行政院國家科學委員會專題研究計畫 成果報告
由雄性激素受體輔助因子在前列腺癌細胞之表現差異來探
討荷爾蒙不依賴性之機轉
計畫類別: 個別型計畫 計畫編號: NSC91-2314-B-002-386- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立臺灣大學醫學院泌尿科 計畫主持人: 張宏江 計畫參與人員: 謝汝敦,陳世乾 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 92 年 10 月 30 日
一、中文摘要
截斷雄性激素能相當有效的抑制前列腺癌細胞之生長。雖然荷爾蒙治療相當有效,但會 漸漸的失去對荷爾蒙治療之感受性。為何前列腺癌細胞會由荷爾蒙依賴型轉成荷爾蒙不 依賴型?現有許多假說及實驗證據解釋其原因。在這其中,以 AR 的輔助因子(cofactor) 最受重視。許多分子生物學的實驗證據顯示在雄性激素相當低量微弱下,AR 的 cofactor 可以增強androgen 與其 receptor 接合後啟動下游基因之生物效應、或甚至在 cofactor 存
在下使estrogen 也能啟動 AR 下游基因之生物效應.第一個被發現的雄性激素受體的輔助
因子是1996 年的 ARA70。基於 AR cofactor 的特別功能,新的 AR 調節的轉錄途徑因此
被發現.在 ARA70 或是 ARA55 輔助下,抗雄性激素藥物例如 Hydroxyflutamide 或 Casodex 可 能 變 成 雄 性 激 素 。 在 AR cofactor ARA70 協 同 作 用 下 , 天 然 的 雌 性 激 素 (17beta-estradiol),可作用類似雄性激素,並活化雄性激素受調控基因。這說明抗雄性激 素藥物從拮抗性變成同質性(agonist)的途徑可受到 AR cofactor 的調節.簡而言之,AR cofactor 很可能就是前列腺癌轉變成雄性素不依賴型的關鍵蛋白質。過去的研究報告, 是屬於基因層次的證據,進一步必須要有細胞生物學及組織學上的佐證。因此本研究將 androgen dependent 的前列腺癌之細胞株(LNCaP)培養於 androgen deprivated Charcoal treated FBS medium,以瞭解細胞生長在缺乏雄性素環境下,這些 AR cofactor 是否其 mRNA 表達的量是否會有變化, 本實驗完全採用 non-isotope methods 進行 Multi-probe RNase protection assay: 選取 AR 相關之 cofactors 的基因序列設計一組多重探針之模版,
可以同時間定量分析一系列之RNA。這種變化,將是支持進一步研究 AR cofactor 是前列
腺癌轉變成雄性素不依賴型的關鍵.
由初步的結果可知當androgen dependent prostate cancer cell (LNCaP)培養於 androgen deprivation 的環境下其 AR 及 ara24 的 mRNA 會增加.而 ara54 ara70 及 RAC3 的 mRNA
則減少. 這個結果不同於已有的文獻報告認為 ara70 可能是前列腺癌轉變成雄性素不依 賴性的關鍵蛋白質的理論.
關鍵詞:雄性素受體,雄性素受體輔助因子,前列腺癌
Abstract
BACKGROUND. There are several hypotheses to explain why prostate cancer cell can
transform from hormone dependent to independent status after androgen deprivation therapy. Several evidences have been shown that during androgen deprivation, the androgen receptor (AR) cofactors still can enhance the transcription activity of AR to activate the AR target gene. So it is very important to know the expression of AR cofactors in androgen deprivation condition. If the expression of AR cofactors in the hormone independent status is increased, that would support the AR cofactors to be the trigger protein. This study investigated the expression patterns of several AR coactivators after androgen deprivation in LNCaP cell line.
Methods An androgen dependent prostate cancer cell line (LNCaP) was cultured in RPMI
medium with charcoal/dextran treated FBS for 28 days. Samples of total RNA collected in one-week interval were analyzed by using a non-isotope multi-probes ribonuclease protection assay system with the hAR multi-probe template set. We simultaneously analyzed the expression changes of 10 interested mRNA (AR, Rb, ara160, ara24, ara54, ara55, ara70, BRCA1, F-SRC-1 and RAC3) of AR and AR associated cofactors in only one hybridization reaction.
Results: From the preliminary data, we found the LNCaP cell in charcoal-treated serum for 28
days clearly show more than 1.5 folds increase in AR, ara160, and ara70 expression. The expression of ara24 and ara54 show increase but the increase below 1.5 folds. Only the RAC3 and F-SRC-1decreased in RNA expression.
Conclusions: This preliminary results support the previous reports that AR cofactors may be
the trigger key factor between androgen dependent and independent. KEY WORDS:
androgen receptor, coactivators, prostate cancer, cell culture, ribonuclease protection assay.
Abbreviations:
AR, androgen receptor; FBS, fetal bovine serum; ARA, AR associated coactivator; SRC-1, steroid receptor coactivator-1; Rb, retinoblastoma protein; HF, hydroxyflutamide; ER, estrogen receptor; E2, 17β-estradiol; CAT, chloramphenicol acetyltransferase; Adiol,
androst-5-ene-3β; 前言
INTRODUCTION
The role of androgen as an important factor in etiology and progression of human prostate cancer has been well documented. However, the molecular mechanism by which androgen modulate the prostate cancer growth is not yet clear. Androgen ablation has been the cornerstone of treatment for advanced prostate cancer, but the effect is often short-lived, as hormone-refractory elements continue to proliferate. However, the mechanisms responsible for androgen-independence remain uncharacterized. Now several studies have attempted to investigate how antiandrogens are converted from antagonists to partial agonists following prolonged androgen ablation therapy in prostate cancer patients. There are several hypotheses to explain why prostate cancer can transform from hormone dependent to independent status. There are several theories to explain this phenomenon of hormone independent status. It include, 1)AR mutations, 2)post-receptor alterations, 3)alterations in down-stream AR target genes, 4)alteration in the autocrine and paracrine loops, et al. But within them, the androgen receptor associated coregulator (AR cofactor) is an important finding since 1996. Several evidences have been shown that during androgen ablation or androgen blockage, the AR cofactors can enhance the ability of AR to activate the AR target gene. By this phenomenon,
the prostate cancer cell can keep growing in androgen deprivation status[1-3].
The AR functions as a ligand activated transcription factor that may play critical roles in prostate cancer growth and sexual development[4,5]. In general, most of the steroid receptors have a N-terminal transactivation domain, a DNA binding domain (DBD), a hinge region, a C-terminal ligand binding domain (LBD). The androgen receptor regulates androgen target genes by binding to androgen response elements with the potential involvement of coactivator or corepressor[6]. It has been demonstrated that receptors may utilize a group of coactivators to effectively stimulate gene transcription. These cofactors would regulate the steroid receptor's function through the transactivation process and may function as a bridge factor between the receptor and the basal transcriptional machinery to promote changes in the chromatin structure.
Several coactivators were found since 1996, e.g. ARA70[7], ARA55[8], and ARA54[9] for the C-terminal AR-ligand binding domain(AR-LBD), ARA160[10] and ARA24[11] for N-terminal. Some other cofactors have also been identified to associate with the AR DNA binding domain or AR N-terminal domain, such as RB, SRC1, CREB binding protein (CBP), a small nuclear ring finger protein (SNURF), a novel nuclear protein kinase (ANPK), TFIIH, etc. These coactivators can enhance DHT-mediated AR transcriptional activity and these interactions between AR and ARA70, ARA55, or ARA54 are androgen-dependent.
Katzenellenboger et al. proposed a new tripartiti system (ligand-receptor-cofactor) to explain the molecular interaction of steroid receptors that may define the potency and biological character of steroid hormones[12]. There are increasing evidences to show that the AR cofactor plays an important role in androgen-independent growth of prostate cancer.
Miyamoto et al. reported that antiandrogens (hydroxyflutamide, casodex) can activate androgen target genes in the presence of ARA70 or ARA55 in DU145 cell line [2]. ARA70 and ARA55 were able to significantly increase the androgenic activity of hydroxyflutamide, a
widely used antiandrogen for the treatment of prostate cancer. AR coactivators may be the trigger key factor between androgen dependent and independent. Hydroxyflutamide inhibits cell proliferation by binding the androgen receptor and thus block binding of its natural ligand, testosterone or dihydrotestosterone. However depending on the cell and tissue context, hydroxyflutamide can also behave as a partial agonist suggesting that its mechanism of action is complex and tissue specific. Furthermore, it is known that prostate cancer cells may develop resistance to hydroxyflutamide or may even cycle between phases of resistance and responsiveness. This varied response and abundant other evidence, suggests that hydroxyflutamide resistance is due to a cellular adaption phenomenon rather than to permanent genetic alterations.
Yeh et al. reported that E2(17β-estradiol)-AR-ARA70 also plays an essential role for the
AR function. ARA70 can induce AR transcriptional activity in the presence of estradiol 10nM, but not diethylstilbestrol. ARA70 is an essential factor to modulate this pathway, without ARA70, this pathway will be closed[13]. ARA70 is the best coactivator for conferring the androgenic activity on E2. This is the first evidence that T/DHT may not be the only ligands
for the AR. E2 represents another important natural ligand for AR that may play an essential
role for the AR function[13-15].
Androstenediol(Adiol), a natural estrogen, can activate AR target genes in the presence of AR, and that ARA70 can further enhance this Adiol-induced AR transcriptional activity. Hydroxyflutamide and casodex fail to block Adiol-mediated induction of AR transcriptional activity in prostate cancer cells[16]. This may raise critical questions about the possible role of Adiol and ARA70 in overcoming the effects of androgen ablation therapy for prostate cancer[14,16,17].
Together, it is suggested that some selective AR coactivators can modulate the specificity of sex hormones and antiandrogens. The post receptor alterations have been suggested that
relative expression of coactivators can modulate androgen and antiandrogen’s regulation for AR transcriptional activity. The agonist/antagonist receptor interactions alone may not be able to control the AR function, and the interaction between ligand-receptor complex and cofactors could be essential for steroid hormone function and sensitivity. So coactivators are very important factor to modulate the AR function. We can strongly believe that AR associated proteins (cofactors) play an important role to promote the prostate cancer cell growth from androgen-dependent into androgen-independent mode after long-term androgen deprivation.
So the expression change of AR cofactors gene either in hormone deprivation or androgen-blockade need to be evaluated in cell line and tissue specimen. The knowledge about the AR cofactors can help us to forecast the responsiveness of androgen-deprivation therapy in prostate cancer, which can be use as a prognostic factor. These findings may facilitate the development of factors or peptides (anti-coactivator) that can interrupt the androgen-mediated AR-ARA interactions to help us to fight the androgen-related prostate cancer.
If the AR cofactors are the trigger protein to transform the prostate cancer cell to be androgen independent, furthermore we need to know whether the expression pattern of AR cofactors is compatible in the androgen independent status. The specific changes in the AR coactivator gene expression profile of the cells could explain the molecular mechanism that prostate cancer cell become to hormone independent.
In summary, AR coactivator could be the key protein to support the prostate cancer cell growth in androgen independent mode. In our hypothesis, the expression of AR coactivators, e.g. ARA70 or ARA 55, may increase in prostate cancer cell after long-term androgen deprivation, so that AR function will be enhanced and AR antagonist will be converted to agonist.
prostate cancer cell line (LNCaP) was cultured in hormone deprivated status. Serial mRNAs were collected to analyze AR cofactors expression. For the purpose of simultaneous analysis with multiple targets, we used a multiple RPA template that includes ten of the AR cofactors and apply the non-isotope multi-probe RNAse protection assay to investigate the effect of androgen deprivation on AR cofactors.
This study is the first multi-probe ribonuclease protection assay (RPA) reported to measure expression of AR and AR associated cofactors mRNA influenced by androgen independent growth. This is also the first report by using non-isotope labeling in multi-probe RPA. From this study, it may help us to compare the expression change of several AR cofactors proteins in hormone deprivated condition. This would provide the evidence whether the increase expression of AR cofactor is compatible with transition from androgen dependence to androgen independence. This data can further clarify our preliminary molecular biology finding [1-3,13]. Furthermore, these finding can help us to forecast the responsiveness of androgen-deprivation therapy in prostate cancer.
研究方法
Materials and Methods Materials
RPMI1640 medium, penicillin/streptomycin, FBS and trypsin/EDTA were purchased from Life Technologies, Inc. (Grand Island, NY). Charcoal/dextran-treated certified FBS (Lot No. AKD11642, C-FBS) was from Hyclone (Logan, UT), with a certificate that the concentration of testosterone was below 3.0 ng/dl, progesterone below 10 ng/dl and 17 β-estradiol below 5 ρg/ml.
Cell culture and RNA isolation
Collection (Rockville, MD). They were grown in phenol red-positive RPMI 1640 medium supplemented with 10% FBS, 1% glutamine and penicillin/streptomycin at 37℃ in an atmosphere of 5% CO2 in air. We developed a subline of LNCaP cells grown in phenol
red-free RPMI1640 medium supplemented with 10% charcoal/Dextran treated FBS (C-FBS). Cell were fed twice per week and split once per week with trypsinization. We developed this subline of LNCaP cell in the condition mimics the androgen deprivated growth, and RNA of this LNCaP subline were collected in serial time course: 7th, 14th, 21st, 28th days.
Total RNA was extracted from the prostate cell line using TRIzol (Life technologies) according to the manufacturer’s specifications. Total RNA was extracted in chloroform, ethanol precipitated and stored at –80 ℃. Total 5 RNA samples were collected for time course analysis in week interval, one before and 4 after androgen deprivation.
Non-isotope Multi-probe ribonuclease protection assay (RPA)
The multi-probe RPA is a highly sensitive and specific method for the detection and quantitation of mRNA species. This novel method can evaluate the time course of several mRNA expressions in cell culture or tissue, and the quantitation of the expression levels can be compared simultaneously. So we can study the multiple mRNA expression patterns of cofactors in androgen deprivation growth and compare them in one hybridization experiment. For the purpose of non-isotope condition, our multi-probe RPA was succeeded by combining the hAR Multiple-Probe Template set (BD Pharmingen), MAXIscript T7TM in vitro transcription Kit (Ambion), Biotin-14-CTP (GIBCOBRL®) for nucleotide labeling, RPAIII kit (Ambion), and BrightStarTM BioDectTM Kit (Ambion). This novel non-radioisotope method can simultaneously and rapidly measure mRNA change of 10 AR associated coactivaors.
The strategy for the development of multi-probe RPA systems is to generate a series of such templates, each of distinct length and each representing a sequence in a distinct mRNA species. The templates are assembled into biologically relevant sets to be used by investigators
for a high-specific-activity, biotin-labeled anti-sense RNA probe set. The hAR Human Androgen Receptor Multi-Probe template set was purchased from BD PharMingen Co., containing DNA templates which can be used for the T7 RNA polymerase-directed synthesis of biotin-labeled, anti-sense RNA probe that can hybridize with target human mRNAs encoding androgen receptor[18], RB[19], ara160[10], ara24[11], ara54[9], ara55[8], ara70[7], BRCA1[20]7, F-SRC-1[21], RAC3[22] as well as the two housekeeping gene products, L32 and GAPDH. The multiple hAR RNA probes was synthesis using the MAXIscript T7TM in vitro transcription Kit (Ambion). 0.4µl Biotin-14-CTP (10 mM, GIBCOBRL®)(in the ratio of 40% biotin-CTP: 60%CTP) was used as label nucleotide in each 20 µl reaction in vitro transcription. NucAwayTM Spin Columns (Ambion, Inc) was used for the removal of unincorporated nucleotides and salts after probe synthesis reactions.
Multi-probe RNase protection assay was performed by using RPAIII kit (Ambion, Inc). For the purpose of non-isotope design, biotin-labeled anti-sense RNA probe set was used as manufacturer’s protocol. The probe set is hybridized in excess to target RNA in solution, after which free probe and other single-stranded RNA are digested with RNase. The remaining RNase-protected probes are purified and analyzed by resolving on denaturing polyacrylamide gels.
Briefly, after denature in 90℃~95℃ for 2~3 minutes,dried total RNA (10 µg) with 1 µl biotin-labeled multi-probe sets was hybridized (56℃; 12 h) in 9µl RPAIII hybridization buffer, Reaction mixtures were then treated with 100µl (500x diluted) RNase A/RNase T1 Mix (Ambion) and incubated 45 minutes at 30℃. Digestion and precipitation were easily performed by adding 150µl RPAIII Inactivation/Precipitation III solution (Ambion). RNA was precipitated from each reaction and pellets were washed with ethanol, air dried, resuspended in Gel loading Buffer II (Ambion) , heated (95℃, 3 min), and resolved on a denaturing 5% polyacrylamide gel at 250V for 3.5 h. The RNase-protected probes on the polyacrylamide gel
was then electroblotted onto a positively charged nylon membrane. The biotinylated RNA probe on the nylon membrane was then detected by BrightStarTM BioDectTM Kit (Ambion, Inc.) and the film was exposed by the light emission for one to two hours. Protected bands on the x-ray film were quantitated by densitometry. Relative level of protected probe were calculated by normalizing the specific band to the ribosomal RNA L32 band included in the multi-probe kits.
For each multi-probe RPA assay, negative controls included 5 µl of probe mixture only and 5 µl of probe mixture incubated with 10 µg of yeast tRNA (Ambion). The probes-only control was treated with 1x RNase Digestion buffer in the absence of RNase A/RNase T1 Mix.
結果與討論
Cell-growth properties of LNCaP cells in Charcoal-treated serum
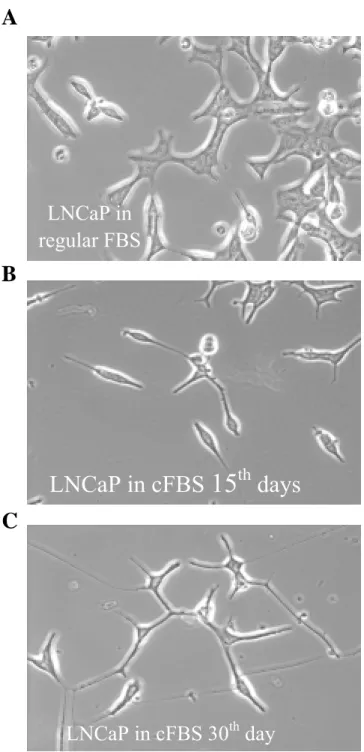
We examined the in vitro growth properties of LNCaP cells under the steroid-reduced culture conditions. By continuous passage of LNCaP parental cells in the medium supplemented with 10% charcoal-treated FBS, we found the cell morphologic feature and growth rate changed gradually (Fig. 1A-C). The LNCaP cell is epithelial cancer cell and the doubling time is 2 days. After 2 weeks in charcoal-treated serum, the cell growth rate became slower and the size of cell body became smaller. At the 4th week, the morphology changes of LNCaP are so obvious. The attachment to cell culture dish is loosening and cell became neuron-like shape with
elongated cytoplasmic process.
Multi-probe RPA study
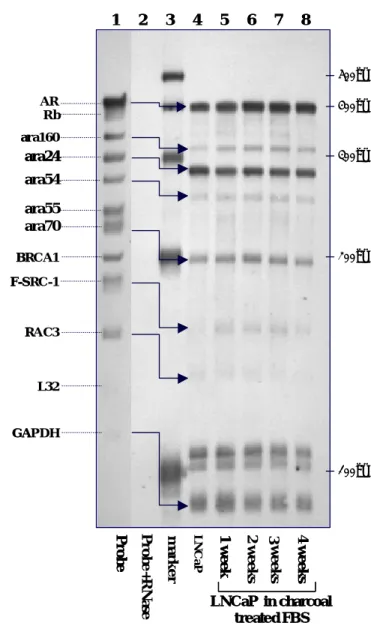
We collected 4 samples of RNA from LNCaP cell cultured in charcoal-treated FBS for 1, 2, 3 and 4 weeks and one sample of RNA from LNCaP in regular FBS. Samples of total RNA from LNCaP were analyzed for distinct mRNA species by using a non-isotope multiple probes ribonuclease protection assay system as the section of methods describes. We used the
BrightStarTM BioDetectTM Kit for nonisotopic chemiluminescent detection of biotinylated RNA probes. The chemiluminescent exposure on film shows the lane of the hAR multi-probe not treated with RNases (lane 1). Also shown are the corresponding RNase-protected probe following hybridization with RNA from LNCaP(10 μ g, Lane 4-8) and BrightStar TM Biotinylated RNA century TM size markers (1μg, Lane 3). Each probe band (Lane 1) migrates slower than its protected band (Lane 4-8). For each multi-probe RPA assay, lane 2 is negative controls. 5 µl of probes were hybridized with 10 µg of yeast tRNA (Ambion) and digested with RNase A/RNase T1 Mix. The probes-only control in lane 1 was treated with 1x RNase Digestion buffer in the absence of RNase A/RNase T1 Mix.
The expressions of mRNA are indicated in relative light unit of CDP-StarTM on chemiluminescent exposure on film. The band of L32 was used to correct the loading RNA amount in each lane. The data were collected from three sets of independent experiments and show as mean ± S.D.
As shown in figure 2, we found that LNCaP cell cultured in charcoal-treated serum for 28 days would induce more than 1.5 folds expression in AR, ara160, and ara70. Another two mRNA, ara24 and ara54, also had increase pattern but the ratio less than 1.5 folds. Only the RAC3 and F-SRC-1decreased in RNA expression. The three bands of Rb, ara55 and BRCA1 are too low to be detected.
Discussion
To date, prostate cancer has become the most frequently diagnosed neoplasm in the United States[23], and the 9th to 10th leading cause of cancer-related death in Taiwan. Androgen ablation has been the cornerstone of treatment for advanced forms of this disease, and a combination therapy of surgical or medical castration with antiandrogen, such as hydroxyflutamide or casodex, is now widely used to reduce the level of endogenous androgens coming from, for example, adrenal sources. Although such “total androgen
blockage” can reduce tumor regression, the effect is often short-lived, as hormone-refractory elements continue to proliferate. It has been suggested that genetic changes of the AR gene may contribute to a poor response to hormone therapy [24]. However, the mechanisms responsible for androgen-independence remain uncharacterized. In recent ten years, the knowledge of AR associated protein has provided a new direction to study the mechanism of androgen independent prostate cancer cell growth. There are several evidences in the molecular biology to support the linkage between AR cofactors and androgen independent growth[2,3,13,15]. We have described several evidences in the introduction section, AR cofactors might be the critical proteins to enhance the effect of minimal androgen, turn on the androgen receptor by estrogen, or promote agonist activity of antiandrogens. Although the evidence is well demonstrated in transcriptional level, we need more data on cell biology to support our hypothesis. We need to investigate the expression of androgen receptor coactivators in the human prostate for a better understanding of androgen deprivation action in prostate cancer. If some of those coactivators were up regulated by the androgen deprivation, that would support the critical role of coactivators in androgen independent growth.
The multi-probe RPA is a novel method for evaluation of mRNA expression, and this can simultaneously evaluate several interesting mRNA in different time points. Only as little as the amount of RNA for one northern blotting, the multi-probe RPA can evaluate ten targets to the most in once procedure. So this can make it possible to compare several targets in only one membrane. In the traditional northern blotting, if we want to evaluate more than one mRNA target, we need to strip the hybridized probe on the membrane for another session of procedure. This would result in some errors of repeat northern blotting for several RNA targets. Besides the one s
This is the first report to study AR coactivators expression pattern related to steroid-reduce environment by using a novel non-isotope multi-probe RPA assay method. This preliminary results support AR coactivators to be the key protein in prostate cancer cell transit from androgen-dependent to androgen-independent growth. From previous reports, it is known that AR cofactors may amplify or change the AR transcription pathway[2,3,13]. When the
androgen dependent LNCaP cell culture in charcoal treated serum, the expression of AR cofactors (e.g. ara160, ara24, ara54, ara70) revealed increase production. From this expression change, these AR coactivators may not only enhance the ordinary AR transcription pathway by the increase amount of AR coactivators protein, but also can activate AR through different pathway or activate other ligand e.g. estradiol or antiandrogen[2,13]. This is the first cell biological data to support the hypothesis of AR cofactors to be the trigger key factors between androgen dependent and independent. Our result can encourage us put more force to further clarify the mechanism of AR coactivators in the prostate cancel growth.
Figure 1.
A
B
C
LNCaP in cFBS 30
thday
LNCaP in cFBS
15
thdays
LNCaP in
regular FBS
Figure 2 A B ara24 ara54 Pr o b e 4 w eek s LNCaP in charcoal treated FBS 3 w eek s 1 w eek AR Rb ara160 ara24 ara55 ara70 BRCA1 F-SRC-1 RAC3 L32 GAPDH Pr o b e ma rk e r LN C a P 100bp 200bp 300bp 400bp 500bp 1 2 3 4 5 6 7 8 w eek s LNCaP in charcoal treated FBS 2 w eek s 1 w eek P ro b e + RNa se ara24 ara54 Pr o b e 4 w eek s LNCaP in charcoal treated FBS 3 w eek s 1 w eek AR Rb ara160 ara24 ara55 ara70 BRCA1 F-SRC-1 RAC3 L32 GAPDH Pr o b e ma rk e r LN C a P 100bp 200bp 300bp 400bp 500bp 1 2 3 4 5 6 7 8 w eek s LNCaP in charcoal treated FBS 2 w eek s 1 w eek P ro b e + RNa se Ra ti o o f re la tiv e li gh t U nit ( C DP -S ta r TM ) No. of weeks in Charcoal treated FBS 0 0.5 1 1.5 2 2.5 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4
AR ara160 ara24 ara54 ara70 F-SRC-1 RAC3
Ra ti o o f re la tiv e li gh t U nit ( C DP -S ta r TM ) No. of weeks in Charcoal treated FBS 0 0.5 1 1.5 2 2.5 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4
Table 1
AR, AR associated cofactors and internal control genes included in the twelve templates in the hAR multi-Probe template set with expected sizes (bp) of full-length probes and protected fragments.
Size (bp) Template
Full-length probe Protected probe
AR 429 400 Rb 381 352 ara160 349 320 ara24 315 286 ara54 284 255 ara55 256 227 ara70 231 202 BRCA1 211 182 F-SRC-1 190 161 RAC3 170 141 L32 141 112 GAPDH 126 97
Legend
Figure 1. Phase-contrast micrographs of different LNCaP cells (200X). A: LNCaP cells were
cultured in RPMI-1640 medium supplemented with 10% FBS. B: LNCaP cells were cultured in RPMI-1640 medium supplemented with 10% charcoal-treated FBS for 2 weeks, C: LNCaP cells were cultured in the same condition as B for 4 weeks.
Figure 2. Multi-probe RPA study for AR and its associated cofactors of LNCaP cell
influenced by steroid reduced culture medium. (A) The LNCaP are cultured in RPMI 1642 with charcoal treated FBS for 4 weeks. Samples of total RNA (10μg, Lane 5-8) were isolated weekly from LNCaP that cultured in RPMI 1642 medium with charcoal treated FBS. These total RNA were analyzed for distinct mRNA species by using Ambion’s RPAIII system with the Pharmingen’s RiBoquantTM hAR multiple-probe template set and non-isotope biotin labeling. The expressions of mRNA are indicated in relative light unit of CDP-StarTM on chemiluminescent film exposure by nylon membrane electrobloted from denaturing polyacrylamide gel. The chemiluminescent exposure on film shows RNase-protected probe following hybridization with RNA from LNCaP (Lane 4-8). Also shown are the RNA probes transcribed from hAR multi-probe template set not digested by RNases (Lane 1), probe hybridized with yeast t-RNA digested by RNase as negative control (Lane 2), BrightStar TM Biotinylated RNA century TM size markers (1μg, Lane 3). Each probe band (Lane 1) migrates slower than its protected band (Lane4-8). A GAPDH probe was used as a control for equivalent mRNA loading. The expressions of mRNA are indicated in relative light unit of CDP-StarTM on chemiluminescent exposure on film. The data were collected from three sets of independent experiments.
(B)Bands from figure 1A were quantified by densitometry. Results are reported as relative CDP activity. The expression of GAPDH gene was analyzed for standardization purposes and
the expression level in lane 4 was set as 100% to compare with Lane 5-8. The data represent the mean ± SD from three independent experiments.
Reference
1. Fujimoto N, Yeh S, Kang HY, Inui S, Chang HC, Mizokami A, Chang C. Cloning and characterization of androgen receptor coactivator, ARA55, in human prostate. J Biol Chem 1999;274:8316-8321.
2. Miyamoto H, Yeh S, Wilding G, Chang C. Promotion of agonist activity of antiandrogens by the androgen receptor coactivator, ARA70, in human prostate cancer DU145 cells. Proc Natl Acad Sci U S A 1998;95:7379-7384.
3. Yeh S, Miyamoto H, Chang C. Hydroxyflutamide may not always be a pure antiandrogen. Lancet 1997;349:852-853.
4. Chang C, Kokontis J. Identification of a new member of the steroid receptor super-family by cloning and sequence analysis. Biochem Biophys Res Commun 1988;155:971-977.
5. Trachtenberg J. Innovative approaches to the hormonal treatment of advanced prostate cancer. Eur Urol 1997;32:78-80.
6. Chang C, Saltzman A, Yeh S, Young W, Keller E, Lee HJ, Wang C, Mizokami A. Androgen receptor: an overview. [Review] [236 refs]. Crit Rev Eukaryot Gene Expr 1995;5:97-125.
7. Yeh S, Chang C. Cloning and characterization of a specific coactivator, ARA70, for the androgen receptor in human prostate cells. Proc Natl Acad Sci U S A 1996;93:5517-5521.
Suzuki R, Sasaki T. Cell adhesion kinase beta forms a complex with a new member, Hic-5, of proteins localized at focal adhesions. J Biol Chem 1998;273:1003-1014.
9. Kang HY, Yeh S, Fujimoto N, Chang C. Cloning and characterization of human prostate coactivator ARA54, a novel protein that associates with the androgen receptor. J Biol Chem 1999;274:8570-8576.
10. Garcia JA, Ou SH, Wu F, Lusis AJ, Sparkes RS, Gaynor RB. Cloning and chromosomal mapping of a human immunodeficiency virus 1 "TATA" element modulatory factor1. Proceedings of the National Academy of Sciences of the United States of America 89(20):9372-6, 1992.
11. Hsiao PW, Lin DL, Nakao R, Chang C. The linkage of Kennedy's neuron disease to ARA24, the first identified androgen receptor polyglutamine region-associated coactivator. J Biol Chem 1999;274:20229-20234.
12. Katzenellenbogen JA, OMalley BW, Katzenellenbogen BS. Tripartite steroid hormone receptor pharmacology: Interaction with multiple effector sites as a basis for the cell- and promoter-specific action of these hormones. Mol Endocrinol 1996;10:119-131.
13. Yeh S, Miyamoto H, Shima H, Chang C. From estrogen to androgen receptor: a new pathway for sex hormones in prostate. Proc Natl Acad Sci U S A 1998;95:5527-5532.
14. Yeh S, Chang HC, Miyamoto H, Takatera H, Rahman M, Kang HY, Thin TH, Lin HK, Chang C. Differential induction of the androgen receptor transcriptional activity by selective androgen receptor coactivators. [Review] [40 refs]. Keio J Med 1999;48:87-92.
15. Yeh S, Kang HY, Miyamoto H, Nishimura K, Chang HC, Ting HJ, Rahman M, Lin HK, Fujimoto N, Hu YC, Mizokami A, Huang KE, Chang C. Differential induction of androgen
receptor transactivation by different androgen receptor coactivators in human prostate cancer DU145 cells. Endocr J 1999;11:195-202.
16. Miyamoto H, Yeh S, Lardy H, Messing E, Chang C. Delta5-androstenediol is a natural hormone with androgenic activity in human prostate cancer cells. Proc Natl Acad Sci U S A 1998;95:11083-11088.
17. Chang HC, Miyamoto H, Marwah P, Lardy H, Yeh S, Huang KE, Chang C. Suppression of Delta(5)-androstenediol-induced androgen receptor transactivation by selective steroids in human prostate cancer cells. Proc Natl Acad Sci U S A 1999;96:11173-11177.
18. Govindan MV. Specific Region in Hormone Binding Domain Is Essential for Hormone Binding and Transactivation by Human Androgen Receptor. Mol Endocrinol
1990;4:417-427.
19. Cordoncardo C, Richon VM. Expression of the Retinoblastoma Protein Is Regulated in Normal Human Tissues. Am J Pathol 1994;144:500-510.
20. Miki Y, Swensen J, Shattuck-Eidens D, Futreal PA, Harshman K, Tavtigian S, Liu Q, Cochran C, Bennett LM, Ding W. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science 1994;266:66-71.
21. Takeshita A, Yen PM, Misiti S, Cardona GR, Liu Y, Chin WW. Molecular cloning and properties of a full-length putative thyroid hormone receptor coactivator5. Endocrinology 137(8):3594-7, 1996.
22. Haataja L, Groffen J, Heisterkamp N. Characterization of RAC3, a novel member of the Rho family. Journal of Biological Chemistry 272(33):20384-8, 1997.
Ca-A Cancer Journal for Clinicians 1998;48:192.
24. Gaddipati JP, Mcleod DG, Heidenberg HB, Sesterhenn IA, Finger MJ, Moul JW,
Srivastava S. Frequent Detection of Codon-877 Mutation in the Androgen Receptor Gene in Advanced Prostate Cancers. Cancer Res 1994;54:2861-2864.
計畫成果自評:
研究內容與原計畫相符、LNCaP cell 培養於 androgen-deprivated medium 生長速度變 緩慢,原計劃期望培養出 stable growth 的 cell line 還未達成,預期尚需一年的時間 努力. 外國文獻已有以同樣方法完成新的 cell line, 因此可以考慮直接向對方索取以 加速實驗的進行.
研究成果的學術或應用價值: 以 Non-isotope method 進行 multiple RPA assay methods 是高難度的定量檢測 RNA 的方法,不但可同時定量多組樣本且可以同時比較一系列多達 十種之 mRNA 這是文獻上尚未見過的報告,值得一般實驗室採用.
主要發現: 由初步的結果可知於 androgen deprivation 的環境下其 AR 及 ara24 的 mRNA 會增加.而 ara54 ara70 及 RAC3 的 mRNA 則減少. 這個結果仍不能支持已有的文獻報告 的理論認為 ara70 可能是前列腺癌轉變成雄性素不依賴型的關鍵蛋白質. 因此如何看待 AR cofactor 在 androgen independent prostate cancer cell 形成過程中扮演的角色, 將需要不同的分子生物學的解釋.本次研究之方法及結果適合在學術期刊發表、