two species are quite hard to describe in writing, and only I can see is the different sclerotizing degree and minor shape changes on distiphallus. In this case, the lateral views (b and b’ in Fig. 6) seem much easier to use in separating these two species.
Morphological methods are the earliest and most intuitive way to
recognize and differentiate different living organisms, and are also the foundation on which most taxonomic systems are based. However, new techniques, such as the molecular methods, developed quickly during these years, and gradually substitute those traditional morphological methods that have long been considered subjective. Fortunately, more advanced morphological methods and instruments are now available for re-evaluating those traditional characters used in taxonomy; and even more, to explore new morphological characters. In this article, morphological characters of Liriomyza species are examined and evaluated; the purpose is under the consideration of using morphological approaches in quarantine examination for a rapid and more accurate species judgment. Using the electronmicrography and computer-aided image enhancement system, we can have image photo pictures of better quality to assist identification; using modern morphometric analysis, we can precisely locate and quantify differences of wing shapes and wing venations. In conclusion, there is no single character whatever can conveniently and successfully differentiate every
Liriomyza species, even in genitalia
characters, without careful comparisons. The reason is partially caused by the highly homogenous morphology within this group. Nevertheless, the highly similar morphology in Liriomyza pest group doesn’t match its diversified ecology; different species with different host preferences, parasite complex, and pesticide resistances do deeply affect the quarantine and control policies. And the preliminary species discrimination is only the first basic step to assist the ongoing related studies to look inside the problems. 五、計畫成果自評 潛蠅科昆蟲種類繁多,傳統分類學描述 及記錄的工作並非短時間內可以完備,目 前雖已初步建立親緣關係,然常態與持續 的進行採集、標本檢查與分析有其絕對的 必要性,我們無法排除在更詳盡種類研究 後系統關係更動的可能性。本研究在形態 與鑑別方面的研究,在應用實務上將來可 直接用在解決害蟲防治上區辨近似種的問 題。在幾何形態測量學之開發應用方面, 本研究已針對傳統形態部份將成果直接在 應用層次上獲得初步的實踐。然而,關於 形態測量學之理論及方法;自我評估對此 新領域的基礎開發及研究上仍嫌不足,對 其與系統學之配合研究方面離完整建立仍 有一段距離;且在與進化推論之關連上仍 未完備;不管在理論與實用層面將來應嘗 試持續與數學、統計或其他生物學領域等 跨學門之國內外學者作更密切的合作。 六、參考文獻
Adams, D. C., and D. J. Funk. 1997. Morphometric inferences on sibling species and sexual dimorphism in
Neochlamisus bebbianae leaf beetles:
multivariate applications of the thin-plate spline. Syst. Biol. 46: 180-194.
Bookstein, F. L. 1989. Principal warps: thin-plate splines and the decomposition of deformation. IEEE Trans. Pattern Anal. Mach. Intell. 11: 567-585.
Bookstein, F. L. 1991. Morphometric tools for landmark data: geometry and biology. Cambridge Univ. Press, Cambridge.
Cheng, C. H. 1994. Damage of the leafminer,
Liriomyza bryoniae Kalt. and its
influence on the fruit quantity and quality. Chinese J. Entomol. 14 : 433-444. (in Chinese)
Chien, C. C. 1997. Current occurrence and control of Liriomyza leafminers in Taiwan, pp. 79-94. In Proceeding of symposium on plant protection in
Taiwan and China, 29-30 May 1997, Taichung, Taiwan.
Chiu, Y. C., W. J. Wu, S. F. Shiao, and C. J. Shih. 2000a. The application of isoenzyme to develop rapid diagnostic techniques for identification of leafminer flies, Liriomyza spp. (Diptera: Agromyzidae). Plant Prot. Bull. 42: 235-247. (in Chinese)
Chiu, Y. C., W. J. Wu, S. F. Shiao, and C. J. Shih. 2000b. The application of RAPD-PCR to develop rapid diagnostic technique for identification of 6 species of Liriomyza spp. (Diptera:
Agromyzidae). Chinese J. Entomol. 20: 293-309. (in Chinese)
Collins, D. W. 1996. The separation of
Liriomyza huidobrensis (Diptera:
Agromyzidae) from related indigenous and non-indigenous species encountered in the United Kingdom using cellulose acetate electrophoresis. Ann. Appl. Biol. 128: 387-398.
(EPPO) European and Mediterranean Plant Protection Organization. 2002. Information on quarantine pests: EPPO A1 and A2 quarantine lists. http://www.eppo.org/ QUARANTINE /quarantine.html.
Griffiths, G. C. D. 1974. Studies on boreal Agromyzidae (Diptera), V-VIII. Quaest Entomol. 10: 35-69; 103-129; 217-222; 295-314.
Iwasaki, A., K. Kasugai, R. Iwaizumi, and M. Sasakawa. 2000. A newly recorded pest,
Liriomyza sativae Blanchard in Japan.
Plant Prot. 54: 142-147. (in Japanese) Kang, L. 1996. Ecology and sustainable
control of serpentine leafminers. Science Press, Peijing, China. (in Chinese)
Kasugai, K., R. Iwaizumi, M. Kumagai, and T. Tsuchiya. 2001. Agromyzidae (Diptera: Agromyzidae) detected in the import plant quarantine of Japan. Res. Bull. Plant Prot. Japan 37: 105-109.
Lee, H. S. 1986. Seasonal occurrence of the important insect pests on cabbage in southern Taiwan. J. Agric. Res. China 35: 530-542. (in Chinese)
Lee, H. S., H. C. Wen, and F. M. Lu. 1990. The occurrence of Liriomyza bryoniae (Kaltenbach) (Diptera: Agromyzidae) on Taiwan. Chinese J. Entomol. 10: 133-142. (in Chinese)
Lin, F. C., and C. L. Wang. 1992. The occurrence of parasitoids of Liriomyza
trifolii (Burgess)in Taiwan. Chinese J.
Entomol. 12:247-257. (in Chinese) Menken, S. B. J., and S. A. Ulenberg. 1986.
Allozymatic diagnosis of four economically importnat Liriomyza species (Diptera, Agromyzidae). Ann. Appl. Biol. 109: 41-47.
Minkenberg, O. P. J. M. 1988. Dispersal of
Liriomyza trifolii. OEPP/EPPO Bull. 18:
173-182.
Nowakowski, J. T. 1962. Introduction to a systematic revision of the family Agromyzidae (Diptera) with some remarks on host plant selection by these flies. Polska Akad. Nauk. Inst. Zool. Ann. Zool. 20: 67-183.
Oudman, L. 1992. Identification of economically important Liriomyza species (Diptera, Agromyzidae) and their parasitoids using enzyme electrophresis. Proc. Exper. & Appl. Entomol. 3:135-139.
Parrella, M. P. 1987. Biology of Liriomyza. Ann. Rev. Entomol. 32: 201-224. Parrella, M. P., and C. B. Keil. 1984. Insect
pest management: the lesson of
Liriomyza. Bull. Entomol. Soc. Am. 30:
22-25.
Parrella, M. P., C. B. Keil, and J. G. Morse. 1984. Insecticide resistance in
Liriomyza trifolii. Calif. Agric. 38:
22-23.
Rohlf, F. J. 1993. Relative warp analysis and an example of its application to mosquito wings, pp. 131-159. In L. F.
Marcus, E. Bello, and A. Garcia-Valdecasas (eds.), Contribution to morphometrics, Vol. 8. Monografia del Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain.
Rohlf, F. J. 1997. NTSYSpc: numerical taxonomy and multivariate analysis system, version 2.01b. Applied Biostatistics Inc., New York.
Rohlf, F. J. 2001a. TpsDig, version 1.31. Dept. of Ecology & Evolution, State University of New York at Stony Brook.
Rohlf, F. J. 2001b. TpsRelw, version 1.24. Dept. of Ecology & Evolution, State University of New York at Stony Brook.
Rohlf, F. J., and D. E. Slice. 1989. GRF: program for generalized rotational fitting. Dept. of Ecology & Evolution, State University of New York at Stony Brook.
Rohlf, F. J., and D. E. Slice. 1990. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst. Zool. 39: 40-59.
Rohlf, F. J., and L. F. Marcus. 1993. A revolution in morphometrics. Trends Ecol. Evol. 8: 129-132.
Sasakawa, M. 1960. A study of the Japanese Agromyzidae (Diptera) Part 1. Sci. Rep. Saikyo Univ. Agric. 12: 76-82.
Sasakawa, M. 1961. A study of the Japanese Agromyzidae (Diptera). Part 2. Pacific Insect 3: 307-472.
Sasakawa, M. 1963. Oriental Agromyzidae (Diptera) in Bishop Museum, Part. 1. Pac. Insects 5: 23-50.
Scheffer, S. J., and M. L. Lewis. 2000. Two nuclear genes confirm mitochondrial evidence of cryptic species within
Liriomyza huidobrensis (Diptera:
Agromyzidae). Ann. Entomol. Soc. Am. 94: 648-653.
Scheffer, S. J., A. Wijesekara, D. Visser, and R. H. Hallett. 2001. Polymerase chain reaction-restriction fragment-length polymorphism method to distinguish
Liriomyza huidobrensis from L. langei
(Diptera: Agromyzidae) applied to three recent leafminer invasions. J. Econ. Entomol. 94:1177-1182.
Shiao, S. F. 1991. Systematic studies on
Liriomyza species in Taiwan. M. S.
thesis, National Taiwan University, Taipei, Taiwan. (in Chinese)
Shiao, S. F., and W. J. Wu. 2000. Liriomyza
huidobrensis (Blanchard), a newly
invaded insect of economic importance to Taiwan (Diptera: Agromyzidae). Plant Prot. Bull. 42: 249-254.
Spencer, K. A. 1973. Agromyzidae (Diptera) of economic importance. Dr. W. Junk B. V., Publishers, The Hague, The Netherlands.
Spencer, K. A. 1987. Agromyzidae, Ch.73. pp. 869-870. In J. F. McAlpine et al. (eds.), Manual of Nearctic Diptera, Vol. 2. Research Branch, Agriculture Canada, Monograph No. 28.
Spencer, K. A. 1989. Leaf miner, Ch. 5. pp. 77-98. In R. P. Kahn (ed.), Plant protection and quarantine, Vol. 2, selected pests and pathogens of quarantine significance. CRC Press, Inc., Boca Raton.
Spencer, K. A. 1990. Host specialization in the world Agromyzidae (Diptera). Kluwer Academic Publishers, The Netherlands.
Spencer, K. A. 1992. Flycatcher: memoirs of an amateur entomologist. SPB Acad. Pub., The Hague, The Netherlands. Spencer, K. A., and G. C. Steyskal. 1986.
Manual of the Agromyzidae (Diptera) of the United States. U. S. Dep. Agric., A. R. S., Agriculture Handbook No. 638.
Thompson, D. W. 1917. On growth and form. Cambridge Univ. Press, Cambridge.
Trumble, J. T. 1981. Liriomyza trifolii: could become a problem on celery. Calif. Agric. 35: 30-31.
Tschirnhaus, M. von. 1971. Unbekannte stridulationsorgame bei Dipteren und ihre bedeutung fur taxonomie und phylogenetik der Agromyziden. Beitr. Entomol. 21: 551-579.
Tschirnhaus, M. von. 1981. Die Halm– und Minierfliegen im Grenzbereich Land– Meer der Nordsee. Spixiana, Suppl. 6. pp. 1-405.
Wang, C. L., and F. C. Lin. 1988. A newly invaded insect pest Liriomyza
trifolii (Diptera: Agromyzidae) in
Taiwan. J. Agric. Res. China 37: 453-457.
Zehnder, G. W., J. T. Trumble, and W. R. White. 1983. Discrimination of
Liriomyza species (Diptera:
Agromyzidae) using electrophoresis and scanning microscopy. Proc. Entomol. Soc. Wash. 85: 564-574.
Zhang, Q., Z. Lin, H. Chen, X. Wang, and X. Chen. 2001. A preliminary study of Agromyzidae (Diptera) in Fujian province, China. Entomol. J. East China 10: 11-19. (in Chinese)
附件:封面格式
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※
近緣種之多食性潛蠅形態進化與系統學研究
※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:█個別型計畫 □整合型計畫
計畫編號:
NSC 90-2313-B-002-334執行期間:90 年 8 月 1 日至 91 年 7 月 31 日
計畫主持人:蕭旭峰
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立台灣大學昆蟲學系
中 華 民 國
91 年 10 月 31 日
11附圖
Fig. 1. Diagrams of abdominal color patterns of six Liriomyza species. A, L. bryoniae; B, L.
huidobrensis; C, L. trifolii; D, L. sativae; E, L. brassicae; F, L. chinensis. I to VI indicate
the first to sixth visible abdominal tergites; a, the third visible tergite is divided by the yellow middle furrow in L. bryoniae; b, the second to fifth tergites are divided by the yellow middle furrow in L. trifolii.
Fig. 2. Arrangements of thoracic microsetae of six Liriomyza species. A and B indicate the location of microsetae on thoracic tergite and their enlargement; C, L. bryoniae; D, L.
huidobrensis; E, L. trifolii; F, L. sativae; G, L. brassicae; H, L. chinensis. Scale bar = 20
μm.
Fig. 3. Local enlargements of thoracic microsetae of Liriomyza species. A, L. bryoniae; B, L.
trifolii; C, L. brassicae; D, L. chinensis. Scale bar = 6 μm.
Fig. 4. Result of generalized least-squares analysis for superimposing 16 landmarks on the wings of six Liriomyza species. First and second principal component axes with one standard deviation length and equal frequency ellipses of two standard deviations were presented for each landmark. Arrows indicate the location of landmarks (#14 & #15) on outer cross vein that have directional variation.
Fig. 5. Minimal spanning tree was superimposed onto the scatterplot of the second relative warp (RW2) against the first relative warp (RW1) of wing shapes of six Liriomyza species. Shape changes of each species were represented by deformation grids. The first and second axes account for 57.6 and 30.2% of the total variance, respectively. BRA, Liriomyza
brassicae; BRY, L. bryoniae; CHI, L. chinensis; HUD, L. huidobrensis; SAT, L. sativae;
TRI, L. trifolii.
Fig. 6. Photo plates of phallus in six Liriomyza species, lateral view. A, L. bryoniae; B, L.
huidobrensis; C, L. trifolii; D, L. sativae; E, L. brassicae; F, L. chinensis. a, a’, b, b’, c, and
d indicate the location of distiphallus. Scale bar = 0.1 mm.
Fig. 7. Photo plates of phallus in six Liriomyza species, ventral view. A, L. bryoniae; B, L.
huidobrensis; C, L. trifolii; D, L. sativae; E, L. brassicae; F, L. chinensis; a, a’, c, c’, d, and
e indicate the location of distiphallus; b and b’ indicate the location of mesophallus. Scale bar = 0.1 mm.
Fig. 1
A B
I
II
III
IV
V
VI
a
C
D
b
E
F
14Fig. 2
A
B
C
D
F
E
G
H
15
A
B
C
D
Fig. 3 16Fig.4 1 2
C
3R
2+3R
4+5 7 4 9 8 10 11 13M
1+2 12 14 16 5 15M
3+4 6 17Fig. 5
BRA
SAT
HUI
CHI
TRI
BRY
Fig. 6
C
D
A
B
d
c
b’
b
a’
a
E
F
19Fig.7