Journal of Ethnopharmacology 115 (2008) 455–462
Suppression of lipopolysaccharide-induced of inducible nitric oxide
synthase and cyclooxygenase-2 by Sanguis Draconis,
a dragon’s blood resin, in RAW 264.7 cells
Cheuk-Sing Choy
a,b, Chien-Ming Hu
a,b, Wen-Ta Chiu
c, Carlos-Shu Kei Lam
a,
Yih Ting
a, Shin-Han Tsai
a, Tzu-Chien Wang
a,∗aEmergency Department of Municipal Wan Fang Hospital, Taipei Medical University, Taipei, Taiwan, ROC bEmergency Department of Taipei Medical University Hospital, Taipei, Taiwan, ROC
cNeurosurgery Department of Municipal Wan Fang Hospital, Taipei Medical University Hospital, Taipei, Taiwan, ROC
Received 4 May 2007; received in revised form 8 October 2007; accepted 14 October 2007 Available online 22 October 2007
Abstract
Sanguis Draconis (SD) is a kind of dragon’s blood resin that is obtained from Daemomorops draco (Palmae). It is used in traditional medicine and has shown anti-inflammatory activity in some diseases. In this study, we examined the effects of Sanguis Dranonis ethanol extract (SDEE) on LPS-induced inflammation using RAW 264.7 cells. Our data indicated that SDEE inhibits LPS-stimulated NO, PGE2, IL-1 and TNF-␣ release,
and iNOS and COX-2 expression. Furthermore, SDEE suppressed the LPS-induced p65 expression of NF-B, which was associated with the inhibition of IB-␣ degradation. We also found that the expression of HO-1 was significantly increased in RAW 264.7 cells by SDEE. These results suggest among possibilities of anti-inflammation that SDEE inhibits the production of NO and PGE2by the down-regulation of iNOS and COX-2
gene expression via the suppression of NF-B (p65) activation. SDEE can induce HO-1 over-expression in macrophage cells, which indicates that it may possess antioxidant properties. This result means that SEDD its anti-inflammatory effects in macrophages may be through a novel mechanism that involves the action of HO-1. Thus, SD could provide a potential therapeutic approach for inflammation-associated disorders. © 2007 Elsevier Ireland Ltd. All rights reserved.
Keywords: Sanguis Draconis; Lipopolysaccharide; Inducible nitric oxide synthase; Cyclooxygenase-2; Nuclear factor-B
1. Introduction
Inflammation is the normal physiological and immune responses to tissue injury. An increased supply of blood, enhanced vascular permeability and migration of immune cells occur at damaged sites. The inflammatory process is a protec-tive response that occurs in response to trauma, infection, tissue
Abbreviations: COX-2, cyclooxygenase-2; eNOS, endothelial nitric oxide
synthase; HO-1, heme oxgenase-1; IB-␣, inhibitory factor B-␣; IL-1, interlukin-1; iNOS, inducible nitric oxide synthase; LPS, lipopolysaccharide; NF-B, nuclear factor-B; NO, nitric oxide; nNOS, neuronal nitric oxide syn-thase; PGE2, prostaglandin E2; SD, Sanguis Draconis; SDEE, Sanguis Draconis
ethanol extract; TNF-␣, tumor necrosis factor-␣.
∗Corresponding author at: Emergency Department of Municipal Wan Fang
Hospital, NO. 111, Section 3, Hsing-Long Road, Taipei Medical University, Taipei 116, Taiwan, ROC. Tel.: +886 2 29307930x1278; fax: +886 2 86621104.
E-mail address:[email protected](T.-C. Wang).
injury or noxious stimuli (Zedler and Faist, 2006; Mariathasan and Monack, 2007). In this process, activated inflammatory cells (neutrophils, eosinophils, mononuclear phagocytes and macrophages) secrete increased amounts of NO, prostaglandin and cytokines, such as IL-1, TNF-␣, IFN-␥ and IL-10 (Vane et al., 1994). All things considered, these substances not only induce cell and tissue damage but also activate macrophages in rheumatoid arthritis (Cheon et al., 2006), chronic hepatitis (Wolf et al., 2005), etc. NO is a major product which is controlled by nitric oxide synthases (NOS), such as iNOS, eNOS and nNOS (Marletta, 1993). Most importantly, iNOS is highly expressed in macrophages, which leads to organ destruction in some inflam-matory and autoimmune diseases (Kleinert et al., 2004). PGE2is also another important mediator which is produced from arachi-donic acid metabolites which are catalyzed by cyclooxygenase-2 (COX-2) in inflammatory responses (Harris et al., 2002).
LPS is an endotoxin, an integral outer membrane component of Gram-negative bacteria, and triggers the most potent micro-0378-8741/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved.
456 C.-S. Choy et al. / Journal of Ethnopharmacology 115 (2008) 455–462
bial initiators of inflammatory response, such as septic shock, fever and microbial invasion (Dobrovolskaia and Vogel, 2002). Generally, the hydrophobic lipid A region of LPS combines with Toll-like receptors and transduces signals into intracel-lular signaling that induce an inflammatory response and the activation of transcription factors such as NF-B, interferon regulatory factors-3 and activating protein-1 (Wright, 1999; Verstak et al., 2007). NF-B is a small protein of intimately governed transcription, and NF-B plays a primary role in the transcriptional regulation of iNOS, COX-2 and TNF-␣ gene expressions (Chen et al., 2002). Nowadays, NF-B has been identified as a heterotrimeric complex composed of p50, p65 and IB-␣, and hides in the cytoplasma of unstimulated cells (Baeuerle and Baltimore, 1996). The phosphorylation of IB-␣ by IB kinase and polyubiquitination by ubiquitin protein ligases leads to NF-B degradation for the formation of active NF-B (p50·p65) dimers. Following the above step, the active NF-B translocates into the nucleus and promotes the transcrip-tion of target genes and initiates gene expression (Ghosh and Karin, 2002).
It is notable that more and more natural plant products are being used as medicine to prevent some acute or chronic dis-eases throughout the world. Dragon’s blood is a kind of red resin that is obtained from several botanical origins, such as Dae-monorops (Palmae), Dracaena (Convallariaceae) and Croton (Euphorbiaceae). Most dragon’s blood that is traded interna-tionally is from Daemonorops (Edwards et al., 2004). Although dragon’s blood is obtained from different species of plants, it was used in the treatment of diarrhea, wounds, tumors, stomach ulcers, and herpes infection, in the itching, pain and swelling of insect bites, rheumatism and with other conditions in the folk medicine (Jones, 2003). In ethnopharmacological studies, the effects of dragon’s blood on various biological activities, such as attenuate visceral nociception, antiviral, antibacterial and antifungal have been reported (Peres et al., 1997; Gurgel et al., 2001; Rao et al., 2007). The pharmaceutical name of dragon’s blood is called Sanguis Draconis (SD) and it was a famous material used for various medical and artistic purposes. The chemical composition of SD probably involves dracoru-bin, dracorhodin (Fig. 1A), dracoresinotannol, abietic acid, etc. The other compositions of SD such as cinnabarone, dracore-sene, dracoresinotannol and benzoic acid have been investigated in detail (Edwards et al., 2004). It is generally believed that a pharmaceutical preparation that is the red resinous excretion from the fruit and stem is harvested during the summer. And it is vapoured or heated into solid resin, then smashed into pow-der. SD is non-odorous and tasteless, and insoluble in water, but soluble in alcohol and ether (Lu et al., 2003). SD fulfil many medical functions, such as improving blood circulation, stop-ping haemorrhages, and healing wounds and cuts, and is also used as an antiseptic (Chen et al., 2003). However, the possible mechanism responsible for its anti-inflammatory effects is still unknown.
In the present study, we investigated the effects of SDEE on LPS-induced inflammatory responses (NO and PGE2release) and further explored the possible mechanisms of these inhibi-tions by SDEE in the macrophage cell line RAW 264.7.
2. Methods
2.1. Plant material
Commercially available plant material (Sanguis Draconis, SD) were purchased from a traditional Chinese medicine drug store and its botanical sources are from Daemonorops draco (Palmae) and the import of habitat was from Malaysia, India and Iran. The authenticity of SD was confirmed by Professor Hui-Wen Cheng of School of Pharmacy, Taipei Medical Uni-versity. A voucher of source was kept at the traditional Chinese medicine drug store and a voucher specimen was deposited at our lab. SD (10 g) was soaked in 200 ml of absolute alcohol at room temperature for 48 h under shade and then was filtered. Ethanolic extract was passed through a 0.22m sterile filter (Millipore; Billerica, MA) and first concentrated using a Yamato vacuum rotary evaporator (Japan) at 40◦C and then freeze-dried at −80◦C in a vacuum freeze-dryer (MicroModulyo, Savant Instruments, USA). Normally, 1.57 g of dried powder can be obtained from 10 g of SD. Its main constituents were reported byEdwards et al. (2004), as shown inFig. 1A.
2.2. Chemicals
Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum (FBS), penicillin and streptomycin were obtained from Life Technologies Inc. (Gibco, Grand Island, NY). iNOS, COX-2, IB-␣, NF-B, -actin monoclonal antibodies and the peroxidase-conjugated secondary antibody were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA). The enzyme immunoassay kits for PGE2, TNF-␣, and IL-1 were purchased from R&D System (Minneapolis, MN). Escherichia coli lipopolysaccharide (LPS), Griess reagent, and 3-(4,5-dime-thylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) were obtained from Sigma Chemical Co. (St. Louis, MO, USA). 2.3. Cell culture
The RAW 264.7 murine macrophage cell line was obtained from the American Type Culture Collection (Rockville, MD). Cells were cultured in DMEM medium with 10% FBS, peni-cillin (100 units/ml) and streptomycin sulfate (100g/ml) in 95% air, and 5% CO2 humidified atmosphere at 37◦C. Cells were counted with a hemocytometer, and the number of viable cells was determined by trypan blue dye exclusion. The cells were incubated with SDEE at various concentrations and treated with LPS 100 ng/ml.
2.4. MTT assay for cell viability
Cell viability was assessed by the mitochondrial respiration-dependent reduction method of MTT to formazan. Cells (1× 104cells/well) in 96-well plates were incubated with var-ious concentrations of SDEE (1, 3, 10, 30, 100, 200, 300 and 1000g/ml) at 37◦C in 5% CO2for 24 h. After treatment, 50l of DMEM medium containing MTT (5 mg/ml) was added to each well, followed by incubation for 4 h at 37◦C. After
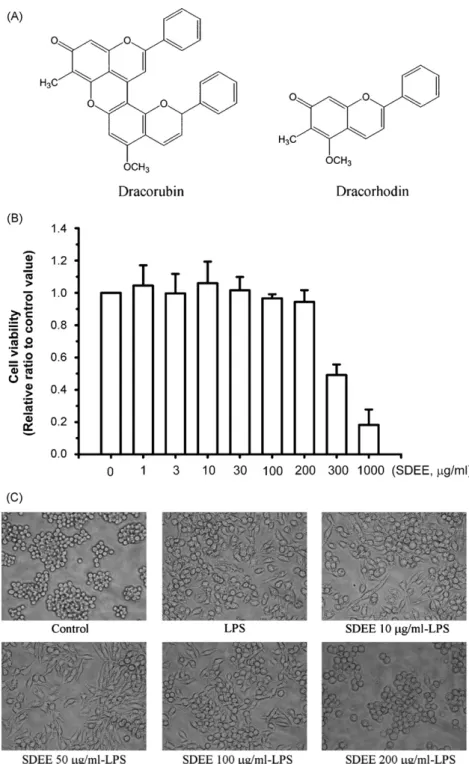
incu-Fig. 1. Effects of SDEE on the viability of a RAW 264.7 murine macrophage cell line. (A) Chemical structures of dracorubin and dracorhodin, the main chemical components of Sanguis Draconis (dragon’s blood resins) from Daemonorops draco. Note: The two structural pictures were acquired fromEdwards et al. (2004). Panel (B) Cells were treated with various concentrations of SDEE for 24 h. Cell viability was determined by MTT assay as described in Section2. (C) Morphology of the LPS-stimulated inflammatory response in RAW264.7 cells in the presence or absence of SDEE for 24 h. Cells were viewed using a video camera (Nikon) attached to a microscope and projected onto a monitor. Images were obtained with a 200× objective. The results are expressed as mean ± S.E. from three independent experiments.
bation, the medium was discarded and the formazan crystals in viable cells were dissolved in DMSO 100l for 10 min. The optical density of each well was measured at 540 nm.
2.5. Determination of nitrite production
As an indicator of NO production, nitrite concentration in the culture medium was determined by Griess reagent (Di Rosa et
al., 1996; Green et al., 1982). The culture supernatant (100l) was mixed with Griess reagent (100l, 1% sulphanilamide, 0.1% N-1-naphthyl ethylenediamine) for 10 min, and then the absorbance of the chromophoric azo-derivative molecule was measured in a microplate reader at 540 nm. Fresh culture medium was used as the blank in all experiments. A range of dilutions of sodium nitrite was used to a standard curve with the amount of nitrite in each sample.
458 C.-S. Choy et al. / Journal of Ethnopharmacology 115 (2008) 455–462
2.6. Measurement of PGE2, TNF-α and IL-1β
The levels of PGE2, TNF-␣ and IL-1 in the RAW 264.7 cell culture medium were measured by enzyme immunoassay (EIA) kits according to the manufacturer’s instructions. The intra- and inter-assay coefficients of variation (CV, %) are recorded fol-lowing: PGE2, 8.5 and 8.8; TNF-␣, 6.6 and 7.5; IL-1, 2.7 and 4.2. The minimum detectable dose of PGE2, TNF-␣ and IL-1 is typically less than 27.5, 5.1 and 3.0 pg/ml, respectively. 2.7. Preparation of total cell lysates
RAW 264.7 cells (5× 105cells/well) in six-well plates were incubated with or without concentrations of SDEE and LPS (100 ng/ml) for 24 h. The total cell lysates were obtained by lysis buffer (250 mM Tris–HCl (pH 6.8), 1% Triton-100, 0.1% SDS, 1 mM Na3VO4, 1 mM EDTA, 5 mM sodium fluoride, 1 mM PMSF, 1g/l leupetin), and cell debris was removed to cen-trifuge at 10,000× g for 10 min at 4◦C and stored at−80◦C until required. The protein content of cell lysates was determined by Bradford assay.
2.8. Preparation of nuclear and cytosolic extract
The nuclear extract was prepared according to Pokharel’s method (2006). Briefly, RAW 264.7 cells (5× 105cells/well) in six-well plates were incubated with or without concentrations of SDEE for 2 h, and then treated with LPS (100 ng/ml) for 30 min. After LPS treatment, cells were removed using a cell scraper and washed with ice-cold PBS, and then centrifuged at 2500× g for 5 min at 4◦C. The cells were then added to 100l lysis buffer (10 mM HEPES pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.5% Nonidet-P40, 1 mM dithioreitol, 0.5 mM PMSF) and vor-texed mildly. Samples were incubated for 10 min on ice and then centrifuged at 2500× g for 5 min at 4◦C. The supernatant was collected and pellets containing crude nuclei were resuspended in 100l extraction buffer (20 mM HEPES pH 7.9, 400 mM NaCl, 1 mM EDTA, 1 mM dithioreitol, 1 mM PMSF) and incu-bated for 30 min on ice, and then centrifuged at 15,800× g for 10 min. The supernatant containing nuclear extracts was col-lected and stored at−80◦C until required.
2.9. Western blot analysis
Equal amounts of cell lysates (30–50g) were electroblot-ted onto a nitrocellulose membrane (Millipore, MA), following separation using 8–12% SDS-polyacrylamide gel electrophore-sis. The transblotted membranes were washed twice with PBS containing 0.1% Tween 20 (PBST) and incubated with block-ing buffer containblock-ing 5% nonfat milk for 1 h. The membranes were incubated overnight at 4◦C with a 1:1000 dilution of primary antibody as anti-iNOS, COX-2, NF-B, IB-␣ and -actin. Blots were washed three times with PBST for 15 min and incubated with a 1:2000 dilution of horseradish peroxidase-conjugated secondary antibody (Santa Cruz, CA) for 1 h at 37◦C. Blots were again washed three times in PBST and developed for visualization using an ECL Plus detection kit
(Pierce Biotechnology Inc., USA). The intensity of each band was quantitatively determined using density analysis software (MetaMorph Imaging System, Meta Imaging Series 4.5), and the density ratio represented the relative intensity of each band against those of controls in each experiment.
2.10. Data and statistical analysis
The results of all experiments were expressed as the mean± S.E. of triple experiments. Data were compared using the Student’s t-test, and P-values less than 0.05 were considered statistically significant.
3. Results
3.1. Effect of SDEE on cell viability
Initially, the cytotoxicity of SDEE to RAW 264.7 cells was measured by MTT assay. As shown inFig. 1B, cell viability was not significantly altered by SDEE at up to 200g/ml. The cell morphology was compared with or without treated LPS 100 ng/ml for 24 h; however, treatment with SDEE 200g/ml can reverse LPS-induced alteration of cell morphology (Fig. 1C). These results suggest that concentrations of SDEE were not toxic to RAW 264.7 cells below 200g/ml, at least. Thus, cells were treated with SDEE in the concentration range 10–200g/ml during follow-up experiments.
3.2. Inhibitory effect of SDEE on LPS-induced NO, PGE2,
TNF-α and IL-1β production
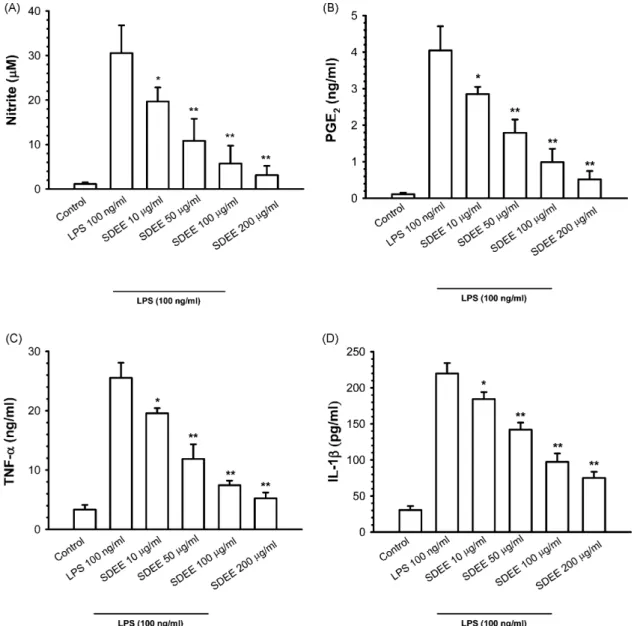
To assess the effects of SDEE on LPS-induced NO, PGE2, TNF-␣ and IL-1 production in RAW 264.7 cells, cell culture media which were treated with or without LPS 100 ng/ml and various concentrations of SDEE were collected and stored at −80◦C. Nitrite production was measured using Griess reagent, and PGE2, TNF-␣ and IL-1 release were determined by EIA kits. As our results show, LPS stimulation caused a signif-icant accumulation of NO in the culture medium; however, pretreatment with SDEE 10 (P < 0.05), 50, 100 and 200g/ml (P < 0.01) significantly attenuated LPS-induced nitrite produc-tion in a dose-dependent manner (Fig. 2A). Since PGE2, TNF-␣ and IL-1 are other inflammatory mediators, the effects of SDEE on PGE2, TNF-␣ and IL-1 production in LPS-stimulated RAW 264.7 cells were measured by culture medium. Consistent with the NO accumulation, coincubation of cells with 10, 50, 100 and 200g/ml of SDEE prior to LPS treatment caused a significant (P < 0.05 or P < 0.01) decrease in a dose-dependent manner in the release of PGE2, TNF-␣ and IL-1 (Fig. 2B–D).
3.3. Effect of SDEE on LPS-induced iNOS and COX-2 protein expression
Western immunoblot analyses were performed to determine whether the inhibitory effects of SDEE on pro-inflammatory mediators, such as NO, PGE2, TNF-␣ and IL-1, are related to changes in the protein levels of iNOS and COX-2, respectively.
Fig. 2. Effects of SDEE on LPS-stimulated inflammatory mediators production in RAW 264.7 cells. (A) Cells were treated with 10, 50, 100 and 200g/ml of SDEE for 2 h, followed by 24 h culture in the presence or absence of 100 ng/ml LPS. The nitrite concentration in the culture medium was determined by Griess reagent. (B–D) PGE2, TNF-␣ and IL-1 production were determined by ELISA kit, as described in Section2. Data are expressed as mean± S.E. from three independent
experiments. Asterisks indicate a significant difference for LPS alone (*P < 0.05,**P < 0.01).
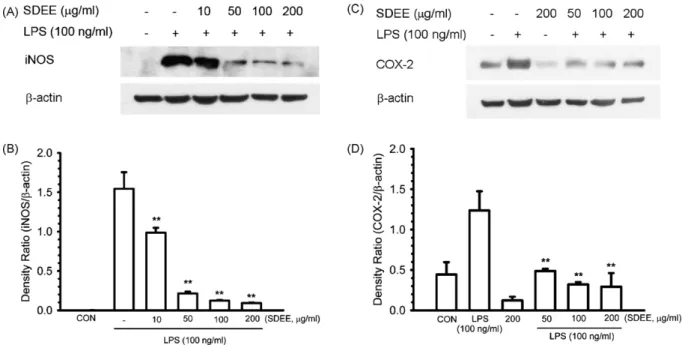
The protein expression of iNOS and COX-2 was obviously increased by stimulation with LPS, and SDEE markedly inhib-ited these protein inductions in a dose-dependent manner (Fig. 3A and C). In quantitative determination,-actin levels as an internal standard were comparable among each sample. The density ratio of iNOS and COX-2 was significantly (P < 0.01) suppressed by SDEE in LPS-induced RAW 264.7 cells (Fig. 3B and D).
3.4. Effect of SDEE on LPS-induced NF-κB expression at translation level
In order to understand the mechanisms underlying the inhibi-tion of iNOS protein expression by SDEE, we further measured the protein level of NF-B (p65) in the nucleus by Western blot-ting. As shown in our data, nuclear p65 protein levels of RAW
264.7 cells were increased after treating cells with LPS at 30 min, and the level of p65 declined in response to LPS pre-treatment with SDEE (10, 50,100 and 200g/ml) for 2 h (Fig. 4A). It is clear that the density ratio of LPS-induced p65 was significantly (P < 0.01) inhibited in a dose-dependent manner by SDEE in the quantitative determination (Fig. 4B).
3.5. Inhibition of SDEE on LPS-induced IκB-α degradation To examine whether SDEE inhibits LPS-induced IB-␣ degradation, we prepared cytoplasmic extracts of RAW 264.7 cells and determined IB-␣ levels by Western blotting. As shown in Fig. 5A, IB-␣ was degraded after treatment with LPS in RAW 246.7 cells for 30 min, and this degradation was markedly inhibited by pre-treatment with SDEE (50,100 and 200g/ml) for 2 h. In addition, we found that SDEE significantly (P < 0.05
460 C.-S. Choy et al. / Journal of Ethnopharmacology 115 (2008) 455–462
Fig. 3. Effects of SDEE on LPS-induced iNOS and COX-2 protein expression. (A and C) Cells (5× 105cells/well) were pretreated with the indicated concentrations of SDEE for 2 h before incubation with 100 ng/ml LPS for 24 h. Proteins (30g) of total cell lysates were analyzed on 8% and 10% SDS-PAGE for anti-iNOS and COX-2, respectively. Immunoblotting was determined as described in Material and methods. (B and D) The blot was stripped of the bound antibody and reprobed with anti- actin to confirm equal loading. Quantification of iNOS and COX-2 expression was performed by densitometric analysis as described in Section2. The relative density was calculated as the ratio of iNOS and COX-2 expression to-action expression. Data are expressed as mean ± S.E. from three independent experiments. Asterisks indicate a significant difference for LPS alone (**P < 0.01).
Fig. 4. Inhibition of the nuclear translocation by SDEE in LPS-stimulated RAW 264.7 cells. (A) Cells (5× 105cells/well) were pretreated with the indicated
con-centrations of SDEE for 2 h before incubation with 100 ng/ml LPS for 30 min. Nuclear extract proteins (50g) were analyzed on 10% SDS-PAGE for anti-NF-B (p65). Immunoblotting was determined as described in Section2.-Actin was used as a loading control. (B) Quantification of NF-B (p65) expression was performed by densitometric analysis as described in Section2. The relative density was calculated as the ratio of NF-B (p65) expression to -action expres-sion. Data are expressed as mean± S.E. from three independent experiments. Asterisks indicate a significant difference for LPS alone (**P < 0.01).
Fig. 5. Effect of SDEE on LPS-stimulated IB-␣ degradation in RAW 264.7 cells. (A) Cells (5× 105cells/well) were pretreated with the indicated
concen-trations of SDEE for 2 h before incubation with 100 ng/ml LPS for 30 min. Cytosolic extract proteins (50g) were subjected on 12% SDS-PAGE to West-ern immunoblot analysis with anti-IB-␣. -Actin was used as a loading control. (B) Quantification of IB-␣ degradation was performed by densitometric anal-ysis as described in Section2. The relative density was calculated as the ratio of IB-␣ degradation to -action expression. Data are expressed as mean ± S.E. from three independent experiments. Asterisks indicate a significant difference for LPS alone (**P < 0.01).
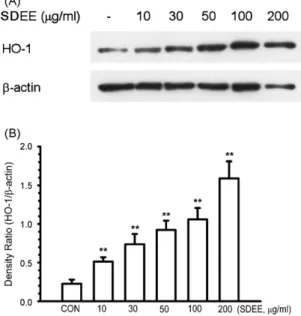
Fig. 6. Effect of SDEE on HO-1 expression in RAW 264.7 cells. (A) Cells (5× 105cells/well) were treated with 10, 30, 50, 100 and 200g/ml SEDD for
24 h. Total cell lysates (30g proteins) were analyzed on 12% SDS-PAGE for anti-HO-1, and immunoblotting was performed as described in Section2. (B) Quantification of HO-1 expression was performed by densitometric analysis as described in Section2. The relative density was calculated as the ratio of HO-1 expression to-action expression. Data are expressed as mean ± S.E. from three independent experiments. Asterisks indicate a significant difference for LPS alone (**P < 0.01).
or P < 0.01) had an inhibitory effect on the LPS-induced degra-dation of IB-␣ in a dose-dependent manner, by quantitative determination (Fig. 5B). These results suggest that SDEE inhib-ited the NF-B translocation to the nucleus through prevention of IB-␣ degradation.
3.6. SDEE induced HO-1 protein expression
RAW 264.7 cells were treated with various concentrations of SDEE (10, 30, 50, 100 and 200g/ml) for 24 h. Cells were harvested by lysis buffer and the total cell lysates were collected; the HO-1 protein level was determined by Western immunoblot analysis. As shown inFig. 6A and B, we unexpectedly found that SDEE significantly (P < 0.01) increased HO-1 protein levels in a dose-dependent manner in RAW 264.7 cells.
4. Discussion
Natural products have played a significant role in drug discovery and development, especially agents against several diseases that have existed from antiquity to the present. In this study, we prepared ethanol extract from SD (SDEE) and exam-ined its effects on the LPS-induced inflammation in a murine macrophage cell line RAW 264.7 model. First, we evaluated the cytotoxicity of SDEE in RAW 264.7 cells by MTT assay, and it was observed that SDEE did not affect cell viability below 200g/ml. In addition, microscopic photos of RAW 264.7 cells showed that the cellular morphology in the LPS-stimulated inflammatory response could be recovered by treatment with a
higher concentration (200g/ml) of SDEE. This result hints that SDEE certainly has some effects in LPS-stimulated inflamma-tion of RAW 264.7 cells.
The massive amounts of NO produced in response to bacte-rial LPS or cytokines play an important role in the inflammatory condition (McCann et al., 2005). The production of PGE2, TNF-␣ and IL-1 is a crucial part of the immune response to some inflammatory stimuli. For example, excessive production of these mediators could be detected in septic and hemorrhagic shock (Jarrar et al., 2002), rheumatoid arthritis, atherosclero-sis, etc. Thus, drug designs that decrease NO production by inhibiting the iNOS gene, major receptors for signaling initi-ated by LPS, or enzyme activity, have a therapeutic effect in the treatment of septic shock, as well as many other inflammatory and infectious disorders (Zhang and Ghosh, 2000). Furthermore, there are many reports of non-steroidal anti-inflammatory drugs inhibiting the production of NO and iNOS by the suppression NF-B activation (DiGirolamo et al., 2003; Tanaka et al., 2002). According to our results, SDEE suppresses the production of NO, PGE2, TNF-␣ and IL-1, and decreases iNOS and COX-2 protein expression levels in LPS-induced RAW COX-264.7 cells. The mechanism by which SDEE inhibits the expression of these mediators seems to involve the inhibition of NF-B activation. These findings suggest that SDEE may prevent inflammation by suppressing the NF-B-mediated inflammatory gene.
It is known that iNOS expression is regulated mainly at the transcriptional level, and p50 and p65 are two major proteins in NF-B activation by LPS in macrophages (Ghosh and Karin, 2002). In the unstimulated condition, NF-B is masked by the inhibitory protein, IB-␣, so that the inactive NF-B cannot translocate into the nucleus (Baldwin, 1996). Following the acti-vation of NF-B by various agents, such as LPS, IL-1, TNF-␣ or cellular cytokine activators, etc., NF-B requires phospho-rylation of IB-␣, which then targets IB-␣ for ubiquitination and degradation (Baeuerle, 1998). After this step, active NF-B is translocated into the nucleus where it activates a variety of genes, including IB-␣. In our data, we found that SDEE inhib-ited the cytosol IB-␣ protein degradation and nuclear NF-B protein expression in LPS-stimulated RAW 264.7 cells. Based on these results, we can ensure that the LPS-stimulated expres-sions of the iNOS and COX-2 genes were inhibited by SDEE possibly through blocking NF-B activation. However, we can-not exclude the possible involvement of the inhibition of other transcription factors.
We also found that hemeoxyganse-1 (HO-1) could be induced by treatment with SDEE. HO-1 seems to be a novel pro-tective gene with potent anti-inflammatory, antioxidant, and antiproliferative effects (Exner et al., 2004). Moreover, the antioxidants, such as polyphenols and flavanols, inhibit the pro-duction of inflammatory mediators through suppression of their gene expression and prevent inflammation (Rahman et al., 2006; Selmi et al., 2006). Accordingly, the inflammatory effect of SDEE by activating HO-1 expression may be involved. Further study is needed to determine the detailed mechanism of SDEE in its antioxidant anti-inflammatory activity.
In this study, we demonstrated that SDEE inhibits the production of NO, PGE2, TNF-␣ and IL-1 in
LPS-462 C.-S. Choy et al. / Journal of Ethnopharmacology 115 (2008) 455–462
stimulated macrophages. Among possible pathway in this anti-inflammatory effect occurs by inhibiting iNOS and COX-2 expression via the blocking of IB-␣ degradation and NF-B activation may be involved. Therefore, SDEE might be a poten-tial therapeutic drug for clinical use in inflammatory diseases.
Acknowledgments
The authors wish to thank Prof. Jaw-Jou Kang (Institute of Toxicology, College of Medicine, National Taiwan University, Taipei, Taiwan, ROC) and Hui-Yi Lin PhD (Institute of Biomed-ical Sciences, Academia Sinica, Taipei, Taiwan, ROC) for their comments on this study.
References
Baeuerle, P.A., 1998. IB-NF-B structures: at the interface of inflammation control. Cell 95, 729–731.
Baeuerle, P.A., Baltimore, D., 1996. NF-B: ten years after. Cell 87, 13–20. Baldwin Jr., A.S., 1996. The NF-B and IB proteins: new discoveries and
insights. Annual Review of Immunology 14, 649–683.
Chen, F., Demers, L.M., Shi, X., 2002. Upstream signal transduction of NF-B activation. Current Drug Targets. Inflammation and Allergy 1, 137–149. Chen, H.H., Sun, C.C., Tseng, M.P., Hsu, C.J., 2003. A patch test study of
27 crude drugs commonly used in Chinese topical medicaments. Contact Dermatitis 49, 8–14.
Cheon, H., Rho, Y.H., Choi, S.J., Lee, Y.H., Song, G.G., Sohn, J., Won, N.H., Ji, J.D., 2006. Prostaglandin E2augments IL-10 signaling and function. Journal
of Immunology 15, 1092–1100.
Di Rosa, M., Lalenti, A., Ianaro, A., Sautebin, L., 1996. Interaction between nitric oxide and cyclooxygenase pathways. Prostaglandins Leukocyte Essen-tail Fatty Acids 54, 229–238.
DiGirolamo, G., Farina, M., Riberio, M.L., Ogando, D., Aisemberg, J., Santos, A.R., de, los., Marti, M.L., Franchi, A.M., 2003. Effects of cyclooxygenase inhibitor pretreatment on nitric oxide production, nNOS and iNOS expres-sion in rat cerebellum. British Journal of Pharmacology 139, 1164–1170. Dobrovolskaia, M.A., Vogel, S.N., 2002. Toll receptors, CD14, and macrophage
activation and deactivation by LPS. Microbes and Infection 4, 903–914. Edwards, H.G.M., Oliveira, L.F.C., Prendergast, H.D.V., 2004. Raman
spectro-scopic analysis of dragon’s blood resins-basis for distinguishing between
Dracaena (Convallariaceae), Daemonorops (Palmae) and Croton
(Euphor-biaceae). Analyst 129, 134–138.
Exner, M., Minar, E., Wagner, O., Schillinger, M., 2004. The role of heme oxygenase-1 promoter polymorphisms in human disease. Free Radical Biol-ogy & Medicine 37, 1097–1104.
Ghosh, S., Karin, M., 2002. Missing pieces in the NF-B puzzle. Cell 109, 81–96.
Green, L.C., Wanger, D.A., Glogowski, J., Skipper, P.L., Wishnok, J.S., Tannen-baum, S.R., 1982. Analysis of nitrate, nitrite, and [15N]nitrate in biological
fluids. Analytical Biochemistry 126, 131–138.
Gurgel, L.A., Sidrim, J.J., Martins, D.T., Cechinel Filho, V., Rao, V.S., 2001. In vitro antifungal activity of dragon’s blood from Croton urucurana against dermatophytes. Journal of Ethnopharmacology 97, 409–412.
Harris, S.G., Padilla, J., Koumas, L., Ray, D., Phipps, R.P., 2002. Prostaglandins as modulators of immunity. Trends in Immunology 23, 144–150. Jarrar, D., Kuebler, J.F., Rue, L.W., Matalon, S., Wang, P., Bland, K.I., Chaudry,
I.H., 2002. Alveolar macrophage activation after trauma-hemorrhage and
sepsis is dependent on NF-B and MAPK/ERK mechanisms. America Journal of Physiology. Lung Cellular and Molecular Physiology 283, L799–L805.
Jones, K., 2003. Review of sangre de drago (Croton lechleri)—a South Amer-ican tree sap in the treatment of diarrhea, inflammation, insect bites, viral infections, and wounds: traditional uses to clinical research. Journal Alter-native and Complementary Medicine 9, 877–896.
Kleinert, H., Pautz, A., Linker, K., Schwarz, P.M., 2004. Regulation of the expression of inducible nitric oxide synthase. European Journal of Phar-macology 500, 255–266.
Lu, H., Teng, J., Wu, H., 2003. A review on the study of Resina Draconis produced in China. Journal of Chinese Medicinal Materials 26, 459, 461. Mariathasan, S., Monack, D.M., 2007. Inflammasome adaptors and sensors:
intracellular regulators of infection and inflammation. Nature Reviews Immunology 7, 31–40.
Marletta, M.A., 1993. Nitric oxide synthase structure and mechanism. The Journal of Biological Chemistry 268, 12231–12234.
McCann, S.M., Mastronardi, C., Laurentiis, A.de., Rettori, V., 2005. The nitric oxide theory of aging revisited. Annals of the New York Academy of Sci-ences 1057, 64–84.
Peres, M.T.L.P., Delle Monache, F., Cruz, A.B., Pizzolatti, M.G., Yunes, R.A., 1997. Chemical composition and antimicrobial activity of Croton urucurana Baillon (Euphorbiaceae). Journal of Ethnopharmacology 56, 223–226. Pokharel, Y.R., Yang, J.W., Kim, J.Y., Oh, H.W., Jeong, H.G., Woo, E.R.,
Kang, K.W., 2006. Potent inhibition of the inductions of inducible nitric oxide synthase and cyclooxygenase-2 by taiwaniaflavone. Nitric Oxide 15, 217–225.
Rahman, I., Biswas, S.K., Kirkham, P.A., 2006. Regulation of inflammation and redox signaling by dietary polyphenols. Biochemical Pharmacology 72, 1439–1452.
Rao, V.S., Gurgel, L.A., Lima-J´unior, R.C., Martins, D.T., Cechinel-Filho, V., Santos, F.A., 2007. Dragon’s blood from Croton urucurana (Baill.) atten-uates visceral nociception in mice. Journal of Ethnopharmacology 113, 357–360.
Selmi, C., Mao, T.K., Keen, C.L., Schmitz, H.H., Eric, G.M., 2006. The anti-inflammatory properties of cocoa flavanols. Journal of Cardiovascular Pharmacology 47, S163–S171.
Tanaka, A., Hase, S., Miyazawa, T., Ohno, R., Takeuchi, K., 2002. Role of cyclooxygenase (COX)-1 and COX-2 inhibition in nonsteroidal anti-inflammatory drug-induced intestinal damage in rats: relation to various pathogenic events. The Journal of Pharmacology and Experimental Thera-peutics 303, 1248–1254.
Vane, J.R., Mitchell, J.A., Appleton, I., Tomlinson, A., Bishop-Bailey, D., Crox-tall, J., Willoughby, D.A., 1994. Inducible isoforms of cyclooxygenase and nitric-oxide synthase in inflammation. Proceedings of the National Academy of Sciences of the United States of America 91, 2046–2050.
Verstak, B., Hertzog, P., Mansell, A., 2007. Toll-like receptor signalling and the clinical benefits that lie within. Inflammation Research 56, 1–10. Wolf, A.M., Wolf, D., Rumpold, H., Ludwiczek, S., Enrich, B., Gastl, G., Weiss,
G., Tilg, H., 2005. The kinase inhibitor imatinib mesylate inhibits TNF-␣ production in vitro and prevents TNF-dependent acute hepatic inflammation. Proceedings of the National Academy of Sciences of the United States of America 20, 13622–13627.
Wright, S.D., 1999. Toll, a new piece in the puzzle of innate immunity. The Journal of Experimental Medicine 189, 605–609.
Zedler, S., Faist, E., 2006. The impact of endogenous triggers on trauma-associated inflammation. Current Opinim in Critical Care 12, 595–601. Zhang, G., Ghosh, S., 2000. Molecular mechanisms of NF-B activation induced
by bacterial lipopolysaccharide through Toll-like receptors. Journal of Endo-toxin Research 6, 453–457.