Protocatechuic Acid from Alpiniaoxyphylla Induces Schwann Cell Migration via ERK1/2, JNK and p38 Activation
Da-Tong Ju1, Wei-Wen Kuo2, Tsung-Jung Ho3,4, Catherine Reena Paul5, Chia-Hua
Kuo6, Fuu-Jen Tsai3, Chien-Chung Lin7, Yueh-Sheng Chen3, Yung-Ming
Chang8,*,Chih-Yang Huang3,5,9,*
1Department of Neurological Surgery,Tri-Service General Hospital, National Defense
Medical Center, Taipei, Taiwan, 2Department of Biological Science and Technology,
China Medical University, Taichung, Taiwan, 3School of Chinese Medicine, China
Medical University, Taichung, Taiwan, 4Chinese Medicine Department, China
Medical University Beigang Hospital, Taiwan, 5Graduate Institute of Basic Medical
Science, China Medical University, Taichung, Taiwan, 6Laboratory of Exercise
Biochemistry, TPEC, Taipei, Taiwan,7Orthopaedic Department, Armed Forces
General Hospital, Taichung, Taiwan, 81PT Biotechnology Co., Ltd. , Taichung,
Taiwan,9Department of Health and Nutrition Biotechnology, Asia University,
Taichung, Taiwan
Running Title: PCA on Schwann cell migration *These authors contributed equally to this work.
Author for correspondence: Chih-Yang Huang, Graduate Institute of Basic Medical Science, China Medical University, No 91, Hsueh-Shih Road, Taichung, 404, Taiwan. Phone number: +886-4-2205-3366 ext. 3313; Fax number: +886-4-2207-0465; E-mail: [email protected]
Abstract
Alpiniaoxyphylla MIQ(AlpinateOxyphyllaeFructus, AOF) is an important traditional
Chinese medicinal herb whose fruits are widely used to prepare tonicsand are used as an aphrodisiac, anti-salivary, anti-diuretic and nerve-protective agent.
Protocatechuic acid (PCA), a simple phenolic compound was isolated from the kernels ofAOF.This study investigatedthe role of PCA in promoting neural regeneration and the underlying molecular mechanisms. Nerve regeneration is a
complex physiological response that takes place after injury. Schwann cells play a crucial role in the endogenous repair of peripheral nerves due to their ability to proliferate and migrate. The role of PCA in Schwann cell migration was determined by assessing the induced migration potential of RSC96 Schwann cells. PCA induced changes in the expression of proteins of three MAPK pathways,as determined using Western blot analysis. In order to determine the roles of MAPK (ERK1/2, JNK and p38) pathways in PCA-induced matrix-degrading proteolytic enzyme (PAs and
MMP2/9) production, the expression of several MAPK-associated proteins was analyzed after siRNA-mediated inhibition assays. Treatment with PCA induced ERK1/2, JNK and p38 phosphorylation that activated the downstream expression of
PAs and MMPs. PCA-stimulated ERK1/2, JNK and p38 phosphorylation was attenuated by individual pretreatment with siRNAs or MAPK inhibitors (U0126, 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
SP600125 and SB203580), resulting in the inhibition of migration and the uPA-related signal pathway.Taken together, our data suggest thatPCA extractregulate the
MAPK (ERK1/2, JNK and p38)/PA (uPA, tPA)/MMP (MMP2, MMP9) mediated regeneration and migration signaling pathwaysin Schwann cells. Therefore, PCA plays a major role in Schwann cell migration and the regeneration of damaged
peripheral nerve.
Key Words:AlpiniaoxyphyllaMIQ. fruit extract; Schwann cell migration; nerve regeneration; protocatechuic acid.
20 21 22 23 24 25 26 27 28
Introduction
Nerve regeneration is a complex phenomenon that has drawn the attention of scientists for many years. Neurons can be separated into central and peripheral
nervous systems (PNS);neurons in these two distinct systemshave different
anatomical structures and regenerative ability. In mammals, the central neurons are difficult to regenerate, whereas the neurons of the PNS have a myelin sheath and can
also regrow more easily. Schwann cells are the support cells of the PNS and can differentiate into the PNS myelin sheath and proliferate and migrate to the distal end
of the injured nerve area . Schwann cell migration is critical for axonal elongation and re-myelination of injured nerves .Peripheral nerve injury usually activates
proximal Schwann cells and macrophages to synthesize a cocktail of neurotrophic factors, adhesion molecules, cytokines and growth-promoting surface molecules .
However, the mechanisms by which these factors regulate Schwann cell migration, proliferation and signal transduction remain unclear. The ability of the Schwann cells to promote regeneration in peripheral neurons has led to an increased interest in
using Schwann cells for peripheral nervous system repair.
The mitogen-activated protein kinase (MAPK) proteins play essential roles in inducing cell migration. JNK, p38 and ERK1/2, which are members of the MAPK family, play crucial roles in the migration of nerve cells . Many experiments have 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48
shown that after injury, rapid increasesin tissue plasminogen activator (tPA) and urokinase plasminogen activator (uPA) expression have been observed in neurons . uPA and tPAare known to cleave plasminogen into plasmin and promote the
degradation of the extracellular matrix (ECM)facilitating cell migration during recovery from injury.
AlpinateOxyphyllaeFructus (AlpiniaoxyphyllaMIQ, AOF) is one of the most important traditional Chinese medicines, and according to the Chinese
pharmacopoeia,it has been used to treat diarrhea, polyuria, ulceration, dementia, tumor proliferationand gastralgia. Several reports have also shown that AOF extracts
potentially exhibitneuro-protectiveeffects. In our search for new active compounds from traditional Chinese medicinal herbs, protocatechuic acid (PCA), a simple phenolic compound, was isolated from the kernels ofAOF. Recently, PCA has been repoted to show strong antioxidant and antitumor effects and further exhibited to induce apoptosis in HL-60 human leukemia cells . The aim of the present study was
to investigate the effects of PCAon Schwann cell migration. 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63
Materials and Methods
Materials and Extraction of Protocatechuic Acid
All the chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwisespecified. The p38 inhibitor, MEK1 and MEK2 inhibitor and JNK
inhibitor were purchased from Promega (Madison, WI, USA). Protocatechuic acid (PCA) was isolated from AOF as previously reported. The purity of the compound was greater than 98% as determined by high-performance liquid chromatography
(HPLC) analysis.
Cell culture and treatments
RSC96 Schwann cells were purchased from the American Type Culture Collection (ATCC) and cultured in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal bovine serum (FBS), 4 mM L-glutamate, 1.5 g/L sodium bicarbonate and 4.5 g/L glucose in humidified atmosphere of 5% CO2 and
95% air. After 4 h of culture in serum-free media, the cells were treated with
different concentrations of PCA for 16~24 h and then harvested for further analysis. Cell viability assay
Cell viability was estimated using a colorimetric assay based on conversion of atetrazolium dye (MTT) into a blue formazan product.Briefly, RSC96Schwann cells
were plated at a density of 2×104 cells/well in 12-well plates. PCA was added to the
wells at different concentrations (0.1-2.0 mM) and the cells were cultured for 24 h. 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84
The culture medium was then replaced with 500 μL of MTT solution. After 4 h of incubation at 37 °C, the MTT solution was removed, and the produced formazan was solubilized in 200 μLof dimethyl sulfoxide (DMSO). The absorbance was measured at 570 nm using an automated microplate reader.
Migration assay
We used a Boyden chamber and polyvinyl-pyrrolidone-free polycarbonate
membranes with 8 μm pores (Neuro Probes, Inc.,Gaithersburg, MD, USA) for the migration assay. The bottom wells of the chambers were filled with DMEMmedium
supplemented with 10% FBS. The wells were covered with a serum-free membrane sheet added onto the top chamber. The membranes were stained with Giemsa stain (Sigma). The cells that migrated through the membrane were counted using a counting grid fitted into the eyepiece of a phase contrast microscope.
Western blotting
Cultured RSC96 cells were collected by scraping and washed, and total cellular protein was extracted as previouslydescribed.The proteins were then separated using 12% SDS-PAGE and transferred to nitrocellulose membranes. Non-specific protein binding was inhibited by treating the membranes in blocking buffer (5% milk,
20 mMTris-HCl, pH 7.6, 150 mMNaCl and 0.1% Tween 21) and then the membranes were blotted with specific antibodies in blocking buffer at 4 °C
overnight. When necessary, the nitrocellulose membranes were stripped with 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104
Restore Western blot stripping buffer for repeated blotting (Pierce Biotechnology, Inc, Rockford, IL, USA) at 37 °C for 15 min.
Zymography
The activities of matrix-degrading proteolytic enzyme (MMP)-2 and MMP-9 were determined using gelatin zymography. RSC96 cells were treated with different PCA concentrations. After 24 h of culture, the medium was collected and electrophoresed on an 8% polyacrylamide gel containing 0.1% gelatin. The gel was washed twice for
30 min in washing buffer (2.5% Triton X-100).The gel was then incubated in incubation buffer (1% NaN3; 2 M Tris-HCl, pH 8.0 and1M CaCl2) at 37 °C for 24 h
with shaking and subsequently stained with Coomassie blue.The presence of MMP-2 and MMP-9 gelatinolytic activity was identifiedas clear bands on a blue
background after destaining. siRNA application
Double-stranded siRNA sequences targeting MEK, p38 and JNK mRNAs were obtained from Dharmacon (Lafayette, CO, USA). A non-specific duplex was used as
a control. RSC96 cells were cultured in DMEM without FBS and transfected with double-stranded siRNA using the DharmaFECT Duo Transfection Reagent
(Dharmacon) according to the manufacturer's instructions. To assess gene silencing the ERK1/2, p38 and JNK protein levels were detected by Western blot.
Statistical analysis 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124
All experiments were performed in triplicates. Significantl differences were assessed using one way-ANOVA. P<0.05 was considered statistically significant. The data are expressed as the mean± SEM.
125 126 127
Results
Effects of PCA on RSC96 Schwann cell viability
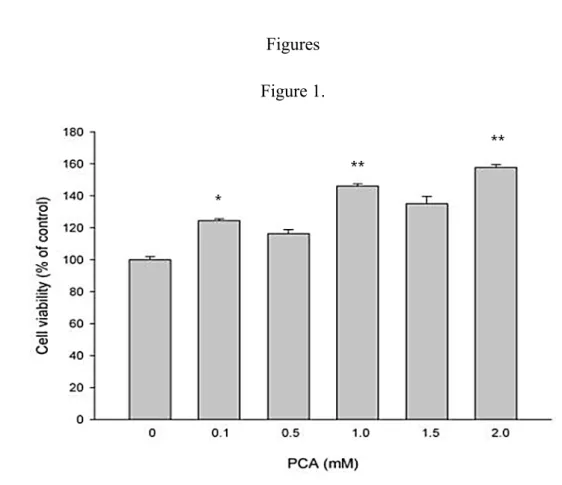
The effect of PCA on the proliferation of RSC96 Schwann cells wasevaluated to assess the ability of PCA to induce the regenerative ability by PCA. Theviability of
the cells significantly increased at the 0.1, 1.0 and 2.0 mM concentrations when measured after 24 h of treatment (Fig. 1). Therefore, these results suggest that treatment with 0.1-2.0 mMPCA for 24 h induces cell proliferation.
PCApromotes the migration of RSC96 Schwann cells
An in vitro experiment was performed using a Boyden chamber system to evaluate the PCA- induced migration potential of RSC96 Schwann cells. The migration
RSC96 cells increased significantly in a dose-dependent manner after PCA treatment (Fig. 2). These results provide evidence that PCA enhances the
proliferation and migrationof RSC96 Schwann cells.
Role of MAPKs in PCA induced RSC96 Schwann Cell Migration
The mechanism of PCA-induced migration in RSC96 Schwann cells wasanalyzed by evaluating the levels of intracellular MAPK signaling components, MEK1/2, ERK1/2, JNK1/2, p38 and their phosphorylatedforms, byWestern blotting. The
phosphorylation of MEK1/2, ERK1/2, JNK1/2 and p38 increased after PCA
treatment(Fig. 3A and B). Therefore, PCAmayinduce the activation of the MEK1/2, 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147
ERK1/2, JNK1/2 and p38 signaling pathways to promote migration in RSC96 Schwann cells. Moreover, the levels of uPAproteins were also increased, and the PAI-1 protein levels decreased(Fig. 3C). PCA also induced the expression of
MMP-9 and MMP-2, and decreased the levels of TIMP-1 and TIMP-2 levels (Fig. 3C). The zymographyresults further demonstrated that the activity of MMP9 and MMP2, increased after PCA treatment (Fig. 3D). These results indicate that PCAmediatesthe activation of the ERK1/2, JNK1/2 and p38 pathways to induce PAs and MMP2/9, resulting in RSC96 Schwann cell migration.
RSC96 Schwann cell migration enhanced by PCA is dependent on ERK1/2, JNK and p38signaling
To determine whether PCA-induced cell migration is indeed mediated through ERK1/2, JNK and p38 MAPK,specific inhibitors of these MAPK cascades were used. Schwann cells were pretreated with U0126, SP600125 and SB203580,
pharmacological inhibitors of MEK1/2, JNK, and p38,and then incubated with 1.0 mMPCA for 24 h. The activation of the downstream signaling substrates of
ERK1/2, JNK and p38 by PCAextract was allsignificantly reduced by pre-treatment with the MAPK inhibitors (Fig. 4). These results wereconfirmed further byknocking down ERK, JNK and p38 protein expression using siRNAs(Fig. 5). After the PCA-treated Schwann cells were transfected with siRNA targeting MEK, JNK or p38, 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166
there was a significant reduction in the levels of p-ERK, p-JNK and p-p38 protein and the correspondingdownstream substrates.Therefore, our results demonstrate that
PCA-induced Schwann cell migration occurs by the activation of the PAs and MMPs and that this activationis dependent on the ERK1/2, JNK and p38 MAPK
pathways. 167 168 169 170 171
Discussion
Schwann cells produce and release adhesion molecules and trophic factors that are essential for successful nerve regeneration following injury . Chinese herbal
medicines have attracted a great deal of attention as alternative and complementary medicines . AOF, which is one of the most important traditional Chinese medicines, has been reported to be a potential neuro-protective agent . Schwann cells in the area
of the injured nerve migrate and form a band ofBungner, supporting axonal regrowth . In our search for new active ingredients from traditional Chinese medicinal herbs that promote nerve regeneration, PCA, a simple phenolic
compound, was isolated from the kernels ofAOF. Recently, PCA has demonstrated a strong antioxidant and antitumor promotion effects and it was shown that PCA induced apoptosis in HL-60 human leukemia cells . This study investigated the
positive effects of PCA on Schwann cell migration.
The migration of RSC96 Schwann cells in the direction of growth is an important phenomenon that is essential for nerve regenerationand helps in repair damaged peripheral nerves. A specific migration pathway that is mediated by PCA in RSC96 Schwann cells, was observed to induce the activation of uPA and tPA through the
ERK, JNK and p38 signaling pathways (Fig.6).PCA treatment of RSC96 Schwann cells resulted in the phosphorylation of ERK1/2, JNK and p38 (Fig. 3A and 3B) with 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191
a corresponding expression ofuPA and tPA in a dose-dependent manner (Fig. 3C), andincreased theactivities ofMMP9 and MMP2 (Fig. 3C and 3D). Furthermore, using chemical inhibitors (Fig. 4) and SiRNAs (Fig. 5), the migratory effects
induced by PCA on RSC96 cells were further identified to be dependent on ERK1/2, JNK and p38 signaling. In addition, Boyden chamber systemexperiments showed
that the increase in the activities of extracellular MMP9 and MMP2proteins was associated with increase in the migration of Schwann cell after PCA treatment(Fig.
2).These results provide evidence that PCA promotes the migration of Schwann cells.
To promote migration, cells secrete proteases, including tPA and uPA,that degrade matrix molecules and cell adhesion factors . The overexpression oftPA regenerates
neurites to a greater extent and facilitates faster migration of cells with complex ECM. PAI-1 is thought to be the major inhibitor of migration. Our data clearly show
that the phosphorylation of ERK1/2, JNK and p38 accompanies the increased expression of uPA and tPA. Conversely, the expression of PAI-1 is gradually decreased (Fig. 3C).
The development and regeneration of the PNS is highly dependent on the migration of Schwann cells and the extension of axons toward their distant targets. PAs are also associated with several neural cell types and they are believed to mediate the 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210
localized degradation of the ECM, thus facilitating cell motility . ECM degradation is associated with neural tissue growth. One of the key regulators of this process is the serine protease uPA, which acts on a wide variety of ECM components . Cell
proliferation and angiogenesis processes are events involving uPA mediateddegradation of ECM. We hypothesize that ERK1/2, JNKand p38 phosphorylation promotesthe expression of uPA and tPA in PCA-treated cells. Another family of proteases, the MMPs, are also implicated in peripheral nerve
regeneration and are involved in cellmigration . MMPs are secreted by many cell types, including neuronsas inactive molecules and require activation via other proteases. Plasmin which are activated by tPA or uPA, activate MMP-9 and MMP-2 . We observed elevated expression and activity of MMP2 and MMP9 (Fig.
3D).However, specific activation of ERK, JNK and p38 kinases either by
overexpression or by using activtors should be further performed to confirm the role of these MAP kinases in Schwann cell migration. In addition, effects of
anymodulations inERK, JNK and p38 kinase activities on 2 and MMP-9activity as well as levels and cell migration have to be determined to conclude the involvement of ERK, JNK and p38 kinase activation in Shwann cell migration.
Based on the availableresults, we suggest that PCA or some of its components may have nerve cell migration and/or regeneration promotion potential. Our results 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229
demonstrate that PCA can stimulate Schwann cell migration and up regulate the expression of PA and MMP2/9 via the ERK1/2, JNK and p38 MAPK pathways. These findings identify potential inducer of neuron regeneration. PCA may serve as a promising migration-inducing and/or therapeutic drug for nerve regeneration. 230
231 232 233
References
An, L.J., Guan S.Guan, Shi G.F.Shi, Bao Y.M.Bao, Duan Y.L. Duan and Jiang B. Jiang. Protocatechuic acid from Alpinia oxyphylla against MPP+-induced
neurotoxicity in PC12 cells. Food Chem. Toxicol.44: 436-443, 2006.
Andreasen, P.A., Kjoller L.Kjoller, Christensen L. Christensenand Duffy M.J. Duffy.The urokinase-type plasminogen activator system in cancer metastasis: A review. International Int.Journal J.of Cancer 72: 1-22, 1997.
Anton, E.S., Sandrock A.W. and Matthew W.D. Merosin Promotes Neurite Growth and Schwann-Cell Migration in-Vitro and Nerve Regeneration in-Vivo - Evidence Using an Antibody to Merosin, Arm-1. Developmental Biology 164: 133-146, 1994.
Ass, K., Ahmad A., Azmi A.S., Sarkar S.H. and Sarkar F.H. Evolving role of uPA/uPAR system in human cancers. Cancer Treatment Reviews 34: 122-136, 2008. Bunge, R.P. Expanding roles for the Schwann cell: ensheathment, myelination, trophism and regeneration. Curr Opin Neurobiol 3: 805-809, 1993.
Chang, Y.-M., Velmurugan B.K., Kuo W.-W., Chen Y.-S., Ho T.-J., Tsai C.-T., Ye C.-X., Tsai C.-H., Tsai F.-J. and Huang C.-Y. Inhibitory effect of alpinate
Oxyphyllae fructus extracts on Ang II-induced cardiac pathological remodeling-related pathways in H9c2 cardiomyoblast cells. BioMedicine 3: 148-152, 2013. Fawcett, J.W. and Keynes R.J. Peripheral nerve regeneration. Annu Rev Neurosci 13: 43-60, 1990.
Feneley, M.R., Fawcett J.W. and Keynes R.J. The Role of Schwann-Cells in the Regeneration of Peripheral-Nerve Axons through Muscle Basal Lamina Grafts.
Experimental Neurology 114: 275-285, 1991.
Hayden, S.M. and Seeds N.W. Modulated expression of plasminogen activator system components in cultured cells from dissociated mouse dorsal root ganglia.
Journal of Neuroscience 16: 2307-2317, 1996.
Hildenbrand, R., Gandhari M., Stroebel P., Marx A., Allgayer H. and Arens N. The urokinase-system - role of cell proliferation and apoptosis. Histology and
Histopathology 23: 227-236, 2008.
Huang, C., Jacobson K. and Schaller M.D. MAP kinases and cell migration. J Cell
Sci 117: 4619-4628, 2004.
Ide, C. Peripheral nerve regeneration. Neurosci Res 25: 101-121, 1996.
Jessen, K.R. and Mirsky R. Schwann cells and their precursors emerge as major regulators of nerve development. Trends in Neurosciences 22: 402-410, 1999. Koo, B.S., Lee W.C., Chang Y.C. and Kim C.H. Protective effects of alpinae oxyphyllae fructus (Alpinia oxyphylla MIQ) water-extracts on neurons from
ischemic damage and neuronal cell toxicity. Phytother Res 18: 142-148, 2004. 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271
Meintanis, S., Thomaidou D., Jessen K.R., Mirsky R. and Matsas R. The neuron-glia signal beta-neuregulin promotes Schwann cell motility via the MAPK pathway. Glia
34: 39-51, 2001.
Michalopoulos, G.K. and DeFrances M.C. Liver regeneration. Science 276: 60-66, 1997.
Muir, D. Metalloproteinase-Dependent Neurite Outgrowth within a Synthetic Extracellular-Matrix Is Induced by Nerve Growth-Factor. Experimental Cell
Research 210: 243-252, 1994.
Muir, E.M., Adcock K.H., Morgenstern D.A., Clayton R., von Stillfried N., Rhodes K., Ellis C., Fawcett J.W. and Rogers J.H. Matrix metalloproteases and their
inhibitors are produced by overlapping populations of activated astrocytes.
Molecular Brain Research 100: 103-117, 2002.
Murphy, G., Atkinson S., Ward R., Gavrilovic J. and Reynolds J.J. The Role of Plasminogen Activators in the Regulation of Connective-Tissue Metalloproteinases.
Annals of the New York Academy of Sciences 667: 1-12, 1992.
Nagase, H. Activation mechanisms of matrix metalloproteinases. Biological
Chemistry 378: 151-160, 1997.
Pittman, R.N. and Dibenedetto A.J. Pc12 Cells Overexpressing Tissue-Plasminogen Activator Regenerate Neurites to a Greater Extent and Migrate Faster Than
Control-Cells in Complex Extracellular-Matrix. Journal of Neurochemistry 64: 566-575, 1995.
Shi, G.F., An L.J., Jiang B., Guan S. and Bao Y.M. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo.
Neurosci Lett 403: 206-210, 2006.
Shoji, N., Umeyama A., Takemoto T. and Ohizumi Y. Isolation of a Cardiotonic Principle from Alpinia-Oxyphylla. Planta Medica 50: 186-187, 1984.
Shui, G., Bao Y.M., Bo J. and An L.J. Protective effect of protocatechuic acid from Alpinia oxyphylla on hydrogen peroxide-induced oxidative PC12 cell death. Eur J
Pharmacol 538: 73-79, 2006.
Siconolfi, L.B. and Seeds N.W. Induction of the plasminogen activator system accompanies peripheral nerve regeneration after sciatic nerve crush. J Neurosci 21:
4336-4347, 2001.
Siconolfi, L.B. and Seeds N.W. Mice lacking tPA, uPA, or plasminogen genes showed delayed functional recovery after sciatic nerve crush. Journal of
Neuroscience 21: 4348-4355, 2001.
Snider, W.D., Zhou F.Q., Zhong J. and Markus A. Signaling the pathway to regeneration. Neuron 35: 13-16, 2002.
Torigoe, K., Tanaka H.F., Takahashi A., Awaya A. and Hashimoto K. Basic 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309
behavior of migratory Schwann cells in peripheral nerve regeneration. Exp Neurol 137: 301-308, 1996.
Tseng, T.H., Hsu J.D., Lo M.H., Chu C.Y., Chou F.P., Huang C.L. and Wang C.J. Inhibitory effect of Hibiscus protocatechuic acid on tumor promotion in mouse skin.
Cancer Letters 126: 199-207, 1998.
Tseng, T.H., Kao T.W., Chu C.Y., Chou F.P., Lin W.L. and Wang C.J. Induction of apoptosis by Hibiscus protocatechuic acid in human leukemia cells via reduction of
retinoblastoma (RB) phosphorylation and Bcl-2 expression. Biochemical
Pharmacology 60: 307-315, 2000.
Tseng, T.H., Wang C.J., Kao E.S. and Chu H.Y. Hibiscus protocatechuic acid protects against oxidative damage induced by tert-butylhydroperoxide in rat primary hepatocytes. Chemico-Biological Interactions 101: 137-148, 1996.
Yu, X., An L., Wang Y., Zhao H. and Gao C. Neuroprotective effect of Alpinia oxyphylla Miq. fruits against glutamate-induced apoptosis in cortical neurons.
Toxicol Lett 144: 205-212, 2003.
Zhang, H.Y. and Tang X.C. Neuroprotective effects of huperzine A: new therapeutic targets for neurodegenerative disease. Trends in Pharmacological Sciences 27:
619-625, 2006. 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329
Figure legends
Figure 1.The effect of PCA extract on RSC96 cell viability. Schwann cells were treated with 0.1-2.0 mM PCA for 24 hours.Cell viability was measured using MTT assay and the data are shown as the mean of three independent experiments ± SE. * denotes significant differences from control values with p<0.05.
Figure 2.The effect of PCA on RSC96 cell migration. Schwann cells were treated with different doses of PCA and the cell migration was measured using Boyden chambers. The data are shown as the mean of three independent experiments ± SE. *
denotes significant differences from control values with p<0.05.
Figure 3. The effect of PCA on RSC96 cell viability is mediated by MAPK signaling. The RSC96 cells were treated with the indicated doses of PCA. The abundance and phosphorylation of MAPK proteins were measured by Western blotting (A-C). α-Tubulin was used as a loading control. The activity of MMP-9 and
MMP-2 was measuredusing gelatin zymography (D).
Figure 4.The effects of PCA on Schwann cell migration are dependent on ERK1/2, JNK and p38 signaling. RSC96 cells were pretreated with U0126 (A), SB203580 (B) or SP600125 (C) for 1 hour and then treated with 1.0 mM AOF PCA for 24
hours. The levels of the indicated proteins were measured using Western blot analysis. α-Tubulin was used as a load control.
330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349
Figure 5. ERK1/2, p38 and JNK signaling are for PCA-induced cell migration. RSC cells were transiently transfected with ERK1/2 siRNA (5 nM), p38 siRNA (5 nM) or JNK siRNA (5 nM) for 8 hours and then treated with 0.1 mM PCA for 24 hours (A-C). A non-specific duplex was used as a control.The levels of the indicated proteins were measured using Western blot analysis. α-Tubulin was used as a loading
control.
Figure 6.Model showing the effects of PCA on RSC96 Schwann cell migration. Stimulation of Schwann cells with PCA activates ERK1/2, JNK and p38 signaling,
which leads to the up-regulation of uPA and tPA contributes to the activation of MMP9 and MMP2, and enhances the migration of RSC96Schwann cell.
350 351 352 353 354 355 356 357 358 359
Figures Figure 1.
Figure 1.The effect of PCA extract on RSC96 cell viability. Schwann cells were treated with 0.1-2.0 mM PCA for 24 hours.Cell viability was measured using MTT assay and the data are shown as the mean of three independent experiments ± SE. * 360 361 362 363 364 365 366
and ** denotes significant differences from control values with p<0.05 and p<0.01, respectively.
367 368
Figure 2. 369
370
Figure 2.The effect of PCA on RSC96 cell migration. Schwann cells were treated with different doses of PCA and the cell migration was measured using Boyden chambers. The data are shown as the mean of three independent experiments ± SE. *, ** and *** denotes significant differences from control values with p<0.05,
p<0.01 and p<0.001, respectively. 372 373 374 375 376
Figure 3.
Figure 3. The effect of PCA on RSC96 cell viability is mediated by MAPK signaling. The RSC96 cells were treated with the indicated doses of PCA. The abundance and phosphorylation of MAPK proteins were measured by Western blotting (A-C). α-Tubulin was used as a loading control. The activity of MMP-9 and
MMP-2 was measuredusing gelatin zymography (D). 377 378 379 380 381 382 383 384
Figure 4. 385
386
Figure 4.The effects of PCA on Schwann cell migration are dependent on ERK1/2, JNK and p38 signaling. RSC96 cells were pretreated with U0126 (A), SB203580 (B) or SP600125 (C) for 1 hour and then treated with 1.0 mM AOF PCA for 24
hours. The levels of the indicated proteins were measured using Western blot analysis. α-Tubulin was used as a load control.
388 389 390 391 392
Figure 5. 393
394
Figure 5. ERK1/2, p38 and JNK signaling are for PCA-induced cell migration. RSC cells were transiently transfected with ERK1/2 siRNA (5 nM), p38 siRNA (5 nM) or JNK siRNA (5 nM) for 8 hours and then treated with 0.1 mM PCA for 24 hours (A-C). A non-specific duplex was used as a control. The levels of the indicated proteins were measured using Western blot analysis. α-tubulin was used as a loading control. 396
397 398 399 400
Figure 6.
Figure 6.Model showing the effects of PCA on RSC96 Schwann cell migration. 401
402
403
Stimulation of Schwann cells with PCA activates ERK1/2, JNK and p38 signaling, which leads to the up-regulation of uPA and tPA contributes to the activation of MMP9 and MMP2, and enhances the migration of RSC96Schwann cell.
405 406 407