Cis-regulator y elements of the mouse Krt1.12 gene
I-Jong Wang¶†§, Eric C. Carlson¶§, Chia-Yang Liu¶, Fung-Rong Hu†, and Winston W. -Y. Kao¶∗
¶Department of Ophthalmology, University of Cincinnati, Cincinnati, Ohio 45267-0527, USA.
†Department of Ophthalmology, National Taiwan University Hospital, Taipei 100, Taiwan
§Authors contributed equally to this manuscript
Running title: Cis-regulatory elements of the mouse Krt1.12 gene
∗Correspondence and reprint requests should be addressed to W. W. -Y. Kao. Department of Ophthalmology, University of Cincinnati Medical Center, Health Professions Building Suite 350, ML0527, 3223 Eden Avenue, Cincinnati, Ohio. TEL: (513) 558-5151, FAX: (513) 558-3108 E-mail: [email protected]
Key wor ds: Keratin 12; gene gun; footprinting; corneal epithelium; electromobility
INTRODUCTION
Keratins are a group of water-insoluble proteins that form the 10 nanometer (nm)
intermediate filaments found in epithelial cells.21 The major function of the cytoskeletal
network formed by keratins is to provide a rigid epithelial cell layer that protects
underlying tissues from the environment.11 The proposition is substantiated by mutations
of K1/K10 and K5/K14, which manifests epidermolysis hyperkeratosis and epidermolysis
bullosa simplex in skin, respectively. Whereas mutations of the K12 gene in human and
ablation of the K12 gene via gene targeting in mice results in fragile corneal epithelium, a
clinical manifestation characteristic of Meesmann’s corneal dystrophy.14,15
The approximately thirty different keratin proteins are subdivided into acidic type I
and basic type II 13,21,25,30. In vivo, a basic keratin usually heterodimerizes or "pairs"
with a particular acidic keratin.8,13,21,25,27,28,30,33,37. The expression of keratin pairs
is tissue-specific, differentiation regulated, and development-dependent. For example,
K5/K14 keratin pair is found in the basal cell layer of all stratified epithelium; whereas,
K1/K10 keratin pair is expressed by suprabasal and superficial epidermal epithelial cells.
The K3/K12 pair is regarded as a marker for cornea-type epithelial differentiation. It should
be noted however that the expression of K3 is not limited to cornea epithelium; it has been
detected in several other tissues.5,7,8,36 We have cloned the mouse cornea-specific K12
5,6,18,22,39,40
Many keratin genes are regulated at the transcriptional level in a coordinate manner
during keratinocyte differentiation. 9,10 Tissue-specific keratin expression is regulated by
a complex collaboration between ubiquitous and tissue-specific transcription factors and is
dependent on the turn-on and -off of a variety of regulatory genes. 11,26,31 There are
several transcriptional factors known for the regulation of keratin gene expression. For
example, AP-2 and SP-1 have been shown to be able to regulate the expression of K1, K3,
K5, K6, and K14. 3,16,17,38 Furthermore, the regulation of a tissue-specific keratin gene
always needs coordinated expression of several transcription factors. 3,16,17,38
Recently, we successfully used Gene Gun, a particle-mediated gene transfer
technique, to deliver K12-promoter reporter genes to rabbit corneal epithelial cells in vivo
and identified a 0.6 kb DNA fragment 5’ flanking Krt1.12 possibly contained corneal
epithelial cell-specific regulatory cis-DNA elements. 29 In the present study, we further
examined the 0.6 kb 5’ flanking region in the K12 gene to determine the cis-regulatory
elements accounting for corneal epithelial cell-specific expression. To do this, we used
both in vitro, DNase I footprinting and electrophoretic mobility shift assays (EMSA), and
in vivo, Gene Gun, techniques to elucidate the epithelial cell-specific cis-regulatory
MATERIALS AND METHODS
Plasmids
Two plasmid DNA constructs, pCMVβ and pNASSβ (Clontech, Palo Alto, CA), were
used as positive and negative control reporter genes. Three reporter gene constructs, 0.2
KZ, 0.4 KZ, and 0.6 KZ were prepared by cloning 127 bp, 327 bp, and 527 bp sequence
plus 40 bp of exon 1 untranslated region of the Krt1.12 gene to pNASSβ plasmid that contained a lac Z reporter gene.
In vivo Par ticle-Mediated Gene Tr ansfer : Gene Gun
All animal experiments were performed according to the ARVO resolution on the use
of animals in vision research. Plasmid DNA purified using Qiagen columns (Qiagen,
Chatsworth, CA) was coated onto 0.6 µm, 1.0 µm or 1.6 µm gold particles (5 µg DNA
per mg of gold), loaded into Tefzel tubing (Biorad, Hercules, CA), and transiently
transfected into New Zealand white rabbit (~2 kg) corneal epithelium using the
HeliosTM Gene Gun System (Biorad) at 200 psi according to Shiraishi et al. 11 Samples
were collected 48 h after delivery and subjected to β-galactosidase activity analysis.
Prepar ation of enzyme extr acts
Excised tissue specimens were minced with a razor blade, and 0.5 ml of extraction
buffer (0.25 M Tris-HCl, pH 7.4, 0.1 % Tween 20) was added. The samples were
were collected by centrifugation at 13,000 x g at 4°C for 10 min.
β-Galactosidase Activity Assay And Whole Mount β-galactosidase Histochemical Staining
Aliquots of supernatant were incubated in a 0.3 ml mixture containing 50 mM
2-mercaptoethanol, 1 mM MgCl2, 1.33 mg/ml o-nitrophenyl β-galactopyranoside and
0.1 M phosphate buffer, pH 7.0 at 37°C for 1 to 5 h. A 0.7 ml aliquot of 1 M Na2CO3
was added to terminate the reaction. The enzyme activity was determined by comparing
the optical density at 460 nm to that of purified β-galactosidase (Boehringer and
Mannheim, Indianapolis, IN). The promoter activity of KZ reporter gene constructs
was calculated as a fold-increase of the enzyme activities derived from the
promoter-less pNASSβ construct.
For whole mount X-gal staining, the eyeball was enucleated, fixed immediately
with 4% paraformaldehyde in PBS at 4°C for 2 h, and washed with PBS buffer (pH 7.0)
containing 137 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4, and 1.4 mM KH2PO4.
Staining was carried out at 30°C for 16 h in a solution of
5-bromo-4-chloro-3-indolyl-β-galactopyronoside (X-gal, Sigma) at a final
concentration of 0.4 mg/ml. Following staining, eyeballs were rinsed with PBS and
Prepar ation of Nuclear Extr acts
Nuclear extracts were prepared as described by Andrews and Faller.1 Briefly,
bovine corneal epithelium was scraped from enucleated eyes into ice-cold PBS, and
then homogenized by Teflon-glass in homogenization buffer (2 ml/gm tissue)
containing 0.1% Triton X-100, 10 mM HEPES (pH 7.6), 25 mM KCl, 1 mM EDTA, 2
M sucrose, 0.5 mM spermidine, 0.15 mM spermine, 10% (v/v) glycerol in DEPC H2O.
The homogenate was then 3-fold diluted in the same buffer without glycerol and
centrifuged at 104,000 x g for 30 min on a 10 ml cushion of homogenization buffer.
Pellets were then combined and resuspended in 15-20 ml of a 9:1 (v/v) mixture of
homogenization buffer and glycerol, using a Teflon-glass homogenizer. This
homogenate was layered on a 10 ml cushion of the 9:1 mixture and centrifuged as
above. The nuclei were homogenized in cold lysis buffer containing 10 mM HEPES
(pH 7.6), 0.1 M KCl, 3 mM MgCl2, 0.1 mM EDTA, 10% glycerol, 1 mM DTT, and 0.1
mM PMSF in DEPC H20 with a glass-glass homogenizer. The absorbance was then
checked with a spectrophotometer at a 260 nm wavelength and diluted in lysis buffer to
10 A260 unit/ml. The nuclear extract was subjected to two steps of ammonium sulfate
fractionation by first incubating for 30 min in a 10% vol of 4 M (NH4)2SO4 (pH 7.9),
followed by centrifugation (100,000 x g for 60 min). The nuclear proteins in the
by centrifugation (100,000 x g, 60 min). The nuclear protein extract was resuspended
and dialyzed in buffer containing 25 mM, 10 mM HEPES, 40 mM KCl, 10% glycerol,
0.1 mM EDTA, 1 mM DTT, in DEPC H2O. The extract was then aliquoted and stored
at -70 °C.
DNase I Footpr inting Analysis
Three DNA probes (-332 bp to –109 bp, -394 bp to –131 bp, and -599 bp to –375 bp,)
were prepared via PCR (primers listed in Table 2) and 5’ labeled with [γ-32P] ATP
using T4 polynucleotide kinase. A 50 µl mixture containing 10 mM Tris-HCl (pH 8.0),
5 mM MgCl2, 5 mM CaCl2, 50 mM KCl, 500 nM dithiothreitol, 0.05 mg/ml bovine
serum albumin, and 40.5 ng/µl poly-dIdC was incubated with crude nuclear extracts
(75 µg) on ice for 15 minutes. The mixture was then incubated for an additional 45
minutes with 100,000 CPM of DNA probe. Variable amounts of DNase I were added to
the mixture and incubated at 25°C for 1 min 30 sec. This reaction was stopped with 50
µl of stop buffer (0.2 M NaCl, 30 mM EDTA, 1% SDS, and 0.1 mg/ml yeast tRNA). Digested DNA probes were purified by phenol extraction and ethanol precipitation and
separated on 6.5% denaturing polyacrylamide gels. Dried gels were exposed to Kodak
XAR film with intensifying screens for 24 hours at –80oC. G and G+A sequencing
reactions were performed to determine the positions of the protected regions. Negative
protected by nuclear proteins were numbered according to the nucleotide positions
relative to the transcription start site of the Krt1.12 gene.
Electrophoretic Mobility Shift Assays
Gel-shift analysis of the potential tissue specific region of the Krt1.12 promoter was performed with 32P labeled double-stranded synthetic oligonucleotide probes (-182 bp
to –111 bp and -256 bp to –193 bp) (Table 3), and crude nuclear extracts prepared from
bovine corneal epithelial cells as previously described. Twenty microliter binding
reactions were performed, which contained 10 µg of nuclear proteins, 20,000 cpm
double strand oligonucleotides end-labeled with [γ-32P] ATP, 12 mM HEPES (pH 7.9),
10% glycerol, 4 mM Tris-HCl (pH 8.0), 1 mM EDTA, 0.3 mg/ml bovine serum albumin,
1 mM dithiothreitol (DTT), 0.1 mg/ml poly-dAdT, and 60 mM KCl. Each labeled probe
was competed with an excess of the same unlabeled double-stranded synthetic oligomer.
Binding mixtures were separated on a nondenaturing 5% polyacrylamide gel and
exposed to Kodak XAR film.
Immunohistochemistr y
Paraffin sections of mouse eyes were deparaffinized and incubated in PBS for 30
min. The tissue section was then blocked with BSA and 3% nonfat milk in PBS for 60
min at room temperature. Rabbit antibodies against mouse c-jun/AP-1 and c/EBPβ
and incubated on the tissue sections at 4°C overnight. The sections were then washed
in PBS. A goat anti-rabbit Alexa Fluor 488 (Molecular Probes Inc. Eugene, OR)
secondary antibody was used for immunoreactivity detection using fluorescence
microscopy.
RESULTS
Analysis of pr omoter activities of the 5'-Flanking Region of the Krt1.12 Gene
It was previously demonstrated that the –0.6 kb 5’flanking DNA fragment
could direct cornea-specific expression of a β-galactosidase gene in vivo 29. In an
attempt to identify the region where cornea-specific 5’ flanking cis-regulatory elements exist in the Krt1.12 gene we tested the 0.2 KZ, 0.4 KZ and 0.6 KZ constructs in vivo. Rabbit corneas, conjunctivas, and skin were transiently transfected with the 0.2, 0.4,
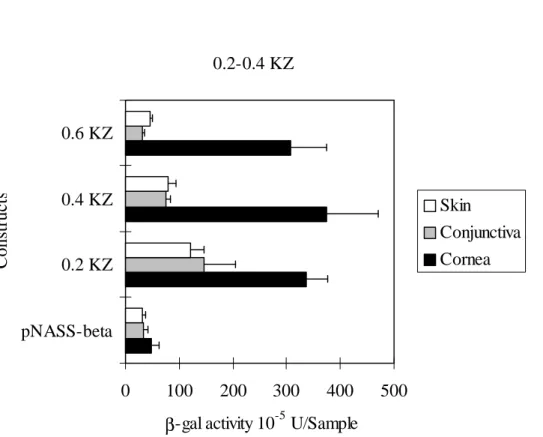
and 0.6 KZ constructs using gene gun. Figure 1 shows that all three promoter
constructs express 7-10 fold β-gal activities above the promoter-less pNASSβ construct
in cornea. In skin and conjunctiva, the 0.4 and 0.6 KZ constructs yield insignificant
β-galactosidase activities in comparison to those of pNASSβ; whereas, the 0.2 KZ construct produces 3-fold β-galactosidase activity above pNASSβ. The observations
suggest that the 0.2 KZ construct does not have cornea-specific cis-regulatory elements for the expression of the β-galactosidase reporter gene. And, the 0.4 KZ and 0.6 KZ
conjunctiva and skin. To further elucidate this suggestion whole mount X-gal staining
was performed with transfected rabbit eyes. Figure 2 demonstrates that the 0.2 KZ
construct (Panel A) shows β-gal expression in both the cornea and conjunctiva. The 0.4
KZ (Panel B) and 0.6 KZ construct (Panel C) show reporter gene expression
exclusively in the cornea. The observation suggested that a cornea-specific regulatory
element(s) might exist in the –327 to +40 bp region of Krt1.12 gene.
DNase I Footpr inting Assay
Since the in vivo data indicates that the 0.4 KZ reporter gene construct has cornea-specific promoter activity, DNase I footprinting was performed to further
identify the DNA sequences that may account for cornea-specific Krt1.12 gene expression. Two footprinting probes (-394 bp to –131 bp, and –331 bp to –109 bp)
were used to identify the possible transcription factor binding site(s) that the previously
described data suggested exist in this region. Bovine epithelial nuclear extract
protection was seen for a 72 bp fragment in the –182 bp to –110 bp region of the
Krt1.12 gene. Also, protection existed in 2 other regions of the Krt1.12 gene, from –231 bp to –193 bp and from –256 to –243 (Fig3).
Electrophoretic Mobility Shift Assays (EMSA)
designed from the regions protected by epithelial nuclear proteins in the footprinting
assays. EMSA of site from –182 to -111 and –256 to –193 showed a binding complex
with nuclear proteins from corneal epithelial cells, but the complex could also be
detected at lower affinity with nuclear proteins of retinal cells (Fig. 4) The radioactive
labeled complex was competed and eliminated by a 400- or 800-fold molar excess of
the unlabeled oligonucleotides termed self-competitor. Sequence analysis suggests that
several transcriptional factors can potentially bind to this region of the of Krt1.12 promoter, which includes c-jun/AP-1 and c/EBPβ binding sites (Fig. 5).
Immunohistochemistr y
Consensus sequence analysis revealed potential c-jun/AP-1 and c/EBPβ (Fig 5)
binding sites in the 5’ flanking Krt1.12 region. Immunohistochemistry was performed to determine if the expression of these two transcription factors may be present in
corneal epithelial cells, but absent in limbal basal epithelial cells and can be correlated
with that of keratin 12 expression. Figure 6 demonstrates the presence of c-jun/AP-1
and c/EBPβ in the nucleus of all mouse corneal epithelial cell layers. However,
c/EBPβ appears to have a significantly lower expression level in the limbal region of
the cornea as compared to c-jun/AP-1. The observation is consistent with the
expression of keratin 12 in corneal epithelial cells, since keratin 12 is only expressed in
Discussion
Although many keratin genes have been cloned, the 5’-upstream sequences of only a
few of them have been found to function as tissue-specific promoters by in vitro analysis of cultured keratinocytes and in vivo in transgenic mice. In the present study, we discovered the 0.4 kb 5’ flanking region of the mouse Krt1.12 gene appeared to contain the cis-regulatory elements necessary for cornea-specific expression as
demonstrated by in vivo transfection of β-galactosidase reporter genes with Gene Gun. Analyses using foot printing and EMSA further identifies two regions –256 bp to –193
bp and –182 bp to –111 bp 5’ to the transcription initiation site that may play a role in
Krt1.12 gene expression by corneal epithelial cells.
Analysis of sequence comparison to the Gene Bank (NCBI GenBank database,
Blast web client software) revealed several possible transcription factor binding sites
including, AP-1 (-240 to –250 bp and –124 bp to –133 bp) and c/EBPβ (-133 bp to –147
bp, -229 to –242 bp, and –226 to –243 bp) and many others in the sense and antisense
orientation as shown in Figure 5.
AP-1 (activation protein 1) was well known to be a regulator of many keratin
genes including K1, K3, K5, K8, K10, and K18, which are expressed by basal and
suprabasal epithelial cells of stratified epithelium, as well as single cell-layer
of the cornea which is consistent with the expression patterns of keratinocyte
differentiation. The retina was also positive for AP-1 (data not shown) which in parts
explains the EMSA data.
Expression of K12 is limbal basal negative until after migration from the limbal
basal layer. Our c/EBPβ transcription factor immunostaining shown in Fig. 6 was
suprabasal positive in the nuclei of corneal epithelial cells, but expression appears
down regulated in the limbal basal region especially with respect to c/EBPβ. C/EBP
family members (α, β, γ, and δ) contain the bZIP region. The bZIP region is
characterized by two motifs, one of which is involved in DNA binding, and the other a
leucine zipper involved in dimerization. c/EBPβ can homodimerize or heterodimerizes
with other family members and other transcription factors (NFκB, p65, p50, and rel
family members) {Stein, Cogswell, et al. 1993 18 /id}. The interaction of c/EBPβ is
believed to be a gene regulation mechanism involving an interaction of various
transcription factors.
In essence this study identifies the 5’ flanking region of the Krt1.12 gene responsible for tissue-specific expression, provides evidence of transcription factor
cooperation for tissue-specific gene expression, and potentially elucidates an in vivo transfection technique for controlled gene delivery and expression. This study further
Table 1. Primers Used for DNase I Footprinting
Probe Length (bp) Sense Primer Antisense Primer -599~-375 ACTTCCAAACA GCTGGCTC CGGTTCAGTATAAG CCAACC -394~-131 GGTTGGCTTATA CTGAACCG TGCATCGTGAAAAT TCACTG -331~-109 GGCTTCGGGAA GGATCCTT CCAGCTCAGCAGGT ATGTGA
Table 2. Probes for Electrophoretic Motility Shift Assays
Probe Length (bp) Sequence
-256~-193 ACTGAAGGTGACAGATTCCTTACGACAGCCTA
TCTGCTCCACCCAGCCTTCTTTCTTGTGTGTC
-182~-111 TGGTAATGGTTATTCGATTATAGCTATATCAGTGAAT TTTCACGATGCATAAATCACATACCTGCTGAGCTG
Figure 1 0.2-0.4 KZ 0 100 200 300 400 500 pNASS-beta 0.2 KZ 0.4 KZ 0.6 KZ Constructs
β-gal activity 10-5 U/Sample
Skin Conjunctiva Cornea
Figur e Legend
Figure 1. In vivo β-gal expression by Krt1.12 promoter-β-gal constructs in rabbit
corneas, conjunctivas and skin. 0.6 µm gold particles coated with Krt1.12 constructs
were bombarded onto rabbit corneas at 150 psi, and corneal tissue were collected 48 h
after delivery. 1.6 µm gold particles coated with Krt1.12 constructs were bombarded
onto rabbit bulbar conjunctivas at 150 psi, and conjunctival tissue were collected 48 h
after delivery. 1.6 µm gold particles coated with Krt1.12 constructs were bombarded
onto rabbit dorsal skin at 400 psi (8 mm in diameter), and skin tissue were collected 48
h after delivery. Values are averaged of β-gal activities ± standard deviations per
specimen.
Figure 2. In situ histochemical staining of β-gal in cornea and conjunctiva. Keratin 12 promoter-β-gal constructs were coated onto 0.6 µm gold particles and delivered to
cornea/conjunctival junctions at 200 psi. The tissues were collected at 48 h and
subjected to staining with X-gal as described in METHODS. pNASSβ generates very
few positive reactions in both cornea and conjunctiva. The 0.2 kb generates positive
reactions in both cornea and conjunctiva. The expression of β-gal by the 0.4 kb
constructs of the 5’flanking sequence of Krt1.12 gene is restricted to the cornea.
-394~-131 and -331~-109 were prepared from polymerase chain reactions after
uniquely labeled at the 5-end of the sense primers, and were digested with DNase I as
described in METHOD. The concentration of nuclease was 100 ng/ml and nuclear
protein was 75 µg. Left lanes showed the control reaction without corneal nuclear
protein.
Figure 4. Nuclear extracts from bovine corneal epithelial cells bind regions shown in
footprinting assays. The synthetic double-stranded DNA probes corresponding to
region of –111 bp to –182 bp and –193 bp to –256 bp were labeled with [γ-32P]dCTP
and used in EMSAs. Crude nuclear extracts were prepared from bovine corneal
epithelial cells by direct scrapping and retinal tissue. In both regions, the bindings were
specific and could be competed with unlabeled probe. However, the probes could also
bind the nuclear extracts from retinal tissue.
Figure 5. Schematic of consensus sequence analysis of the sense and antisense regions
(-111 to –182 bp and –193 to –256 bp) of the Krt1.12 gene showing the transcription factor binding sites generated by computer analysis using the NCBI GenBank database
and Blast web client software.
Figure 6. Immunohistochemical staining of mouse eye paraffin sections of central
pre-immune control. Immunoreactivity was detected by immunofluorescence.
Reference List
1. N. C. Andrews and D. V. Faller (5-11-1991) Nucleic Acids Res. 19, 2499.
2. W. J. Boyle, T. Smeal, L. H. Defize, P. Angel, J. R. Woodgett, M. Karin, and T. Hunter (2-8-1991) Cell 64, 573.
3. C. Byrne and E. Fuchs (1993) Mol.Cell Biol. 13, 3176.
4. J. Casatorres, J. M. Navarro, M. Blessing, and J. L. Jorcano (8-12-1994) J.Biol.Chem. 269, 20489.
5. C. Chaloin-Dufau, T. T. Sun, and D. Dhouailly (12-1-1990) Cell Differ.Dev. 32, 97.
6. W. Y. Chen, M. M. Mui, W. W. Kao, C. Y. Liu, and S. C. Tseng (1994) Curr.Eye Res. 13, 765.
7. D. Cooper, A. Schermer, and T. T. Sun (1985) Lab Invest 52, 243.
8. D. Cooper and T. T. Sun (4-5-1986) J.Biol.Chem. 261, 4646.
9. R. L. Eckert, J. F. Crish, and N. A. Robinson (1997) Physiol Rev. 77, 397.
10. R. L. Eckert and J. F. Welter (1996) Mol.Biol.Rep. 23, 59.
11. E. Fuchs (1990) J.Cell Biol. 111, 2807.
12. H. Gille, A. D. Sharrocks, and P. E. Shaw (7-30-1992) Nature 358, 414.
13. S. M. Hollenberg, R. Sternglanz, P. F. Cheng, and H. Weintraub (1995) Mol.Cell Biol. 15, 3813.
14. A. D. Irvine, L. D. Corden, O. Swensson, B. Swensson, J. E. Moore, D. G. Frazer, F. J. Smith, R. G. Knowlton, E. Christophers, R. Rochels, J. Uitto, and W. H. McLean (1997) Nat.Genet. 16, 184.
15. W. W. Kao, C. Y. Liu, R. L. Converse, A. Shiraishi, C. W. Kao, M. Ishizaki, T. Doetschman, and J. Duffy (1996) Invest Ophthalmol.Vis.Sci. 37, 2572.
16. A. Leask, C. Byrne, and E. Fuchs (9-15-1991) Proc.Natl.Acad.Sci.U.S.A 88, 7948.
17. A. Leask, M. Rosenberg, R. Vassar, and E. Fuchs (1990) Genes Dev. 4, 1985.
18. C. Y. Liu, G. Zhu, R. Converse, C. W. Kao, H. Nakamura, S. C. Tseng, M. M. Mui, J. Seyer, M. J. Justice, M. E. Stech, and . (10-7-1994) J.Biol.Chem. 269, 24627.
19. B. Lu, J. A. Rothnagel, M. A. Longley, S. Y. Tsai, and D. R. Roop (3-11-1994) J.Biol.Chem. 269, 7443.
20. R. Marais, J. Wynne, and R. Treisman (4-23-1993) Cell 73, 381.
21. R. Moll, W. W. Franke, D. L. Schiller, B. Geiger, and R. Krepler (1982) Cell 31, 11.
22. P. D. Moyer, A. H. Kaufman, Z. Zhang, C. W. Kao, A. G. Spaulding, and W. W. Kao (1996) Differentiation 60, 31.
23. R. G. Oshima, L. Abrams, and D. Kulesh (1990) Genes Dev. 4, 835.
24. B. J. Pulverer, J. M. Kyriakis, J. Avruch, E. Nikolakaki, and J. R. Woodgett (10-17-1991) Nature 353, 670.
25. R. A. Quinlan, D. L. Schiller, M. Hatzfeld, T. Achtstatter, R. Moll, J. L. Jorcano, T. M. Magin, and W. W. Franke (1985) Ann.N.Y.Acad.Sci. 455, 282.
26. R. H. Rice and H. Green (1977) Cell 11, 417.
27. A. Schermer, S. Galvin, and T. T. Sun (1986) J.Cell Biol. 103, 49.
28. A. Schermer, J. V. Jester, C. Hardy, D. Milano, and T. T. Sun (1989) Differentiation 42, 103.
29. A. Shiraishi, R. L. Converse, C. Y. Liu, F. Zhou, C. W. Kao, and W. W. Kao (1998) Invest Ophthalmol.Vis.Sci. 39, 2554.
30. P. M. Steinert and D. R. Roop (1988) Annu.Rev.Biochem. 57, 593.
31. A. Stoler, R. Kopan, M. Duvic, and E. Fuchs (1988) J.Cell Biol. 107, 427.
32. B. Su, E. Jacinto, M. Hibi, T. Kallunki, M. Karin, and Y. Ben Neriah (6-3-1994) Cell 77, 727.
(1985) Ann.N.Y.Acad.Sci. 455, 307.
34. Y. Takemoto, Y. Fujimura, M. Matsumoto, Y. Tamai, T. Morita, A. Matsushiro, and M. Nozaki (5-25-1991) Nucleic Acids Res. 19, 2761.
35. Y. Tamai, Y. Takemoto, M. Matsumoto, T. Morita, A. Matsushiro, and M. Nozaki (8-15-1991) Gene 104, 169.
36. Z. G. Wei, R. L. Wu, R. M. Lavker, and T. T. Sun (1993) Invest Ophthalmol.Vis.Sci. 34, 1814.
37. R. A. Weiss, R. Eichner, and T. T. Sun (1984) J.Cell Biol. 98, 1397.
38. R. L. Wu, S. Galvin, S. K. Wu, C. Xu, M. Blumenberg, and T. T. Sun (1993) J.Cell Sci. 105 ( Pt 2), 303.
39. R. L. Wu, G. Zhu, S. Galvin, C. Xu, T. Haseba, C. Chaloin-Dufau, D. Dhouailly, Z. G. Wei, R. M. Lavker, W. Y. Kao, and . (1994) Differentiation 55, 137.