International Journal of
Molecular Sciences
ISSN 1422-0067 www.mdpi.com/journal/ijms ArticleUsing the internal ribosome entry site of tau and amyloid-
precursor protein as target to search potential nature products
for Alzheimer’s disease
Yun-Chieh Tasi1, †, Ting-Yu Chin2,5, †, Ying-Ju Chen2, Chun-Chih Huang 3, Shou-Lun Lee4,* and Tzong-Yuan Wu2,5*
1 Graduate Institute of Life Sciences, National Defense Medical Center, Taipei, Taiwan
2 Department of Bioscience Technology, Chung Yuan Christian University, Taoyuan, Taiwan 3 New Bellus Enterprise Co., Ltd., Tainan, Taiwan
4 Department of Biological Science and Technology, China Medical University, Taichung, Taiwan 5 Program and Center of Nanoscience Technology, Chung Yuan Christian University, Taoyuan,
Taiwan
† These authors contributed equally to this work.
* Author to whom correspondence should be addressed; E-Mail: [email protected] (T.Y. Wu);
[email protected] (S.L. Lee); Tel.: +886-3-2653520 (T.Y. Wu); Tel.: +886-4-22053366x2526
(S.L. Lee); Fax: +866-3-2653599 (T.Y. Wu); Fax: +886-4-22051507 (S.L. Lee) Academic Editor:
Received: / Accepted: / Published:
Abstract: Overexpression of the amyloid precursor protein (APP) and the hyperphosphorylation of the tau protein are vital in the understanding of the cause of Alzheimer’s disease (AD). As a consequence, regulation of the expression of both APP and tau proteins is one important approach in combating AD. The APP and tau proteins can be targeted at the levels of transcription, translation and protein structural integrity. This paper reports the utilization of a bi-cistronic vector containing either APP or tau IRES elements flanked by-galactosidase gene (cap-dependent) and secreted alkaline phosphatase (SEAP) (cap-independent) to discern the mechanism of action of memantine, an N-methyl-D- aspartate (NMDA) receptor antagonist. Results indicated that memantine was capable of reducing the actvity of both the APP and tau IRES at a concentration of ~10 M 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
monitored by the -galactosidase assay. Western blot analysis of the tau protein in neuroblastoma (N2A) and rat hippocampal cells confirmed the halting of the expression of the tau proteins. We also employed this approach to identify a preparation named NB34, extracts of Boussingaultia baselloides (madeira-vine) fermented with Lactobacillus spp., which can work like the memantine in both IRES of APP and Tau. Water maze test demonstrated that NB34 could improve the spatial memory of high fat diet induced neurodegeneration in apolipoprotein E-knockout (ApoE-/- ) mice. These results revealed
that the bi-cistronic vector provided a simple, and effective platform in screening and establishing the mechanistic action of potential compounds for the treatment and management of AD.
Keywords: Memantine, amyloid precursor protein, tau, Alzheimer’s disease, internal ribosome entry sites, bi-cistronic
1. Introduction
Alzheimer’s disease (AD) is considered as the most common neurodegenerative malady in the modern but senile society [1]. This condition is characterized primarily by dementia which attacked an estimated 35.6 million people worldwide and the numbers are estimated to be doubling every 20 years [2, 3]. Anatomical dissections and analysis of the brain of AD patients lead to the identification of two hallmarks defining the neuropathological characteristics of this disease: neuritic plaques and neurofibrillary tangles (NFTs). Under the electron microscope, abnormal amyloid-like filaments were found in the plaques and neurofibrillary tangles [4, 5]. The specific sites of plaques and tangles are different: plaque filaments are extracellular but most of the tangled filaments are present intracellularly; deposited in nerve cell bodies, as well as in neurites of neuron. The major molecular compositions of the plaques and tangles are also different: amyloid- (A) peptide [6] is the major plaque component while the tau protein [7] is the major tangle component. The 40-42 amino acid A peptide is derived from the sequential cleavage of amyloid precursor protein (APP), a type 1 transmembrane protein, by two proteases, - and -secretase [1, 2]. Tau protein, on the other hand, is one of the main neuronal microtubule-associated protein and functions importantly in the modulation of microtubule organization during morphogenesis and process outgrowth in neurons [8]. Transgenic mouse models of AD that target the APP and tau genes also confirmed the pathogenetic factors [9-12]. All these studies indicated that the control of the expressions of APP and tau in the brain may be a good target for drugs that can potentially be used in the treatment of AD.
Aside from the canonical cap-dependent model of recognition and ribosomal scanning, there is an alternative method of translation initiation and named as cap-independent translation is first described in members of the family Picornaviridae [13, 14] and subsequently for a growing subset of cellular mRNAs [15, 16]. In the cap-independent mechanism of translation initiation, ribosomes are recruited to the mRNA by RNA structural elements called internal ribosome entry sites (IRESes) [17]. Since it had been shown that many cellular mRNAs contain IRESes, it is likely that up to 10% of all mRNAs have the capability to initiate translation by the cap-independent mechanism [18, 19]. It is also 2 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75
apparent that genes involved in a diverse range of cellular activities, including proliferation, growth and apoptosis employ this alternative mechanism resulting to the consideration of internal initiation through IRES as an important cellular mechanism and not just a specialized viral strategy [15]. Recent studies demonstrated that the APP mRNA may be translated through an IRES wherein APP mRNA was found to be one of the several mRNAs which may remain associated with polyribosomes during mitosis, when cap-dependent translation initiation is greatly diminished [20]. Interestingly, it has also been reported that the 5’ leader in the human tau mRNA contains an IRES and that the IRES-dependent translation plays a significant role in the generation of the tau protein [21].
Memantine is a US Food and Drug Administration-approved, uncompetitive N-methyl-D-aspartate
(NMDA) receptor antagonist and reduces the clinical deterioration in moderate to severe AD. Preclinical evaluations regarding the use of memantine as an NMDA receptor antagonist were reviewed and reported extensively [22-26]. Besides, memantine is also capable of halting and reversing the protein phosphatase (PP)-2A inhibition-induced abnormal hyperphosphorylation of tau/neurofibrillary degeneration [27]; protects the neurons from microglial-inflammatory responses that result to cell death [28]. Recently, our laboratory linked NMDA receptor antagonists, amantadine and memantine, [22] with the down-regulation of the IRES of enterovirus 71 and encephalomyocarditis virus [29, 30].
In this study, we report that the NMDA receptor antagonist, memantine can suppress the expression of neuronal APP and tau proteins through the novel cap-independent translational initiation mechanism. Based on this observation, we also employed the IRESes of APP and Tau as the potential targets for AD to screen Lactobacillus spp. fermented traditional Chinese herbs and identified a preparation that can inhibit the translational activity of the Tau IRES. Water maze test demonstrated that this fermented preparation could improve the spatial memory of high-fat diet (HFD) induced neurodegeneration in ApoE-/- mice.
2. Results
2.1. The Amyloid precursor protein and tau IRESes construct
The expression of APP and tau proteins had been demonstrated in a number of reported studies to be mediated by IRES, an atypical translational initiation mechanism, aside from the conventional cap-dependent translation initiation [20, 21].
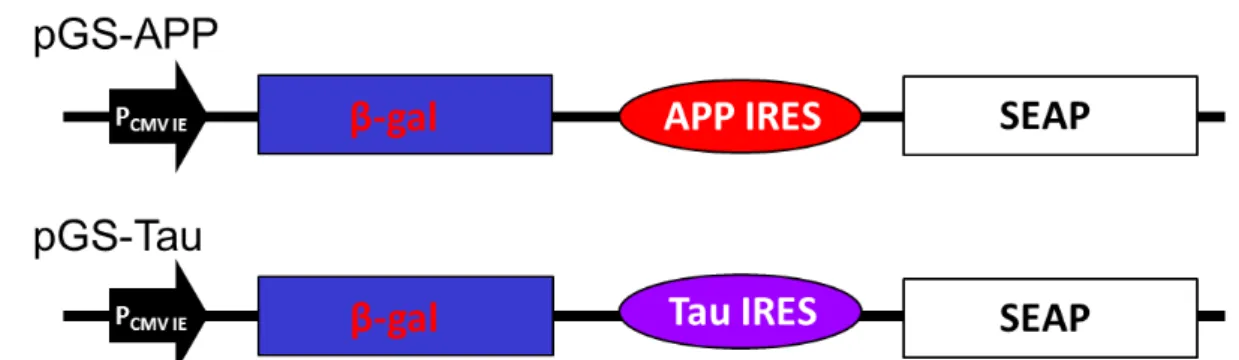
A previous study had shown that amantadine can inhibit the translation activity of IRES derived from HAV, enterovirus 71 or encephalomyocarditis virus [30]. It is interesting to note that the chemical structure of amantadine is similar to memantine, a therapeutic drug for moderate to severe AD and both are tricyclic symmetric amines. The plasmid pTriEx4 containing either the genes for the -galactosidase or the secreted alkaline phosphatase were used for the construction of the two cistronic vectors as shown in Figure 1A and 1B (as described in Materials and methods). The bi-cistronic vectors were generated by inserting either the APP (pGS-APP) or Tau (pGS-Tau) IRES of DNA fragment in between the β-galactosidase (-Gal) and secreted alkaline phosphatase (SEAP) reporter genes (Fig. 1).
5 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114
Fig 1. Construct of pGS-APP and pGS-Tau. The bi-cistronic vectors contain the genes for beta-galactosidase (-Gal) and secretory alkaline phosphatase (SEAP), IRES element from the APP or the Tau genes, APP IRES and Tau IRES, respectively. PCMV IE is the
promoter of human cytomegavirus immediately early promoter.
2.2. The tissue tropism of APP and tau IRESes and the effect of memantine on APP and tau IRESes Literatures report that IRESes’ ability to initiate translation varies greatly in cells of different origin. Therefore, we presumed that both APP and tau IRESes could drive more efficient cap-independent translation in neuron-like cells, i.e. N2A, than in non-neuronal cells such as COS-1 and CHO cells. To test this presumption, we performed transient transfection assays on different cell lines using the plasmids listed in Fig. 1.
It was observed that the CHO cells gave a two-fold -Gal activity upon transfection with pGS-APP plasmid signifying an efficient transfection of the plasmid DNA as compared to the N2A and the COS-1 cells (Figure 2A). However, considering the activity of the reporter protein- SEAP, it is apparent that the N2A cells showed a significantly increased secreted alkaline phosphatase activity that is thrice as the measured activity in CHO cells whereas, the SEAP activity was not observed in COS-1 cells. After normalizing the SEAP activity using the -Gal assay, results clearly indicated that APP IRES favored the cap-independent translation specifically to the N2A neuron-like cells. Similar data were obtained after cell lines were transfected with the plasmid containing the tau IRES (Figure 2B).
Fig 2. Determination of cell tropism of the APP and tau IRES. Transient transfection with the corresponding plasmid was performed on mammalian cells (~9 x 104 cells/well) of
different types using Lipofectamine 2000. Cap-dependent mechanism of translation was assessed by -galactosidase activity in the cell lysate while the IRES-dependent mechanism of translation was ascertained through the SEAP activity in the cell medium. Normalization was carried out by beta-galactosidase assay. (A) Comparison of the APP IRES and (B) Tau IRES activity in mouse N2A, CHO and COS-1 cells.
8 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140
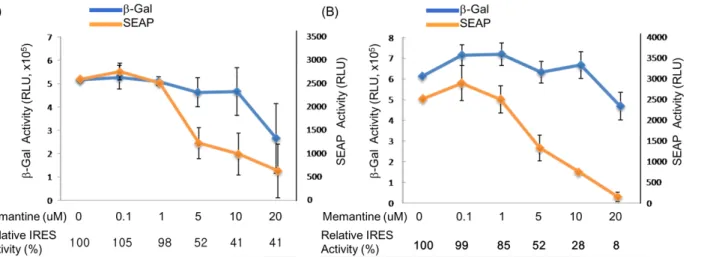
It is conspicuously seen in this result that the tau IRES is significantly more functional in the N2A cells since the other cell lines tested gave a very little or no SEAP activity at all. In addition, the tau IRES was also found to be more functional than the APP IRES in N2A cells considering the six-fold increase of the SEAP activity in the cell medium of the N2A cells transfected with the plasmid containing the tau IRES. The results presented herein were consistent with the tissue tropism [31] of either APP or Tau IRESes [32]. More interestingly, data revealed that memantine was able to inhibit the IRES activity of APP or tau without interfering with the cap-dependent translation. At a memantine concentration of 5 M, a significant decrease equivalent to 52% relative APP (Figure 3A) and tau (Figure 3B) IRES activity was observed after 24 hours treatment. This result implied that memantine, comparable to amantadine [30] can also act as an inhibitor of IRESes and on this note, a regulator of the translation of the APP and tau proteins. This in turn further supports the previously reported hypothesis [29] that memantine has a dual action in the management and treatment of Alzheimer’s disease. Memantine does not only obstruct the excitotoxicity of NMDA receptors [33-35] but also halts the expression of APP and tau proteins through IRES. The inhibition of the APP IRES and tau IRES by memantine might imply for the diminished A production and tauopathies as well as an antagonist of NMDA receptors in AD.
11 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158
Fig 3. Effect of various concentration of memantine on mouse N2A neuroblastoma cells. The N2A cells (~9 x 104 cells/well) were seeded onto a 24-well plate prior to
transfection. Lipofectamine (2 L) was used to transfect the plasmid DNA (1 g) into the neuroblastoma cells. At 24 hrs post treatment with memantine, the culture medium from each well was harvested and cells were lysed subsequent SEAP and beta-galactosidase assay were done, respectively. The effect of memantine on (A) APP IRES and (B) tau IRES, as evaluated by beta-galactosidase (cap-dependent) and SEAP (IRES-dependent) activities in murine neuroblastoma (N2A) cells.
2.3. Effect of memantine on the amyloidprecursor protein and tau expressions in neuronal cells
Aggregated A affects neurons and induces NFTs formation and neuronal loss, eventually leading to dementia [36]. In a number of cell culture based studies on the assessment of the pharmacological effects of memantine, it had been revealed that memantine exerted significant activity at a concentration range between 1 M and 20 M [35, 37, 38].
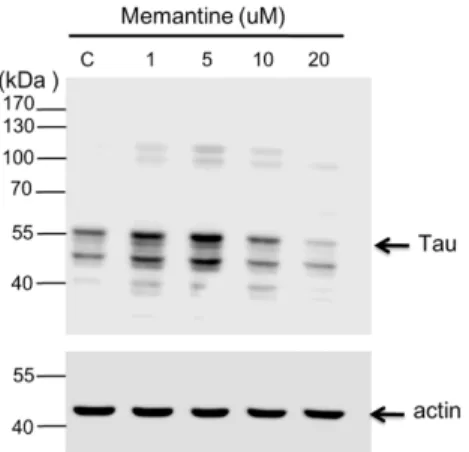
In this study, concentrations: 1, 5, 10 and 20 M of memantine were used to treat mouse N2A (Figure 4) and rat hippocampal (Figure 5) neuronal cells and to monitor the expression of tau and APP proteins. Tau is a single copy gene in both humans and rats that goes through alternative message splicing resulting to multiple isoforms in adults [39]. These isoforms are post-translationally modified via phosphorylation [40]. The monoclonal antibody for Tau-1 was used in this study identifies an amino acid sequence from 192-204 in humans and 180-198 in rats when all of the four serine residues are unphosphorylated [41]. Western blot analysis confirmed the presence of tau-1 proteins from both neuronal (mouse N2A and rat hippocampus) cell lysates treated with various concentrations of memantine (1, 5, 10 and 20 M) as revealed by bands at around 52-68 kDa. It is clearly depicted in Figure 4 that memantine was able to halt the expression of the tau-1 protein in the murine model as seen through the observed faint bands at approximately 52-55 kDa corresponding to different isoforms of mouse tau protein [42] after treatment with 10 M memantine. This is in agreement with the result obtained upon treatment of the rat hippocampal cells with memantine wherein the appearance of a diminished bandwidth along ~55 kDa at a concentration of 10 M signifies the stalling of the tau protein expression (Figure 5). Furthermore, densitometer scan of gel bands in Figure 5 indicated as the 10 or 20 µM of memantine were added in the culture medium of hippocampal cells, the tau proteins 14 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188
expression were inhibited about 25%. It is also evident in Figure 5 that dephosphorylation in the tau-protein may have happened after it was treated with 20 M memantine as shown by the two visible bands at lower molecular weight regions. The experimental results therefore provided initial evidences for the potential use of the IRES of the tau protein as a novel target for the screening of compounds for the treatment of AD and other tauopathies.
Fig 4. Western blot analysis showing the effect of memantine on tau protein expression in the N2A cell lysate. The neuroblastoma cells (~9 x 104 ) were treated with
different concentrations of memantine (1, 5, 10 and 20 M). The PVDF membrane was exposed to anti-tau-1 (clone PC1C6) antibody to detect the endogenous tau protein from N2A cells. Each lane corresponds to the concentration (1 – 20 M) of memantine treatment; Lane C is for the untreated cell lysate. Tau protein is visible as bands between the 40 and 55 kDa regions. Molecular weight marker is shown left most. Beta-actin was used as a loading control.
Fig. 5 Western blot analysis showing the effect of memantine on tau protein expression in the rat hippocampal cell lysate. The hippocampal cells (5 x 104/cm2) were
treated with different concentrations of memantine (1, 5, 10 and 20 M). The PVDF membrane was incubated with anti-tau-1 (clone PC1C6) antibody to identify the endogenous tau protein from N2A cells. Each lane corresponds to the concentration (1 – 20 M) of memantine treatment; Lane C is for the untreated cell lysate. Tau protein is visible as bands in between the 43 and 55 kDa regions. Molecular weight marker is shown left most. 17 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210
We also evaluated the effect of memantine on the expression of APP by Western analysis and revealed a minimal dose-dependent increase in the intracellular APP as the concentration approaches 10 M. It is noteworthy to mention that at a concentration of 20 M, the intracellular APP was increased markedly (data not shown). This result is consistent with the reports by Ray et al., [43], they have observed a significant increase in intracellular APP levels with 20μM dose of memantine in the human neuroblastoma cells. However, they also show that memantine treatment decreases levels of secreted APP and Aβ peptide in their reports [43]. These results may implied that memantine can enhance APP proteins expression through uncharacterized mechanism, although memantine can inhibit APP IRES activity. Such a novel property of memantine demands further study to clarify this conflicts and define its beneficial effect on AD.
2.4. Identification of NB34 as a potent inhibitor of Tau IRES
Mementine can inhibit both APP and Tau IRES revealed that both the IRES could be potential targets for compounds screening for AD. Traditional Chinese medicines (TCM) have been widely investigated for the treatment of Alzheimer’s disease [44]. And there were reports indicated fermentation of these Chinese herbs by microbes, like Lactobacillus spp., could dramatically enhance the concentration of active compounds. Thus, we tried to prepare 92 different preparations that were derived from Eleutherococcus senticosus 、 Lycium chinense Miller 、 Panax ginseg 、 Curcuma longa 、 Radix notoginseng and Gastrodia elata after fermented by Lactobacillus spp.. All the preparations were named NB1 to NB92. Fig. 6A shows that the preparation, named NB34, could inhibit the Tau IRES mediated translation activity in N2A cells. We further studied the dose response of NB 34 on the Tau IRES and APP IRES activity. Fig. 6B shows that the NB34 could inhibit the translational activity of Tau IRES as low as 0.02 mg/ml, although its effect on translational activity of APP IRES was only obviously on 0.05 mg/ml. Thus, we were interesting to investigate whether the NB34 could work like the Mementine be benefit to AD. NB34 was the product of Radix notoginseng fermented by Lactobacillus spp. Interesting, previous study has demonstrated anti-aging effect of Radix notoginseng on cultured neurons or rats with AD [45]. Thus, we tried to evaluate this novel NB34 preparation could be benefit for memory on mice by Morris water maze task.
20 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238
Fig. 6 Identification of NB34 as an inhibitor for Tau IRES in N2A cells. (A) The N2A cells (~9 x 104 cells/well) were seeded onto a 24-well plate prior to transfection.
Lipofectamine (1 L) was used to transfect the pGS-Tau plasmid DNA (1 g) into the neuroblastoma cells. At 24 hrs post treatment with various preparation (0.05 mg/ml) of Chinese herbs fermented by Lactobacillus spp., (NB1-92, and only the NB33-45 are shown) the culture medium from each well was harvested and cells were lysed subsequent SEAP and beta-galactosidase assay were done, respectively. (B) The dose dependent of NB34 on translation activity of APP IRES and Tau IRES in N2A cells. M, 0.25 μM memantine.
2.5. NB34 inhibits impairment of spatial learning induced by high fat diets during memory acquisition in ApoE-/- mice
ApoE4 allele is recognized as a prominent risk factor for the development of AD in human [46, 47]. In mice, ApoE protects against neuropathology induced by HFD and mice deficient in ApoE display disturbances in learning and memory function like the long-term potentiation [43], lose of synapses with age, or cytoskeleton alternations [50]. Thus, we employed the HFD feeding ApoE-/- mice to
address whether the NB34 could improve the spatial memory of HFD-fed ApoE-/- mice.
To assess whether simultaneously intake of NB34 could reverse HFD-induced learning and memory impairments in ApoE-/- mice, a battery of behavioral tests was conducted. Both groups learned to find
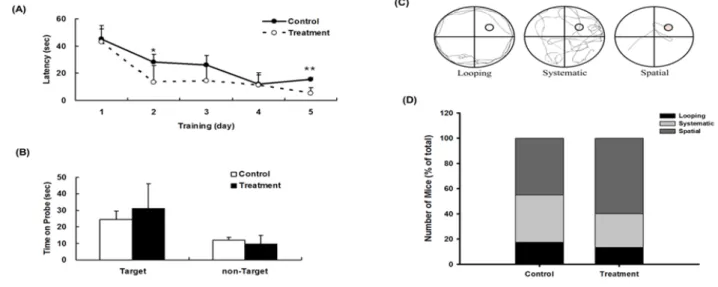
the hidden platform during the acquisition phase of training (Days 1–5), which was shown by a progressive decrease in latency to reach the platform. NB34-treated mice appeared to reach the platform faster than control mice at day 2 (p < 0.05; Fig. 7A) and day 5 (p < 0.005; Fig. 7A). However, in the probe trial, there was no significant difference in the time spent in the target quadrant between the two groups (Fig. 7B).
23 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262
To assess the effect of diet on spatial learning, mice were fed diets containing NB34, we also analyzed the respective search patterns shown by the mice to locate the hidden platform on each day of the first acquisition period in the Morris water maze test. We categorized the behavior of individual mice according to the incidence of distinct search strategies, an outcome that is less influenced by locomotion deficits. Fig. 7C indicated the individual mouse strategies reveal three learning phases, e.g., spatial strategy, systemic strategy, and looping strategy, following previously published criteria [51-53]. Three different search strategies were defined as follows: swimming directly to the correct target quadrant and searching was called the spatial strategy; systematic strategy was defined as searching interior portion of or entire tank; and if more than 70% of the swim trace was outside the circle and swimming around the wall of tank, it was defined as the looping strategy. The search strategies were examined for first of the 4 trials on different training days. ApoE-/- mice with NB34
treatment displayed increase spatial type (60% vs 45% from control mice) and reduced looping type (13.3% vs 17.5% from control mice).The mice with spatial strategy for platform indicating better learning behavior than those with looping strategy. Comparison of swimming strategies indicated that
NB34 treated mice had increased spatial strategies for platform indicating better learning while control
mice despite having increased systemic approaches to platform, they had also increased looping approaches indicating poor learning (Fig. 7D).
Fig. 7 Effects of NB34 on Spatial memory performance in the Morris water maze (MWM). (A) Acquisition trials. The time spent to reach the platform (escape latency).*p<0.05 when compared with the corresponding control groups; **p<0.005 when compared with the corresponding control groups. (B) Probe trial. Probe test performed 24 h after the hidden platform acquisition period. Time spent in the target quadrant was similar in two groups. (C) Examples of search strategies. (D) Comparison of swimming strategies during acquisition phase of MWM between control and NB34 treated ApoE-/- mice combine with
HFD. Search strategy was examined for first of the 4 trials on different acquisition days.
3. Discussion 26 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290
The bi-cistronic assay is considered as the gold standard to define internal initiation of translation and is one of the most widely used method for testing supposed IRES sequences [31, 54, 55]. Thus, we constructed bi-cistronic plasmids to monitor the activity of the 5’-UTR of both the APP and tau as well as to verify the neurotropism of these IRESes. It is noteworthy to mention that the ribosomes and other components of the translation machinery were also found in neuron’s dendritic processes although at minimal amounts compared to those found in the cell body [56-58]. Discoveries indicated that the APP [20, 59] and Tau [21] mRNAs can be translated through an IRES. It has been well documented that APP 5’leader contains an IRES and showed that IRES-dependent translation is a mechanism by which endogenous APP mRNA is translated [59]. Same is true with the tau protein wherein it exhibits the characteristics of a viral IRES that contains a relatively lengthy 5’ leader at 250 bp with a high guanine/cytosine (G/C) content and this IRES in turn functions in the regulation of the synthesis of the tau protein [21]. Therefore, these reports indicated that IRES are good targets for AD treatment due to the reason that the internal initiation of translation of the APP and tau mRNAs is an important mode for the synthesis of both APP and tau, a mechanism which is controlled by conditions that also contribute to AD pathology [59, 60]. Based on these observations, we also identified NB34, a preparation of Radix notoginseng fermented by Lactobacillus spp. that like memantine could inhibit the translational activity of Tau IRES. Interestingly, NB34 could rescue HFD-induced learning and memory impairments in ApoE-/- mice (Fig. 7). Although the functional components in NB34 that can
inhibit the translational activity of Tau IRES is unknown, the preparation of Radix notoginseng fermented by Lactobacillus spp. might be developed as a potential health food in the future. And, identification of the compound(s) that are responsible for NB34 function in learning and memory of ApoE-/- mice would be critical and conducted in the future.
In our study, the NMDA-receptor antagonist memantine was shown to inhibit both the APP and tau IRES without restraining the cap-dependent translation when the concentration of memantine is below 10 M as revealed through the monitoring of the activity of beta-galactosidase in N2A cells (Figures 3A and 3B). The result of our study is in good agreement with the recent study of Ray and co-workers [61] where treated human neuroblastoma SK-N-SH cells with 10 M memantine decreased the measured levels of secreted total APP (sAPP), APPisoform and A(1–40) in a time dependent
manner for up to 24 hours.
Result of transient transfection studies in different cell lines clearly implied the preferential activity of both APP and tau IRESes on neuronal cells than on CHO and COS-1 cells (Figures 2A and 2B) that could be attributed to the tissue tropism of the APP and tau IRESes. Anti-A(-amyloid) therapy has been a very ideal approach toward the development of treatment against AD. The secretion of Ain AD leads to the production of highly reactive oxygen species (ROS) and mitochondrial defects [62]. Thereby, the continuous secretion of Atogether with high levels of oxidative stress leads to cascade of events resulting to degeneration of neurons and eventually cell death. Memantine prevented A-induced memory impairment in rats that received bilateral microinjections of aggregated A1-40 into the
(Cornu Ammonis, CA) CA1 and CA3 subfields of the rat hippocampus had been reported. Nakamura and co-workers [63] reported that subcutaneous infusion of memantine at doses of 10 and 20 mg/kg/day for 6 weeks starting 24 hours before aggregated A1-40 significantly prevented learning
29 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330
debilitating Alzheimer’s disease, excitotoxicity is considered as a contributing factor specifically on the induction of neuronal cell death [33]. The over-activation of the NMDA type glutamate receptor increases the ability of the calcium ions (Ca2+) to enter the cytosol, thus acting as one of the culprits of
cell injury, damage and even cell death (details are discussed in the review of Lipton [64]). Many studies in vitro also suggest that glutamate receptors specifically the NMDA receptors contribute to neuronal toxicity produced by the accumulation of the -amyloid peptide [65, 66].
In our present study, memantine is also demonstrated to down regulate the expression of tau that is consistent with our previous report [29]. The biological activity of tau is controlled by the degree of its phosphorylation. The abnormally hyperphosphorylated tausequesters the microtubule associated proteins (MAP-1 and MAP-2) or normal tau to resulting in the breakdown of the microtubule networks, subsequently neurofibrillary degeneration and other tauopathies may develop [67]. Studies have shown that memantine is capable of modulating the signaling pathways of the protein phosphatase -2A, an enzyme responsible for the phosphorylation of tau [68, 69].
Interestingly, no direct evidence indicated the higher activity of APP and Tau IRES result in higher release of A and tau extracellularly and intracellularly, respectively. However, Han et al., analyzed published data on AD blood transcriptome had revealed that the perturbation of cellular functional units could lead to the up regulation of cellular IRES activity [70]. This in silico study had implied that aberrant expression of APP and Tau IRES might increase the release of A and tau extracellularly and intracellularly, respectively.
Based from the results obtained in our study, memantine can be used as a potential IRES-dependent translational inhibitor since it is capable of stalling the activity of tau-IRES leading to a diminished tau protein expression. Thus, the action of memantine on AD may be contributed by its action on block of NMDA receptor-induced excitotoxicity as well as through the inhibition of IRES-mediated translation. And, our finding also offers a facile method for screening biologically important compounds that may play a significant role in arresting the development of Alzheimer’s disease. However, the precise mechanism on how memantine inhibits the IRES of both tau and APP remains elusive. These IRESes may form a specific RNA structures that operate as aptamers or molecular switches in response to the direct binding of various compounds to regulate and control gene translation. Further studies are yet to be conducted on an in-depth understanding of the mechanism of the IRES-mediated translation in the pathogenesis of AD.
4. Experimental Section
4.1. Culturing of cells, plasmids construction and transfection studies on mammalian cells
The cell lines used in the experiment were COS-1 (African green monkey kidney fibroblast-like cells, CHO (Chinese hamster ovary cells) and N2A (Mouse neuroblastoma). The COS-1 and CHO cells were grown in Dulbecco’s medium (DMEM) (Invitrogen) while the N2A cells were grown in Minimum Essential Medium (MEM) (Invitrogen) both supplied with 10% fetal bovine serum. The plasmid pUC57 (NCBI No. NM016835) containing the tau IRES DNA fragment was synthesized by PROTECH Technology Company, Taiwan. The 240 bp tau IRES was cut from pUC57 by enzymatic digestion with Not1 and cloned into the Not1 treated plasmid-pGS-EMCV [70] to replace the EMCV IRES fragment. The APP IRES DNA fragment was amplified by PCR using two pairs of primers: 32 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372
(1Forward: ATTGCGGCCGCAGTTTCCTCGGCAGCGGTAGGCGAGAGCACGCGGAGGAGCGTGCGC; 2Reverse: TCTGCCCGCGCCGCCACCGCCGCCGTCTCCCGGGGCCCCCGCGCACGCTCCTCCGCGT; 3Forward: TGGCGGCGCGGGCAGAGCAAGGACGCGGCGGATCCCADTCGCACAGCAGCGCACTC; 4Reverse: TATGCGGCCGCCGCGACCCTGCGCGGGGCACCGAGTGCGCTGCTGTGCGA). The restriction sites in Not1I are underlined. The APP IRES containing DNA fragment was digestion with Not1 and cloned into the Not1 treated plasmid-pGS-EMCV [71] to replace the EMCV IRES fragment. The plasmids were generated as pGS-APP and pGS-Tau, respectively (Fig. 1). In each plasmid, the IRES element from the genes of either APP or tau that was flanked correspondingly by the reporter genes: -galactosidase and secreted human alkaline phosphatase (SEAP). Prior to transfection, cells from the corresponding cell line were seeded onto a 24-well plate at a density ~9.0 x 104 cells/well.
The cells were washed repeatedly with serum-free medium to remove all traces of sera. Plasmids were then transfected into the respective cell line using Lipofectamine 2000 reagent (Invitrogen). Briefly, the plasmid DNA (1g) was diluted with serum-free either DMEM or MEM (50 L) then the Lipofectamine 2000 reagent (1 L) was added and allowed to form the complexation product for 15~20 minutes, followed by transfection to the respective cells and incubated at 37oC with 95% air and
5% CO2. After five hours, the transfection medium was removed, the adherent cells were washed with
PBS and then replaced with a fresh medium (with 10% FBS and antibiotics) and the corresponding memantine dosage.
4.2. IRES reporter assay
Memantine solutions were prepared at various concentrations (0.1, 1, 5, 10 and 20 M) by dilution with DMEM or MEM to treat the transfected COS-1, CHO and N2A cells. Twenty-four hours post-addition of memantine, the cell culture medium from each well was pipetted out and was kept frozen at -20 oC until used for SEAP activity measurement using BD Great EscApe SEAP detection kit
(Clonetech). Subsequently, remaining cells were lysed for 10 minutes in 120 L of lysis buffer (CytobusterTM Protein Extraction Reagent, Novagen) and was subjected to centrifugation at 12,800
rpm for 10 minutes the lysate supernatant was then assayed for -galactosidase activity using Luminiscent -Galactosidase Detection Kit II (BD Biosciences). The chemiluminescence intensity (relative light units, RLU) and was measured with Mithras LB 940 (Berthold Technologies) chemical luminescence counter.
4.3. Western Blot Analysis of the APP and tau proteins in neuronal cells
Primary hippocampal neurons were dissociated from the rat fetuses at 18th embryonic day according
to the procedures previously described [72, 73]. Inhibition of the expression of the tau protein was conducted in neuronal cells (N2A and hippocampal neurons). The hippocampal cells (5 x 104
cells/cm2) were cultured in MEM (Invitrogen) supplemented with 5% horse serum, 5% FBS, 0.5 mM
glutamine and penicillin/streptomycin (PS, all from Invitrogen) and was allowed to grow for 24 hours 35 373 374 375 376 377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399 400 401 402 403 404 405 406 407 408 409 410
cells were then lysed using CytobusterTM and were subsequently analyzed for endogenous tau protein
expression. Bicinchoninic acid assay (BCA) was done to determine the total protein concentration and protein separation was carried out with SDS-PAGE. Western blot analysis proceeded using the monoclonal antibodies (1:2500) for tau-1 clone PC1C6 (Millipore) and rabbit polyclonal antibody against APP (ab207, ABCAM Company) to detect the target proteins from the cell lysates.
4.4. Fermentation of traditional Chinese herb
All the Lactobacillus spp. used in this study were purchased from Bioresources Collection and Research Center (BCRC), Hsin Tue, Taiwan and were preserved in Lactobacilli MRS broth (DIFCO, Detroit, Michigan, USA) (-80 oC, with 15% glycerol). To prepare the products of Lactobacillus spp.
fermented Chinese herb, these Lactobacillus spp. were first activated in Lactobacilli MRS broth with 0.05% L-cysteine (100 ml, 37 oC) for 24 hours. The Chinese herbs (700 g) were added with the
activated Lactobacillus spp. and incubated in a 10 liter fermenter with 7 liter culture broth for 24 hours at 37 oC. The composition of the culture broth contained: peptone (20 g/l); yeastolate (10 g/l) and
glucose (70 g/l) in potassium phosphate buffer pH 7.4. 4.5. Preparation of the fermentation products
After the Chinese herbs including: Eleutherococcus senticosus 、Lycium chinense Miller、Panax ginseg 、 Curcuma longa 、 Radix notoginseng and Gastrodia elata were fermented with the Lactobacillus spp., the ferments were extracted with 70% alcohol for two days and filtrated twice. The filtrates were collected and concentrated with rotary evaporator and then with lyophilizer to remove remained water. The powders of these Lactobacillus spp. fermented Chinese herbs were stored in -20
oC freezer before use.
4.6. Animals
All procedures were performed according to the National Institutes of Health Guidelines for the Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee of Chung Tuan Christian University, Taoyuan, Taiwan. ApoE-/- mice with the C57BL/6 genetic background were
provided by National Taiwan University (Prof. S. W. Lin). Mice (male, 10 weeks old) were randomly divided into two groups (n =8 per group) and were fed either HFD containing 60% of kilocalories from fat (TestDiet; LabDiet Cat no. 58Y1) or HFD plus NB-34 for 12 weeks. The NB-34 group received 4 mg/kg body weight (bw)/day NB-34 via a gavage of gastric tube for 12 weeks, with dose adjustment weekly according to body weight.
4.7. Morris Water Maze (MWM) task
The spatial learning performance of the control or NB-34 intaking ApoE-/- mice was assessed in a
circular white water tank (diameter 120 cm, depth 45 cm) filled with tap water (25 ± 0.5℃). The water tank was located in a test room that contained several cues around the maze and remained unchanged during the test. Each mouse was subjected to a series trial, 4 trials per day. For each trial, the place where the mouse was put in the water differed with four different positions. If the mouse could not 38 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430 431 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447
find the platform within 60 seconds, it was guided to the platform with a sieve, and after it on the platform for 20 seconds was then put into its cage. After completing the 5 days learning of the MWM, memory recall was determined by a probe test. This probe test was performed 24 h after the acquisition, and measured the ability to consolidate spatial memories. For the acquisition trial the latency of these control or NB34 intaking mice reach the platform were recorded and compared. For the probe trial, the platform was removed from the water tank and the time of the mice swam in the four quarters were recoded and analysis using a video-tracking software (Ethovision XT 7, Noldus Information Technology, Netherlands). Water maze search strategy analysis was performed as described previously [51, 52]. The search strategies were analyzed the first of the 4 trials of Morris Water Maze for 6 days. The percentage of each strategy in each group was calculated. Swim strategies were characterized as spatial, systematic, or looping and representative strategies are provided (Figure 7C).
4.8. Data analysis
All data were presented as the means +/- SEM. Statistical comparisons were performed by paired or unpaired Student’s t-tests, and one-way analysis of variance (ANOVA) or two way ANOVA for repeated experiments followed by Fisher’s protected least significant different test. P<0.05 was considered to indicated a statistically significant difference.
Acknowledgments
This research was supported by Grants from the Conventional Industry Technology Development (E10200049-092) and the Ministry of Science and Technology (NSC 102-2632-M-033-001-MY3). Author Contributions
All authors read and approved the final manuscript; TYC, SLL and TYW designed methods and experiments, analyzed the data, and interpreted the results and wrote the manuscript. YCT, CCH and YJC carried out experiments.
Conflicts of Interest
The authors declare no conflict of interest. References
1. Goedert, M.; Spillantini, M.G. A century of Alzheimer's disease. Science 2006, 314, 777-781. 2. Roberson, E.D.; Mucke, L. 100 years and counting: prospects for defeating Alzheimer's
disease. Science 2006,314, 781-784.
3. World Alzheimer Report 2009, M. Prince and J. Jackson, Editors. 2009, Alzheimer's Disease International. 41 448 449 450 451 452 453 454 455 456 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475 476 477 478 479
197, 192-193.
5. Terry, R.; Gonatas, N.; Weiss, M. Ultrastructure studies in Alzheimer's presenile dementia. Am J Pathol 1964, 44, 269-297.
6. Zetterberg, H.; Blennow, K.; Hanse, E. Amyloid and APP as biomarkers for Alzheimer's disease. Exp Gerontol 2010, 45, 23-29.
7. Brandt, R.; Hundelt, M.; Shahani, N. Tau alteration and neuronal degeneration in tauopathies: mechanisms and models. Biochim Biophys Acta 2005, 1739, 331-354.
8. Zhang, Y.; Tian, Q.; Zhang, Q.; Zhou, X.; Liu, S.; Wang, J. Z. Hyperphosphorylation of microtubule-associated tau protein plays dual role in neurodegeneration and neuroprotection. Pathophysiology 2009, 16, 311-316.
9. Games, D.; Adams, D.; Alessandrini, R.; Barbour, R.; Borthelette, P.; Blackwell, C.; Carr, T.; Clemens, J.; Donaldson, T.; Gillespie, F.; Guido, T.; Hagopian, S.; Johnson-Wood, K.; Khan, K. Lee M Leibowitz P Lieberburg I Little S Masliah E McConlogue L Montoya-Zavala M Mucke L.; Paganini, L.; Penniman, E.; Power, M.; Schenk, D.; Seubert, P.; Snyder, B.; Soriano, F.; Tan, H.; Vitale, J.; Wadsworth, S.; Wolozin, B.; Zhao, J. Alzheimer-type neuropathology in transgenic mice overexpressing V717F [beta]-amyloid precursor protein. Nature 1995, 373, 523-527.
10. Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y .; Younkin, S.; Yang, F.; Cole, G. Correlative Memory Deficits, A Elevation, and Amyloid Plaques in Transgenic Mice. Science 1996, 274, 99-103.
11. Cirrito, J. R.; Disabato, B. M.; Restivo, J. L.; Verges, D. K.; Goebel, W. D.; Sathyan, A.; Hayreh, D.; D'Angelo, G.; Benzinger, T.; Yoon, H.; Kim, J.; Morris, J. C.; Mintun, M. A.; Sheline, Y. I. Serotonin signaling is associated with lower amyloid- levels and plaques in transgenic mice and humans. Proc Natl Acad Sci 2011, 108, 14968-14973.
12. Ando, K.; Leroy, K.; Heraud, C.; Yilmaz, Z.; Authelet, M.; Suain, V.; De Decker, R.; Brion, J. P. Accelerated Human Mutant Tau Aggregation by Knocking Out Murine Tau in a Transgenic Mouse Model. Am J Pathol 2011, 178, 803-816.
13. Pelletier, J.; Sonenberg, N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature 1988, 334, 320-325.
14. Pelletier, J.; Kaplan, G.; Racaniello, V. R.; Sonenberg, N. Cap-independent translation of poliovirus mRNA is conferred by sequence elements within the 5' non-coding region. Mol Cell Biochem 1988, 8, 1103-1112.
15. Stoneley, M.; Willis, A. E. Cellular internal ribosome entry segments: structures, trans-actinng factors and regulation of gene expression. Oncogene 2004, 23, 3200-3207.
44 481 482 483 484 485 486 487 488 489 490 491 492 493 494 495 496 497 498 499 500 501 502 503 504 505 506 507 508 509 510 511 512 513 514
16. Komar, A. A.; Hatzoglou, M. Internal Ribosome Entry Sites in Cellular mRNAs: Mystery of Their Existence. J Biol Chem 2005, 280, 23425-23428.
17. Kerry, D.; Fitzgerald, B. L. S. Bridging IRES elements in mRNAs to the eukaryotic translation apparatus. Biochim Biophys Acta 2009, 1789, 518 - 528.
18. William, C. M. Cap-dependent and cap-independent translation in eukaryotic systems. Gene 2004, 332, 1-11.
19. Laurent, B.; Ricardo, S. R.; Ricci, E. P.; Decimo, D.; Ohlmann, T. Structural and functional diversity of viral IRESes. Biochimica et Biophysica Acta 2009, 1789, 542-557.
20. Qin, X.; Sarnow, P. Preferential translation of internal ribosome entry site-containing mRNAs during the mitotic cycle in mammalian cells. J Biol Chem 2004, 279, 13721-13728.
21. Veo, B. L.; Krushel, L. A. Translation Initiation of the Human Tau mRNA Through an Internal Ribosomal Entry Site. J Alzheimers Dis 2009, 16, 271-275.
22. Danysz, W.; Parsons, C. G.; Konhuber, J.; Schmidt, W. J.; Quack, G. Aminoadamantanes as NMDA Receptor Antagonists and Antiparkinsonian Agents-Preclinical Studies. Neurosci Biobehav Rev 1997, 21, 455-468.
23. Lipton, S. Failures and successes of NMDA receptor antagonists: Molecular basis for the use of open-channel blockers like memantine in the treatment of acute and chronic neurologic insults. Neurotherapeutics 2004, 1, 101-110.
24. Chen, H. S. V.; Lipton, S. A. The chemical biology of clinically tolerated NMDA receptor antagonists. J Neurochem 2006, 97, 1611-1626.
25. Gilling, K. E.; Jatzke, C.; Hechenberger, M.; Parsons, C. G. Potency, voltage-dependency, agonist concentration-dependency, blocking kinetics and partial untrapping of the uncompetitive N-methyl-d-aspartate (NMDA) channel blocker memantine at human NMDA (GluN1/GluN2A) receptors. Neuropharmacology 2009, 56, 866-875.
26. Reus, G. Z.; Stringari, R. B.; Kirsch, T. R.; Fries, G. R.; Kapczinski, F.; Roesler, R.; Quevedo, J. Neurochemical and behavioural effects of acute and chronic memantine administration in rats: Further support for NMDA as a new pharmacological target for the treatment of depression? Brain Res Bull 2011, 81, 585-589.
27. Li, L.; Sengupta, A.; Haque, N.; Iqbal-Grundke, I.; Iqbal, K. Memantine inhibits and reverses the Alzheimer's type abnormal hyperphosphorylation of tau and associated neurodegeneration. FEBS Letters 2004, 566, 261-269.
28. Floden, A.; Li, S.; Combs, C. K. -Amyloid-stimulated microglia induce neuron death via synergistic stimulation of tumor necrosis factor and NMDA receptors. J Neurosci 2005, 25, 47 515 516 517 518 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547
29. Wu, T. Y.; Chen, C. P. Dual action of memantine in Alzheimer's disease: A hypothesis. Taiwan J Obstet Gynecol 2009, 48, 273-277.
30. Chen, Y.J.; Hsu, J.T.; Horng, J. T.; Yang, H. M.; Shih, S. R.; Chu, Y. T.; Wu, T.Y. Amantadine as regulator of intenal ribosome entry site. Act Pharmacol Sin 2008, 29, 1327-1333.
31. Borman, A. M.; Le Mercier, P.; Girard, M.; Kean, K. M. Comparison of Picornaviral IRES-Driven internal initiation of translation in cultured cells of different origins. Nucleic Acids Res 1997, 25, 925-932.
32. Nothias, F.; Boyne, L.; Murray, M.; Tessler, A.; Fischer, I. The expression and distribution of tau proteins and messenger RNA in rat dorsal root ganglion neurons during development and regeneration. Neuroscience 1995, 66, 707-719.
33. Danysz, W.; Parsons, C. G. The NMDA receptor antagonist memantine as a symptomatological and neuroprotective treatment for Alzheimer's disease:preclinical evidence. Int J Geriatr Psychiatry 2003, 18, S23-S32.
34. Rogawski, M. A.; Wenk, G. L. The Neuropharmacological Basis for the Use of Memantine in the Treatment of Alzheimer's Disease. CNS Drug Rev 2003, 9, 275-308.
35. Johnson, J. W.; Kotermanski, S. E. Mechanism of action of memantine. Curr Opin Pharmacol 2006, 6, 61-67.
36. Takashima, A. Amyloid-, Tau and Dementia. J Alzheimers Dis 2009, 17, 729 - 736.
37. Alley, G. M.; Bailey, J. A.; Chen, D.; Ray, B.; Puli, L. K.; Tanila, H.; Banerjee, P. K.; Lahiri, D. K. Memantine lowers amyloid-β peptide levels in neuronal cultures and in APP/PS1 transgenic mice. J Neurosci Res 2010, 88, 143-154.
38. Seeman, P.; Caruso, C.; Lasaga, M. Memantine agonist action at dopamine D2High receptors.
Synapse 2008, 62, 149 - 153.
39. Hanes, J.; Zilka, N.; Bartkova, M.; Caletkova, M.; Dobrota, D.; Novak, M. Rat tau proteome consists of six tau isoforms: implication for animal models of human tauopathies. J Neurochem 2009, 108, 1167-1176.
40. Deshpande, A.; Win, K. M.; Busciglio, J. Tau isoform expression and regulation in human cortical neurons. The FASEB Journal 2008, 22, 2357-2367.
41. Combs, C. K.; Coleman, P. D.; O'Banion, M. K. Developmental regulation and PKC dependence of Alzheimer's-type tau phosphorylations in cultured fetal rat hippocampal neurons. Dev Brain Res 1998, 107, 143 - 158.
42. Taku, H.; Arawaka, S.; Mori, H. Isoforms changes of tau protein during development in various species. Dev Brain Res 2003, 142, 121-127.
50 549 550 551 552 553 554 555 556 557 558 559 560 561 562 563 564 565 566 567 568 569 570 571 572 573 574 575 576 577 578 579 580 581 582
43. Ray, B.; Banerjee, P. K.; Greig, N. H.; Lahiri, D. K. Memantine treatment decreases levels of secreted Alzheimer's amyloid precursor protein (APP) and amyloid beta (A) peptide in the human neuroblastoma cells. Neurosci Lett 2010, 470, 1-5.
44. Wu, T. Y.; Chen, C. P.; Jinn, T. R. Traditional Chinese medicines and Alzheimer's disease. Taiwan J Obstet Gynecol 2011, 50, 131-5.
45. Choi, R. C.; Zhu, J. T.; Leung, K. W.; Chu, G. K.; Xie, H. Q.; Chen, V. P.; Zheng, K. Y.; Lau, D. T.; Dong, T. T.; Chow, P. C.; Han, Y. F.; Wang, Z. T.; Tsim, K. W. A flavonol glycoside, isolated from roots of Panax notoginseng, reduces amyloid-beta-induced neurotoxicity in cultured neurons: signaling transduction and drug development for Alzheimer's disease. J Alzheimers Dis 2010, 19, 795-811.
46. Corder, E. H.; Saunders, A. M.; Strittmatter, W. J.; Schmechel, D. E.; Gaskell, P. C.; Small, G. W.; Roses, A. D.; Haines, J. L.; Pericak-Vance, M. A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer's disease in late onset families. Science 1993, 261, 921-3. 47. Saunders, A. M.; Schmader, K.; Breitner, J. C.; Benson, M. D.; Brown, W. T.; Goldfarb, L.;
Goldgaber, D.; Manwaring, M. G.; Szymanski, M. H.; McCown, N. et al. Apolipoprotein E epsilon 4 allele distributions in late-onset Alzheimer's disease and in other amyloid-forming diseases. Lancet 1993, 342, 710-1.
48. Krugers, H. J.; Mulder, M.; Korf, J.; Havekes, L.; de Kloet, E. R.; Joëls, M. Altered synaptic plasticity in hippocampal CA1 area of apolipoprotein E deficient mice. Neuroreport 1997, 8, 2505-10.
49. Veinbergs, I.; Masliah, E. Synaptic alterations in apolipoprotein E knockout mice. Neuroscience 1999, 91, 401-3.
50. Masliah, E.; Mallory, M.; Ge, N.; Alford, M.; Veinbergs, I.; Roses, A. D. Neurodegeneration in the central nervous system of apoE-deficient mice. Exp Neurol 1995, 136, 107-22.
51. Wu, J.; Zhao, Z.; Sabirzhanov, B. X.; Stoica, B. A.; Kumar, A.; Luo ,T.; Skovira, J.; Faden, A. I. Spinal cord injury causes brain inflammation associated with cognitive and affective changes: role of cell cycle pathways. J Neurosci 2014, 34, 10989-11006.
52. Zhao, Z.; Loane, D. J.; Murray, M. G.; Stoica, B. A.; Faden, A. I. Comparing the predictive value of multiple cognitive, Affective, and motor tasks after rodent traumatic brain injury. J Neurotrauma 2012, 29, 2475-2489.
53. Brody, D. L.; Holtzman, D. M. Morris water maze search strategy analysis in PDAPP mice before and after experimental traumatic brain injury. Exp Neurol 2006, 197, 330-340.
54. Domier, L. L.; McCoppin, N. K. In vivo activity of Rhopalosiphum padi virus internal 53 583 584 585 586 587 588 589 590 591 592 593 594 595 596 597 598 599 600 601 602 603 604 605 606 607 608 609 610 611 612 613 614 615
55. Chen, Y. J.; Chen, W. S.; Wu, T. Y. Development of a bi-cistronic baculovirus expression vector by the Rhopalosiphum padi virus 5' internal ribosome entry site. Biochem Biophys Res Com 2005, 335, 616-23.
56. Gardiol, A.; Racca, C.; Triller, A. Dendritic and postsynaptic protein synthetic machinery. J Neurosci 1999, 19, 168-79.
57. Tiedge, H. Brosius, J. Translational machinery in dendrites of hippocampal neurons in culture. J Neurosci 1996, 16, 7171-81.
58. Steward, O.; Levy, W. B. Preferential localization of polyribosomes under the base of dendritic spines in granule cells of the dentate gyrus. J Neurosci 1982, 2, 284-91.
59. Beaudoin, M. E.; Poirel, V. J.; Krushel, L. A. Regulating amyloid precursor protein synthesis through an internal ribosomal entry site. Nucleic Acids Res 2008, 36, 6835-6847.
60. Aronov, S.; Aranda, G.; Behar, L.; Ginzburg, I. Axonal tau mRNA localization coincides with tau protein in living neuronal cells and depends on axonal targeting signal. J Neurosci 2001, 21, 6577-6587.
61. Jackson, C. E.; Snyder, P. J. Electroencephalography and event-related potentials as biomarkers of mild cognitive impairment and mild Alzheimer's disease. Alzheimer's and Dementia 2008, 4, S137-S143.
62. Petersen, R. B.; Nunomura, A.; Lee, H. G.; Casadesus, G.; Perry, G.; Smith, M. A.; Zhu, X. Signal Transduction Cascades Associated with Oxidative Stress in Alzheimer's Disease. J Alzheimers Dis 2007, 11, 143-152.
63. Nakamura, S.; Murayama, N.; Noshita, T.; Katsuragi, R.; Ohno, T. Cognitive dysfunction induced by sequential injection of amyloid- and ibotenate into the bilateral hippocampus; protection by memantine and MK-801. Eur J Pharmacol 2006, 548, 115-122.
64. Lipton, S. A. The molecular basis of memantine action in Alzheimer's Disease and other neurologic disorders: Low affinity, uncompetitive antagonism. Current Alzheimer Research 2005, 2, 155-165.
65. Qiu, Z.; Gruol, D. L. Interleukin-6, -amyloid peptide and NMDA interactions in rat cortical neurons. J Neuroimmunol 2003, 139, 51-57.
66. Koh, J. Y.; Yang, L. L.; Cotman, C. W. -Amyloid protein increases the vulnerability of cultured cortical neurons to excitotoxic damage. Brain Research 1990, 533, 315-320.
67. Alonso, A. C.; Grundke-Iqbal, I.; Barra, H. S.; Iqbal, K. Abnormal phosphorylation of tau and the mechanism of Alzheimer neurofibrillary degeneration: Sequestration of microtubule-associated proteins 1 and 2 and the disassembly of microtubules by the abnormal tau. Proc Natl Acad Sci 1997, 94, 298-303.
56 617 618 619 620 621 622 623 624 625 626 627 628 629 630 631 632 633 634 635 636 637 638 639 640 641 642 643 644 645 646 647 648 649 650
68. Chohan, M. O.; Khatoon, S.; Iqbal, I. G.; Iqbal, K. Involvement of in the abnormal hyperphosphorylation of tau and its reversal by Memantine. FEBS Letters 2006, 580, 3973-3979.
69. Li, L.; Sengupta, A.; Haque, N.; Grundke-Iqbal, I.; Iqbal, K. Memantine inhibits and reverses the Alzheimer type abnormal hyperphosphorylation of tau and associated neurodegeneration. FEBS Letters 2004, 566, 261-269.
70. Han, G.; Wang, J.; Zeng, F.; Feng, X.; Yu, J.; Cao, H.Y.; Yi, X.; Zhou, H.; Jin, L.W.; Duan, Y.; Wang, Y.J.; Lei, H. Characteristic Transformation of Blood Transcriptome in Alzheimer's Disease. J Alzheimers Dis 2013, 35, 373-386.
71. Lee, J. C.; Wu, T. Y.; Huang, C. F.; Yang, F. M.; Shih, S. R.; Hsu, J. T.A. High-efficiency protein expression mediated by enterovirus 71 internal ribosome entry site. Biotechnol Bioeng 2005, 90, 656-662.
72. Brewer, G. J.; Torricelli, J. R.; Evege, E. K.; Price, P. J. Optimized survival of hippocampal neurons in B27-supplemented Neurobasal, a new serum-free medium combination. J Neurosci Res 1993, 35, 567-76.
73. Cheng, H. H.; Huang, Z. H.; Lin, W. H.; Chow, W. Y. Chang, Y.C. Cold-induced exodus of postsynaptic proteins from dendritic spines. J Neurosci Res 2009, 87, 460-9.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/). 59 651 652 653 654 655 656 657 658 659 660 661 662 663 664 665 666 667 668 669 670