行政院國家科學委員會專題研究計畫 期中進度報告
以 MPTP 誘發巴金森症之動物模式來探討其對睡眠的影響及
相關機制(1/2)
計畫類別: 個別型計畫 計畫編號: NSC92-2314-B-039-014- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 中國醫藥大學神經部 計畫主持人: 張芳嘉 計畫參與人員: 尹珮璐、林峻邦 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 5 月 26 日
行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
█期中進度報告
以 MPTP 誘發巴金森症之動物模式來探討其對睡眠的影響及相關機制 (1/2)
計畫類別:█ 個別型計畫 □ 整合型計畫
計畫編號:NSC 92-2314-B-039-014-
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日
計畫主持人:
張芳嘉
共同主持人:
計畫參與人員:
尹珮璐、林峻邦
成果報告類型(依經費核定清單規定繳交):█精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列
管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:
中國醫藥大學
中 華 民 國 93 年 05 月 20 日
行政院國家科學委員會專題研究計畫期中進度報告
以 MPTP 誘發巴金森症之動物模式來探討其對睡眠的影響及相關機
制 (1/2)
計畫編號:
NSC 92-2314-B-039-014-
執行期限:
92 年 08 月 01 日至 93 年 07 月 31 日
主持人:張芳嘉 中國醫藥大學神經部
一、中文摘要 Parkinson’s disease (PD) 目前已知是 一種中樞神經細胞退化所造成的疾病,主 要 是 由 於 substantia nigra 之 pars compacta 區域之神經元細胞損害,以致 striatum 內之神經細胞缺乏 dopamine 所 導致。在臨床上 PD 的病人,經常會有睡眠 方 面 的 問 題 , 例 如 : 失 眠 (insomnia) 、 parasomnias 、 日 間 嗜 睡 [daytime somnolence; 特 別 是 excessive day time sleepiness (EDS) 和 sleep attack]。雖然有 足夠的臨床資料顯示 PD 病人的睡眠形式 與正常人不同,但至今仍缺乏導致睡眠形 式不同之基礎細胞訊息傳遞機轉。在文獻 上顯示:PD 病人的 nigrostriatal 區域以及 脊 髓 液 中 cytokines 的 濃 度 有 明 顯 的 增 加,且一些 neurotrophins 有顯著的減少。 此外,在 PD 的病人,其一些與 apoptosis 相關的蛋白質在 striatum 內有增加的趨 勢。而導致 nigrostriatal dopamine neurons 損害的主因可能為中樞 cytokines 濃度增 加、減少 neurotrophins, 以致於細胞凋亡 (programmed cell death; apoptosis)。本計劃 是要來探討 cytokines 和 neurotrophins 在 PD 形成中之細胞內訊息傳遞的機制,以及 parkinsonism 老鼠之睡眠模式的改變,包 括 在 wakefulness 、 slow wave sleep (SWS) 、 以 及 rapid eye movement sleep (REMS) 的時間、睡眠深度 (slow wave activity)、以及睡眠結構 (sleep architecture) 之改變。我們初步的結果顯示:在以 MPTP 所誘發之 PD 老鼠中,在連續兩次 MPTP 靜脈注射後 15 天,SWS 在黑暗期有稍微 的增加,12 小時 SWS 總量由 20.2±1.6%增 加至 25.3±1.6% (n=6);而 REMS 在黑暗期 則 顯 著 地 由 2.7±0.7% 增 加 至 8.0±1.3% (p<0.05, n=6) ; 相 對 的 WAKE 總 量 由 77.1±1.9% 下 降 至 72.3±2.6% (p<0.05, n=6)。雖然 MPTP 會改變老鼠睡眠,但當 我們再確認組織學傷害時發現 MPTP 在大 鼠 並 不 會 造 成 substantia nigra 之 pars compacta 區 域 之 神 經 元 細 胞 損 害 , 且 cytokine 在 MPTP 處理過的老鼠中並無顯 著變化。這些結果顯示 MPTP 對大鼠而言 並非產生 PD 的最佳模式。因此我們另外使 用 同 樣 是 屬 於 mitochondrial electron transport chain complex I inhibitor 的 rotenone 來誘發 PD。我們發現 rotenone 能 有 效 地 減 少 substantia nigra 之 pars compacta 區域之神經元細胞、減少 pole test score、以及降低 locomotion,為一產生 PD 的較佳模式。對睡眠的影響,我們的結 果顯示:在長期少量給予 rotenone 15 天 後,SWS 及 REMS 在黑暗期會顯著地增 加,相對的 WAKE 量下降。另外,我們初 步 的 結 果 顯 示 : 給 予 IL-1 受 體 拮 抗 劑 IL-1ra 能抑制 rotenone 對老鼠睡眠的改 變 。 而 且 我 們 發 現 在 老 鼠 striatum 中 pro-IL-1 濃度在剛進入黑暗期時有明顯的 增加。這些初步結果認為:在以 rotenone 誘發 PD 的模式對睡眠改變的影響可能是 藉由 IL-1 而來。下一步將探討 neurotrophins 可能扮演的角色及分析一些與 apoptosis 相 關的蛋白質(如:bcl-xl, bcl-2, bax, Fas 和 caspase-8) mRNA 在 striatum 之變化,並 使用 nuclear factor-κB (NF-κB) 抑制劑: pyrrolidine dithiocarbamate (PDTC) 、 或 activating protein-1 (AP-1) 抑 制 劑 : curcumin,來探討 IL−1經由 NF-κB 或 AP-1 之訊息傳遞在此動物模式中所扮演的角 色。neurotrophins、訊息傳遞
Abstract
The predominant lesion in Parkinsonism, observed in Parkinson’s disease (PD) is cell degeneration and loss of pigmented neurons in the pars compacta of the substantia nigra and the resulting loss of nerve terminals accompanied by dopamine (DA) deficiency in the striatum. Patients with PD clinically can experience a number of sleep disorders, including insomnia, parasomnias and daytime somnolence, especially excessive day time sleepiness (EDS) and sleep attacks. Although a body of clinical evidence suggests that sleep pattern may be altered in PD patients, there is a lack of basic cellular mechanisms. It has been previously reported that cytokines concentrations markedly increase and certain neurotrophins decrease in the nigrostriatal DA regions and ventricular and lumbar cerebrospinal fluid of PD patients. In addition, it has been found that the levels of the apoptosis-related proteins such as bcl-2 and soluble Fas (sFas) in the striatum of PD patients are elevated. Accordingly it has been hypothesized that central cytokines and neurotrophins may play an important role in the development of PD. This project is designed to elucidate the possible cellular mechanisms for cytokines and neurotrophins in PD development by pharmacology and molecular biology methods and to determine the mechanism for alteration in sleep pattern, including the total amount of time spending in distinct stages [wakefulness, slow wave sleep (SWS), and rapid eye movement sleep (REM sleep)], sleep intensity (slow wave activity), and sleep architectures. Our preliminary results indicate that fifteen days after two-dose IV administration of MPTP SWS and REMS significantly increase during the 15th-day dark period, with a mirrored effect of WAKE decrease. Although sleep is altered after MPTP treatment, there is no significant dopaminergic neuronal loss in the pars compacta of substantia nigra confirmed by tyrosine hydroxylase (TH) stain. In addition, our results also showed that cytokine concentrations in several brain regions are not altered after MPTP treatment. This
evidence that MPTP is not a suitable substance to induce parkinsonism in rats. The toxicity of MPTP appears to result from inhibition of complex I of the mitochondrial electron transport chain by its metabolite, MPP+. Therefore we chose the other potent and highly specific inhibitor of complex I, rotenone, which was chronically and systemically administered (for 14 days) through an abdominal osmotic minipump. We found that rotenone causes dopaminergic neuronal loss in pars compacta of substantia nigra, significantly reduces pole test score, and attenuates locomotive activity during the dark period in the 15th-day. This indicates rotenone induces parkinsonism in rats. Our preliminary data have shown that SWS and REMS significantly increase during the 15th-day dark period with a mirrored effect of WAKE decrease, after chronic rotenone treatment. In addition, intracerebroventricular (ICV) administration of IL-1 receptor antagonist (IL-1ra) significantly blocks rotenone-induce sleep-wake alteration. Analysis of IL-1 expression in several brain regions found that pro-IL-1 (35 KD) is significantly increased in striatum after 14-day of rotenone. These results suggest that IL-1 mediates rotenone-induced sleep-wake alterations. We will next examine the involvement of neurotrophin in this animal model. In addition, we will further analyze the expression of certain apoptotic proteins, such as bcl-xl, bcl-2, bax, Fas and caspase-8, in striatum after chronic rotenone treatment, and determine the effects of inhibiting nuclear factor-κB (NF-κB) or activating protein-1 (AP-1) by pyrrolidine dithiocarbamate (PDTC) or curcumin, respectively, to elucidate the cellular signal transduction involving in the rotenone- induced sleep alterations.
Keywords: parkinsonism, sleep, cytokine,
neurotrophin, signal transduction 二、緣由與目的
The predominant lesion in Parkinsonism, observed in Parkinson’s disease (PD) is cell degeneration and loss of pigmented neurons in the pars compacta of the substantia nigra and the resulting
loss of nerve terminals accompanied by dopamine (DA) deficiency in the striatum. Inflammation and the associated release of inflammatory cytokines such as interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-α) have been hypothesized that these mediators may be a component of neurodegenerative diseases associated with aging (i.e. Alzheimer’s disease and Parkinson’s disease) (1;2;4;7) or chronic human immunodeficiency virus (HIV) infection [review (6)]. One hypothesis of the cause of the degeneration of the nigrostriatal dopamine neurons, although there are several distinct risk factors, is that increased central cytokine expressions and/or decreased neurotrophins induce programmed cell death (apoptosis) contributing to the cause of Parkinson’s disease (4;5). Little evidence however in literatures elucidates the involvement of cytokines and neurotrophins in PD. Nagatsu et al. have reported that cytokine concentrations, such as TNF-α, IL-1β, IL-2, IL-4, IL-6 markedly increase and certain neurotrophins [i.e. brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF)] decrease in the nigrostriatal DA regions and ventricular and lumbar cerebrospinal fluid of PD patients. In addition, they also found the levels of the apoptosis-related proteins such as bcl-2 and soluble Fas (sFas) in the striatum of PD patients are elevated (4;5). Accordingly they suggest that increased central cytokines and decreased neurotrophins may play an important role in apoptosis and in the degeneration of nigrostriatal DA neurons in development of PD. However, the role for cytokines contributing to PD is still not full understood.
Most of neurons that are affected under aforementioned conditions, i.e. inflammation or infection require a constant supply of trophic factors such as NGF for survival. NGF binds to a high affinity receptor - TrkA and a low affinity receptor - p75 (NTR), a member of the TNF-α receptor (TNFR-1) superfamily, and generates signals via the activation of nuclear factor (NF)-κB contributing to
neuronal cell survival, differentiation, and programmed cell death. The mechanism(s) underlying NGF-mediated rescue of neurons from apoptosis may mediate through the effect of high affinity NGF receptor TrkA. Yoon et al. had reported that TrkA-mediated mitogen-activated protein kinase (MAPK) activation and suppression of c-Jun kinase activity initiated by p75 prevent cells from apoptosis (8). These results imply that the interaction between tyrosine kinase and cytokine receptors provides a novel mechanism that achieves alternative cellular responses by merging signals from different ligand-receptor systems.
TNF is a potent proinflammatory cytokine that plays an important role in immunity and inflammation, as well as in the control of cell proliferation, differentiation and apoptosis. TNF-α exerts its effects through two distinct receptors, TNFR1 and TNFR2 (3). Collectively, TNFR1-mediated signals could transduce apoptotic and anti-inflammtory signals through the recruitment of Fas-associated death domain protein (FADD) and Caspase-8, and also mediate anti-apoptotic and inflammatory responses through the recruitment of receptor-interacting protein 1 (RIP1) and TNF-receptor-associated factor 2 (TRAF2). Occupancy of TNFR2 results in direct recruitment of TRAF2, which in turn recruit TRAF1. Signals mediated by TNFR2 transmit anti-apoptotic and inflammatory actions [see review (3)]. Furthermore, IL-1β via IL-1 receptor type 1 activates adenylate cyclase activity and recruits TRAF 6, which subsequently activate NF-κB. NF-kB target genes can inhibit Caspase activation and suppress the apoptosis. Therefore, NF-κB as an evolutionarily conserved biochemical and molecular cascades may play an important role in prevention of cell death and/or promote neuronal plasticity.
TNF-α and IL-1 have been well known as a sleep promoter and NGF also has been reported exhibiting actions of sleep enhancement, but limit information could be provided for the involvement of TNF-α, IL-1 and NGF in the sleep attack with PD patients.
Therefore, the present study was designed to elucidate the role of interplay between central immune system (TNF-α or IL-1) and neural system (NGF) on the MPTP- or rotenone-induced sleep disturbance.
三、結果與討論
Our result indicates that, fifteen days after two-dose IV administration of MPTP (5 mg/kg/day), SWS and REMS significantly increase during the 15th-day dark period with a mirrored effect of WAKE decrease. The time spent in SWS increase from 20.2±1.6% to 25.3±1.6% (p>0.05, n=6) during the 12-h dark period, REMS increases from 2.7±0.7% to 8.0±1.3% (p>0.05, n=6), and WAKE decreases from 77.1±1.9% to 72.3±2.6% (p>0.05, n=6; Figure 1). Although sleep is altered after MPTP treatment, there is no significant dopaminergic neuronal loss in the pars compacta of substantia nigra confirmed by tyrosine hydroxylase (TH) stain. In addition, our results also showed that cytokine concentrations in several brain regions are not altered after MPTP treatment. This evidence that MPTP is not a suitable substance to induce parkinsonism in rats. The toxicity of MPTP appears to result from inhibition of complex I of the mitochondrial electron transport chain by its metabolite, MPP+. Therefore we chose the other potent and highly specific inhibitor of complex I, rotenone, which was chronically and systemically administered through an abdominal osmotic minipump (3 mg/kg/day, total volume of a minipump is 2 ml, infusion rate at 5 µl/hour for 14 days). We found that rotenone causes dopaminergic neuronal loss in pars compacta of substantia nigra (since the image is still in process, the data have not shown here), significantly reduces pole test score (Table 1), and attenuates locomotive activity during the dark period in the 15th-day (Figure 2). This indicates rotenone successfully induces parkinsonism in rats. Our preliminary data have shown that SWS and REMS significantly increase during the 15th-day dark period with a mirrored effect of WAKE decrease, after chronic rotenone treatment. SWS increases from 20.0±1.6% after vehicle (pyrogen-free saline; PFS) to 26.7±1.7% after rotenone, REMS increases
from 3.8±0.7% after PFS to 7.9±1.0% after rotenone, and WAKE decreases from 76.3±1.9% after PFS to 65.4±2.5% after rotenone (p<0.05, n=7; Figure 3). Intracerebroventricular (ICV) administration of IL-1 receptor antagonist (IL-1ra) significantly blocks rotenone-induce sleep-wake alteration in a dose-dependent manner. SWS decreases from 26.7±1.7% after rotenone to 19.5±2.5% and 16.3±1.4% after 100 and 200 ng of IL-1ra, respectively (p<0.05, n=7; Figure 3). REMS decreases from 7.9±1.0% after rotenone to 4.2±0.8% and 3.8±0.7% after 100 and 200 ng of IL-1ra, respectively (p<0.05, n=7; Figure 3). Analysis of IL-1 expression in several brain regions found that pro-IL-1 (35 KD) is significantly increased in striatum at the first-hour of the dark period after 14-day of rotenone, however the active form of IL-1 (17 KD) is not altered (Figure 4). The explanation for the unaltered expression in active IL-1 after rotenone is that the level of IL-1 is at the lowest of its circadian fluctuation at the beginning of the dark period, the enhancement may occur later since we have found the increase of pro-IL-1. These results suggest that IL-1 mediates rotenone-induced sleep-wake alterations. We will examine the involvement of neurotrophin in this animal model during the second year. In addition, we will further analyze the expression of certain apoptotic proteins, such as bcl-xl, bcl-2, bax, Fas and caspase-8, in striatum after chronic rotenone treatment, and determine the effects of inhibiting nuclear factor-κB (NF-κB) or activating protein-1 (AP-1) by pyrrolidine dithiocarbamate (PDTC) or curcumin, respectively, to elucidate the cellular signal transduction involving in the rotenone- induced sleep alterations.
四、計畫成果自評
At the beginning of this project we try to establish a parkinsonism model in rat by IV administration twice MPTP, however our result suggest that MPTP fails to induce dopaminergic neuronal loss in substantia nigra and fails to induce deficit in locomotor activity and limb weakness test in rats. Although our results showed that sleep-wake
activity was altered after administration of MPTP, this sleep disturbance may be simply due to the effect of MPTP toxicity itself rather than the consequence of parkinsonism. Then we selected another more potent and highly specific inhibitor of complex I, rotenone, administered chronically and systemically for 14 days through an abdominal osmotic minipump, and we successfully established parkinsonism in rats. Our results indicate pro-IL-1 expression in striatum increases and IL-1ra blocks sleep-wake alteration in rotenone-induced parkinsonism rats, suggesting the involvement of IL-1 in sleep disturbance in parkinson’s disease. During the second year we will further elucidate the cellular mechanism of NF-κB and apoptotic proteins in this sleep disturbance, and the role of neurotrophins.
五、參考文獻
1. Grunblatt E, Mandel S, and Youdim M. MPTP and 6-hydroxydopamine
-induced neurodegeneration as models for Parkinson's disease: neuroprotective strategies. Journal of Neurology
247(Suppl 2), 95-102. 2000.
2. Mandel S, Grunblatt E, and Youdim M. cDNA microarray to study gene
expression of dopaminergic
neurodegeneration and neuroprotection in MPTP and 6-hydroxydopamine models: implications for idiopathic Parkinson's disease. Journal of Neural Transmission.Supplementum 60, 117-124. 2000.
3. Mattson MP, Culmsee C, Yu Z, and Camandola S. Role of nuclear factor kB in neuronal survival and plasticity. Journal of Neurochemistry 74, 443-456. 2000.
4. Nagatsu T, Mogi M, Ichinose H, and Togari A. Changes in cytokines and neurotrophins in Parkinson's disease [Review]. Journal of Neural
Transmission.Supplementum 60, 277-290. 2000.
5. Nagatsu T, Mogi M, Ichinose H, and Togari A. Cytokines in Parkinson's disease. Journal of Neural
Transmission.Supplementum 58, 143-151. 2000.
6. Poli G and Fauci AS. The effect of cytokines and pharmacological agents on chronic HIV infection. AIDS Research & Human Retroviruses 8(2), 191-197. 1992.
7. Tong L, Toliver-Kinsky T, Taglialatela G, Werrbach-Perez K, Wood T, and Perez-Polo JR. Signal transduction in neuronal death. Journal of
Neurochemistry 71, 447-459. 1998. 8. Yoon SO, Casaccia-Bonnefil P, Carter
B, and Chao MV. Competitive
signaling between TrkA and p75 nerve growth factor receptors determines cell survival. Journal of Neuroscience 18(9), 3273-3281. 1998.
Table 1. Behavioral test before and after chronic rotenone infusion.
Before rotenone After rotenone Pole test 2.8±0.2 (n=6) 1.5±0.2* (n=6) Traction test 2.8±0.2 (n=6) 2.5±0.2 (n=6)
Figure 1. The alteration of SWS and REMS after MPTP treatment
% of SWS 0 10 20 30 40 50 Time (hour) 1 2 3 4 5 6 7 8 9 10 11 12 % of REMS -10 0 10 20 30
The open circle represents the values obtained before MPTP treatment and the close circle indicates the values obtained at the 15th-day after twice administered MPTP (5mg/kg/day).
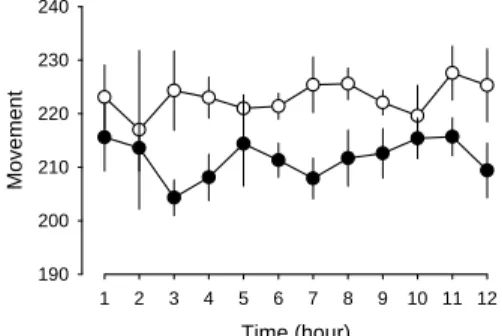
Figure 2. Alteration of locomotive activity after chronic rotenone infusion.
Time (hour) 1 2 3 4 5 6 7 8 9 10 11 12 Mo ve men t 190 200 210 220 230 240
The open circle represents the locomotion activity obtained before rotenone treatment and the close circle indicates the values obtained after 14-day chronic infusion of rotenone (3 mg/kg/day).
Figure 3. Sleep alterations after chronic rotenone treatment and IL-1 receptor antagonist, IL-1ra blocks rotenone-induced sleep disturbance.
% of SW S 0 10 20 30 40 50 Time (hour) 1 2 3 4 5 6 7 8 9 10 11 12 % of RE MS -5 0 5 10 15 20 25 0 5 10 15 20 25 30 0 2 4 6 8 10
*
*
# # # #The shade area represent the control values, the open circle depicts the values after 14-day chronic infusion of rotenone (3mg/kg/day), and the close circle indicates the values obtained after ICV administration of IL-1ra (200 ng). Insert bar graphs are the summary for rotenone and IL-1ra+rotenone effects; open bar: control, close bar: 14-day rotenone, hatch bar: 100 ng IL-1ra, close bar: 200 ng IL-1ra.
Figure 4. Pro-IL-1 expression in striatum is altered after chronic rotenone treatment.
35 KD is the molecular weight for pro-IL-1β, numbers 1-4 represents the rat numbers.