Characterization of a pea gene responsive to low temperature
Cheng-Che S. Kung, Kai-Wun Yeh, Chu-Yung Lin, and Yih-Ming Chen
1 Department of Botany, National Taiwan University, Taipei, Taiwan 106, Republic of China(Received April 12, 1997; Accepted September 11, 1997)
Abstract. A cold-induced gene, peaci11.8, was isolated by differential screening of a subtracted cDNA library from cold-acclimated etiolated seedlings of pea (Pisum sativum L.). The expression of peaci11.8 gene was investigated. The accumulation of the transcripts began within 6 h of cold treatment and peaked between 4 and 10 days of treatment. The expression of this gene was rapidly down-regulated after the acclimated seedlings were transferred to ambient temperature. This indicated the correlation of the peaci11.8 gene expression and cold treatment. Further studies showed that the expression was also highly stimulated by water stress. The amino acid sequence deduced from the nucleotide sequence of peaci11.8 shared significant homology with several plant cold-induced genes. In addition, the sequence analysis revealed the presence of two common features of many late embryogenesis abundant (LEA) proteins, the lysine-rich motif and the serine cluster. The availibility of peaci11.8 will facilitate the structure-function studies of the cold-inducible gene product.
Keywords: Cold acclimation; Gene expression; Pisum sativum.
Abbreviations: LEA, late embryogenesis abundant; NTS, nuclear-targeting signal; ORF, open reading frame; SSC, 150 mM NaCl/15 mM sodium citrate.
Recently, we found a 19-kDa thylakoid membrane-asso-ciating protein increased remarkably when pea (Pisum sativum) plants were exposed to low temperature (Ma et al., 1990). To obtain a better insight into the mechanisms of cellular acclimation, efforts were made to isolate and characterize cDNA clones corresponding to cold-induced transcripts from pea. In this report, we describe the clon-ing and characterization of a novel gene of pea that re-sponds to low temperature.
Materials and Methods
Plant Material and TreatmentsSeeds of pea (Pisum sativum L. cv. Taichung No. 9) were rinsed with tap water and germinated in rolls of moist paper towel at ambient temperature (28°C) in a dark growth chamber. Two-day-old seedlings were either har-vested without cotyledons or used for cold (4°C) treatment. For cold acclimation, seedlings were rinsed with ice-cold water, then transferred to cold in dark and harvested at prescribed time intervals. For deacclimation, seedlings acclimated for 4 days were transferred back to ambient temperature and harvested at a specified time. For other stresses, seedlings were harvested 1 day after being trans-ferred to 20% PEG 6,000 or 100 mM ABA at ambient tem-perature, respectively.
RNA Isolation
Total and poly(A)+ RNA were extracted according to
the method of Zurfluh and Guilfoyle (1982), except that the poly(A)+ RNA was precipitated in the presence of 0.3
M ammonium acetate and two volumes of ethanol.
Introduction
Temperature is one of the major environmental factors affecting nutrient distribution, growth, and development of plants. Cold-sensitive plants, especially those of tropi-cal or subtropitropi-cal origins, are readily injured or killed at low, nonfreezing temperatures (Levitt, 1980). In contrast, plants of temperate areas are more tolerant to freezing when previously exposed to low positive temperatures, a process known as cold acclimation. Comparative analy-ses have revealed that various biochemical and physiologi-cal changes occur during cold acclimation, including alterations in lipid and carbohydrate compositions as well as protein contents and enzyme activities (Guy, 1990; Thomashow, 1990).
It has been well-demonstrated that changes in gene ex-pression and protein synthesis are associated with induc-tion of freezing tolerance during cold acclimainduc-tion (Gilmour et al., 1988; Guy et al., 1985). Thus far, cold-acclima-tion-specific transcripts have been identified in various plant species including Arabidopsis thaliana (Kurkela and Franck, 1990), Brassica napus (Orr et al., 1992), Hordeum vulgare (Cativelli and Bartels, 1990), Medicago sativa (Wolfraim and Dhindsa, 1993), Oryza sativa (Aguan et al., 1991), Spinacia oleracea (Neven et al., 1993), and Triticum aestivum (Chauvin et al., 1993). Although func-tions of these proteins and the molecular mechanisms regu-lating expression of these genes remain largely unknown, they are thought to be closely related to cold acclimation.
1Corresponding author. Fax: 886-2-23919096; E-mail:
Construction and Screening of a cDNA Library A subtracted cDNA library, constructed by using the Subtractor II kit (Invitrogen, NV Leek, Netherlands) ac-cording to the manufacturers instructions, was used for differential screening. Poly(A)+ RNA from etiolated pea
seedlings cold-acclimated for 3 days was used to construct a subtracted cDNA library. Single-stranded cDNA was synthesized and hybridized to an excess of photobiotinylated poly(A)+ RNA from nonacclimated
seedlings (grown at 28°C). The resulting RNA/cDNA hybrids were complexed with streptavidin and removed from the hybridization mixture by phenol/chloroform ex-traction.
The subtracted cDNA was tailed with dGTP by termi-nal deoxynucleotidyl transferase (Boehringer Mannheim, Mannheim, Germany) and used for synthesis of second strand cDNA primed with oligo(dC). The double-stranded cDNA was ligated to EcoRI linkers, cloned into the EcoRI site of the λgt10 vector (Promega, Madison, WI, USA) using Escherichia coli as the host cells.
Differential screening was carried out as follows: Du-plicate plaque lifts of the library were prepared using Hybond-C extra filters (Amersham). Filters were then hybridized with [32P]dCTP-labeled single-stranded cDNA
generated from poly(A)+ RNA of nonacclimated plants and
plants acclimated for 3 days, separately. Recombinant λgt10 clones that hybridized preferentially to the probe prepared from acclimated plants were isolated, and the cDNA inserts were then subcloned into pGEM-11Zf(+) (Promega) for further analysis.
Northern Hybridization
Total RNA was heat-denatured in the presence of 0.3× electrophoresis buffer (1× electrophoresis buffer equals 200 mM 3-[N-morpholino]propanesulfonic acid/50 mM sodium acetate, pH 6.0/10 mM EDTA), 0.2% formalde-hyde, and 15% formamide at 65°C for 10 min and sub-jected to electrophoresis through a 1% agarose gel containing 1× electrophoresis buffer and 1.2% formalde-hyde. RNA samples were transferred onto Hybond-C ex-tra membranes, hybridized with 32P-labeled probes,
washed, and then subjected to autoradiography by the stan-dard procedures (Sambrook et al., 1989). To prepare the probe used for Northern analyses, EcoRI fragment of cDNA was gel-purified and labeled with [32P]-dCTP by
using Prime-a-Gene system (Promega). DNA Sequencing and Analysis
DNA sequence was determined by the dideoxy method of Sanger et al. (1977). Sequence analysis and alignment were performed with DNASTAR software (DNASTAR, Madison, WI, USA) or the BESTFIT program.
Isolation and Southern Analysis of Genomic DNA Two grams fresh weight of frozen pea seedlings were ground to a powder in the presence of liquid nitrogen and dispersed in 8 ml of extraction buffer (50 mM Tris-HCl
[pH 8.0]/50 mM NaCl/20 mM EDTA/100 µg/ml protein-ase K). After adding N-lauroyl sarcosine to a final con-centration of 1% (w/v), the homogenate was incubated for 1 to 2 h at 55°C. The mixture was then extracted with an equal volume of water-saturated phenol, and twice more with an equal volume of phenol:chloroform:isoamyl al-cohol (25:24:1). Total DNA was precipitated with a 0.7 volume of isopropanol. The recovered pellet was washed with 70% ethanol and resuspended in water. RNA was removed by adding ribonuclease to a final concentration of 10 µg/ml and incubating at 37°C for 1 h. Proteins were removed by adding 50 mM NaCl and 100 µg/ml protein-ase K and incubating for another 2 h at 50°C. The solu-tion was extracted with phenol:chloroform:isoamyl alcohol, then precipitated with ethanol. The pellet was resuspended in water. DNA samples were digested with appropriate restriction endonucleases prior to agarose gel electrophoresis. Southern blot was carried out and ana-lyzed according to standard procedures (Sambrook et al., 1989).
Results
Isolation and Expression of the peaci11.8 Gene cDNAs corresponding to cold-inducible mRNAs were isolated by differential screening and confirmed by north-ern blot analysis. One clone, peaci11.8, was isolated from 1.4 × 105 plaques of the subtracted cDNA library. By
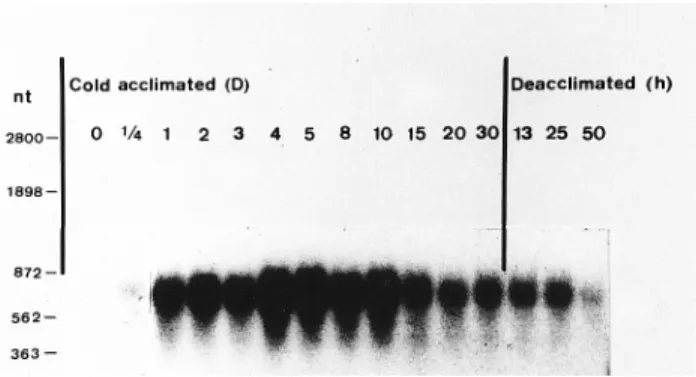
us-ing randomly primed insert of peaci11.8 as a probe, north-ern analysis including nonacclimated and cold-acclimated samples identified a single transcript of approximately 750 nucleotides in the RNA from acclimated seedlings. Fig-ure 1 shows steady-state expression of peaci11.8 mRNA during cold acclimation and deacclimation. No hybrid-ization in RNA prepared from nonacclimated seedlings was detectable (0 acclimated). The mRNA appeared dur-ing the first 6 h of cold treatment and peaked between 4 and 10 days. Subsequently, the accumulation of the tran-scripts gradually decreased and remained steady for at least
Figure 1. Cold acclimation induced the expression of peaci11.8 gene in pea. Forty micrograms of total RNA prepared from seed-lings grown at ambient temperature (28°C) and seedseed-lings treated as indicated was subjected to northern blot analysis. Deacclimation was executed after the seedlings were cold-ac-climated for 4 days. Molecular standards are indicated in nucle-otides at the left.
Figure 3. A, Nucleotides and deduced amino acid sequences of cloned peaci11.8 cDNA. The nucleotides and amino acids are numbered at the ends of lines. The direct repeats of the dodenucleotide are dashed underlined. The glycine-histidine-rich clusters are shown in bold. The NTS-like sequence is singly underlined, and the serine cluster is doubly underlined. The stop codon is indicated by asterisks, and the cis elements in the polyadenylation signal are shown in bold italics. B, Hydropathy profile of PEACI11.8 protein generated according to the procedure of Kyte and Doolittle (1982) using a 6-amino acid averaging window. The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide sequence Databases under the accession number U24398.
30 days. When 4-day-acclimated seedlings were trans-ferred back to ambient temperature, the level of transcripts declined but remained detectable for at least 50 h.
Induction of peaci11.8 responding to various stresses was also investigated. Figure 2 reveals that in addition to cold, the expression of peaci11.8 was also induced by wa-ter stress and exogenous ABA. When the seedlings were treated with 20% PEG 6,000, the accumulation of peaci11.8 transcript was identical to that of cold treatment. The gene expression was induced by exogenous ABA, but to a lower level when compared with cold treatment. About one-tenth of the transcripts induced by cold were detected when seedlings were incubated with 100 µM ABA at ambient temperature.
Sequence Analysis of the peaci11.8 cDNA
The DNA sequence was determined to be 607 bp, not including the poly(A) tail. Sequence analyses of peaci11.8
Figure 2. Effect of various stress treatments on the accumula-tion of the peaci11.8 mRNA. Forty micrograms of total RNA was subjected to northern blot analyses. Lane 1, seedlings grown at ambient temperature; Lane 2, seedlings cold-acclimated for 1 day; Lanes 3 and 4, seedlings treated with 100 µM ABA or 20% PEG 6000, respectively, at ambient temperature for 1 day.
revealed a putative open reading frame (ORF). The nucle-otide sequence and the deduced amino acid sequence are shown in Figure 3A. The 5' untranslated region contained two direct repeats of 5'-AATTCCGGAATT-3'. Polyadenylation signals, including a near-upstream ele-ment 5'-AATTATAA-3', located 17 bp upstream from the poly(A) tail, and two copies of far-upstream element 5'-TTGTA-3', were identified. The ORF, starting at nucle-otide 36 and ending with the stop codon at nuclenucle-otide 368, encoded a 110-residue polypeptide with predicted molecu-lar mass of 11.8 kDa and isoelectric point of 6.9. This deduced polypeptide, designated PEACI11.8, was rich in glycine (22.7%) and contained a high percentage of resi-dues such as histidine (15.4%), lysine (13.6%), and gluta-mate (11.8%). These four kinds of residues accounted for 63.6% of the polypeptide. On the other hand, cysteine, threonine, and tryptophan were absent from the polypep-tide. Possible motif survey of the PEACI11.8 sequence revealed two glycine-histidine-rich clusters in the middle, a serine cluster at the C-terminal end, and a nuclear
tar-geting signal (NTS)-like sequence (Cai et al., 1995; Monroy et al., 1993) at the C-terminal region. The puta-tive NTS sequence contained a bipartite motif, which started at amino acid 72, KVKHKKKKKERKKR, with nine spacer residues. The hydrophathy profile sug-gested it was hydrophilic (Figure 3B).
Search of the sequence data base indicated that PEACI11.8 showed significant homology with CAS15B (69.1%) from M. sativa (Monroy et al., 1993), WSI724 (42.7%) from O. sativa (Takahashi et al., 1994), and COR19 (44.1%) and COR11 (45.2%) from Poncirus trifoliata (Cai et al., 1995), all of which were responsive to low temperature induction. The alignment of the de-rived amino acid sequences of these genes is shown in Fig-ure 4. The regions of highest homology were located at both termini. Among these, the N-terminal 20 residues of these five polypeptides were almost identical. Addi-tionally, they all possessed NTS-like sequence and serine clusters at the C-terminal ends.
Figure 4. Comparison of the deduced amino acid sequences of PEACI11.8, CAS15B (accession number: L12461), COR19 (L39004), COR11 (L39005), and WSI724 (D26538). The amino acids are numbered at the ends of lines. Dashes present insertions in the deduced polypeptide sequences to increase sequence alignment. Identical amino acids are indicated below the sequences. Marks below are for amino acids identical in three (.) and four (:) proteins compared, respectively. Other symbols are indicated as in Figure 3.
Genomic Characterization of peaci11.8 Gene Southern blot analysis of P. sativum genomic DNA pro-vided insight into the organization of the gene. Genomic DNA was purified, digested with restriction endonucle-ases and probed with peaci11.8. The hybridization pat-tern is shown in Figure 5; only a single band was observed in the genomic DNA digested with the restriction enzymes used. From these results and the fact that the peaci11.8 transcript was polyadenylated, it was suggested that peaci11.8 exists as a single nuclear gene.
gested that water stress may also cause the cold tolerance response in pea. Although treatment with exogenous ABA induced freezing tolerance and the expression of cold-regulated genes in Arabidopsis thaliana (Chandler and Robertson, 1994), only a low level of peaci11.8 transcript was induced in pea seedlings treated with ABA at ambi-ent temperature when compared with cold treatmambi-ent. Therefore, the expression of the peaci11.8 gene in pea seems to be regulated in a different manner. This obser-vation is consistent with the hypothesis that independent but converging signal pathways may regulate the expres-sion of genes in response to low temperatures and exog-enous ABA treatment (Nordin et al., 1991).
Polyadenylation signals of plant genes are composed of several cis elements: far-upstream element, near-up-stream element, the actual polyadenylation site and/or downstream element (Hunt, 1994). Two copies of 5'-TTGTA-3', occurring in the far-upstream element in the pea rbcS-E9 gene (Mogen et al., 1992), were found between 40 and 107 upstream from the polyadenylation site in peaci11.8 cDNA. An AATAAA-like motif, 5'-AATTATAA-3', in the near-upstream element in peaci11.8 gene was proposed.
According to DNA sequence analysis, this gene could potentially encode a polypeptide, PEACI11.8. This ORF appeared to encode a small, hydrophilic protein with abun-dant glycine, histidine, lysine, and glutamate residues. Examination of the deduced amino acid sequence revealed three structural features. First, the arrangement of the ba-sic amino acids in the putative bipartite motif near the carboxy terminus of PEACI11.8 was highly homologous with those of NTS sequences (Cai et al., 1995; Monroy et al., 1993). The spacer between the two basic regions of the NTS is known to be variable in length and sequence.
Figure 6. Comparison of lysine-rich motif between PEACI11.8 and a selected group 2 LEA-relative proteins: COR47 from A. thaliana (Gilmour et al., 1992), D-11 from Gossypium hirsutum (Baker et al., 1988), B-17 from H. vulgare (Close et al., 1989), LE4 from Lycopersicon esculentum (Kahn et al., 1993), RAB21 from O. sativa (Mundy and Chua, 1988), COR19 and COR11 from Poncirus trifoliata (Cai et al., 1995) and CAP85 from S. oleracea (Neven et al., 1993).
Discussion
A cDNA clone, peaci11.8, corresponding to a cold-in-duced transcript was isolated from P. sativum by differen-tial screening of a subtracted cDNA library. The results of northern analyses clearly showed a relationship between expression of the peaci11.8 gene and cold treatment. Ac-cumulation of the corresponding transcript was observed within the first 6 h of cold treatment and remained at a high level for 10 days. When seedlings were returned back to ambient temperature after 4 days of cold treatment, the amount of the mRNA diminished rapidly. These obser-vations indicated that pea responds to cold treatment quickly and the modulation of the peaci11.8 gene is strin-gently regulated at low temperature. The turnover of peaci11.8 mRNA was slow during deacclimation. The level of this mRNA remained detectable after deacclimation for up to 50 h.
In addition to cold, peaci11.8 was responsive to water stress and exogenous ABA. It has been reported (Amundson et al., 1993) that cold tolerance could be contributed to by moderate water stress. Their results
sug-Figure 5. Southern blot analysis of P. sativum genomic DNA. Lanes B, E, H, S, and X, repesent genomic DNA digested with BamHI, EcoRI, HindIII, SalI, and XbaI, respectively. Twenty micrograms of DNA was used in each lane. Molecular standards are indicated in nucleotides at the left.
It is highly likely that the protein product is transported to nuclei. The second feature was the glycine-histidine-rich sequence tracts. Such a structure has not yet been observed in other cold-induced proteins. The third feature was the serine cluster located at the C-terminus. This is a com-mon feature of the five homologous cold-induced proteins represented in Figure 3 and is common in many LEA-like proteins, but in a different position. The glycine-histidine cluster was demonstrated in one of the fsh proteins (Haynes et al., 1989) and the pipsqueak protein (Weber et al., 1995) of Drosophila; the serine cluster motif was also found in the fsh protein. Both Drosophila proteins are critical in regulating the expression of other genes during develop-ment. Whether or not these characters in PEACI11.8 play the same regulatory function as pipsqueak and fsh proteins requires further investigation. Moreover, a lysine-rich motif, well known in group 2 LEA proteins, was found. A comparison of the lysine-rich motifs between peaci11.8 and selected LEA-relative proteins is shown in Figure 6. These sequence characterizations together with the expres-sion pattern of peaci11.8 and the hydrophilicity of PEACI11.8 suggest that it could be one of the LEA pro-teins.
Although it remains unclear how the expression of peaci11.8 contributes to the development of freezing tol-erance during the cold acclimation of pea, it is possible for its product to play an important function in plant me-tabolism during cold acclimation. The availability of peaci11.8 cDNA clone should facilitate further investiga-tion into mechanisms of cold adaptainvestiga-tion and modulainvestiga-tion of gene expression of plants at low temperatures.
Acknowledgments. The authors are grateful to Dr. Charles Y. Tsai and Dr. Linda P.-F. Chang for critical reading of the manu-script. This research was supported, in part, by National Sci-ence Council, R.O.C., Grants NSC 82-0211-B-002-318, NSC 83-0211-B-002-292, NSC 84-2311-B-002-011-B01, and NSC 85-2311-B-002-007-B13 to Y.-M. Chen.
Literature Cited
Aguan, K., K. Sugawara, N. Suzuki, and T. Kusano. 1991. Iso-lation of genes for low-temperature-induced proteins in rice by a simple subtractive method. Plant Cell Physiol. 32: 12851298.
Amundson, R.G., R.J. Kohut, J.A. Laurence, S. Fellows, and L.J. Colavito. 1993. Moderate water stress alters carbohy-drate content and cold tolerance of red spruce foliage. Environ. Exp. Bot. 33: 383390.
Baker, J., C. Steel, and L. Dure III. 1988. Sequence and charac-terization of 6 Lea proteins and their genes from cotton. Plant Mol. Biol. 11: 277291.
Cai, Q., G.A. Moore, and C.L. Guy. 1995. An unusual group 2 LEA gene family in citrus responsive to low temperature. Plant Mol. Biol. 29: 1123.
Cativelli, L. and D. Bartels. 1990. Molecular cloning and char-acterization of cold-regulated genes in barley. Plant Physiol. 93: 15041510.
Chandler, P.M. and M. Robertson. 1994. Gene expression
regu-lated by abscisic acid and its rrelation to stress tolerance. Annu. Rev. Plant Physiol. 45: 113141.
Chauvin, L.-P., M. Houde, and F. Sarhan. 1993. A leaf-specific gene stimulated by light during wheat acclimation to low temperature. Plant Mol. Biol. 23: 255265.
Close, T.J., A.A. Kortt, and P.M. Chandler. 1989. A cDNA-based comparison of dehydration-induced proteins (dehydrins) in barley and corn. Plant Mol. Biol. 13: 95108.
Gilmour, S.J., N.N. Artus, and M.F. Thomashow. 1992. cDNA sequence analysis and expression of two cold-regulated genes of Arabidopsis thaliana. Plant Mol. Biol. 18: 1321. Gilmour, S.J., R.K. Hajela, and M.F. Thomashow. 1988. Cold acclimation in Arabidopsis thaliana. Plant Physiol. 87: 745750.
Guy, C.L. 1990. Cold acclimation and freezing stress tolerance: Role of protein metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41: 187223.
Guy, C.L., K.J. Niemi, and R. Brambl. 1985. Altered gene ex-pression during cold acclimation of spinach. Proc. Natl. Acad. Sci. USA 82: 36733677.
Haynes, S.R., B.A. Mozer, N. Bhatia-Dey, and I.B. Dawid. 1989. The Drosophila fsh locus, a maternal effect homeotic gene, encodes apparent membrane proteins. Dev. Biol. 134: 246257.
Hunt, A.G. 1994. Messenger RNA 3' end formation in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 45: 4760. Kahn, T.L., S.E. Fender, E.A. Bray, and M.A. OConnell. 1993.
Characterization of expression of drought- and abscisic acid-regulated tomato genes in the drought-resistant species Lycopersicon pennellii. Plant Physiol. 103: 597605. Kurkela, S. and M. Franck. 1990. Cloning and characterization
of a cold- and ABA-inducible Arabidopsis gene. Plant Mol. Biol. 15: 137144.
Kyte, J. and R.F. Doolittle. 1982. A simple method for display-ing the hydropathic character of a protein. J. Mol. Biol. 157: 105132.
Levitt, J. 1980. Responses of plants to environmental stresses. I. Chilling, freezing and high temperature stresses. 2nd edn. Academic Press, Orlando, FL., pp. 2364.
Ma, S.F., C.Y. Lin, and Y.M. Chen. 1990. Comparative studies of chilling stress on alterations of chloroplast ultrastructure and protein synthesis in the leaves of chilling-sensitive (mung bean) and -insensitive (pea) seedlings. Bot. Bull. Acad. Sin. 31: 263272.
Monroy, A.F., Y. Castonguay, S. Laberge, F. Sarhan, L.P. Vezina, and R.S. Dhindsa. 1993. A new cold-induced alfalfa gene is associated with enhanced hardening at subzero tempera-ture. Plant Physiol. 102: 873879.
Mogen, B.D., M.H. MacDonald, G. Leggewie, and A.G. Hunt. 1992. Several distinct types of sequence elements are re-quired for efficient mRNA 3' end formation in a pea rbcS gene. Mol. Cell. Biol. 12: 54065414.
Mundy, J. and N.H. Chua. 1988. Abscisic acid and water stress induce the expression of a novel rice gene. EMBO J. 7: 22792286.
Neven, L., D.W. Haskell, A. Hofig, Q.-B. Li, and C.L. Guy. 1993. Characterization of a spinach gene responsive to low temperature and water stress. Plant Mol. Biol. 21: 291305. Nordin, K., P. Heino, and E.T. Palva. 1991. Separate signal path-ways regulate the expression of a low-temperature-induced
gene in Arabidopsis thaliana (L.) Heynh. Plant Mol. Biol. 16: 10611071.
Orr, W., B. Iu, T.C. White, L.S. Robert, and J. Singh. 1992. Complementary DNA sequence of a low temperature-in-duced Brassica napus gene with homology to the Arabidopsis thaliana kin1 gene. Plant Physiol. 98: 1532 1534.
Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: A Laboratory Manual. 2nd edn. Cold Spring Har-bor LaHar-boratory Press, Cold Spring HarHar-bor, N.Y. ISBN 0-87969-309-6.
Sanger, F., S. Nicklen, and A.R. Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA 74: 54635467.
Takahashi, R., N. Joshee, and Y. Kitagawa. 1994. Induction of chilling resistance by water stress, and cDNA sequence
analysis and expression of water stress-regulated genes in rice. Plant Mol. Biol. 26: 339352.
Thomashow, M.F. 1990. Molecular genetics of cold acclimation in higher plants. Adv. Genet. 28: 99131.
Weber, U., V. Siegel, and M. Mlodzik. 1995. Pipsqueak encodes a novel nuclear protein required downstream of seven-up for the development of photoreceptors R3 and R4. EMBO J. 14: 62476257.
Wolfraim, L.A. and R.S. Dhindsa. 1993. Cloning and sequenc-ing of the cDNA for cas17, a cold acclimation-specific gene of alfalfa. Plant Physiol. 103: 667668.
Zurfluh, L.L. and T.J. Guilfoyle. 1982. Auxin-induced changes in the population of translatable messenger RNA in elon-gating sections of soybean hypocotyl. Plant Physiol. 69: 332337.
‰ ¤§¤ §C•¯»⁄
ƒ]⁄§flS'˚⁄ “R
¯˙‹F›ı¡@‚›¶}•¯¡@“L‹ ”a¡@‡flflq'œ
Œ¥ »O˘W⁄j ˙· “« ˙¤t
⁄@›‰¤§ (Pisum sativum L.) ¤ §C•¯»⁄ ⁄§ ƒ] peaci11.8¡A ‚g¥ „ §R¿ ƒ]fiw¶iƒ fit†§¿z¿ ƒ ‹D¿
–o¤ ¡C‚ ƒ]ƒb‰ ¤§¥fi›]¤ ¤ 4¢J §C•¯‡B†z 6 ⁄pfi ⁄”«K¶}'l“ †{¡A¤ ´ ¿ †£“««h' §C•¯‡B†z 4 ƒ 10 ⁄ † ¿nƒ ‡ “¡Aƒ ¥fi›]ƒbƒ^•¯⁄§«Æ¡A‚ ƒ]⁄§“ †{« § §Y –⁄ ¡A¯ª¥ ¤ “ †{'M§C•¯ f„ ƒ‡‹ • ” –K“”ˆ
`p¡Cƒ„¥~¡A⁄ ¶ f„ ¥ ¥i»⁄ ‚ ƒ]« ““”“ †{¶q¡Cƒ peaci11.8 fi – »˜§˙ƒC– ⁄§ i »˜§˙ƒC»P⁄@¤˙ · “«¤ §C•¯‰ ––⁄§ ƒ]¤ªƒ‡•¥ ““”ƒP•‰'˚¡C¤ ⁄⁄¥]‹Aƒb›F›L o¥˝– ·`·I§t (late embryogenesis abundant¡ALEA) ‡J¥ ‰Ł–‘¤£“” “¿ fi »˜ (lysine-rich) ˇ‹q⁄˛ •fi »˜ƒŒ´O (serine cluster) ⁄G” flS…x¡C
peaci11.8⁄§ o†{–Nƒ‡§U' · “«§C•¯»⁄ ‡J¥ ‰Ł»P§C•¯§ '˚⁄§¥˝†z¥\fl ‹ª¤s¡C