Association analysis of dopaminergic gene variants (Comt, Drd4 And Dat1) with
Alzheimer's disease
Wei-Yong Lin a, b Bor-Tsang Wu c Chun-Cheng Lee b, d Jim Jinn-Chyuan Sheu b, e Shu-Hsiang Liu f Wen-Fu Wang g Chon-Haw Tsai h, i Hsin-Ping Liu j Fuu-Jen Tsai b, e, f
a Graduate Institute of Integrated Medicine, China Medical University, Taichung, Taiwan b Department of Medical Research, China Medical University Hospital, Taichung, Taiwan c Department of Physical Therapy, China Medical University, Taichung, Taiwan
d School of Medicine, China Medical University, Taichung, Taiwan
e School of Chinese Medicine, China Medical University, Taichung, Taiwan f Department of Biotechnology, Asia University, Taichung, Taiwan
g Department of Neurology, Chang-Hua Christian Hospital, Chang-Hua, Taiwan
h Graduate Institute of Neural and Cognitive Sciences, China Medical University, Taichung, Taiwan
i Department of Neurology, China Medical University Hospital, Taichung, Taiwan
j Graduate Institute of Acupuncture Science, China Medical University, Taichung, Taiwan
Wei-Yong Lin and Bor-Tsang Wu contributed equally to this work. Corresponding authors:
Dr. Hsin-Ping Liu Graduate Institute of Acupuncture Science, China Medical University, No.91 Hsueh-Shih Road, Taichung 404, Taiwan. Tel. +886 4 22053366 ext 3603, Fax. +886 4 22035191, E-mail [email protected]
Dr. Fuu-Jen Tsai Department of Medical Research, China Medical University Hospital, No. 2 Yuh Der Road, Taichung 404, Taiwan. Tel. +886 4 22062121 ext 7076, Fax. +886 4 22033295, E-mail [email protected]
Abstract
Background: Defects in dopaminergic transmission play important roles in the
disturbance of synaptic plasticity and even in advanced cognitive behavior. However, the relationship between genes involved in the regulation of dopamine levels and predisposition for Alzheimer’s disease (AD) remains unclear. Aims: The potential association of dopamine-modulating gene polymorphisms with AD was evaluated.
Methods: We performed a case-control study with 120 patients and 86 healthy
controls. Two catechol-O-methyltransferase (COMT) single-nucleotide polymorphisms (SNPs) (rs2020917 and rs4646312), two dopamine D4 receptor (DRD4) SNPs (rs3758653 and rs916455), and four dopamine transporter (DAT1) SNPs (rs2937639, rs6347, rs12516948 and rs11133762) were investigated. Results: The T allele at the DRD4 SNP (rs3758653) was found to be significantly associated with AD. Our results also showed that haplotype frequencies, observed from the analyzed SNPs, were distributed significantly differently in AD patients verse control subjects. Moreover, a strong association was observed between the A allele at rs6347 of DAT1 and moderate stage of dementia. Conclusion: These observations suggest that genetic variations in the dopamine-modulating genes, COMT, DRD4 and DAT1, may contribute to AD pathogenesis in the Taiwanese population.
Introduction
Alzheimer’s disease (AD) is the most common form of dementia worldwide. Its clinical symptoms include progressive neurodegeneration and cognitive dysfunction, and its incidence is strongly age-related [1]. The brains of patients with AD display two neuropathological features: aggregates of senile plaques and neurofibrillary tangles, which are primarily composed of amyloid-beta (Aβ) peptides and hyperphosphorylated Tau proteins, respectively [2, 3]. Although the pathology of AD has been well characterized, most cases are sporadic and genetically complicated, which indicates that other genetic factors are responsible for the development of AD.
Certain neurotransmitters and their receptors and transporters are considered to be possible risk factors for AD. One such neurotransmitter is acetylcholine. Defects in cholinergic neuron activity may account for the cognitive decline in AD patients [4-6]. Treating this dysfunction is a current strategy in AD treatment. Another excitatory neurotransmitter system is the glutamate. Oligomeric Aβ peptides may disturb synaptic plasticity by altering the activity, number and trafficking of AMPA and NMDA glutamate receptors [7, 8]. Parameshwaran et al., provided evidence that Aβ peptides inhibit both the amplitude and frequency of postsynaptic currents through AMPA receptors (AMPARs) and decrease the probability of channel opening in hippocampal CA1 pyramidal neurons [9]. Immunoprecipitation analyses have demonstrated that assembly forms of Aβ peptides can associate with NMDA receptors (NMDARs) at their NR1 and NR2A subunits [10]. A peptides were found to induce massive calcium ion permeability in a rat study, resulting in excitotoxicity and neuronal death, and this effect was alleviated by MK-801, which is an NMDAR antagonist [11]. Through selective interactions between NMDARs and Aβ peptides, changes in synaptic plasticity, such as deficits in long-term potentiation (LTP) [12,
13] and long-term depression (LTD) induction, have been detected in rat hippocampus [14, 15]. These results seem to demonstrate that A-impaired synaptic plasticity may have deleterious effects on neuronal activity, leading to loss of synapses or dendrites [16-18], and this impairment might be responsible for the neurodegenerative process and cognitive dysfunction of AD.
Previous studies have suggested that more than just the cholinergic and glutamatergic neurotransmission systems influence synaptic failure and neuronal death in AD. Stimulation of dopamine receptors has been shown to promote AMPAR trafficking from the cytosol to the synaptic membrane, implying that newly recruited surface AMPARs may facilitate LTP induction and demonstrating the critical role of dopaminergic transmission in neuronal plasticity [19, 20]. Another report indicated that activation of dopamine receptors could prevent internalization of AMPA and NMDA receptors that is caused by oligomeric A peptides [21]. These changes in synaptic plasticity might be essential for advanced cognitive behavior and memory formation. Therefore, we explored the relationship between some genes involved in dopaminergic transmission and susceptibility to AD progression. This type of associative relationship has profound implications regarding correlations between genetic markers in the catechol-O-methyltransferase (COMT), dopamine D4 receptor (DRD4) and dopamine transporter (DAT1) genes and disease progression in the Taiwanese population.
Materials and Methods
Subjects
This study recruited 120 patients with late-onset of AD (mean age at onset, 74.9 ± 6.7, 54.2% female) and 86 age-matched normal control subjects (mean age 65.9 ± 8.7, 73% female). Participants were enrolled from the Chang-Hua Christian Hospital and China Medical University Hospital. All patients were clinically diagnosed according to guidelines from the National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer’s Disease and Related Disorders Association (NINCDS–ADRDA). Dementia was diagnosed based on Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) criteria. An institutional ethics committee approved this study, and informed consent was obtained from all subjects.
Genotyping of gene variants in COMT, DRD4 and DAT1
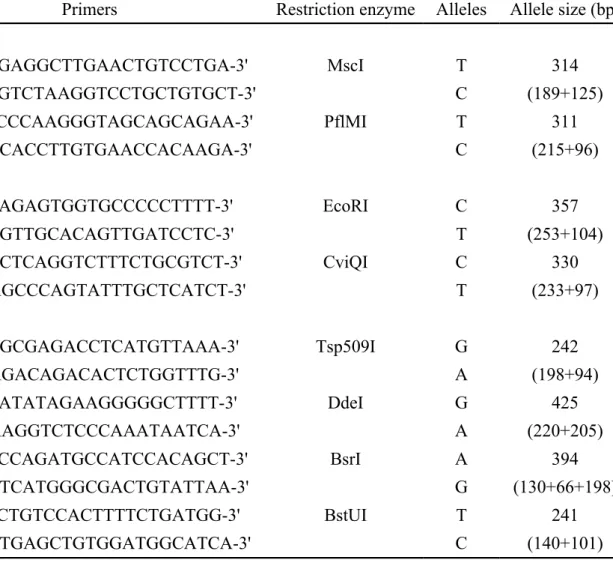
Genomic DNA was extracted from peripheral blood samples according to a standard protocol (Genomic DNA kit, Qiagen, CA). For analysis of COMT gene variants, two polymorphic sites located upstream of the coding region, 5’ distal promoter region, were selected for this study from the public dbSNP database: C/T (rs2020917) and T/C (rs4646312). To study variants in the dopamine D4 receptor (DRD4), two SNPs located in the 5’ region near the gene T/C (rs3758653) and C/T (rs916455) were selected for this study. To genotype dopamine transporter DAT1 (locus symbol: SLC6A3) variants, four SNPs located in the gene region were chosen. One is A/G SNP in intron 1 (rs2937639), one is a synonymous 1336 A/G SNP in exon 9 (rs6347), and A/G (rs12516948) and C/T (rs11133762) are two SNPs in the 3’ region near the gene. To identify the allele preference for these SNPs, PCR-Restriction Fragment Length Polymorphism (PCR-RFLP) analyses were performed. Specific primers and restriction enzymes for each PCR-RFLP reaction are shown in
table 1. In brief, PCR amplification was carried out in a total volume of 25 µl with 5 ng genomic DNA and primer pairs. PCR reactions were carried out at 95°C for 5 min, followed by 40 cycles of 95°C for 30 sec, specific Tm temperatures for each primer pair for 30 sec, and 72°C for 40 sec, with a final elongation step at 72°C for 7 min. Five microliters of the PCR amplicons was digested overnight by various restriction enzymes (New England Biolabs, MA) at 37°C in a total volume of 20 μl. The sizes of the PCR products and digestion fragments were determined by agarose gel electrophoresis and visualized directly by ethidium bromide staining.
Statistical analyses
The distributions of allele and genotype frequencies for each SNP were evaluated by χ2 test using 2 x 2 or 2 x 3 contingency tables. Statistical analysis of the odds ratio (OR) and 95% confidence interval (CI) were carried out using SPSS version 10.0 software (Chicago, IL) based on the presence of the reference allele and genotype frequencies. P values less than 0.05 were considered statistically significant. Adherence to Hardy-Weinberg equilibrium was tested using Pearson’s χ2 test with one degree of freedom. The haplotype approach for each individual was analyzed with Phase v2.1 software program using Bayesian statistical method [22, 23]. P values were determined by comparing a given haplotype with a combination of all other haplotypes.
Results
A total of 206 subjects of Taiwan Han Chinese comprised this cohort. One hundred and twenty subjects were diagnosed as having AD, and eighty-six subjects served as healthy controls. The genotype distributions within each gene variant in this study did not deviate significantly from Hardy-Weinberg equilibrium (all p > 0.1, table 2). After analyzing genotype and allele frequencies, neither of the COMT gene polymorphisms (rs2020917 and rs4646312) showed a statistically significant association with AD (table 2). Genotypic analyses indicated that only one DAT1 gene variant (rs11133762) had a borderline significant allele frequency (p = 0.055). This C/T gene polymorphism located at the 3’ end region of the DAT1 gene was C:T 61.3%:38.7% in AD patients, compared with 51.8%:48.2% in controls (table 2). With genotyping two polymorphisms of DRD4 gene (rs3758653 and rs916455), we found that gene variant, rs3758653, located at the 5’ region, attained statistical difference at the 0.05 level in allele frequency. At this SNP, the T:C ratio was 73.9%:26.1% in the subjects with AD and 62.5%:37.5% in controls. The odds ratio (OR) of the T allele indicated a 1.70-fold higher risk than with the C allele (95% CI, 1.11 to 2.61) (table 2), suggesting that the T allele at rs3758653 is a risk factor that correlates with AD.
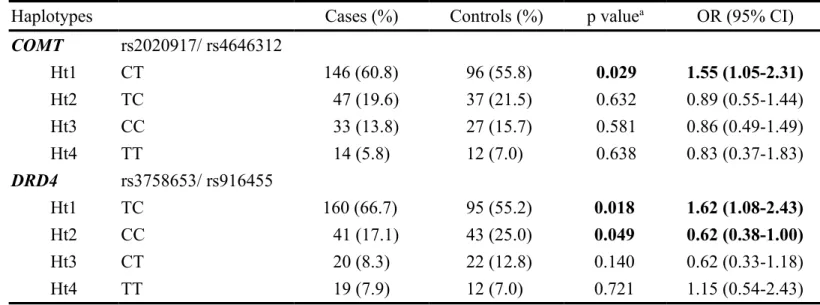
We also analyzed associations between the haplotype distributions of patients and controls. The haplotype frequencies of the COMT gene at two polymorphic loci are shown in table 3, and four haplotypes were observed in both AD patients and controls. The frequency of the most common haplotype (Ht1-CT) in the patients was 60.8%, compared with 55.8% in the controls. After haplotype-specific analysis, this Ht1-CT haplotype appeared to be a significant ‘risk’ haplotype (p = 0.029, OR = 1.55, 95% CI, 1.05 to 2.31) compared with the non-Ht1 haplotype in AD patients and control groups. Similar haplotype-specific analyses showed no significant differences
between the two groups of subjects for the three other haplotypes, Ht2-CC, Ht3-TT and Ht4-TC. In an association analysis of DAD4 gene haplotype distributions, subjects with the Ht1-TC haplotype at the two assayed polymorphisms may have a higher incidence of pathogenic AD (p = 0.018, OR = 1.62, 95% CI, 1.08 to 2.43). Conversely, subjects with the Ht2-CC haplotype may have a lower incidence of AD (p = 0.049, OR = 0.62, 95% CI, 0.38 to 1.00) (table 3).
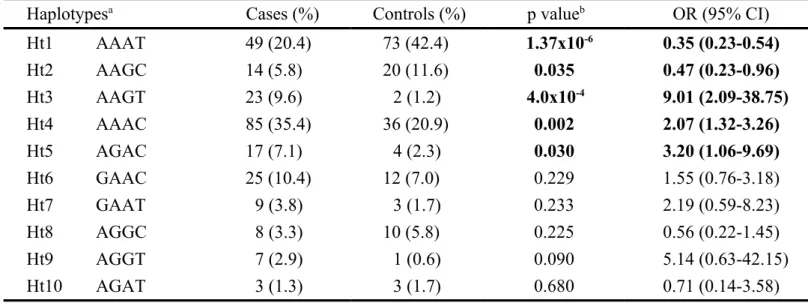
Of ten observed haplotypes from four analyzed polymorphisms in the DAT1 gene, three appeared to show significant risk effects (Ht3-AAGT, p < 0.001; Ht4-AAAC, p = 0.015; and Ht5-AGAC, p = 0.030) between the AD patients and control groups (table 4). This result may be a subtle hint that individuals carrying these three haplotypes have a higher risk of developing AD. Additionally, our results indicate that two observed haplotypes were significantly different between patients and controls, suggesting that subjects carrying the Ht1-AAAT and Ht2-AAGC haplotypes (p < 0.0001 and p = 0.035, respectively) may be protected against AD progression.
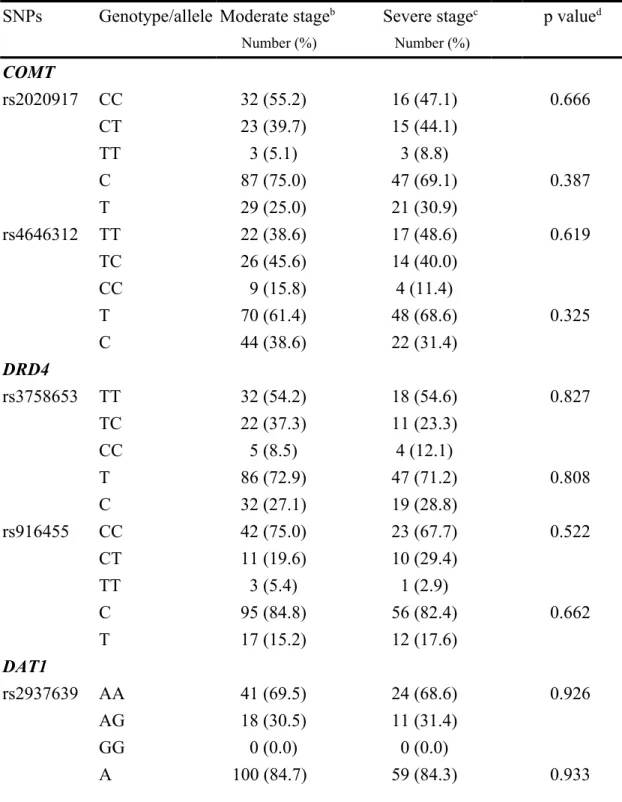
We next asked whether dopamine-modulating gene variants showed associations with the clinical stages of AD. The patients were divided into three groups: a mild stage group with Mini-Mental State Examination (MMSE) scores above 20, a moderate stage group with MMSE scores between 10 and 20, and a severe stage group with MMSE scores less than 10. In the analysis of allele frequencies between the moderate and severe stage groups, strong association was observed between severe stage of AD and the A allele at rs6347 in DAT1 gene (p = 0.025, OR = 2.67, 95% CI, 1.11 to 6.42) (table 5). This result suggests that the A allele could be a protective factor for patients, contributing to a lower risk of developing severe dementia. However, genotypic and allelic analyses of the other genetic variants failed to detect any significant correlations with the different clinical stages.
Discussion
Previous studies have discovered that mutations in genes encoding proteins involved in amyloidogenic processing, including amyloid precursor protein, presenilin 1 and presenilin 2, predominantly lead to amyloid plaque production in early-onset familial cases of AD [24-26]. Excessive A peptides demonstrably increase the risk of synaptic loss and dysfunction, which may disturb brain functions, including advanced cognitive behavior and memory formation, before neuronal death [27, 28]. Most cases of AD are sporadic, and the factors important in disease development are unknown, which motivates us to identify additional genetic risk factors that are also responsible for AD progression. To evaluate whether genetic variations in dopamine-modulating genes, COMT, DRD4 and DAT1 are associated with AD in the Taiwanese population, we performed a case-control study and found these genetic variants and their effects on dopaminergic transmission might be the risk factors in the etiology of AD.
Previous findings indicated that dopaminergic neurotransmission regulates synaptic plasticity. Activation of the dopamine D1/D5 receptor consolidates the late-phase LTP in the CA1 of rat hippocampal slices [29], and reverses the LTD induced by NMDA or low-frequency stimulation [20]. The other study also found that direct injections of dopamine receptor agonists can enhance spatial memory performance [30]. These results suggest that defects in dopaminergic neurotransmission might disturb synaptic plasticity and even advanced cognitive behavior, indicating that dopamine could be a candidate risk factor for AD progression. Within the brain, the transmission efficiency of dopaminergic neurons is obviously associated with dopamine-modulating genes, which are thought to regulate dopamine levels by its production and clearance. COMT is a dopamine-modulating gene that plays an
essential role in dopamine clearance by catalyzing the methylation of the hydroxyl group of dopamine [31]. In our study, the allele and genotype distributions of COMT were found to be insignificantly associated with AD. However, one haplotype, observed involving the two assayed polymorphisms, was associated a significant increase in the risk of AD. Another gene involved in the dopamine-modulating system is the dopamine transporter (DAT1). Because DAT1 functions to control dopamine levels in the synaptic cleft, which limits dopamine receptor activation [32], we tried to explore the relationship between its SNPs and AD, and found that no allele or genotype frequencies of DAT1 showed a significant correlation with AD. Among the ten haplotypes observed involving four analyzed DAT1 gene SNPs, five haplotype distributions showed significant differences between the AD patient and control groups. Interestingly, the A allele at rs6347 in the DAT1 gene was found to be strongly associated with clinical stage, suggesting that patients with an A allele had a lower risk of developing severe dementia. Our data also indicate that the dopamine receptor DRD4 gene variant is a genetic risk factor for AD, and analyses of allele frequency and haplotype distributions indicate that gene variations in DRD4 are significant risk factors for having AD.
The influence of the activities of COMT, DRD4 and DAT1 on AD remains unclear. AD patients with more severe stages of dementia frequently suffer from depression [33] and previous studies have also found that variations in the dopaminergic system may be risk factors for AD and/or depression. For example, a functional genetic variant in COMT, wherein valine to methionine substitution at codon 108/158 decreases its enzyme activity, which increases susceptibility to psychosis in AD [34, 35] and schizophrenia [36]. Functional genomic experiments have indicated that SNPs located at distal promoter region of the COMT gene alter its transcriptional
efficiency, potentially changing the COMT activity [37]. A study by Grunblatt et al. showed that allelic repeats in the DRD4 gene appeared to have no significant influences on the outcome of AD or depressive symptoms in their European population [38]. Previous reports have found that DAT1 gene polymorphism has no correlation with AD etiology [39], but a demonstrable association with schizophrenia phenotype was detected [40]. Here, we determined that COMT, DRD4 and DAT1 gene variants have associations with AD in the Taiwanese population. In contrast with prior studies, we found significant associations between these dopamine-modulating genes and AD development. Our data provide some guidance for understanding how dopaminergic-system gene variations lead to AD susceptibility and identification of AD-modulating genes opens a new path for understanding the pathogenesis of this disease.
Acknowledgements This study was supported by grants from the China Medical University and Hospital (CMU98-S-21, DMR-93-001) and supported in part by Taiwan Department of Health Clinical Trial and Research Center of Excellence (DOH100-TD-B-111-004).
References
1 Kalaria RN, Maestre GE, Arizaga R, Friedland RP, Galasko D, Hall K, Luchsinger JA, Ogunniyi A, Perry EK, Potocnik F, Prince M, Stewart R, Wimo A, Zhang ZX, Antuono P: Alzheimer's disease and vascular dementia in developing countries: Prevalence, management, and risk factors. Lancet Neurol 2008;7:812-826.
2 Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K: Amyloid plaque core protein in alzheimer disease and down syndrome. Proc Natl Acad Sci U S A 1985;82:4245-4249.
3 Goedert M, Wischik CM, Crowther RA, Walker JE, Klug A: Cloning and sequencing of the cdna encoding a core protein of the paired helical filament of alzheimer disease: Identification as the microtubule-associated protein tau. Proc Natl Acad Sci U S A 1988;85:4051-4055.
4 Terry AV, Jr., Buccafusco JJ: The cholinergic hypothesis of age and alzheimer's disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J Pharmacol Exp Ther 2003;306:821-827.
5 Bartus RT, Dean RL, 3rd, Beer B, Lippa AS: The cholinergic hypothesis of geriatric memory dysfunction. Science 1982;217:408-414.
6 Sivaprakasam K: Towards a unifying hypothesis of alzheimer's disease: Cholinergic system linked to plaques, tangles and neuroinflammation. Curr Med Chem 2006;13:2179-2188.
7 Gasparini L, Dityatev A: Beta-amyloid and glutamate receptors. Exp Neurol 2008;212:1-4.
8 Parameshwaran K, Dhanasekaran M, Suppiramaniam V: Amyloid beta peptides and glutamatergic synaptic dysregulation. Exp Neurol 2008;210:7-13.
9 Parameshwaran K, Sims C, Kanju P, Vaithianathan T, Shonesy BC, Dhanasekaran M, Bahr BA, Suppiramaniam V: Amyloid beta-peptide abeta(1-42) but not abeta(1-40) attenuates synaptic ampa receptor function. Synapse 2007;61:367-374.
10 Venkitaramani DV, Chin J, Netzer WJ, Gouras GK, Lesne S, Malinow R, Lombroso PJ: Beta-amyloid modulation of synaptic transmission and plasticity. J Neurosci 2007;27:11832-11837.
11 Harkany T, Abraham I, Timmerman W, Laskay G, Toth B, Sasvari M, Konya C, Sebens JB, Korf J, Nyakas C, Zarandi M, Soos K, Penke B, Luiten PG: Beta-amyloid neurotoxicity is mediated by a glutamate-triggered excitotoxic cascade in rat nucleus basalis. Eur J Neurosci 2000;12:2735-2745.
12 Larson J, Lynch G, Games D, Seubert P: Alterations in synaptic transmission and long-term potentiation in hippocampal slices from young and aged pdapp mice.
Brain Res 1999;840:23-35.
13 Walsh DM, Klyubin I, Fadeeva JV, Cullen WK, Anwyl R, Wolfe MS, Rowan MJ, Selkoe DJ: Naturally secreted oligomers of amyloid beta protein potently inhibit hippocampal long-term potentiation in vivo. Nature 2002;416:535-539.
14 Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, Smith I, Brett FM, Farrell MA, Rowan MJ, Lemere CA, Regan CM, Walsh DM, Sabatini BL, Selkoe DJ: Amyloid-beta protein dimers isolated directly from alzheimer's brains impair synaptic plasticity and memory. Nat Med 2008;14:837-842.
15 Li S, Hong S, Shepardson NE, Walsh DM, Shankar GM, Selkoe D: Soluble oligomers of amyloid beta protein facilitate hippocampal long-term depression by disrupting neuronal glutamate uptake. Neuron 2009;62:788-801.
16 Mucke L, Masliah E, Yu GQ, Mallory M, Rockenstein EM, Tatsuno G, Hu K, Kholodenko D, Johnson-Wood K, McConlogue L: High-level neuronal expression of abeta 1-42 in wild-type human amyloid protein precursor transgenic mice: Synaptotoxicity without plaque formation. J Neurosci 2000;20:4050-4058.
17 Chapman PF, White GL, Jones MW, Cooper-Blacketer D, Marshall VJ, Irizarry M, Younkin L, Good MA, Bliss TV, Hyman BT, Younkin SG, Hsiao KK: Impaired synaptic plasticity and learning in aged amyloid precursor protein transgenic mice. Nat Neurosci 1999;2:271-276.
18 Walsh DM, Klyubin I, Fadeeva JV, Rowan MJ, Selkoe DJ: Amyloid-beta oligomers: Their production, toxicity and therapeutic inhibition. Biochem Soc Trans 2002;30:552-557.
19 Gao C, Sun X, Wolf ME: Activation of d1 dopamine receptors increases surface expression of ampa receptors and facilitates their synaptic incorporation in cultured hippocampal neurons. J Neurochem 2006;98:1664-1677.
20 Mockett BG, Guevremont D, Williams JM, Abraham WC: Dopamine d1/d5 receptor activation reverses nmda receptor-dependent long-term depression in rat hippocampus. J Neurosci 2007;27:2918-2926.
21 Jurgensen S, Antonio LL, Mussi GE, Brito-Moreira J, Bomfim TR, De Felice FG, Garrido-Sanabria ER, Cavalheiro EA, Ferreira ST: Activation of d1/d5 dopamine receptors protects neurons from synapse dysfunction induced by amyloid-beta oligomers. J Biol Chem 2011;286:3270-3276.
22 Stephens M, Donnelly P: A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am J Hum Genet 2003;73:1162-1169.
23 Stephens M, Smith NJ, Donnelly P: A new statistical method for haplotype reconstruction from population data. Am J Hum Genet 2001;68:978-989.
A pathogenic mutation for probable alzheimer's disease in the app gene at the n-terminus of beta-amyloid. Nat Genet 1992;1:345-347.
25 Sherrington R, Rogaev EI, Liang Y, Rogaeva EA, Levesque G, Ikeda M, Chi H, Lin C, Li G, Holman K, Tsuda T, Mar L, Foncin JF, Bruni AC, Montesi MP, Sorbi S, Rainero I, Pinessi L, Nee L, Chumakov I, Pollen D, Brookes A, Sanseau P, Polinsky RJ, Wasco W, Da Silva HA, Haines JL, Perkicak-Vance MA, Tanzi RE, Roses AD, Fraser PE, Rommens JM, St George-Hyslop PH: Cloning of a gene bearing missense mutations in early-onset familial alzheimer's disease. Nature 1995;375:754-760.
26 Steiner H, Romig H, Grim MG, Philipp U, Pesold B, Citron M, Baumeister R, Haass C: The biological and pathological function of the presenilin-1 deltaexon 9 mutation is independent of its defect to undergo proteolytic processing. J Biol Chem 1999;274:7615-7618.
27 Kim JH, Anwyl R, Suh YH, Djamgoz MB, Rowan MJ: Use-dependent effects of amyloidogenic fragments of (beta)-amyloid precursor protein on synaptic plasticity in rat hippocampus in vivo. J Neurosci 2001;21:1327-1333.
28 Yoshiyama Y, Higuchi M, Zhang B, Huang SM, Iwata N, Saido TC, Maeda J, Suhara T, Trojanowski JQ, Lee VM: Synapse loss and microglial activation precede tangles in a p301s tauopathy mouse model. Neuron 2007;53:337-351. 29 Navakkode S, Sajikumar S, Frey JU: Synergistic requirements for the induction of
dopaminergic d1/d5-receptor-mediated ltp in hippocampal slices of rat ca1 in vitro. Neuropharmacology 2007;52:1547-1554.
30 White NM, Packard MG, Seamans J: Memory enhancement by post-training peripheral administration of low doses of dopamine agonists: Possible autoreceptor effect. Behav Neural Biol 1993;59:230-241.
31 Mannisto PT, Kaakkola S: Catechol-o-methyltransferase (comt): Biochemistry, molecular biology, pharmacology, and clinical efficacy of the new selective comt inhibitors. Pharmacol Rev 1999;51:593-628.
32 Vandenbergh DJ, Persico AM, Hawkins AL, Griffin CA, Li X, Jabs EW, Uhl GR: Human dopamine transporter gene (dat1) maps to chromosome 5p15.3 and displays a vntr. Genomics 1992;14:1104-1106.
33 Fischer P, Simanyi M, Danielczyk W: Depression in dementia of the alzheimer type and in multi-infarct dementia. Am J Psychiatry 1990;147:1484-1487.
34 Borroni B, Agosti C, Archetti S, Costanzi C, Bonomi S, Ghianda D, Lenzi GL, Caimi L, Di Luca M, Padovani A: Catechol-o-methyltransferase gene polymorphism is associated with risk of psychosis in alzheimer disease. Neurosci Lett 2004;370:127-129.
Chowdari KV, DeKosky ST, Ferrell RE: Catechol-o-methyltransferase haplotypes are associated with psychosis in alzheimer disease. Mol Psychiatry 2005;10:1026-1036.
36 Glatt SJ, Faraone SV, Tsuang MT: Association between a functional catechol o-methyltransferase gene polymorphism and schizophrenia: Meta-analysis of case-control and family-based studies. Am J Psychiatry 2003;160:469-476.
37 Ji Y, Olson J, Zhang J, Hildebrandt M, Wang L, Ingle J, Fredericksen Z, Sellers T, Miller W, Dixon JM, Brauch H, Eichelbaum M, Justenhoven C, Hamann U, Ko Y, Bruning T, Chang-Claude J, Wang-Gohrke S, Schaid D, Weinshilboum R: Breast cancer risk reduction and membrane-bound catechol o-methyltransferase genetic polymorphisms. Cancer Res 2008;68:5997-6005.
38 Grunblatt E, Zehetmayer S, Bartl J, Loffler C, Wichart I, Rainer MK, Jungwirth S, Bauer P, Danielczyk W, Tragl KH, Riederer P, Fischer P: Genetic risk factors and markers for alzheimer's disease and/or depression in the vita study. J Psychiatr Res 2009;43:298-308.
39 Nicholl DJ, Bennett P, Hiller L, Bonifati V, Vanacore N, Fabbrini G, Marconi R, Colosimo C, Lamberti P, Stocchi F, Bonuccelli U, Vieregge P, Ramsden DB, Meco G, Williams AC: A study of five candidate genes in parkinson's disease and related neurodegenerative disorders. European study group on atypical parkinsonism. Neurology 1999;53:1415-1421.
40 Cordeiro Q, Siqueira-Roberto J, Vallada H: Association between the slc6a3 a1343g polymorphism and schizophrenia. Arq Neuropsiquiatr 2010;68:716-719.
Table 1. Primers information for testing SNPs in the COMT, DRD4 and DAT1 genes by PCR-RFLP
SNPs Primers Restriction enzyme Alleles Allele size (bp)
COMT rs2020917 F: 5'-ATGAGGCTTGAACTGTCCTGA-3' MscI T 314 R: 5'-TTGTCTAAGGTCCTGCTGTGCT-3' C (189+125) rs4646312 F: 5'-TTCCCAAGGGTAGCAGCAGAA-3' PflMI T 311 R: 5'-TGCACCTTGTGAACCACAAGA-3' C (215+96) DRD4 rs3758653 F: 5'-ACAGAGTGGTGCCCCCTTTT-3' EcoRI C 357 R: 5'-CCGTTGCACAGTTGATCCTC-3' T (253+104) rs916455 F: 5'-AGCTCAGGTCTTTCTGCGTCT-3' CviQI C 330 R: 5'-GAGCCCAGTATTTGCTCATCT-3' T (233+97) DAT1 rs2937639 F: 5'-GAGCGAGACCTCATGTTAAA-3' Tsp509I G 242 R: 5'-AAGACAGACACTCTGGTTTG-3' A (198+94) rs6347 F: 5'-ACATATAGAAGGGGGCTTTT-3' DdeI G 425 R: 5'-AAAGGTCTCCCAAATAATCA-3' A (220+205) rs12516948 F: 5'-TACCAGATGCCATCCACAGCT-3' BsrI A 394 R: 5'-CCTCATGGGCGACTGTATTAA-3' G (130+66+198) rs11133762 F: 5'-CTCTGTCCACTTTTCTGATGG-3' BstUI T 241 R: 5'-TGTGAGCTGTGGATGGCATCA-3' C (140+101)
Table 2. Genotype and allele frequencies of the tested SNPs in the COMT, DAT1 and
DRD4 genes in AD patients and controls
SNPs Genotype/allele Cases HWEa Controls HWE p valueb OR (95% CI)
Number (%) Number (%) COMT rs2020917 CC 63 (54.3) 0.713 41 (48.8) 0.545 0.740 1.32 (0.41-4.20) CT 46 (39.7) 37 (44.0) 1.07 (0.33-3.44) TT 7 (6.0) 6 (7.2) 1 C 172 (74.1) 119 (70.8) 0.464 1.18 (0.76-1.84) T 60 (25.9) 49 (29.2) 1 rs4646312 TT 55 (47.0) 0.329 31 (36.0) 0.180 0.169 1.06 (0.42-2.72) TC 47 (40.2) 46 (53.5) 0.61 (0.24-1.54) CC 15 (12.8) 9 (10.5) 1 T 157 (67.1) 108 (62.8) 0.368 1.21 (0.80-1.82) C 77 (32.9) 64 (37.2) 1 DAT1 rs2937639 AA 84 (70.6) 0.219 59 (72.0) 0.140 0.702 1.06 (0.56-2.02) AG 34 (28.6) 23 (29.0) 1 GG 1 (0.8) 0 (0.0) A 202 (84.9) 141 (86.0) 0.759 0.92 (0.52-1.61) G 36 (15.1) 23 (14.0) 1 rs6347 AA 76 (71.1) 0.421 62 (75.6) 0.733 0.515 0.31 (0.03-2.81) AG 27 (25.2) 19 (23.2) 0.36 (0.04-3.43) GG 4 (3.7) 1 (1.2) 1 A 179 (83.6) 143 (87.2) 0.336 0.75 (0.42-1.35) G 35 (16.4) 21 (12.8) 1 rs12516948 AA 71 (60.2) 0.231 53 (61.6) 0.615 0.879 0.74 (0.24-2.35) AG 38 (32.2) 28 (32.6) 0.75 (0.23-2.50) GG 9 (7.6) 5 (5.8) 1 A 180 (76.3) 134 (77.9) 0.698 0.91 (0.57-1.46) G 56 (23.7) 38 (22.1) 1 rs11133762 CC 47 (39.5) 0.391 24 (28.6) 0.520 0.183 2.06 (0.94-4.51) CT 52 (43.7) 39 (46.4) 1.40 (0.67-2.93) TT 20 (16.8) 21 (25.0) 1 C 146 (61.3) 87 (51.8) 0.055 1.48 (0.99-2.20)
T 92 (38.7) 81 (48.2) 1 DRD4 rs3758653 TT 67 (57.3) 0.152 34 (40.5) 0.580 0.057 2.33 (0.94-5.74) TC 39 (33.3) 37 (44.0) 1.25 (0.50-3.13) CC 11 (9.4) 13 (15.5) 1 T 173 (73.9) 105 (62.5) 0.014 1.70 (1.11-2.61) C 61 (26.1) 63 (37.5) 1 rs916455 CC 79 (69.9) 0.281 54 (64.3) 0.705 0.698 1.17 (0.30-4.56) CT 29 (25.7) 26 (31.0) 0.89 (0.22-3.68) TT 5 (4.4) 4 (4.8) 1 C 187 (82.7) 134 (79.8) 0.451 1.22 (0.73-2.03) T 39 (17.3) 34 (20.2) 1
a HWE, p values of deviation from Hardy-Weinberg equilibrium constant. b p value was compared by 2 test. p values less than 0.05 are highlighted in bold. OR, odds ratio; 95% CI, 95% confidence interval.
Table 3. Haplotype frequencies of COMT and DRD4 polymorphisms between AD patients and controls
Haplotypes Cases (%) Controls (%) p valuea OR (95% CI)
COMT rs2020917/ rs4646312 Ht1 CT 146 (60.8) 96 (55.8) 0.029 1.55 (1.05-2.31) Ht2 TC 47 (19.6) 37 (21.5) 0.632 0.89 (0.55-1.44) Ht3 CC 33 (13.8) 27 (15.7) 0.581 0.86 (0.49-1.49) Ht4 TT 14 (5.8) 12 (7.0) 0.638 0.83 (0.37-1.83) DRD4 rs3758653/ rs916455 Ht1 TC 160 (66.7) 95 (55.2) 0.018 1.62 (1.08-2.43) Ht2 CC 41 (17.1) 43 (25.0) 0.049 0.62 (0.38-1.00) Ht3 CT 20 (8.3) 22 (12.8) 0.140 0.62 (0.33-1.18) Ht4 TT 19 (7.9) 12 (7.0) 0.721 1.15 (0.54-2.43)
a p value was compared by 2 test. p values less than 0.05 are highlighted in bold. OR, odds ratio; 95% CI, 95% confidence interval.
Table 4. Haplotype frequencies of DAT1 polymorphisms in AD patients and controls
Haplotypesa Cases (%) Controls (%) p valueb OR (95% CI)
Ht1 AAAT 49 (20.4) 73 (42.4) 1.37x10-6 0.35 (0.23-0.54) Ht2 AAGC 14 (5.8) 20 (11.6) 0.035 0.47 (0.23-0.96) Ht3 AAGT 23 (9.6) 2 (1.2) 4.0x10-4 9.01 (2.09-38.75) Ht4 AAAC 85 (35.4) 36 (20.9) 0.002 2.07 (1.32-3.26) Ht5 AGAC 17 (7.1) 4 (2.3) 0.030 3.20 (1.06-9.69) Ht6 GAAC 25 (10.4) 12 (7.0) 0.229 1.55 (0.76-3.18) Ht7 GAAT 9 (3.8) 3 (1.7) 0.233 2.19 (0.59-8.23) Ht8 AGGC 8 (3.3) 10 (5.8) 0.225 0.56 (0.22-1.45) Ht9 AGGT 7 (2.9) 1 (0.6) 0.090 5.14 (0.63-42.15) Ht10 AGAT 3 (1.3) 3 (1.7) 0.680 0.71 (0.14-3.58)
a Haplotypes are shown in sequence: rs2937639G/A, rs6347G/A, rs12516948A/G, and rs11133762T/C. b p value was compared by 2 test. p values less than 0.05 are
Table 5. Genotype and allele frequencies of the tested SNPs in the COMT, DRD4 and
DAT1 genes in AD patients at different clinical stagesa
SNPs Genotype/allele Moderate stageb Severe stagec p valued
Number (%) Number (%) COMT rs2020917 CC 32 (55.2) 16 (47.1) 0.666 CT 23 (39.7) 15 (44.1) TT 3 (5.1) 3 (8.8) C 87 (75.0) 47 (69.1) 0.387 T 29 (25.0) 21 (30.9) rs4646312 TT 22 (38.6) 17 (48.6) 0.619 TC 26 (45.6) 14 (40.0) CC 9 (15.8) 4 (11.4) T 70 (61.4) 48 (68.6) 0.325 C 44 (38.6) 22 (31.4) DRD4 rs3758653 TT 32 (54.2) 18 (54.6) 0.827 TC 22 (37.3) 11 (23.3) CC 5 (8.5) 4 (12.1) T 86 (72.9) 47 (71.2) 0.808 C 32 (27.1) 19 (28.8) rs916455 CC 42 (75.0) 23 (67.7) 0.522 CT 11 (19.6) 10 (29.4) TT 3 (5.4) 1 (2.9) C 95 (84.8) 56 (82.4) 0.662 T 17 (15.2) 12 (17.6) DAT1 rs2937639 AA 41 (69.5) 24 (68.6) 0.926 AG 18 (30.5) 11 (31.4) GG 0 (0.0) 0 (0.0) A 100 (84.7) 59 (84.3) 0.933
G 18 (15.3) 11 (15.7) rs6347 AA 44 (81.5) 17 (60.8) 0.106 AG 9 (16.7) 9 (32.1) GG 1 (1.8) 2 (7.1) A 97 (89.8) 43 (76.8) 0.025 G 11 (10.2) 13 (23.2) rs12516948 AA 37 (63.8) 19 (54.3) 0.600 AG 17 (29.3) 12 (34.3) GG 4 (6.9) 4 (11.4) A 91 (78.4) 50 (71.4) 0.279 G 25 (21.6) 20 (28.6) rs11133762 CC 21 (36.2) 16 (44.4) 0.680 CT 26 (44.8) 15 (41.7) TT 11 (20.0) 5 (13.9) C 68 (58.6) 47 (65.3) 0.363 T 48 (41.4) 25 (34.7)
a Mini-Mental State Examination (MMSE) score. b Moderate stage, patients with MMSE scores 10-20. c Severe stage, patients with MMSE scores below 10. d p value was compared by 2 test. p values less than 0.05 are highlighted in bold.