中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/50313
51
0
0
全文
(2) . . II .
(3) . Catalog Catalog. -------------------------------------------------------------------------------------III. List of Abbreviations. --------------------------------------------------------------------V. Chinese abstract --------------------------------------------------------------------------VII English abstract. --------------------------------------------------------------------------VIII. Text Chapter One: Introduction. ----------------------------------------------------------------1. Section One: Background. ------------------------------------------------------3. 1.1.1 Nicardipine 1.1.2 Microglial cells 1.1.3 Activated microglia and neuroinflammation in CNS 1.1.4 MAPK/Akt pathway Section Two: Purpose ------------------------------------------------------------8 Chapter Two: Material and Methods Section One: Materials. --------------------------------------------------------14. 2.1.1 Cell line 2.1.2 Cell culture 2.1.3 Migration assay 2.1.4 Reverse Transcriptase-PCR 2.1.5 Western blot 2.1.6 Antibody 2.1.7 Inhibitor Section Two: . Methods. ------------------------------------------------------18 III .
(4) . 2.2.1 Reagents and antibodies 2.2.2 Cell culture 2.2.3 Animals 2.2.4 Lipopolysaccharide (LPS) injection and tissue preparation 2.2.5 Immunohistochemistry 2.2.6 Western blot 2.2.7 Migration assay 2.2.8 Reverse transcription-PCR and quantitative real-time PCR 2.2.9 Statistical analysis Chapter Three: Results. ------------------------------------------------------------------23. 3.1. Effects of nicardipine on BV-2 microglia viability 3.2. Effects of nicardipine on ATP-cellular migration in microglial cells 3.3. Effects of nicardipine on LPS/IFN-γ-induced or peptidoglycan-induced NO production in microglial cells 3.4. Effect of nicardipine on LPS/IFN-γ-induced iNOS and COX-2 expression in BV-2 microglia 3.5. Effect of nicardipine on LPS/IFN-γ-induced IL-6 and IL-1β production and expression in BV-2 microglia 3.6. Nicardipine inhibits LPS/IFN-γ-stimulated phosphorylation of MAP kinases and Akt in BV-2 microglia 3.7. Nicardipine inhibits activation of microglia induced by LPS in mice Chapter Four: Discussion and conclusion. --------------------------------------------28. Figures ------------------------------------------------------------------------------------------30 Reference. . -------------------------------------------------------------------------------------39. IV .
(5) . List of Abbreviations Aβ: β-amyloid APS: Ammonium persulfate CCB: Calcium channel blocker CNS: Central nervous system COX-2: Cyclooxygenase-2 DAMPs: Danger associated molecular patterns ddH2O: double distilled water DMEM: Dulbecco's modified eagle's medium FBS: Fetal bovine serum Iba-1: Ionized calcium binding adaptor molecule-1 ICE: Interleukin-1 converting enzyme ICH: Intracerebral hemorrhage IFN: Interferon IL-1β: Interleukin-1β IL-6: Interleukin-6 iNOS: Induced nitric oxide synthase. . V .
(6) . IP: Intraperitoneal LPS: Lipopolysaccharide MAPK: Mitogen-activated protein kinase MHC: Major histocompatibility complex NO: Nitric oxide PBS: Phosphate buffered saline PGE2: Prostaglandin E2 PKB: Protein kinase B P/S: Penicillin/Streptomycin ROS: Reactive oxygen species TLRs: Toll-like receptors TNF-α: Tumor necrosis factor-α. . VI .
(7) . Chinese Abstract Nicardipine 是一種鈣離子阻斷劑,有很好的降血壓的效果。它常被用來 治療因為缺血性中風、外傷性腦損傷及出血性腦中風後引起的危害性高血壓。而 且因為降血壓效果穩定且快速,又沒有其他心肺方面的抑制效果,所以逐漸被廣 泛且常規的使用在神經加護照顧。但是這個藥物在神經細胞的作用機制目前仍不 清楚。我們利用老鼠的微膠細胞 (microglia) 培養實驗,發現 nicardipine 有明顯 的抑制神經細胞發炎反應的效果。而且在動物實驗上也發現 nicardipine 能抑制微 膠細胞 (microglia) 的活化達到神經保護的效果,我們同時也發現 nicardipine 會 抑制微膠細胞因 ATP 所活化的移行反應。Nicardipine 會減少經由 LPS/IFN-γ 所 刺激的微膠細胞 (microglia) 之 NO 生成量,同時也會抑制 iNOS 和 COX-2 的 表現,對於前驅發炎細胞因子 IL-6 及 IL-1β 也有抑制的效果。由進一步的實驗 顯示,nicardipine 有可能是藉由抑制 MAPK/Akt pathway 中的 Akt, p38、p65 及 cJun 之磷酸化作用,進一步抑制 NF-κB pathway 之活化來達到抗神經性發炎的 效果。. . VII .
(8) . English Abstract Nicardipine hydrochloride, a calcium channel blocker (CCB), has strong anti-hypertensive activity. It has been used to control blood pressure in severe hypertension after ischemic stroke, traumatic brain injury, and intracerebral hemorrhage (ICH) and increasingly widely applied in neurointensive care. But the definite response of nicardipine on neuron cells is unclear. Using the murine BV-2 microglia in culture, we demonstrated the nicardipine significantly inhibited LPS/IFN-γ-induced neuroinflammatory response. The neuroprotective effect of nicardipine was attributed to the inhibition of microglial activation in animal model, and significantly inhibited the production of nitric oxide (NO). We also found that nicardipine inhibited the migration of microglia cells which were activated by ATP. Nicardipine down-regulates LPS/IFN-γ induced nitric oxide synthase (iNOS) and cyclo-oxygenase-2 (COX-2) expression in BV-2 microglia, too. Hereafter nicardipine effectively suppressed pro-inflammtory cytokines expression such as IL-6 and IL-1β. Furthermore, nicardipine inhibited the phosphorylated of p38 and Akt in BV-2 microglia and suppressed the phosphorylated of p65 and cJun. Our data showed that nicardipine hydrochloride significantly suppress MAPK/Akt signaling pathway activation, which is involved in anti-neuroinflammation.. . VIII .
(9) . Chapter One: Introduction Anti-hypertensive agents have been used to the patients who were suffered from ischemic stroke, intracerebral hemorrhage, and traumatic brain injury routinely because of uncontrolled hypertensive crisis will be developed frequently (Andrews et al., 2012; Gianino and Afuwape, 2012; Gross et al., 2012). There are several antihypertensive agents, which were suggested including nitroprusside, β-blocker, and calcium-channel-blockers. Nicardipine hydrochloride, a calcium channel blocker, has been used to treat the severe hypertension increasingly because of it is feasible and safe (Qureshi et al., 2006). Moreover, nicardipine has less effect of cardiopulmonary suppression than β-blockers. In some recent studies, nicardipine was used in treatment of acute hypertension with acute brain disease carried better outcome (Kim et al., 2012; Narotam et al., 2008; Qureshi et al., 2012; Qureshi and Palesch, 2011; Sato et al., 2012). But the cell response of nicardipine on neurons is not clear until now. Acute brain disease that is defined as ischemic stroke, intracerebral hemorrhage and traumatic brain injury (Narotam et al., 2008) often result in the poor prognosis and the secondary insult play the important role in poor outcome. The neuroinflammatory response after neurons damage is the leading cause of secondary insult (Aronowski and Zhao, 2011). Microglia are the main form of active defense system and tissue repair in central nervous system. In pathologic conditions, activated microglia have been implicated as the predominant type and been in charge of inflammation-mediated neuronal damage (Aloisi, 1999). Upon chronic activation, microglia are capable of producing a variety of pro-inflammatory mediators and . 1 .
(10) . potentially neurotoxic agents. Overactivation of microglial cells may cause severe brain tissue damage in many neurologic diseases. Glial activation involves changes in cell phenotype and the expression of special proteins, such as iNOS and COX-2. The mechanism by which activated glia induce neuronal death has been proved to involve NO (Brown, 2001) , reactive oxygen species (ROS) and pro-inflammatory cytokines (Beckman et al., 1994). LPS and IFN-γ potently increases the levels of cellular MAP kinase phosphorylation, thus enhancing pro-inflammatory responses in microglia. In this designed study, we address the questions whether nicardipine downregulates inflammatory responses and how nicardipine exhibits anti-inflammatory responses in microglia.. . 2 .
(11) . Section One: Background 1.1.1 Nicardipine hydrochloride Nicardipine hydrochloride (Cardene®) is a dihydropyridine type calcium channel-blocking agent used for treatment of vascular disorders like stable angina and hypertension. It is available in oral and intravenous formulations. The mechanism and clinical effects of nicardipine closely resemble those of nifedipine and the other dihydropyridines, except that nicardipine is more selective for cerebral and coronary blood vessels. Furthermore, nicardipine does not intrinsically decrease myocardial contractility unlike the other anti-hypertensive agent of β-blocker. Nicardipine also has a longer half-life than nifedipine and has greatest vasodilating effect between all calcium channel blockers, especially in coronary artery. So Nicardipine has been used to control blood pressure in severe hypertension after acute brain injury, which is including of ischemic stroke, traumatic brain injury, ICH, and subarachnoid hemorrhage (Haley et al., 1994; Narotam et al., 2008; Powers et al., 2001). Nicardipine inhibits the transmembrane influx of calcium ions into cardiac muscle and smooth muscle without changing serum calcium concentrations. The effects of nicardipine are more selective to vascular smooth muscle than cardiac muscle. The structure is showed as the Figure 4A. Calcium channel blockers bind to receptors on voltage-gated Ca2+ channels including L, N, T subtypes. All of these subtypes keep these channels in an inactive state and decreased intracellular Ca2+ ion. All clinically useful calcium channel blockers could bind to alpha1 subunit of L-type Ca2+ channel and decreased Ca2+ influx into cells. . 3 .
(12) . Although nicardipine has been administrated routinely in patients of acute brain injury, but there is few literature to discuss the biochemical response in neurons. Some authors mentioned that nicardipine shows the different regulatory reaction on glia to calcium-channel blocker (Koizumi et al., 2002; Komagiri et al., 2011). In clinical practice, Nicardipine was used in treatment of acute hypertension with acute brain disease carried better outcome (Kim et al., 2012; Narotam et al., 2008; Qureshi et al., 2012; Qureshi and Palesch, 2011; Sato et al., 2012). Interestingly, Amenta et al. proposed the neuroprotective effect of nicardipine in animal model and in vitro study (Amenta et al., 1996; Amenta and Tomassoni, 2004). The definite mechanism of nicardipine acting in CNS is not understood.. 1.1.2 Microglial cells Microglia are a type of glial cell that are the resident macrophages of the central nervous system (CNS) and act as the first and main form of active immune defense. Microglia constitute 20% of the total glial cell population within the brain. Under the normal condition, microglia serve a critical role of immune surveillance and provide immune protection against CNS infection. Since this process must be done quickly to prevent potentially fatal damage, microglia are extremely sensitive to even small pathological changes in the CNS (Dissing-Olesen et al., 2007). However, in pathologic conditions, microglia become activated and undergo impressive morphologic changes, metamorphosing form resting ramified microglia into activated amoeboid microglia (Aloisi, 1999). The expression of many surface markers, such as complement receptors and major histocompatibility complex (MHC) . 4 .
(13) . molecules increases in activated microglia (Ng and Ling, 1997) . Microglial cells express receptors typically described for brain-specific communication such as neurotransmitter receptors and those first discovered as immune cell-specific such as for cytokines. Microglial cells are the most susceptible sensors of brain pathology. Upon any detection of signs for brain lesions or neurons dysfunction, microglial cells undergo a complex and multiple stages activation process that converts them into the activated microglial cells (Figure 1). This cell form has the capacity to release a large number of substances that can act detrimental or beneficial for the surrounding cells. Activated microglial cells can migrate to the site of injury, proliferate, and phagocytic cells and cellular compartments (Kettenmann et al., 2011).. 1.1.3 Activated microglial cells and neuroinflammation The majority of factors produced by activated microglia, including tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), arachidonic acid metabolities prostaglandin E2 (PGE2), reactive oxygen species (ROS) and nitric oxide (NO), are all pro-inflammatory and neurotoic molecules (Lee et al., 1993; Liu et al., 2000; Liu and Hong, 2003). The overproduction and accumulation of these pro-inflammatory factors are toxic to neighboring neurons, which further activated microglia and damage the remaining neurons, thus set in motion a self-propelling progressive cycle of inflammation and neuronal damage (Figure 2) (Block and Hong, 2005). Therefore, inhibition of microglial activation and their production of neurotoxic factors would be a potential therapeutic approach to stop or decrease the progression of acute brain . 5 .
(14) . injury. S. aureus is one of the main pathogens of CNS parenchymal infection leading to the establishment of brain abscess (Kielian, 2004). However, the mechanisms by which S. aureus influences immune activation in the CNS remain to be fully defined. Previous studies from our laboratory have shown that both S. aureus and PGN enhance microglial PRR expression, which may serve to potentiate the antibacterial immune response in infected brain parenchyma prior to leukocyte infiltration (Kielian et al., 2002,2005). In addition, upon exposure to S. aureus and PGN, microglia respond with the robust synthesis and release of numerous proinflammatory mediators as well as internalizing and killing S. aureus in vitro (Kielian et al., 2002, 2005). Although previous studies have demonstrated that PGN binds to CD14 and enhances TLR2-mediated signaling in macrophages (Iwaki et al., 2002; Koedel et al., 2003; Schwandner et al., 1999), the fundamental role of CD14 in S. aureus- and PGN-dependent microglial activation is not known. In response to injury or inflammatory stimuli, the resting microglia can be rapidly activated to participate in pathological responses, including migration to the affected site, release of various inflammatory molecules, and clearing of cellular debris. ATP released from damaged neural cells has been considered as the major chemokine for inducing rapid process extension and cell body migration of microglial cells. However, ATP leaking from the local injured cells may not be able to diffuse extensively to effectively induce migration of remote microglia, because extracellular nucleotides are rapidly degraded by ecto-ATPases in the brain. It seems likely that ATP released from injured neural tissue may evoke a regenerative ATP-induced ATP . 6 .
(15) . release from surrounding cells to establish the long-range extracellular ATP gradient required for the chemotaxis of remote microglia.. 1.1.4 MAP kinase pathway and Akt pathway Mitogen-activated protein kinases (MAPKs) are a family of serine/threonine protein kinases that mediate important biological processes and cellular responses to external stress signals. MAP kinase has been shown to play a critical role in the regulation of energy metabolism under both physiological and pathological conditions. MAP kinases are stimulated by many inflammatory stimuli, including of LPS, IFN-γ, and TNF-α. Furthermore, MAP kinases are involved in LPS-induced iNOS and COX-2 production via control of NF-κB activation in microglial cells (Caivano, 1998). Activation of the three MAPKs (ERK1/2, p38, JNK) are linked with induction of iNOS expression in microglia (Figure3) (Moon et al., 2007). Akt pathway, also known as protein kinase B (PKB), is a serine/threoninespecific protein kinase that plays an important role in multiple cellular processes such as glucose metabolism, apoptosis, and cell proliferation, transcription and cell migration. pAkt play a predominant roles in pro-inflammatory gene expression, too.. . 7 .
(16) . Section Two: Purpose Nicardipine is an anti-hypertensive agent to apply in the patients who are suffered from crisis hypertension resulted from acute brain injury. To compare with other anti-hypertensive agents, nicardipine shows more effective on blood pressure control and has less cardiopulmonary effect. Hence, nicardipine is more popular application in clinical practice for the recent years. Acute brain injury, including ischemia stoke, spontaneous ICH and traumatic brain injury, are critical problems. In addition to surgery and removal the first pathology, the severe brain injury due to secondary insults was the major issue. A series of neuroinflammatory responses result in secondary insults and activated microglial cells play the most important roles. Some literatures mentioned nicardipine demonstrated neuro-proective effect. Except for anti-hypertensive function, the other biologic mechanism of nicardipine on CNS is not clear. We purposed the nicardipine suppress the neuroinflammation responses in microglial cells and to discuss the mechanism.. . 8 .
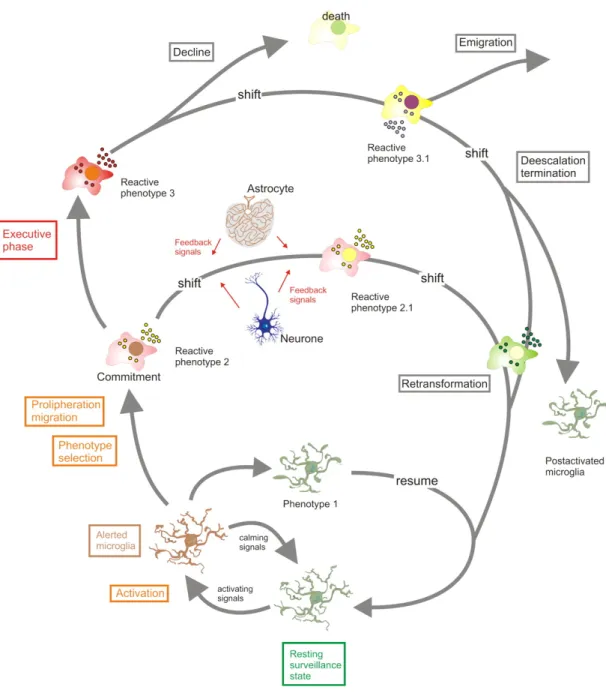
(17) . Figure 1 Microglial activity states throughout an activation process. The “resting” microglia constantly and actively scan their environment for exogenous or endogenous signals indicating a threat to the homeostasis. Sudden appearance of “activating” signals or a loss of constitutively “calming” inputs can then trigger transitions to alerted and activated states. Cells can commit to distinct reactive phenotypes depending on the challenging stimuli and the situational context. Initial response profiles may further shift as instructed by additional influences. Not only. . 9 .
(18) . resident CNS cells, but also invading immune cells would exert such modulating influences. Initial reactive phenotypes with defense orientation may convert to repair-orientated activity profiles. Cells may eventually return to a resting state or stay “experienced”. Experienced microglia could reveal altered responsiveness and exert distinct responses upon rechallenge (Hanisch and Kettenmann, 2011).. . 10 .
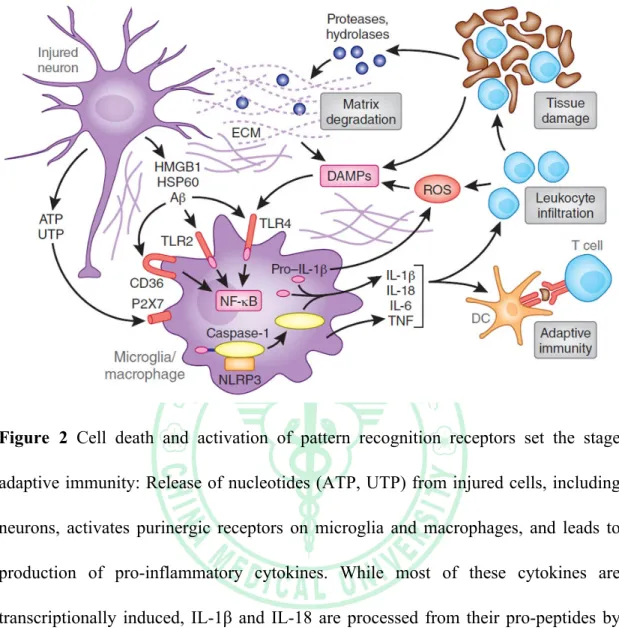
(19) . Figure 2 Cell death and activation of pattern recognition receptors set the stage adaptive immunity: Release of nucleotides (ATP, UTP) from injured cells, including neurons, activates purinergic receptors on microglia and macrophages, and leads to production of pro-inflammatory cytokines. While most of these cytokines are transcriptionally induced, IL-1β and IL-18 are processed from their pro-peptides by the activity of interleukin-1 converting enzyme (ICE; caspase 1). ICE is embedded in a multi-protein complex (NLRP3 or inflammasome) and is activated by microglial P2X7 receptors. Ischemic cell death leads to the formation of danger associated molecular patterns (DAMPs) molecules, which activate TLRs, especially TLR2 and 4. DAMPs released by ischemia include HMGB1, an intracellular DNA binding protein released after cellular injury, HSP60, and β-amyloid (Aβ) among others. TLRs in concert with scavenger receptors such as CD36, upregulate pro-inflammatory gene. . 11 .
(20) . expression through the transcription factor NF-κB. DAMPs also derive from matrix breakdown by lytic enzymes released from dead cells and by the action of reactive oxygen species (ROS) on lipids. The cytokine production and complement activation resulting from these events leads to increased leukocyte infiltration and enhances tissue damage, which, in turn, produces more DAMPs. Antigens unveiled by tissue damage are presented to T cells, setting the stage for adaptive immunity (Iadecola and Anrather, 2011).. . 12 .
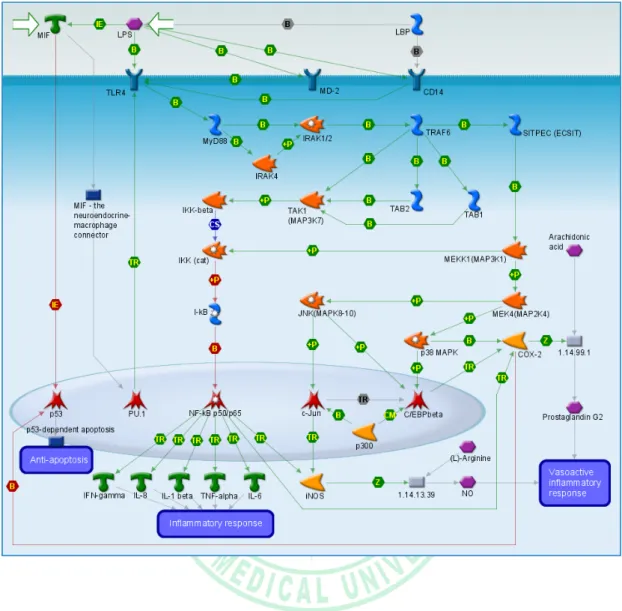
(21) . Figure 3: Proposed MAPK and NF-κβ pathways (Annual review of immunology 2003;21:335-76 ). . 13 .
(22) . Chapter Two: Materials and Methods Section One: Materials 2.1.1 Cell line BV-2 microglia (mouse, C57BL/6, brain, microglial cells). 2.1.2 Cell culture 1. Dulbecco's modified eagle's medium (Gibco® 12100-061) 2. Fetal bovine serum (Gibco® 16000-044) 3. Penicillin/Streptomycin (Gibco® 15140-122) 4. Phosphate buffered saline 5. 0.25% trypsin-EDTA (Gibco® 25200-144). 2.1.3 Migration assay 1.. Transwell (Corning® 3422). 2.. 20% methanol. 3.. 0.05% crystal violet (SIGMA C3886). 2.1.4 Reverse transcriptase-PCR . . 1.. Trizol (Invitrogen™ 15596-018). 2.. Chloroform (Merck 1.02444.1000). 14 .
(23) . 3.. Isopropanol (MARESCO,Inc). 4.. 75% alcohol. 5.. double distilled water. 6.. Oligo dT (MDBio,Inc.). 7.. dNTP (MDBio,Inc.). 8.. 10x RT buffer (MDBio,Inc.). 9.. MgCl2 (MDBio,Inc.). 10.. DTT (Invitrogen™ P2325). 11.. M-MLV enzyme (Invitrogen™ 28025-013). 2.1.5 Western blot. . 1.. Polyacrylamide (MDBio,Inc.). 2.. 1.5M Tris-HCl. 3.. 0.5M Tris-HCl. 4.. 10% SDS (MARESCO,Inc). 5.. Ammonium persulfate (MDBio,Inc.). 6.. Running buffer. 7.. Transfer buffer. 15 .
(24) . 8.. Methanol. 9.. PVDF membrane (Millipore). 10.. ECL (Millipore). 2.1.6 Antibody 1.. β-actin (Santa Cruz, CA). 2.. MMP-13 (Santa Cruz, CA). 3.. MMP-9 (Cell Signaling Technology, Inc). 4.. p-ERK (Santa Cruz, CA). 5.. ERK (Santa Cruz, CA). 6.. p-p38 (Cell Signaling Technology, Inc). 7.. p38 (Santa Cruz, CA). 8.. p-JNK (Cell Signaling Technology, Inc). 9.. JNK (Santa Cruz, CA). 10.. p-c-Jun (Santa Cruz, CA). 11.. c-Jun (Santa Cruz, CA) . 2.1.7 Inhibitor 1.. . PD98059 (ERK inhibitor, Sigma-Aldrich). 16 .
(25) . . 2.. SP600125 (JNK inhibitor, Torcis Bioscience). 3.. SB203580 (P38 inhibitor, Sigma-Aldrich). 17 .
(26) . Section Two: Methods 2.2.1 Reagents and antibodies The antibody against ionized calcium binding adaptor molecule-1 (Iba-1) was purchased from Wako Pure Chemical Industries (Osaka, Japan). Primary antibodies against β-actin, p65, c-Jun, ERK2, p38, JNK and phospho-ERK1/2 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Primary antibodies against phospho-c-Jun, phospho-p65, phospho-p38 and phospho-JNK were purchased from Cell Signaling and Neuroscience (Danvers, MA). The primary antibody against iNOS was purchased from BD Transduction Lab (Lexington, KY). The primary antibody against COX-2 was purchased from Cayman Chemicals (Ann Arbor, MI).. 2.2.2 Cell culture The murine microglial cell line BV-2 was originally generated by infecting primary microglial cell cultures with a v-raf/v-myc oncogene carrying a retrovirus (J2). Since BV-2 microglia retain most of the morphological, phenotypical, and functional properties described for freshly isolated microglial cells, they can be considered as immortalized active microglial cells (Blasi et al, 1990). Cells were cultured in dulbecco's modified eagle's medium (DMEM) with 10% fetal bovine serum (FBS) at 37°C, and passaged by trypsinization.. 2.2.3 Animals All mice were manipulated in accordance with the Animal Care and Use Guidelines of the China Medical University (Taichung, Taiwan). Eight-week-old . 18 .
(27) . male imprinting control region (ICR) mice were purchased from the National Laboratory Animal Center (Taipei, Taiwan). The animals were housed in temperature and humidity controlled environment and given access to food and water ad libitum. Mice were acclimated to their environment for 7 days before the experiments.. 2.2.4 Lipopolysaccharide (LPS) injection and tissue preparation Eight-week-old mice received intraperitoneal injections of saline or 20 mg/kg LPS (E. coli, serotype 0127:B8) for 24 h. After performing rotarod tests, mice were deeply anesthetized, transcardially perfused with 10% formaldehyde, and brains were postfixed overnight. Brain samples were stored in a 30% sucrose solution at 4°C until they sank.. 2.2.5 Immunohistochemistry Coronal serial sections (30 µm) were sliced on a freezing sliding microtome cryostat (Leica CM305S). Free-floating slices were quenched with endogenous peroxidases with hydrogen peroxide, blocked by nonspecific binding with non-fat milk, permeabilized with Triton X-100, and then incubated with primary antibody against Iba-1 overnight at 4°C. Following incubation with biotinylated secondary antibody, the sections were stained using the avidin-biotin complex (Vector Laboratories), and developed with diaminobenzidine (DAB). To determine Iba-1-positive microglia across different treatments, the cerebral cortex and hippocampus were digitally captured at 10× and 40× magnifications under a light microscope.. . 19 .
(28) . 2.2.6 Western blot Cells were lysed briefly in homogenizing buffer for 30 mins on ice. Equal amounts of the samples were loaded in each lane. The membranes were blocked with non-fat milk in phosphate-buffered saline (PBS) and then probed with primary antibodies. After undergoing three PBS washes, the membranes were incubated with secondary antibodies. The blots were visualized by enhanced chemiluminescence using FUJI Medical X-Ray Film (FUJIFILM Corporation, TOKYO).. 2.2.7 Migration assay In vitro migration and invasion assays were performed using Costar Transwell inserts (Costar, NY; pore size, 8 µm) in 24-well plates as described previously (Chen et al., 2011; Lu et al., 2010). Approximately 1 × 104 cells in 200 µl of serum-free medium were placed in the upper chamber, and 300 µl of the same medium containing ATP was placed in the lower chamber. Before performing the migration assay, cells were pre-treated for 30 mins with nicardipine followed by treatment with ATP for 24 h. The plates were incubated for 24 h at 37°C in 5% CO2, and then cells were stained with 0.05% crystal violet and 2% methanol. Non-migratory cells on the upper surface of the filters were removed by wiping with a cotton swab. The cell number in three fields per well was counted under a microscope at 100× magnification. Images of migratory cells were observed and acquired with a digital camera and light microscope.. 2.2.8 Reverse transcription-PCR and quantitative real-time PCR. . 20 .
(29) . Total RNA was extracted from primary cultured microglia or BV-2 microglia using a TRIzol kit (Invitrogen™). The reverse transcription reaction was performed using 2 µg of total RNA that was reverse transcribed into cDNA using the oligo(dT) primer. Quantitative real-time PCR using SYBR Green I Master Mix was analyzed with a model 7900 Sequence Detector System (Applied Biosystems, Foster City, CA). After preincubation at 50°C for 2 mins and 95°C for 10 mins, the PCR was performed as 40 cycles of 95°C for 10 s and 60°C for 1 mins. The threshold was set above the non-template control background and within the linear phase of target gene amplification to calculate the cycle number at which the transcript was detected (denoted as CT). The oligonucleotide primers were iNOS . 5’-CCCAGAGTTCCAGCTTCTGG-3’ 5’- CCAAGCCCCTCACCATTATCT-3’ . COX-2 . 5’-TGGGGTGATGAGCAACTATT-3' 5’-AAGGAGCTGGGTCAAACT-3’ . IL-1β . 5’-TGGGGGAGATTCTCACTTTG-3’ 5’-CCATCAGCGTTCCCATACTT-3’ . IL-6 . 5’-CCAGTTGCCTTCTTGGGACTG-3’ 5’-CAGGTCTGTTGGGAGTGGTATCC-3’ . . 21 .
(30) . TNF-α . 5’-AAAATTCGAGTGACAAGCCTGTAG-3’ 5’-CCCTTGAAGAGAACCTGGGAGTAG-3’ . GAPDH . 5’-CTCAACTACATGGTCTACATGTTCCA-3’ 5’-CTTCCCATTCTCAGCCTTGACT-3’ . 2.2.9 Statistical analysis The values given are means ± S.E.M. The significance of difference between the experimental group and control was assessed by Student’s t test. The difference was considered to be significant if the p value is less than 0.05.. . 22 .
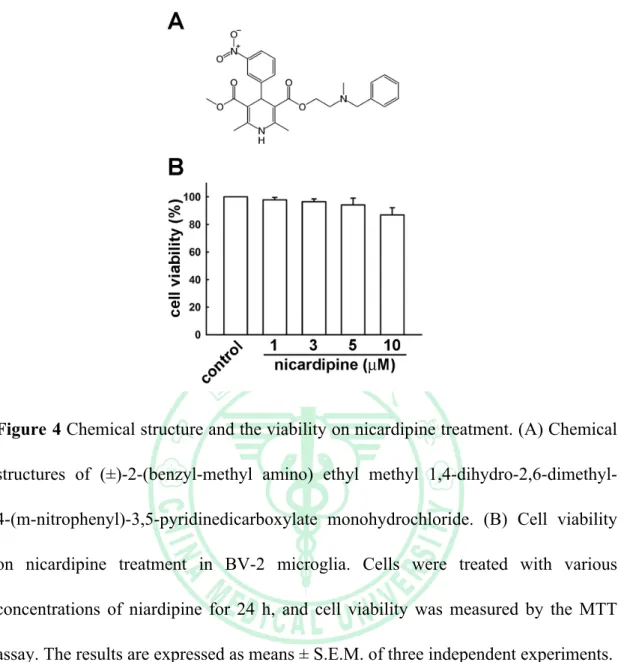
(31) . Chapter Three: Results 3.1. Effects of nicardipine on BV-2 microglia viability The MTT assay was performed at 24 h after treatment with the indicated concentrations of nicardipine on BV-2 microglia viability. Nicardipine at 1-10 µM did not have a cytotoxic effect on BV-2 microglia (Figure 4B). Therefore, a concentration of nicardipine within this range was applied in the remaining experiments.. 3.2. Effects of nicardipine on ATP-cellular migration in microglial cells In vitro migration activity study, BV-2 microglia were pre-incubated with or without nicardipine (10 µM) for 60 mins followed by incubation with ATP (100 or 300 µM) for another 24 h and the migrated cells were visualized by phase-contrast imaging. The results demonstrated that ATP enhanced the migrative activity on BV-2 microglia in a concentration-dependent manner. Furthermore, nicardipine suppressed the ATP-cellular migration on BV-2 microglia(Figure 5B). The migration fold of BV-2 microglia were decreased from 1.5 to 0.5 in the nicardipine treatment group (Figure 5A).. 3.3. Effects of nicardipine on LPS/IFN-γ-induced or peptidoglycaninduced NO production in microglial cells To evaluate the effect of nicardipine on NO production, BV-2 microglia were stimulated with LPS (10 ng/ml)/IFN-γ (10 ng/ml) or peptidoglycan (10 µg/ml) for 24 h after pre-treatment with 1, 3, 5, 10 µM of nicardipine for 60 mins. Cell supernatants . 23 .
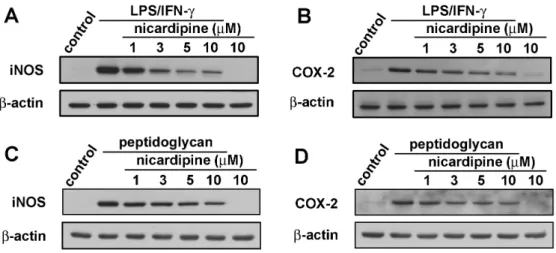
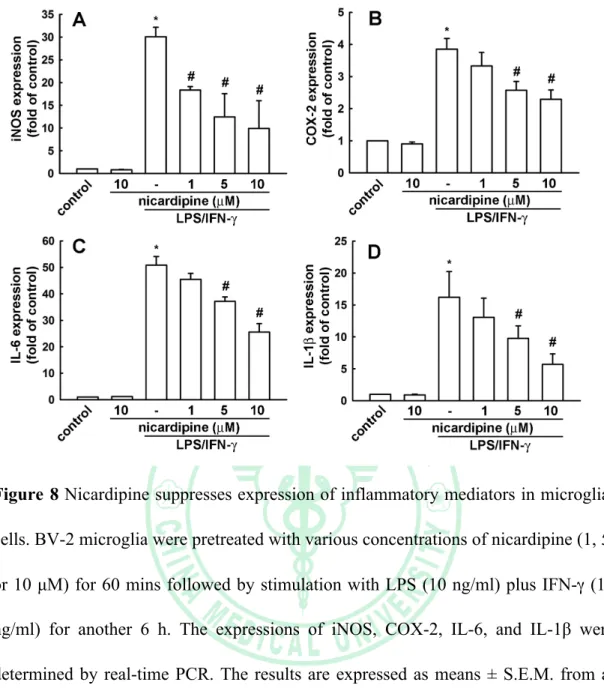
(32) . were collected and assayed for NO production using Griess reaction assay and ELISA. As shown in Figure 6A, treatment with LPS/IFN-γ resulted in marked NO production form cells as compared with that generated by the control. However, pre-treatment with nicardipine significantly repressed the levels of NO in LPS/IFN-γ-stimulated BV-2 microglia in a concentration-dependent manner. The similarity results were showed in the peptidoglycan-stimulated cells (Figure 6B).. 3.4. Effect of nicardipine on LPS/IFN-γ-induced iNOS and COX-2 expression in BV-2 microglia We performed western blot analysis and real-time PCR to detect of protein and mRNA levels to examine whether inhibiting NO production by nicardipine was associated with decreased levels of inducible NO synthase and COX-2 expression. The western blot data showed that treatment with LPS/IFN-γ significantly increased iNOS and COX-2 protein expression at 24 h; however, nicardipine markedly inhibited iNOS and COX-2 protein expression in LPS/IFN-γ-stimulated BV-2 microglia in a concentration-dependent manner (Figure 7A and B). Sequentially, treatment with PGN increased iNOS and COX-2 protein expression at 24 h; however, nicardipine markedly inhibited iNOS and COX-2 protein expression in PGN-stimulated BV-2 microglia in a concentration-dependent manner (Figure 7C and D). Consistent with the results of the western blot analysis, real-time PCR data indicated that treatment with LPS/IFN-γ significantly increased iNOS and COX-2 mRNA expression after 24 h (Figure 8A and B). However, treatment with nicardipine resulted in a significantly decrease in iNOS and COX-2 mRNA expression. Taken together, these data indicate. . 24 .
(33) . that nicardipine inhibit upregulation of LPS/IFN-γ-stimulated NO production by suppressing iNOS and COX-2 expression.. 3.5. Effect of nicardipine on LPS/IFN-γ-induced IL-6 and IL-1β production and expression in BV-2 microglia We next investigated whether nicardipine regulated the production of pro-inflammatory cytokines, such as IL-6 and IL-1β, and their mRNA levels in LPS/IFN-γ-stimulated BV-2 microglia. BV-2 microglia were pre-treated with 1, 5, 10 µM of nicardipine for 1h, before LPS (10 ng/ml) plus IFN-γ (10 ng/ml) stimulation for 24 h, real-time PCR analysis were preformed to determine the effect of nicardipine on IL-6 and IL-1β gene expression. Consistent with IL-6 and IL-1β production, the real-time PCR data showed that nicardipine suppressed IL-6 and IL-1β expression in LPS stimulated BV-2 microglia (Figure 8C and D). These data indicated that nicardipine regulate LPS-stimulated IL-6 and IL-1β release at transcriptional level.. 3.6. Nicardipine inhibits LPS/IFN-γ-stimulated phosphorylation of MAP kinases and Akt in BV-2 microglia. Because MAP kinases are the most important signaling molecules involved in regulating the synthesis and release inflammatory mediators and cytokines by activated microglia (Kim et al., 2004; Zhang and Dong, 2007), we next examined the effect of nicardipine on LPS/INF-γ-induced activation of MAP kinases, including extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK) and p38. The western blotting results indicated that LPS/IFN-γ rapidly activated MAP kinases . 25 .
(34) . within 1 h after stimulation. However, nicardipine pre-treatment significantly inhibited phosphorylation of p38 in LPS/IFN-γ-stimulated BV-2 microglia (Figure 9), suggesting that nicardipine are capable of disrupting the key signal transduction pathways activated by LPS/IFN-γ in BV-2 microglia. Activation of NF-κB is closely related to regulation of iNOS and COX-2 genes in activated BV-2 microglia. Therefore, we next investigated whether nicardipine modulate the NF-κB activation in BV-2 microglia in response to LPS/IFN-γ using western blot analysis. The Western blot data showed a marked increase in the amount of NF-κB p65 in the nucleus after exposure to LPS/IFN-γ. However, the LPS/IFN-γ-induced p65 level in the nuclear fractions decrease by nicardipine (10 µM) pre-treatment (Figure 10). Recent findings have indicated that Akt signaling molecule prompts NF-κB activation (Lu et al., 2013; Madrid et al., 2000). Therefore, we investigated that the effect of nicardipine on LPS/IFN-γ-induced phosphorylation of Akt. As shown in Figure 9, Akt phosphorylation increased markedly within 1 h after LPS/IFN-γ stimulation and nicardipine pre-treatment resulted in significant blockage of LPS/IFN-γ-induced Akt phosphorylation, suggesting that Akt phosphorylation is involved in the inhibitory effect of nicardipine on LPS/IFN-γ-induced inflammatory response in BV-2 microglia.. 3.7. Nicardipine inhibits activation of microglia induced by LPS in mice. To confirm the suppression of neuroinflammatory response of nicardipine, we . 26 .
(35) . design an in vivo study of mice. The mice were divided into four groups: control, LPS, LPS plus nicardipine (5 mg/kg) and LPS plus nicardipine (50 mg/kg). The control group received normal saline intraperitoneal (IP) and LPS group received LPS (20 mg/kg) IP. In the treatment group, IP injections of nicardipine and the dose divide to 5 mg/kg and 50 mg/kg for 3 days first. Two hours later in the third day of injection of nicardipine, LPS (20 mg/kg) was injected IP. All animals were sacrificed 24 h after completed injections. The immunohistostaining showed LPS IP induce activation of microglia by Iba-1 staining comparing to control group (Figure 11). These cells generally take on a large, ameboid shape, although some variance has been observed. In the treatment group, activated microglia were predominant suppressed and cell shape became thin and small. The result showed nicardipine suppress the activation of microglia induced by LPS in mice.. . 27 .
(36) . Chapter Four: Discussion and Conclusion Nicardipine is a dihydropyridine-type calcium channel blocker (CCB) with a peculiar cerebrovascular profile developed approximately 30 years ago and the most frequent indication is antihypertensive agent for acute brain injury (Degoute, 2007; Gianino and Afuwape, 2012; Kim et al., 2012; Narotam et al., 2008; Qureshi et al., 2006; Sato et al., 2012). Nicardipine is widely administrated in the patients of intensive care because it shows effective antihypertensive effect and more safety. In comparison with other antihypertensive agents, their effects are similar to or even better than those exerted by other drugs. In clinical practice, nicardipine is superior to other antihypertensive agents in less cardiopulmonary suppression. Some studies show nicardipine may also play a role as neuroprotectants (Amenta et al., 1996; Amenta and Tomassoni, 2004; Amenta et al., 2008; Inzitari and Poggesi, 2005; Sato et al., 2012) but the definite mechanisms between nicardipine and glia in unknown. Acute brain injuries include ischemia stroke, spontaneous intracerebral hemorrhage and traumatic brain injury. All of these conditions often carried the crisis hypertensive due to autoregulation in CNS. The injuries within brain parenchyma trigger a series of adverse events causing secondary insults and severe neurological deficits. The neuroinflammatory response is the key mechanism in secondary insults and microglia play the most important role in neuroinflammation (Aloisi, 1999; Aronowski and Zhao, 2011; Block and Hong, 2005; Kettenmann et al., 2011).. . 28 .
(37) . We design a series study including in vivo and in vitro method and the result demonstrate that nicardipine could suppress neuroinflammatory response through MAPK/Akt pathway and inhibit the activation of microglia. These results maybe explain the neuroprotective effect in clinical practice of nicardipine.. . 29 .
(38) . Figures. . 30 .
(39) . Figure 4 Chemical structure and the viability on nicardipine treatment. (A) Chemical structures of (±)-2-(benzyl-methyl amino) ethyl methyl 1,4-dihydro-2,6-dimethyl4-(m-nitrophenyl)-3,5-pyridinedicarboxylate monohydrochloride. (B) Cell viability on nicardipine treatment in BV-2 microglia. Cells were treated with various concentrations of niardipine for 24 h, and cell viability was measured by the MTT assay. The results are expressed as means ± S.E.M. of three independent experiments.. . 31 .
(40) . Figure 5 Effect of nicardipine on ATP-cellular migration in microglia. (A) BV-2 microglia were pre-incubated with or without nicardipine for 60 mins followed by incubation with ATP (100 or 300 µM) for another 24 h. In vitro migratory activities were examined using cell culture inserts system. The results are expressed as means ± S.E.M. of three independent experiments. The migrated cells were visualized by phase-contrast imaging (B). * means p value <0.05 compared with control group. # means p< 0.05 compared with the ATP-treatment group.. . 32 .
(41) . Figure 6 Inhibitory effect of cytokine expressions in LPS/IFN-γ-stimulated BV-2 microglia. (A) Cells were pretreated with various concentrations of nicardipine (1, 3, 5, or 10 µM) for 60 mins before application of LPS (10 ng/ml) plus IFN-γ (10 ng/ml) for another 24 h. (B) Cells were pretreated with various concentrations of nicardipine (1, 3, 5, or 10 µM) for 60 mins before application of peptidoglycan (10 µg/ml) for another 24 h. The culture media were collected and analyzed by NO production. The results are expressed as means ± S.E.M. of three independent experiments. * means p<0.05 compared with control group.. . 33 .
(42) . Figure 7 Inhibitory effect of iNOS and COX-2 expressions in LPS/IFN-γ-stimulated BV-2 microglia. (A and B) Cells were pretreated with various concentrations of nicardipine (1, 3, 5, or 10 µM) for 60 mins before application of LPS (10 ng/ml) plus IFN-γ (10 ng/ml) for another 24 h. (C and D) Cells were pretreated with various concentrations of nicardipine (1, 3, 5, or 10 µM) for 60 mins before application of peptidoglycan (10 µg/ml) for another 24 h. Whole cell lysis protein was extracted and subjected to western blot analysis for iNOS (A and C) and COX-2 (B and D) expression, respectively. The results are expressed as means ± S.E.M. of three independent experiments. The results are representative of three independent experiments.. . 34 .
(43) . Figure 8 Nicardipine suppresses expression of inflammatory mediators in microglial cells. BV-2 microglia were pretreated with various concentrations of nicardipine (1, 5, or 10 µM) for 60 mins followed by stimulation with LPS (10 ng/ml) plus IFN-γ (10 ng/ml) for another 6 h. The expressions of iNOS, COX-2, IL-6, and IL-1β were determined by real-time PCR. The results are expressed as means ± S.E.M. from at least three independent experiments. * means p<0.05 compared with control group. # means p<0.05 compared with the LPS/IFN-γ-treatment group.. . 35 .
(44) . Figure 9 Nicardipine suppresses LPS plus IFN-γ-induced MAPK and Akt signaling pathways. BV-2 microglia were pretreated with nicardipine (10 µM) for 60 mins followed by stimulation with LPS (10 ng/ml) plus IFN-γ (10 ng/ml) for another 60 mins. Whole cell lysis protein was extracted and subjected to western blot analysis. The results are representative of three independent experiments.. . 36 .
(45) . Figure 10 Nicardipine suppresses LPS plus IFN-γ-induced NF-κB and AP-1 signaling pathways. BV-2 microglia were pretreated with nicardipine (10 µM) for 60 mins followed by stimulation with LPS (10 ng/ml) plus IFN-γ (10 ng/ml) for another 90 mins. Whole cell lysis protein was extracted and subjected to western blot analysis. The results are representative of three independent experiments.. . 37 .
(46) . Figure 11 Inhibitory effect of nicardipine prevents LPS-induced microglial activation. Mice were treated with 5 or 50 mg/kg nicardipine intraperitoneally once daily for three consecutive days before a single intraperitoneal injection of 20 mg/kg LPS. Microglial morphology was visualized by DAB immunohistostaining with the anti-Iba-1 antibody. Each group n= 3.. . 38 .
(47) . References Aloisi, F., 1999. The role of microglia and astrocytes in CNS immune surveillance and immunopathology. Adv Exp Med Biol. 468, 123-33. Amenta, F., Strocchi, P., Sabbatini, M., 1996. Vascular and neuronal hypertensive brain damage: protective effect of treatment with nicardipine. J Hypertens Suppl. 14, S29-35. Amenta, F., Tomassoni, D., 2004. Treatment with nicardipine protects brain in an animal model of hypertension-induced damage. Clin Exp Hypertens. 26, 351-61. Amenta, F., et al., 2008. Nicardipine: a hypotensive dihydropyridine-type calcium antagonist with a peculiar cerebrovascular profile. Clin Exp Hypertens. 30, 808-26. Andrews, C.M., et al., 2012. Emergency neurological life support: intracerebral hemorrhage. Neurocrit Care. 17 Suppl 1, S37-46. Aronowski, J., Zhao, X., 2011. Molecular pathophysiology of cerebral hemorrhage: secondary brain injury. Stroke. 42, 1781-6. Beckman, J.S., et al., 1994. Reactions of nitric oxide, superoxide and peroxynitrite with superoxide dismutase in neurodegeneration. Prog Brain Res. 103, 371-80. Block,. M.L.,. Hong,. J.S.,. 2005.. Microglia. and. inflammation-mediated. neurodegeneration: multiple triggers with a common mechanism. Prog. . 39 .
(48) . Neurobiol. 76, 77-98. Brown, D.R., 2001. Microglia and prion disease. Microsc Res Tech. 54, 71-80. Caivano, M., 1998. Role of MAP kinase cascades in inducing arginine transporters and nitric oxide synthetase in RAW264 macrophages. FEBS Lett. 429, 249-53. Chen, J.H., et al., 2011. Ghrelin induces cell migration through GHS-R, CaMKII, AMPK, and NF-kappaB signaling pathway in glioma cells. J Cell Biochem. 112, 2931-41. Degoute, C.S., 2007. Controlled hypotension: a guide to drug choice. Drugs. 67, 1053-76. Dissing-Olesen, L., et al., 2007. Axonal lesion-induced microglial proliferation and microglial cluster formation in the mouse. Neuroscience. 149, 112-22. Gianino, J.W., Afuwape, L.O., 2012. Evidence-based guidelines for the management of traumatic brain injury. Mo Med. 109, 384-7. Gross, H., et al., 2012. Emergency neurological life support: acute ischemic stroke. Neurocrit Care. 17 Suppl 1, S29-36. Haley, E.C., Jr., et al., 1994. A randomized trial of two doses of nicardipine in aneurysmal subarachnoid hemorrhage. A report of the Cooperative Aneurysm Study. J Neurosurg. 80, 788-96. Iadecola, C., Anrather, J., 2011. The immunology of stroke: from mechanisms to . 40 .
(49) . translation. Nat Med. 17, 796-808. Inzitari, D., Poggesi, A., 2005. Calcium channel blockers and stroke. Aging Clin Exp Res. 17, 16-30. Kettenmann, H., et al., 2011. Physiology of microglia. Physiol Rev. 91, 461-553. Kim, S.H., Smith, C.J., Van Eldik, L.J., 2004. Importance of MAPK pathways for microglial pro-inflammatory cytokine IL-1 beta production. Neurobiol Aging. 25, 431-9. Kim, S.Y., et al., 2012. Effectiveness of nicardipine for blood pressure control in patients with subarachnoid hemorrhage. J Cerebrovasc Endovasc Neurosurg. 14, 84-9. Koizumi, S., et al., 2002. Spatial and temporal aspects of Ca2+ signaling mediated by P2Y receptors in cultured rat hippocampal astrocytes. Life Sci. 72, 431-42. Komagiri, Y., Nakamura, K., Kubokawa, M., 2011. A nicardipine-sensitive Ca2+ entry contributes to the hypotonicity-induced increase in [Ca2+]i of principal cells in rat cortical collecting duct. Cell Calcium. 49, 35-42. Lee, S.C., et al., 1993. Cytokine production by human fetal microglia and astrocytes. Differential induction by lipopolysaccharide and IL-1 beta. J Immunol. 150, 2659-67. Liu, B., Du, L., Hong, J.S., 2000. Naloxone protects rat dopaminergic neurons against inflammatory damage through inhibition of microglia activation and. . 41 .
(50) . superoxide generation. J Pharmacol Exp Ther. 293, 607-17. Liu, B., Hong, J.S., 2003. Role of microglia in inflammation-mediated neurodegenerative diseases: mechanisms and strategies for therapeutic intervention. J Pharmacol Exp Ther. 304, 1-7. Lu, D.Y., et al., 2010. Glial cell line-derived neurotrophic factor induces cell migration and matrix metalloproteinase-13 expression in glioma cells. Biochem Pharmacol. 80, 1201-9. Lu, D.Y., Leung, Y.M., Su, K.P., 2013. Interferon-alpha induces nitric oxide synthase expression and haem oxygenase-1 down-regulation in microglia: implications of. cellular. mechanism. of. IFN-alpha-induced. depression.. Int. J. Neuropsychopharmacol. 16, 433-44. Madrid, L.V., et al., 2000. Akt suppresses apoptosis by stimulating the transactivation potential of the RelA/p65 subunit of NF-kappaB. Mol Cell Biol. 20, 1626-38. Moon, D.O., et al., 2007. Bee venom and melittin reduce proinflammatory mediators in lipopolysaccharide-stimulated BV2 microglia. Int Immunopharmacol. 7, 1092-101. Narotam, P.K., et al., 2008. Management of hypertensive emergencies in acute brain disease: evaluation of the treatment effects of intravenous nicardipine on cerebral oxygenation. J Neurosurg. 109, 1065-74. Ng, Y.K., Ling, E.A., 1997. Induction of major histocompatibility class II antigen on microglial cells in postnatal and adult rats following intraperitoneal injections . 42 .
(51) . of lipopolysaccharide. Neurosci Res. 28, 111-8. Powers, W.J., et al., 2001. Autoregulation of cerebral blood flow surrounding acute (6 to 22 hours) intracerebral hemorrhage. Neurology. 57, 18-24. Qureshi, A., Palesch, Y., Investigators, A.I., 2012. Expansion of recruitment time window in antihypertensive treatment of acute cerebral hemorrhage (ATACH) II trial. J Vasc Interv Neurol. 5, 6-9. Qureshi, A.I., et al., 2006. Treatment of acute hypertension in patients with intracerebral hemorrhage using American Heart Association guidelines. Crit Care Med. 34, 1975-80. Qureshi, A.I., Palesch, Y.Y., 2011. Antihypertensive Treatment of Acute Cerebral Hemorrhage (ATACH) II: design, methods, and rationale. Neurocrit Care. 15, 559-76. Sato, S., et al., 2012. [Antihypertensive Treatment of Acute Cerebral Hemorrhage (ATACH)-II at Japan site: study design and advance construction of domestic research network]. Rinsho Shinkeigaku. 52, 642-50. Zhang, Y., Dong, C., 2007. Regulatory mechanisms of mitogen-activated kinase signaling. Cell Mol Life Sci. 64, 2771-89.. . 43 .
(52)
數據
+7




相關文件
In this section we define a general model that will encompass both register and variable automata and study its query evaluation problem over graphs. The model is essentially a
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
2 Center for Theoretical Sciences and Center for Quantum Science and Engineering, National Taiwan University, Taipei 10617, Taiwan!. ⇤ Author to whom correspondence should
2 Center for Theoretical Sciences and Center for Quantum Science and Engineering, National Taiwan University, Taipei 10617, Taiwan..
2 Center for Theoretical Sciences and Center for Quantum Science and Engineering, National Taiwan University, Taipei 10617, Taiwan..
2 Department of Materials Science and Engineering, National Chung Hsing University, Taichung, Taiwan.. 3 Department of Materials Science and Engineering, National Tsing Hua
北京中醫藥大學 天津中醫藥大學 上海中醫藥大學 遼寧中醫藥大學 南京中醫藥大學 浙江中醫藥大學 廈門大學 福建中醫藥大學 江西中醫藥大學 暨南大學
For a 4-connected plane triangulation G with at least four exterior vertices, the size of the grid can be reduced to (n/2 − 1) × (n/2) [13], [24], which is optimal in the sense
