行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 尋找與人類肝癌有關的抑癌基因之研究 ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:■個別型計畫
□整合型計畫
計畫編號:NSC90-2323-B-002-008-
執行期間:九十年八月一日至九十一年七月三十一日
執行單位:國立台灣大學醫學院內科
計畫主持人:許金川
共同主持人:
中
華
民
國
91 年
10
月
21
日
年度研究成果報告內容
中文摘要
肝細胞癌是全世界最好發的癌症之一,在台灣也是國人癌症死因的第一位,目前真正的 致病機轉仍不清楚。肝癌的發生通常伴隨基因的變化,其中包括致癌基因的活化、或癌抑制基 因的不活化。目前已知有多種人類腫瘤均與癌抑制基因之變異有關。近年來利用染色體中的微 小衛星可以有系統而快速的分析比較每一染色體中基因的變化,更可進一步尋找可能的癌抑制 基因。 我們的研究結果顯示肝癌病患在 16q 常有雜合子丟失(LOH)的情形,且細微定位的結果 靠近 16q12.1,16q21-22 和 16q24.3。因此表示這些位置可能有癌抑制基因的存在。目前,我們 利用位於 16q12.1 的標記(D16S415, D16S419, D16S409, D16S3080及 D16S3034) 開始篩選人類 之細菌人工染色體基因庫(Bacterial Artificial Chromosome Library, BAC Library),並配合表現 序列捕捉系統(exon trapping system)找到表現序列的 clones。總共選殖到 15 個可能的表現序 列,與 NCBI ( national center for Biotechnology Infornation )的資料庫比對的結果,有二個序列 與最近被發現的 KIAA1005 基因序列完全相同。而 KIAA 1005 基因在肝癌組織中有同源性丟 失的現象,反轉錄-聚 合 連 鎖反應(RT-PCR)的結果也顯示 mRNA 的表現有差異,因此我們 推測 KIAA1005 基因可能是位於 16q12.1 附近的假想抑癌基因。我們進一步分析所選殖基因的 功能發現它具有抑制肝癌細胞生長。另一方面我們利用微陣列晶片分析肝癌,找到一百多個 在肝癌細胞失去調控的基因將進一步分析。Abstr act
Key words: hepatocellular carcinoma, micorsatellite analysis, tumor suppressor gene
Hepatocellular carcinoma (HCC) is one of the most common cancer in the world and is the leading cause of cancer death in Taiwan. The prognosis of this cancer is extremely poor with survival of only several months after symptoms occurred. Elucidation of the basic genetic changes of HCC is important for the understanding and treatment of this cancer. Cancer is usually accompanied with genetic alternations either through the activation of cellular oncogene or the inactivation of cancer suppressor gene. The recently identified short tandem repeat, the microsatellite, which is widely distributed throughout the whole human genome. Identification of disease genes as well as tumor suppressor genes by microsatellite polymorphism. have been published recently.
In this study, we have used 35 microsatellite markers for further fine mapping of LOH. We have confirmed the most frequent regions of LOH for HCC are 16q12.1, 16q22, and 16q24. After analyzing these information, we started to screen the human BAC(Bacterial Artificial chromosome)library by these markers at 16q12.1 and we identified 15 clones. Exon trapping system is used to search the putative exon sequences of BAC genomic clones. Two exon-like sequences are identical to the KIAA1005 gene. Homozygous deletion of KIAA1005 was found in 37%(10 /27) HCCs. These data suggested that the KIAA1005 might be the putative tumor suppressor genes at chromosome 16q12.1.
To identify genes involved in the development of familial HCC, we analyzed the transcription profiles of six familial HCC specimens using Human Genome GEM 1 cDNA microarray consisting 9766 elements. Comparison of expression patterns between HCC tumors and the corresponding nontumor tissues made us to identify genes that were commonly upregulated (72 genes) and genes that were downregulated (47 genes) in the tumor cells. Hierarchical clustering analysis of the six familial HCC cases with 119 gene expression ratios demonstrated two distinct expression patterns and might reflect tumors evolved from different liver status. Furthermore, we used laser capture microdissection (LCM) combined with real-time quantitative polymerase chain reaction (RT-QPCR) to confirm microarray data and screen laminin-binding protein and afamin expression levels in two panels of HCC tissues. The expression profiles of 6 familial HCC provided valuable information for elucidation of hepatocarcinogenesis, and revealed the potential biomarkers implicated in this disease.
一、計畫緣起。
Hepatocellular carcinoma (HCC) is the leading cause of cancer death in Taiwan. Chronic hepatitis B and recently the hepatitis C viral infection are thought related to the development of HCC. However, the basic molecular mechanism remained to be clarified. Recent studies have suggested that accumulation of these genetic changes, which affect the expression of oncogenes and tumor suppressor genes, occur in a stepwise manner and during the development of HCC. Many tumor suppressor genes are inactivated by intragenic mutations in one allele accompanied by the loss of a chromosomal region containing the other allele, termed loss of heterozygosity (LOH). Mapping of homozygous deletions within regions showing a high frequency of LOH has been a critical step in the discovery of several tumor suppressor genes.
In the past years, we have using 231 microsatellite markers to study the LOH of HCC. We included 30 cases of HCC and we have found the most common site of LOH for HCC is 16q(43.3%), 13q(36.7%), 17p(30%), 5q(20%), 11p(20%), 11q(16.7%). These data provides as basis for further fine mapping of the common site of LOH. In our previous project, we further study the LOH of HCC by narrowing down the common site of LOH on chromosome 16q. The most frequent regions of LOH for HCC are 16q12.1, 16q22, and 16q24. Fifteen clones were identified by screening the human BAC (Bacterial Artificial chromosome) by microsatellite markers at 16q12.1. Exon trapping system is used to search the putative exon sequences of BAC genomic clones. One putative tumor suppressor gene on chromosome 16q12.1 is found.
In this proposal, We will prepare the tissue sample by LCM and extract the DNA or RNA. Then, we will apply microarray system to monitor the expression of ESTs and search the putative tumor suppressor genes on chromosome 16q (16q12.1, 16q22, and 16q24) in HCC patients.
Furthermore, we hope that functional assay of putative tumor suppressor genes in vitro will be
established. Specific genes that are differentially expressed between tumor and non-tumor part will be analyzed to elucidate gene function and their role in hepatocarcinogenesis.
for prevention and improvement of treatment. We hope through this project, we can have a better understanding of the genetic change of HCC and wishing to identify possible tumor suppressor genes of this cancer.。
二、計畫目的
Specific aims
l
Objectives
(long term):
1. Improvement of liver cancer treatment. 2. As cancer diagnostic marker
l Focus
(short term)1. ESTs on chromosome 16q will be analyzed between non-tumor parts and tumor parts by microarray. Deletion mapping, BAC library screening, exon trapping system will be applied to analysis the other chromosomes regions with high frequency LOH.(first year)
2. Functional assay of candidate genes will be established.(second year)
Results
Deletion mapping of chromosome 16q and characterization of the interest region
Eighty-eight tumor samples derived from 88 HCC patients were subjected to analysis of LOH in 35 microsatellite loci on chromosome 16q (Table3). LOH at one or more loci was observed in 59 (67%) of these tumors. Seven independent regions of frequent LOH were defined in these 59 tumors
with partial deletions. The first region was between D16S517 and D16S409 loci, encompassed by a
3-cM region. The second region was between D16S419 and D16S415, encompassed approximately
by a 0.5-cM region. The third region was between D16S496 and D16S514, encompassed
approximately by a 5-cM region. The fourth region was between D16S512 and D16S515,
encompassed approximately by a 3-cM region. The fifth region was near D16S504, encompassed
approximately by a 5-cM region. The sixth region was between D16S505 and D16S422,
encompassed approximately by a 5.5-cM region. The seventh region was near D16S3048,
encompassed approximately by a 4-cM region.
Deletion mapping showed a high frequency of LOH within the area of interest (16q12.1) affecting 20-30% of the HCCs and 5 tumor cases with a homozygous deletion at D16S415. We followed one main approach to characterize the chromosomal region of interest to isolate the putative tumor suppressor genes. After building BAC contig spanning the D16S415 region, we used exon-trapping system to search putative exon sequences in these BACs. Two exon-like sequences were identified and matched to the UniGene AB023222 (KIAA1005 partial CDS).
Isolation and Characterization of KIAA1005
Using available information about AB023222, we recovered a 4.5-kb fragment after RT-PCR with EST primers from normal tissue. To clone full-length cDNA, we used the 5’ and 3’ rapid
amplification of cDNA ends PCR method. Three different clones were identified and sequenced. One clone has additional N-terminal extra 550 nucleotides of KIAA1005 (4.5kb) and the other two clones were 2.4kb and 3.4kb, respectively (Figure 4). Recently, NCBI annotation project also identified three different splicing forms of KIAA1005. The sequences are the same as our results. The 2.4kb cDNA showed a consensus sequences of 2426 bp long with a predicted ORF of 1595 bp, a 206-bp-long 5’UTR, and a 3’ UTR 625 bp long (GenBank accessin no. XM_051197). The putative start ATG codon is located with a adequate Kozak sequence (CCCatgg), which contains a highly conserved G residue (position +4) and pyrimidine (Y residue at position –3; ref14). The three cDNAs were compared with the genomic DNA sequence to identify evidence exon-intron structure. Only the 2.4kb cDNA showed the features and contains 12 exons. However, the two cDNAs (5kb and 3.4 kb) had the exon-intron structure but the sequences were split to two regions on chromosome 16q12 (about 1.3 cM). Some of these sequences were not confirmed.
Northern blot analysis with a probe derived from 1kb of KIAA1005 revealed four bands of mRNA (Figure 4 )(i.e., 1.6kb, 2.4kb, 3.4kb, and 5kb, respectively). The major band is 2.4kb. Analysis of the KIAA1005 expression pattern in normal human tissues showed that expression was highest in heart, kidney, spleen, and significantly lower in the other examined tissues (i.e. muscle, liver, and small intestine).
To examine whether the translation of the ORF of KIAA1005 (2.4kb form) produced the predicted
protein, we used an in vitro transcription-translation system. SDS-PAGE analysis of the translated
product revealed a single protein product of about 82kDa. This agree with the predicted molecular mass of KIAA based on its amino acid sequence (i.e., Da)
Homozygous deletion of KIAA1005
We also analyzed the expression of KIAA1005 (2.3kb) in HCC cell lines and HCC tissues by RT-PCR. The KIAA1005 were not expressed in some of the HCC cell lines and tissues . Homozygous deletion of KIAA1005 was found in 37% (10/27) HCCs. In the remaining specimens that showed specific band, the band intensity was almost the as that of the non-tumor part. These data suggest that the KIAA1005 might be the putative tumor suppressor gene at chromosome 16q12.1. By Southern blot analysis, homozygous deletion of KIAA1005 was confirmed in 10 tumors of the 27 HCCs.
KIAA1005 Protein Structure
The 1596-bp KIAA1005 ORF encoded for a 532 amino acid protein. The BLAST and PSIBLAST algorithms were used to search for matches in GenBank databases. Interesting, the N-terminus of the putative contains a C2 domain.
Overexpression of KIAA1005 in hepatoma
We observed the significant suppression of growth in Hep3B cells after introduction of the KIAA1005 gene(Figure 5).
Table 1 Clinical characterization of 6 familial HCC patients (consisted of 3 different families) studied by cDNA microarray
Case no. Gender Age HBsAg Anti-HCV AFP Tumor grade Tumor size Liver status
1a M 63 + - 40.5 3 5.0×4.5×3.0 cm cirrhosis
2a M 59 + - 139 clear cell 2.6×2.0×1.0 cm cirrhosis
3b M 46 + - 426.9 4 8.0×3.0×3.0 cm cirrhosis
4b M 42 + - 129.8 2 5.2×3.8×3.0 cm fatty change and CPH
5c M 53 + - 1370 3 8.0×7.5×6.5 cm portal fibrosis and PVT
6c M 27 + - >70000 3 16×9.0×9.0 cm portal lymphocyte infiltration
M, male; HBsAg, hepatitis B surface antigen; Anti-HCV, hepatitis C virus antibody; AFP, α-fetoprotein; CPH, chronic persistent hepatitis; PVT, portal vein
7
-Table 2. Representative list of candidate genes that expressed up-regulated in HCC by microarray
Human RNA for NADH dehydrogenase. Human moesin mRNA, complete cds.
Human cDNA: FLJ21171 fis, clone CAS10962.
Human IL-17B receptor (IL17BR) mRNA, complete cds. Human mRNA for titin protein (clone hsk1-hsk19).
Human cytoplasmic dynein light chain 1 (hdlc1) mRNA, complete cds. Human S19 ribosomal protein mRNA, complete cds.
Human gene for phosphate carrier.
Human ADP-ribosylation factor 1 gene, exons 2-5.
Human (chromosome X) glypican (GPC3) mRNA, complete cds. Human HALPHA44 gene for alpha-tubulin, exons 1-3.
Human mRNA for putative p64 CLCP protein.
Human 1-8D gene from interferon-inducible gene family. Human HSPC224 mRNA, complete cds.
Human CpG island DNA genomic Mse1 fragment, clone 97h11, reverse read cpg97h11.rt1a.
Human Cctg mRNA for chaperonin.
Human myelin basic protein (MBP) mRNA, complete cds. Sequence 28 from Patent WO9954448.
Human liver glucose transporter-like protein (GLUT2), complete cds. Human protein phosphatase 2A B56-delta (PP2A) mRNA, complete cds. Human mRNA for polyA binding protein.
Human ubiquitin gene.
Human DNA for ribosomal protein S13, complete cds, U14 small nucleolar RNA, complete sequence.
Human mRNA for KIAA1293 gene, complete cds.
Human mitochondrial carrier homolog 1 isoform a mRNA, complete cds; nuclear gene for mitochondrial product.
Human ADP-ribosylation factor 1 gene, exons 2-5. Incyte EST
Human 90-kDa heat-shock protein gene, cDNA, complete cds. Human mRNA for cytochrome oxidase subunit VIb.
Human cDNA: FLJ22916 fis, clone KAT06406, highly similar to HSCYCR Human mRNA for T-cell cyclophilin.
Human sin3 associated polypeptide (SAP18) mRNA, complete cds. Human mRNA for diubiquitin.
Human cDNA: FLJ22980 fis, clone KAT11387, highly similar to HSCYC1 Human mRNA for cytochrome c1.
Human mRNA for non-muscle type cofilin. Human mRNA for lipocortin II, complete cds.
8
-Human lysosomal membrane glycoprotein CD63 mRNA.
Human MHC class I HLA-C-alpha-2 chain and alternative mRNA, complete cds, clones 4 and 10.
Human genomic DNA, chromosome 21q, section 37/105. Human nuclear protein SDK3 mRNA, complete cds. Human PRO1608 mRNA, complete cds.
Human flotillin-1 mRNA, complete cds.
Human hum-a-tub2 alpha-tubulin mRNA, complete cds. Human mRNA for myosin regulatory light chain.
Human mRNA for KIAA0703 protein, complete cds.
Human colon carcinoma laminin-binding protein mRNA, complete cds. Human mRNA for Nop10p, complete cds.
Human cDNA: FLJ23506 fis, clone LNG03055. Human sorcin CP-22 mRNA, complete cds.
Human nonmuscle/smooth muscle alkali myosin light chain gene, complete cds. Human ADP/ATP carrier protein mRNA, complete cds.
Human mRNA for ribosomal protein S18.
Human cDNA: FLJ22915 fis, clone KAT06354, highly similar to Human Wilm's tumor-related protein (QM) mRNA.
Human profilin mRNA, complete cds.
Human FK506-binding protein (FKBP) mRNA, complete cds. Human full length insert cDNA clone YB24D08
Table 3. Representative list of candidate genes that expressed down-regulated in by microarray
Human mineralocorticoid receptor (MLR), exon 8. Incyte EST
Incyte EST
Human aldehyde oxidase (hAOX) mRNA, complete cds.
Human flavin-containing monooxygenase 3 (FMO3) gene, exon 7. Human cytochrome P450IIE1 (ethanol-inducible) gene, complete cds. Human afamin mRNA, complete cds.
Human 4q13 genomic sequence.
Human nicotinamide N-methyltransferase gene, exon 1 and 5' flanking region. Human HSPC062 mRNA, complete cds.
Sequence 13 from Patent WO9951727.
cytochrome P450 2C [Macaca fascicularis=monkeys, liver, mRNA, 1901 nt]. Sequence 92 from Patent WO9955858.
RhoE=26 kda GTPase homolog [Human, HeLa cell line, mRNA, 833 nt].
Human class II alcohol dehydrogenase (ADH4) pi subunit mRNA, complete cds. Human histidine-rich glycoprotein mRNA, complete cds.
9
-Human cytochrome P450IIA3 (CYP2A3) mRNA, complete cds. Human mRNA for thrombospondin.
Human 11-beta-hydroxysteroid dehydrogenase (HSD11) gene, exon 6. Human mRNA for hBD-1 protein.
Human cytokine (GRO-beta) mRNA, complete cds. Sequence 12 from Patent WO9954460.
Human pre-B-cell stimulating factor homologue (SDF1b) mRNA, complete cds. Human fetal liver cytochrome P450 (P-450 HFLa), complete cds.
Human mRNA for organic cation transporter, liver.
Human putative glycine-N-acyltransferase mRNA, complete cds. Human coagulation factor IX mRNA, complete cds.
Table 4. Upregulation of laminin-binding protein (LBP) gene and downregulation of afamin (AFM) gene identified in GEM1 microarray screen was independently tested using RT-QPCR analysis. All RT-QPCR reactions were performed in duplicate each experiment, and average expression ratio of two independent experiments was represented. We use an expression ratio to represent target gene mRNA quantities differences in tumor over nontumor cells. The cases from 1 to 10 are HCC with familial history, and those from 11 to 20 are nonfamilial cases.
Case no. LBP/RT-QPCR (T/NT) AFM/RT-QPCR (NT/T) 1 1.2 1.54 2 0.51 1.86 3 0.83 deleted 4 2.47 1.23 5 1.48 1.43 6 4.17 8.65 7 2.26 2.82 8 15.06 13.36 9 0.6 6.41 10 0.52 0.95 11 0.88 2.91 12 1.43 10.09 13 2.37 6.29 14 7.7 1.08 15 0.79 2.47 16 0.64 1.56 17 2.13 3.5 18 0.97 1.6 19 1.26 7.05 20 6.1 10.25
10
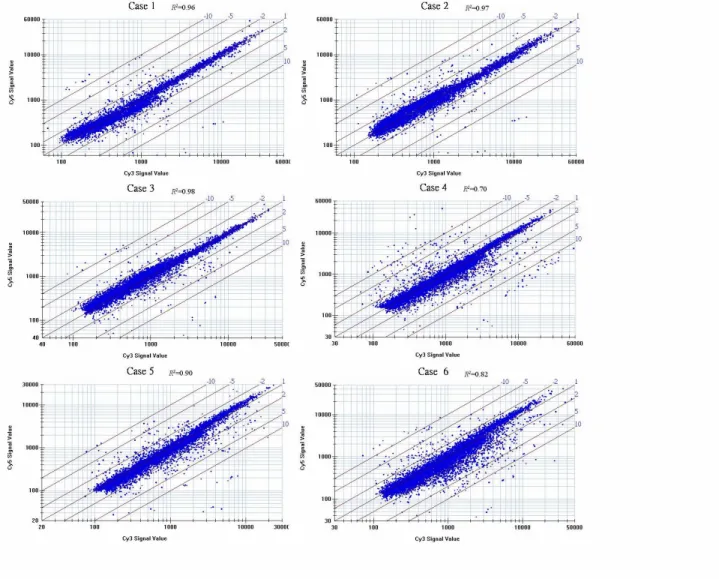
-Figur e 1 Scatterplot analysis of 6 cDNA microarray experiments. Each case of tumor mRNA
was labeled with with Cy3 and nontumor mRNA with Cy5. The graphs are presented in a log
11
-(A) laminin-binding protein
0 5 10 15 20 0 5 10 15 20
RT-QPCR fold change (tumor/nontumor) cDNA microarray ratio (tumor/nontumor)
(B) afamin 0 2 4 6 8 10 12 14 16 0 2 4 6 8 10
RT-QPCR fold change (nontumor/tumor) cDNA microarray ratio (nontumor/tumor)
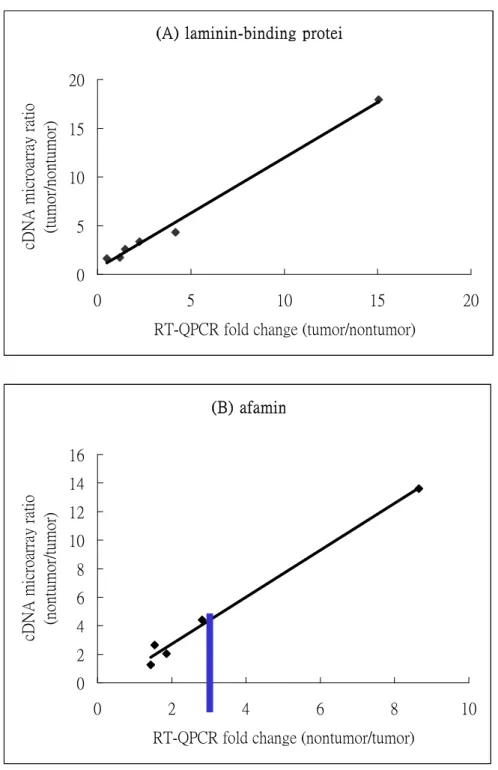
Figur e 3 Comparison of the relative fold-change of two gene expressions by cDNA microarray and
RT-QPCR. The Cy3/Cy5 ratio detected by cDNA microarray represents the tumor/nontumor tissue expression of mRNA. The values of relative quantification of target mRNA by RT-QPCR are the averages of results from two experiments. (A) laminin-binding protein. (B) afamin. The negative
values are due to down-regulated in tumor tissues. Highly correlation of coefficient (R2) of 0.99 was
obtained in laminin-binding protein gene test. The same R2 value was also observed in afamin gene
12
-Table 5. Loss of heterozygosity at 35 microsatellite loci on chromosome 16q in HCCs
Locus Symbol
Chromosomal
location (LOH/informative cases) Frequency of LOH(%) D16S409 16q11-12 14/44 31.8 D16S517 16q11.2 16/56 28.6 D16S261 16q12.1 4/79 5 D16S3044 16q12.1 2/84 2.5 D16S3080 16q12.1 19/77 24.7 D16S416 16q12.1 3/43 7 D16S419 16q12.1 15/64 23.4 D16S415 16q12.1-q13 10/44 22.7 D16S390 16q12.1-q13 16/71 22.5 D16S3034 16q12.1-q13 13/44 30.2 D16S3137 16q12.1-q13 18/77 23.4 D16S3053 16q12.1-q13 10/68 14.7 D16S3112 16q13 14/80 17.5 D16S3039 16q13 18/60 30 D16S408 16q12.1-21 10/56 17.9 D16S494 16q21 10/67 14.9 D16S514 16q21 16/66 24.2 D16S496 16q22.1 17/42 40.5 D16S398 16q22.1 9/68 13.2 D16S397 16q22.1 5/68 7.35 D16S512 16q22.2 10/67 14.9 D16S3018 16q22.2 19/79 24 D16S515 16q22.3 15/73 20.5 D16S266 16q23.1 11/62 17.7 D16S516 16q24.1 9/69 13.04 D16S504 16q24.1 20/59 33.9 D16S507 16q24.1 12/78 15.4 D16S505 16q24.1 5/71 7.04 HSD17B2 16q24.1-24.2 13/58 22.4 D16S422 16q24.2 9/49 18.4 D16S402 16q24.2 10/64 15.6 D16S3037 16q24.3 6/54 11.1 D16S520 16q24.3 7/79 8.9 D16S3048 16q24.3 16/78 20.5 D16S413 16q24.3 6/34 17.6
13 -MTT ASSAY 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0 1 2 3 4 5 6 DAY OD490 3B 1501 , pool 2162 , pool
1 2 3 4 5 6 7 8 9 10 11 12
7.5 kb 4.4 kb 2.4 kb 1.35 kb 2.4 kb 1.35 kbactin
KIAA1005
Figure 4. Northern blot of KIAA1005 in various tissue