Abstract.
Phenethyl isothiocyanate (PEITC), one of many
compounds found in cruciferous vegetables, has been
reported as a potential anticancer agent. In earlier studies,
PEITC was shown to inhibit cell growth and induction of
apoptosis in many cancer cell lines. However, no report has
shown whether PEITC can induce apoptosis in human
prostate cancer cells. Herein, we aimed to determine whether
PEITC has anticancer activity in DU 145 human prostate
cancer cells. As a result, we found that PEITC induced a
dose-dependent decrease in cell viability through induction
of cell apoptosis and cell cycle arrest in the G
2/M phase of
DU 145 cells. PEITC induced morphological changes and
DNA damage in DU 145 cells. The induction of G
2/M phase
arrest was mediated by the increase of p53 and WEE1 and it
reduced the level of CDC25C protein. The induction of
apoptosis was mediated by the activation of caspase-8-,
caspase-9- and caspase-3-depedent pathways. Results also
showed that PEITC caused mitochondrial dysfunction,
increasing the release of cytochrome c and Endo G from
mitochondria, and led cell apoptosis through a
mitochondria-dependent signaling pathway. This study showed that PEITC
might exhibit anticancer activity and become a potent agent
for human prostate cancer cells in the future.
Prostate cancer is not a single disease and is an umbrella
which under a plethora of heterogeneous diseases is
included. These heterogeneous diseases include indolent
localized tumors and aggressive metastatic diseases (1). In
the developed world, prostate cancer is the primary male
cancer, accounting for 24% of all diagnosed male cancer
cases, and is the second leading cause of death from cancer
(2). In Taiwan, prostate cancer is the seventh most common
cancer in males and about 8.0 men per 100 thousand die
annually from prostate cancer based on reports from the
Department of Health, Executive Yuan, R.O.C (Taiwan) in
*These authors contributed equally to this study.Correspondence to: Jing-Gung Chung, Department of Biological
Science and Technology, China Medical University, No. 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C. Tel: +886 422053366 ext. 2161, Fax: +886 422053764, e-mail: [email protected]
Key Words: Phenethyl isothiocyanate, cell cycle arrest, apoptosis,
mitochondria, caspases, prostate cancer DU 145 cells.
ANTICANCER RESEARCH 31: 1691-1702 (2011)
Phenethyl Isothiocyanate (PEITC) Promotes G
2
/M
Phase Arrest
via p53 Expression and Induces Apoptosis
through Caspase- and Mitochondria-dependent Signaling
Pathways in Human Prostate Cancer DU 145 Cells
NOU-YING TANG
1*, YA-TING HUANG
2*, CHUN-SHU YU
3, YANG-CHING KO
4,
SHIN-HWAR WU
5,6, BIN-CHUAN JI
7, JAI-SING YANG
8, JIUN-LONG YANG
9,
TE-CHUN HSIA
1,10, YA-YIN CHEN
11,12and JING-GUNG CHUNG
13,14*Schools of
1Chinese Medicine and
3Pharmacy, Departments of
2Medical Laboratory Science and Biotechnology,
8
Pharmacology and
13Biological Science and Technology, Graduate Institutes of
6Clinical Medical Science,
9Chinese Pharmaceutical Science, China Medical University, Taichung 404, Taiwan, R.O.C.;
4
Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine,
St. Martin De Porres Hospital, Chiayi 600, Taiwan, R.O.C.;
5
Division of Critical Care Medicine, Department of Internal Medicine, and
7Division of Respiratory Care Center, Department of Medicine,
Changhua Christian Hospital, Changhua 500, Taiwan, R.O.C.;
10
Department of Internal Medicine, China Medical University Hospital, Taichung 404, Taiwan, R.O.C.;
11Department of Chinese-Western Medicine Integration,
Chung Shan Medical University Hospital, Taichung 402, Taiwan, R.O.C.;
12
School of Medicine, Chung Shan Medical University, Taichung 402, Taiwan, R.O.C.;
14Department of Biotechnology, Asia University, Taichung 413, Taiwan, R.O.C.
2009. About 90% of advanced prostate cancer patients
developing skeletal metastases and in advanced prostate
cancer patients is morbidity from bone metastasis is high (3),
despite treatments including radiation, chemotherapy, or
combination of radiotherapy with chemotherapy.
The majority of cancer-targeting drugs cause aberrant
cellular growth through the induction of apoptosis (4-6).
Apoptosis, programmed cell death type I, is regulated by
multifactor and signaling pathways to remove unwanted
cells (7, 8). Active caspases are known to be biologically
important and associated with apoptosis (9).
Poly-ADP-ribose polymerase (PARP) is cleaved by both caspase-3 and
caspase-7, and is an abundant DNA-binding enzyme that
detects and signals DNA strand breaks (10). Caspase-3 in
particular is the main executor of apoptosis and caspase
activation is considered to be a key hallmark of apoptosis
(11). Cytochrome c release from the internal part of the
mitochondrial membrane into the cytosol leads to the
activation of caspase-9, -3, -6, and -7, and then to apoptosis
(12-14).
Traditional drugs obtained from various plant types with
active natural compounds such as licochalcone-A (15),
xanthoangelol (16) and chalcones (17) have been reported to
act as potential agents for the treatment of cancer. Dietary
intake of cruciferous vegetables can reduce the risk of various
types of malignancies (18). Phenethyl isothiocyanate (PEITC)
is one of such important compounds from cruciferous
vegetables. Several mechanisms have been shown to be
involved in the antitumor effect of PEITC, including the
inhibition of cytochrome p450 enzymes, the induction of
phase II detoxification enzymes (19) and the reduction of
azoxymethane-induced colonic aberrant crypt foci formation
(20). PEITC also inhibited
4-(methylnitrosamino)-1-(3-pyridyl)-1-butone-induced pulmonary neoplasia in A/J mouse
lung (21). In addition, PEITC can act as a cancer
chemopreventive in rat (22). Recently, it was reported that
PEITC inhibits oxidative phosphorylation to trigger reactive
oxygen species-mediated death of human prostate cancer cells
(23).
Although many reports stated that PEITC induced cell
death through induction of apoptosis, there is no report to
address whether this occurs in human prostate cancer cells.
In this study, we investigated the effects of PEITC on cell
growth of human prostate cancer cells.
Materials and Methods
Reagents. PEITC, RPMI-1640 medium, fetal bovine serum, and
penicillin and streptomycin antibiotic mixture were purchased from Invitrogen Life Technologies (Grand Island, NY, USA). Propidium iodide (PI), dimethyl sulfoxide (DMSO), RNase A, and 4,6-diamidino-2-phenylindole (DAPI) were from Sigma-Aldrich Corp. (St. Louis, MO, USA). The antibodies against BAK, BID, BCL-2, cell division cycle 25C (CDC25C), and apoptosis inducing effect
(AIF) were from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA), and the antibodies against cytochrome c, Glucose-regulated protein 78 (GRP78), and PARP were from BD PharMingen (Palo Alto, CA, USA). 6-Carboxy-2,7-dichlorodihydrofluorescein diacetate (H2DCFDA), Fluo-3/AM and 3,3’-dihexyloxacarbocyanine iodide (DiOC6)were purchased from Invitrogen Life Technologies. The caspase-9 inhibitor (Z-LEHD-FMK), caspase-8 inhibitor (Z-IETD-FMK) and caspase-3 inhibitor (Z-DEVD-(Z-IETD-FMK) were obtained from R&D Systems Inc. (Minneapolis, MN, USA). All chemicals were reagent grade.
Cell culture. The human prostate carcinoma cell line (DU 145)
was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC). DU 145 cells were maintained in RPMI-1640 medium with 2 mM L-glutamine supplemented with 10% heat-inactivated fetal bovine serum, 100 μg/ml penicillin and 100 U⁄ml streptomycin. Cells (1×105cells/ml) were
maintained at 5% CO2and 37˚C in humidified 5% CO2and 95% air (24).
Cell morphology and cell viability assay. DU 145 cells were
maintained on 12-well plates and then incubation with different concentrations (0, 1, 5, 10, 15 and 20 μM) of PEITC for different time periods (24 and 48 h). For examination of cell morphological, treated and negative-control cells were observed at 24 h-incubation under a phase-contrast microscope and photomicrographs were taken. Cell viability was determined by 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The DU 145 (1×104cells/well/100 μl) cells were cultured in 96-well plates. After
incubation overnight, the RPMI-1640 medium in each well was replaced with different concentrated solutions of PEITC and cells further incubated for 24 and 48 h. At the end of incubation, 10 μl of MTT (5 mg/ml in phosphate-buffered saline (PBS)) (Sigma-Aldrich Corp.) were added individually to each well for 4 h incubation at 37˚C, and then removed media after 100 μl of DMSO were added. The absorbance was then measured by Microplate Reader (Bio-Rad, Hercules, CA, USA) at 570 nm (25, 26). In addition, cultured DU 145 cells with or without treatment with 25 μM of caspase-8 inhibitor (Z-IETD-FMK), caspase-9 inhibitor (Z-LEHD-FMK) or caspase-3 inhibitor (Z-DEVE-FMK), respectively, for 3 h prior to the addition 10 μM of PEITC were then analyzed for cell viability as described previously (26, 27).
Flow cytometric analysis for DNA content. DU 145 cells were
plated at a density of 2×105cells on 12-well plates for 24 h, and
exposed to 0, 1, 5, 10, 15 and 20 μM of PEITC for different time periods. At the end of incubation, cells were trypsinized and centrifuged then the cell pellet from each treatment was resuspended with 1 ml of PI staining buffer containing 4 mM sodium citrate, 0.1% Triton X-100, 50 μg/ml PI and 200 μg/ml RNAse and incubated for 30 min at 37˚C in the dark. All samples were analyzed for cell cycle distribution and sub-G1 proportion (apoptosis) by using a FACSCalibur flow cytometer and BD CellQuest Acquisition software (San Jose, CA, USA) as described previously (24, 27). Data in each phase were reported as the percent age cells in each phase of the cell cycle.
DAPI staining. Morphological changes of apoptosis were examined by
staining DU 145 cell nuclei with DAPI. Cells (5×104cells) were
of PEITC for 24 h. The cells were washed twice with PBS then were fixed with 4% paraformaldehyde-PBS solution for 15 min and stained with DAPI (300 nM) for 30 min at room temperature. The cells were washed with PBS and mounted. Images of DAPI fluorescence were examined and photographed under a fluorescence microscope as described previously (28, 29).
Comet assay. DU 145 cells (1×105cells) were treated with PEITC
(0, 1, 5, 10, 15 and 20 μM) for 24 h and then cells were embedded onto 0.6% low melting point agar then immersed in ice-cold cell lysis solution (2.5 mM NaCl, 100 mM ethylenediaminetetra acetic acid (EDTA), 10 mM Tris (pH 10), 1% N-laurylsarcosine, 1% Triton X-100 and 10% DMSO). Cells were then washed and digested with Tang et al: Phenethyl Isothiocyanate Triggers Apoptosis in Human Prostate Cancer Cells
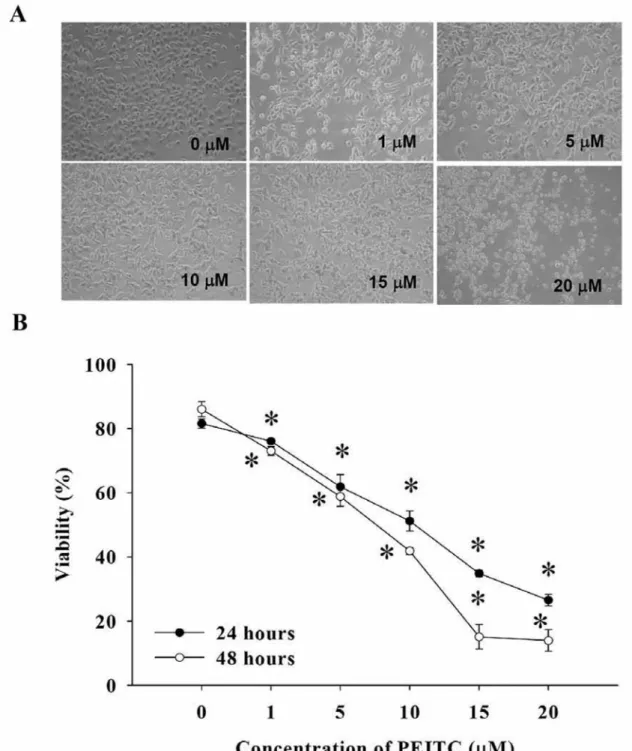
Figure 1. PEITC affected on cell morphology and viability in DU 145 cells. Cells were cultured in RPMI-1640 medium and 10% FBS with PEITC
for 24 and 48 h. Morphological changes were examined and cells photographed under a phase-contrast microscope (×200) (A). Cells viability was measured by flow cytometric assays as described in the Materials and Methods (B). *p<0.05 Significantly different from the control.
two units of endonuclease III in the same buffer for 1 h at 37˚C then slides were denatured with 0.3 mM NaOH, 1 mM EDTA (pH 13.4) for 20 min and electrophoresis was carried out at 25 V, 300 mA for 25 min. Cellular DNA was stained with 4 μg/ml PI and examined under a fluorescence microscope as described previously (26, 30).
Flow cytometric detection of reactive oxygen species (ROS), mitochondrial membrane potential (ΔΨm) and intracellular Ca2+
levels. DU 145 cells were plated at a density of 2×105cells on 12-well
plates for 24 h, and then PEITC (10 μM) was added to the well and cells incubated for different time periods. At the end of incubation, cells were trypsinized and centrifuged. The cells from each treatment were collected and washed twice with PBS, re-suspended in 500 μl of H2DCFDA (10 μM) for ROS, 500 μl of DiOC6(1 μM) for ΔΨmand 500 μl of Fluo-3/AM (2.5 μg/ml) for intracellular Ca2+ release. All
samples were incubated with these specific fluorochromes at 37˚C for
30 min and the differential expressions from each treatment were analyzed by flow cytometry (27, 31).
Caspase-3, -8 and -9 activity assays. DU 145 cells were plated at a
density of 2×105cells on 12-well plates for 24 h, and then PEITC (10
μM) was added to the well and cells incubated for different time periods. The cells from each treatment were trypsinized and centrifuged, washed twice with PBS and were re-suspended in 50 μl of 10 μM substrate solution (PhiPhiLux-G1D1 for caspase-3, CaspaLux8-L1D2for caspase-8 and CaspaLux9-M1D2for caspase-9, OncoImmunin, Inc. Gaithersburg, MD, USA) before being incubated at 37˚C for 60 min. The cells were washed again by PBS and were analyzed by flow cytometry as described previously (29, 30).
Western blotting analysis. Western blotting was carried out as
previously described (30). DU 145 cells were seeded at a density of 2×106cells/ml in RPMI-1640 medium with 10% fetal bovine serum
for 24 h. Cells were treated with or without 10 μM PEITC for 0, 6, 12, 24 and 48 h. Cells from each treatment were collected and protein was extracted into the PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea), and protein concentrations were determined by a BCA Protein Assay kit (Bio-Rad Laboratories, Hercules, CA, USA) (26). Proteins were resolved on a polyacrylamide gel via electrophoresis (SDS-PAGE). After SDS-PAGE transfer, the Polyvinylidene Fluoride (PVDF) membrane (Immobilon-P; Millipore, Bedford, MA, USA) was stained by primary antibody (anti-CDC25C, WEE1, p53, FAS, Fad ligand, BCL-2, BAK, BID, PARP, cytochrome c, AIF, GRP78 and GADD153), washed and then stained by appropriate horseradish peroxidase–conjugated secondary antibodies (GE Healthcare Pewaukee, WI, USA) for enhanced chemiluminescence (ECL) reagent (Millipore) as described previously (29, 32).
Immunofluorescence staining and confocal laser scanning microscopy. DU 145 cells (5×104 cells/well) were placed on 4-well
chamber slides before being treated with 10 μM PEITC for 24 h. Cells were then fixed in 3% formaldehyde in PBS for 15 min, permeabilized with 0.1% Triton X-100 in PBS for 1 h with blocking of non-specific binding sites using 2% bovine serum albumin (BSA) as described previously (29, 33). Fixed cells were stained with primary antibodies to GADD153 and endonuclease G (Endo G) (1:200 dilution) overnight and then stained with fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG secondary antibody at 1:100 dilution) (green fluorescence) followed by mitochondria and nuclei as counterstaining with PI (red fluorescence). Photomicrographs were obtained using a Leica TCS SP2 confocal spectral microscope.
Statistical analysis. The data are presented as the mean±S.D. of three
independent experiments. The statistical significance of the mean difference between the control and treated groups was determined by a paired t-test. P<0.05 was considered statistically significant.
Results
PEITC induced morphological changes and reduced the
percentage of viable DU 145 cells. To evaluate the effect of
PEITC on DU 145 prostate cancer cells, we determined and
measured the cell morphological changes and viability using a
phase-contrast microscope and the MTT assay, respectively. The
Figure 2. PEITC affected the cell cycle and associated protein levels inDU 145 cells. Cells were placed in RPMI-1640 and 10% FBS with PEITC for 48 h then were harvested for analysis of cell cycle distribution (A) by flow cytometric assay. Cells were harvested for examination of the G2/M phase arrest-associated protein levels and were assayed by Western blotting (B) as described in the Materials and Methods.
cells were dose-dependently treated with PEITC and
morphological changes of DU 145 cells after treatment with
PEITC are shown in Figure 1A. Compared with the control
(untreated) cells, the majority of the PEITC-treated DU 145 cells
became irregularly shaped spinouts and less cell number (Figure
1A). These morphological changes demonstrated cell damage
after PEITC treatment. As shown in Figure 1B, PEITC
significantly reduced the viability of DU 145 cells in a time- and
dose-dependent manner. A sharp decrease in cell viability was
present at 5 μM of PEITC treatment time. The cytotoxic effect
was more evident at 48 h. The viability of DU 145 cells after
exposure to 10 μM of PEITC for 48 h decreased by almost 50%.
PEITC induced cell cycle arrest and apoptosis in DU 145
cells. The effects of PEITC on the cell cycle progression in
DU 145 cells were determined by flow cytometry and PI
staining. In DU 145 cells, treatment with PEITC for 48 h
resulted in a dose-dependent increase in the percentage of
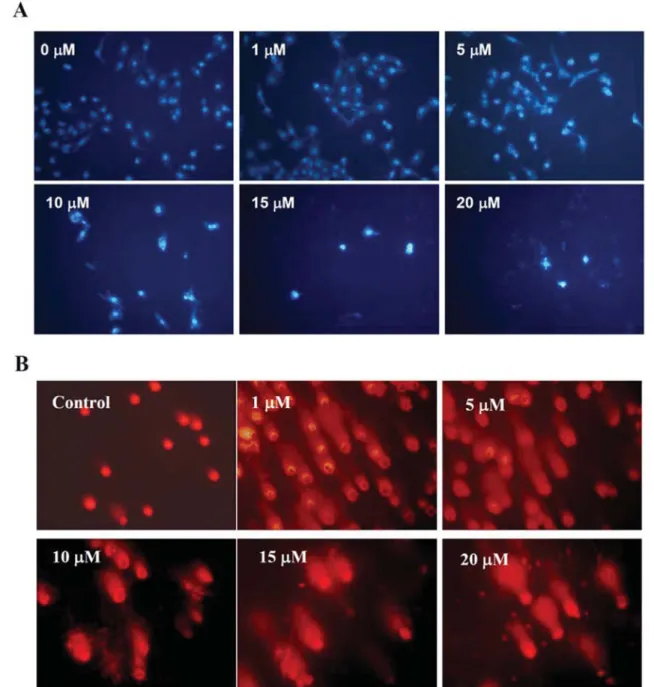
Tang et al: Phenethyl Isothiocyanate Triggers Apoptosis in Human Prostate Cancer CellsFigure 3. PEITC induced apoptosis and DNA damage in DU 145 cells. Cells were treated with PEITC at various doses and then were incubated for
24 h. Cells were harvested for DAPI staining then were examined and photographed under a fluorescence microscope for examination the apoptosis (A). Cells were stained with PI to examine DNA damage by Comet assay (B) as described in the Materials and methods.
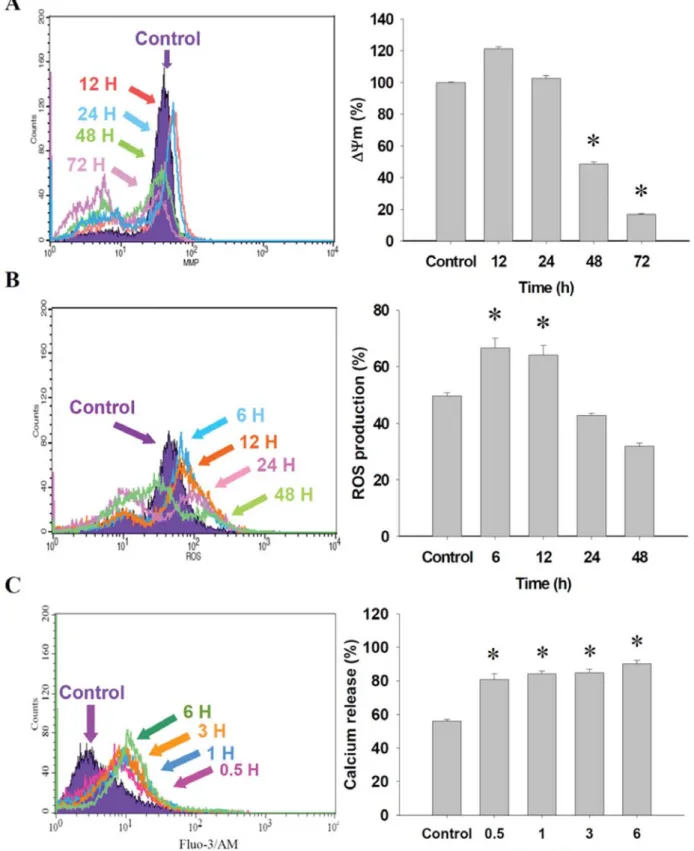
Figure 4. PEITC affected the levels of mitochondria membrane potential (ΔΨm), production of reactive oxygen species (ROS) and Ca2+and in DU
145 cells. Cells were treated with 10 μM of PEITC for various time periods then were collected and stained by DiOC6 for the ΔΨmlevels (A), with
2,7-dichlorodihydrofluorescein diacetate for ROS production (B) and by Fluo-3 for Ca2+level (C) as described in the Materials and Methods. Data
Tang et al: Phenethyl Isothiocyanate Triggers Apoptosis in Human Prostate Cancer Cells
Figure 5. PEITC induced caspase-3, -8 and -9 activities in DU 145
cells. Cells were treated with 10 μM of PEITC for 12, 24, 48 or 72 h and cells were collected for determination activities of caspase-3 (A), caspase-8 (B) and caspase-9 (C) as described in the Materials and Methods. Data represents the mean±S.D. of three experiments. *p<0.05, significantly different from the control.
Figure 6. Caspase inhibitors protected against PEITC-induced
cytotoxicy in DU 145 cells. Cells were pretreated with the caspase-9 LEHD-FMK) (A), caspase-8 IETD-FMK) (B) and caspase-3 (Z-DEVE-FMK) (C) inhibitors, and then were treated with 10 μM PEITC for 24 h before determination of viable cells as described in the Materials and Methods. Data represent the mean±S.D. of three experiments. *p<0.05, significantly different from the control.
cells in the G
2/M phase and a concomitant reduction of cell
numbers in the S phase (Figure 2A). Higher concentrations
of PEITC (15 and 20 μM) retuned the proportion of G
2/M
cells back to control levels. However, the proportion of G
0/G
1phase cells increased at treatment of PEITC at 15 and 20 μM.
The sub-G
1phase was clearly to increase when the treatment
of PEITC was increased and in a dose-dependent manner
(Figure 2A). This was further verified by microscopic
examination that showed a decrease of viable cell number in
treated samples. Results from Western blotting shown in
Figure 2B also show that PEITC increased the protein level of
p53, but it inhibited the protein levels of CDC25C and WEE1.
Taken together, these data indicated that PEITC induced
apoptosis and arrested cell cycle progression at the G
2/M
stage in DU 145 cells.
PEITC induced apoptosis and DNA damage in DU 145 cells.
We found that PEITC induced apoptosis in a dose-dependent
manner in DU 145 cells (Figure 3A). PEITC-induced DNA
damage was examined by Comet assay in DU 145 cells
(Figure 3B). Using this assay, we found that PEITC-induced
DNA damage in a dose-dependent manner in DU 145 cells.
PEITC reduced the level of ΔΨ
mand increased the production
of ROS and Ca
2+in DU 145 cells. To investigate whether
PEITC induced cytotoxic effects involved in ΔΨ
m, ROS and
Ca
2+in DU 145 cells, the levels of ΔΨ
mand ROS and Ca
2+were measured by flow cytometry and the results are shown in
Figure 4. The data indicate that PEITC significantly reduced
the level of ΔΨ
mwithin 48 and 72 h treatment of PEITC
(Figure 4A) and promoted ROS production at 6 and 12 h
treatment of PEITC (Figure 4B). However, treatments for 24 h
led to a decrease the ROS levels when compared to the control
group, but not significantly so. Furthermore, PEITC increased
the intracellular Ca
2+level at 0.5-6 h treatment (Figure 4C) in
DU 145 cells.
PEITC promoted the activities of caspase-3, -8 and -9 in DU
145 cells. To determine whether PEITC-induced apoptosis
was associated with caspase activation in DU 145 cells, the
activities of caspase-3, -8 and -9 in DU 145 cells after
exposure to PEITC were assayed by flow cytometry and the
results are shown in Figure 5. The results indicate that
PEITC promoted the activities of caspase-3 at 24-72 h in a
time-dependent manner (Figure 5A), increased the caspase-8
Figure 7. Representative Western blotting showing changes in the levels of apoptosis-associated protein level of DU 145 cells after exposure toPEITC. Cells were treated with 10 μM of PEITC for 0, 6, 12, 24 and 48 h before the total proteins were determined, as described in Materials and Methods. The levels of associated protein expressions (A: FAS ligand and FAS; B: BCL-2, BAK and BID; C: PARP, AIF and cytochrome c; D: GRP78 and GADD153) were estimated by Western blotting, as described in the Materials and Methods.
activity at 48 h, (Figure 5B), and that of caspase-9 at 24 h
(Figure 5C). Based on these observations, we suggest that
caspases might be involved in the induction of apoptosis in
DU 145 cells after exposure to PEITC.
Caspase-specific inhibitors protected against PEITC-induced
cell death in DU 145 cells. DU 145 cells were pretreated with
the caspase-3 inhibitor DEVE-FMK), caspase-8 inhibitor
(Z-IETD-FMK) and caspase-9 inhibitor (Z-LEHD-FMK),
Tang et al: Phenethyl Isothiocyanate Triggers Apoptosis in Human Prostate Cancer CellsFigure 8. PEITC altered the GADD153 and Endo-G distributions in DU 145 cells. Cells were incubated with 10 μM of PEITC for 24 h, and then
were fixed and stained with primary antibodies to GADD153 (A) and Endo G (B) which were stained by FITC-labeled secondary antibodies (green fluorescence) and the proteins were detected by a confocal laser microscopic system. The nuclei were stained by PI (red fluorescence). Areas of co-localization of GADD153 and Endo G expressions in the merged panels are yellow. Scale bar, 20 μm.
respectively, and then cells were harvested for measuring the
percentage of viability and the results are shown in Figure 6.
These data indicate that caspase-8 inhibitor (Z-IETD-FMK)
significantly increased the percentage of viable cells (Figure 6B),
while the other two inhibitors did not (Figure 6A and C) after
exposure to PEITC.
PEITC affected the apoptosis-associated protein levels in DU
145 cells. The effects of PEITC on the apoptotic associated
protein levels in DU 145 cells were assayed by Western
blotting and the results are shown in Figure 7. The results
indicate that PEITC increased the protein levels of FAS
ligand, FAS (Figure 7A), BAK (Figure 7B), AIF, cytochrome
c (Figure 7C) and GRP78 (Figure 7D), but reduced the levels
of BCL-2 and BID (Figure 7B), PARP (Figure 7C) and did
not affect GADD153 (Figure 7D) level in DU 145 cells.
PEITC altered the distribution of GADD153 and Endo G in
DU 145 cells. The effects of PEITC on the levels and
distribution of apoptosis-associated proteins GADD153 and
Endo G in DU 145 cells were examined by confocal laser
microscope and the results are shown in Figure 8. PEITC
increased the protein levels of GADD153 (Figure 8A) and
Endo G (Figure 8B) and increased their translocation to nuclei.
Discussion
PEITC has been isolated from various common
vegetables, especially cruciferous, and has anticancer
activities against many human cancer cell types in vitro
and in vivo (34-39). In this study, we found that PEITC
exhibited antitumor activity as it significantly inhibited
prostate cancer cell growth and induced apoptosis, and
more importantly, through mitochondrialdependent and
-independent pathways.
It is well known that cell proliferation and viability are
one of major observations for the assessment of cytotoxicity
and screening of anticancer drugs. Our results also show that
PEITC induced morphological changes and reduced the
percentage of viable of DU 145 cells (Figure 1A and B).
Apoptosis is one form of programmed cell death and is
characterized by morphological changes, chromatin
condensation and extensive DNA fragmentation. The
frequency and time of appearance of apoptosis depend on the
apoptosis-inducing signal and cell types (40, 41). It was
reported that failure of apoptosis in cancer cells may promote
survival and accumulation of cells to form tumors (42).
Therefore, the induction of apoptosis in cancer cells has
become a strategy for their elimination (43, 44).
In the present study, we investigated the effect of PEITC
on apoptosis in DU 145 human prostate cancer cells in vitro.
Flow cytometric assay for sub-G
1phase examination, DAPI
staining and the comet assay were used to study the
morphological changes of apoptosis and DNA damage,
respectively. Results showed that PEITC induced sub-G
1phase in DU 145 cells and the data from DAPI staining also
confirmed that PEITC induced DU 145 cells apoptosis and
that the DNA damage occurred in the cell apoptotic process.
To further verify the DNA damage induced by PEITC, the
single-cell gel electrophoresis assay (Comet assay) was
performed and the results showed that PEITC induced DNA
damages dose dependently. We suggest that DNA damage
might be one of the means by with apoptosis is induced by
PEITC in DU 145 cells.
It is also reported that ROS are involved in apoptosis in
cancer cells (45). Thus, we used flow cytometric assay and
found that PEITC promoted the release of ROS in DU 145
cells (Figure 4B) at 6-12 h treatment. The results also
showed that PEITC reduced the levels of ΔΨ
mand
resulted in Ca
2+increased in DU 145 cells (Figure 4A and
C). These observations indicate that PEITC-induced
apoptosis also involves mitochondria. It is well known that
caspases also play important roles in apoptosis, and we
investigated the activities of caspases and found that
PEITC significantly activated of caspase-3, -8 and -9
(Figure 6A, B and C). PEITC-induced apoptosis involves a
caspase-dependent pathway. Caspase-3 is a critical enzyme
in the execution phase of apoptosis and a hallmark of
apoptosis (46). Our findings are in agreement with this
showing a significant increase in caspase-3 activity in DU
145 cells treated with 10 μM PEITC for 24 h, but no
change in caspase-8 protein expression (Figure 5B). This
shows that the treatment time was insufficient for an
increase in protein expression but sufficient for
enhancement of activity.
The BCL-2 family of proteins are the most important
regulators of apoptosis. BAX is an apoptotic agonist that
promotes cells’ apoptosis. BCL-2 is an intracellular
suppressor of apoptosis, which prolongs cell survival.
Therefore, the ratio of BCL-2/BAX of the cells will
determine whether or not apoptosis occurs (47).
In conclusion, our results clearly indicate that PEITC
promoted the expression of BAX but inhibited the expression
of BCL-2 in DU 145 cells thus contributing to the activation
of caspase-3 and the induction of apoptosis via the
mitochondrial apoptosis pathway (Figure 9).
Acknowledgements
This study was supported in part by Taiwan Department of Health Clinical Trial and Research Center of Excellence (DOH100-TD-B-111-004).
References
1 Bracarda S, de Cobelli O, Greco C, Prayer-Galetti T, Valdagni R, Gatta G, de Braud F and Bartsch G: Cancer of the prostate. Crit Rev Oncol Hematol 56: 379-396, 2005.
2 Jemal A, Siegel R, Ward E, Hao Y, Xu J and Thun MJ: Cancer statistics, 2009. CA Cancer J Clin 59: 225-249, 2009.
3 Taichman RS, Cooper C, Keller ET, Pienta KJ, Taichman NS and McCauley LK: Use of the stromal cell-derived factor-1/CXCR4 pathway in prostate cancer metastasis to bone. Cancer Res 62: 1832-1837, 2002.
4 Woynarowska BA and Woynarowski JM: Preferential targeting of apoptosis in tumor versus normal cells. Biochim Biophys Acta
1587: 309-317, 2002.
5 Debatin K: Activation of apoptosis pathways by anticancer treatment. Toxicol Lett 112-113: 41-48, 2000.
6 Jaffrezou JP, Bettaieb A, Levade T and Laurent G: Antitumor agent-induced apoptosis in myeloid leukemia cells: a controlled suicide. Leuk Lymphoma 29: 453-463, 1998.
7 Arends MJ and Wyllie AH: Apoptosis: mechanisms and roles in pathology. Int Rev Exp Pathol 32: 223-254, 1991.
8 Strasser A, O’Connor L and Dixit VM: Apoptosis signaling. Annu Rev Biochem 69: 217-245, 2000.
9 Degterev A, Boyce M and Yuan J: A decade of caspases. Oncogene 22: 8543-8567, 2003.
10 Decker P and Muller S: Modulating poly (ADP-ribose) polymerase activity: potential for the prevention and therapy of pathogenic situations involving DNA damage and oxidative stress. Curr Pharm Biotechnol 3: 275-283, 2002.
11 Earnshaw WC, Martins LM and Kaufmann SH: Mammalian caspases: structure, activation, substrates, and functions during apoptosis. Annu Rev Biochem 68: 383-424, 1999.
12 Kluza J, Clark AM and Bailly C: Apoptosis induced by the alkaloid sampangine in HL-60 leukemia cells: correlation between the effects on the cell cycle progression and changes of mitochondrial potential. Ann N Y Acad Sci 1010: 331-334, 2003. 13 Mathur A, Hong Y, Kemp BK, Barrientos AA and Erusalimsky JD: Evaluation of fluorescent dyes for the detection of mitochondrial membrane potential changes in cultured cardiomyocytes. Cardiovasc Res 46: 126-138, 2000.
14 Isenberg JS and Klaunig JE: Role of the mitochondrial membrane permeability transition (MPT) in rotenone-induced apoptosis in liver cells. Toxicol Sci 53: 340-351, 2000. 15 Fu Y, Hsieh TC, Guo J, Kunicki J, Lee MY, Darzynkiewicz Z
and Wu JM: Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem Biophys Res Commun 322: 263-270, 2004.
16 Tabata K, Motani K, Takayanagi N, Nishimura R, Asami S, Kimura Y, Ukiya M, Hasegawa D, Akihisa T and Suzuki T: Xanthoangelol, a major chalcone constituent of Angelica keiskei, induces apoptosis in neuroblastoma and leukemia cells. Biol Pharm Bull 28: 1404-1407, 2005.
17 Hsu YL, Kuo PL, Tzeng WS and Lin CC: Chalcone inhibits the proliferation of human breast cancer cell by blocking cell cycle progression and inducing apoptosis. Food Chem Toxicol 44: 704-713, 2006.
18 Conaway CC, Yang YM and Chung FL: Isothiocyanates as cancer chemopreventive agents: their biological activities and metabolism in rodents and humans. Curr Drug Metab 3: 233-255, 2002. Tang et al: Phenethyl Isothiocyanate Triggers Apoptosis in Human Prostate Cancer Cells
19 Zhang Y: Cancer-preventive isothiocyanates: measurement of human exposure and mechanism of action. Mutat Res 555: 173-190, 2004.
20 Zhang Y, Kensler TW, Cho CG, Posner GH and Talalay P: Anticarcinogenic activities of sulforaphane and structurally related synthetic norbornyl isothiocyanates. Proc Natl Acad Sci USA 91: 3147-3150, 1994.
21 Morse MA, Amin SG, Hecht SS and Chung FL: Effects of aromatic isothiocyanates on tumorigenicity, O6-methylguanine
formation, and metabolism of the tobacco-specific nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone in A/J mouse lung. Cancer Res 49: 2894-2897, 1989.
22 Stoner GD, Morrissey DT, Heur YH, Daniel EM, Galati AJ and Wagner SA: Inhibitory effects of phenethyl isothiocyanate on N-nitrosobenzylmethylamine carcinogenesis in the rat esophagus. Cancer Res 51: 2063-2068, 1991.
23 Xiao D, Powolny AA, Moura MB, Kelley EE, Bommareddy A, Kim SH, Hahm ER, Normolle D, Van Houten B and Singh SV: Phenethyl isothiocyanate inhibits oxidative phosphorylation to trigger reactive oxygen species-mediated death of human prostate cancer cells. J Biol Chem 285: 26558-26569, 2010. 24 Liu KC, Huang YT, Wu PP, Ji BC, Yang JS, Yang JL, Chiu TH,
Chueh FS and Chung JG: The roles of AIF and Endo G in the apoptotic effects of benzyl isothiocyanate on DU 145 human prostate cancer cells via the mitochondrial signaling pathway. Int J Oncol 38: 787-796, 2011.
25 Lan YH, Wu YC, Wu KW, Chung JG, Lu CC, Chen YL, Wu TS and Yang JS: Death receptor 5-mediated TNFR family signaling pathways modulate gamma-humulene-induced apoptosis in human colorectal cancer HT29 cells. Oncol Rep 25: 419-424, 2011. 26 Yang JS, Hour MJ, Huang WW, Lin KL, Kuo SC and Chung JG:
MJ-29 inhibits tubulin polymerization, induces mitotic arrest, and triggers apoptosis via cyclin-dependent kinase 1-mediated Bcl-2 phosphorylation in human leukemia U937 cells. J Pharmacol Exp Ther 334: 477-488,
27 Wu SH, Hang LW, Yang JS, Chen HY, Lin HY, Chiang JH, Lu CC, Yang JL, Lai TY, Ko YC and Chung JG: Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways. Anticancer Res 30: 2125-2133, 2010. 28 Lu HF, Wang HL, Chuang YY, Tang YJ, Yang JS, Ma YS,
Chiang JH, Lu CC, Yang JL, Lai TY, Wu CC and Chung JG: Danthron induced apoptosis through mitochondria- and caspase-3-dependent pathways in human brain glioblastoma multiforms GBM 8401 cells. Neurochem Res 35: 390-398, 2010.
29 Chiang JH, Yang JS, Ma CY, Yang MD, Huang HY, Hsia TC, Kuo HM, Wu PP, Lee TH and Chung JG: Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chem Res Toxicol 24: 20-29, 2011. 30 Ji BC, Hsu WH, Yang JS, Hsia TC, Lu CC, Chiang JH, Yang JL,
Lin CH, Lin JJ, Suen LJ, Gibson Wood W and Chung JG: Gallic acid induces apoptosis via caspase-3 and mitochondrion-dependent pathways in vitro and suppresses lung xenograft tumor growth in vivo. J Agric Food Chem 57: 7596-7604, 2009. 31 Lu CC, Yang JS, Huang AC, Hsia TC, Chou ST, Kuo CL, Lu HF, Lee TH, Wood WG and Chung JG: Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Mol Nutr Food Res 54: 967-976, 2010.
32 Yu FS, Yang JS, Yu CS, Lu CC, Chiang JH, Lin CW and Chung JG: Safrole induces apoptosis in human oral cancer HSC-3 cells. J Dent Res 90: 168-174, 2011.
33 Chen JC, Lu KW, Tsai ML, Hsu SC, Kuo CL, Yang JS, Hsia TC, Yu CS, Chou ST, Kao MC, Chung JG and Wood WG: Gypenosides induced G0/G1 arrest via CHk2 and apoptosis through endoplasmic reticulum stress and mitochondria-dependent pathways in human tongue cancer SCC-4 cells. Oral Oncol 45: 273-283, 2009.
34 Bommareddy A, Hahm ER, Xiao D, Powolny AA, Fisher AL, Jiang Y and Singh SV: Atg5 regulates phenethyl isothiocyanate-induced autophagic and apoptotic cell death in human prostate cancer cells. Cancer Res 69: 3704-3712, 2009.
35 Moon YJ, Brazeau DA and Morris ME: Dietary phenethyl isothiocyanate alters gene expression in human breast cancer cells. Evid Based Complement Alternat Med 2011, 2011. 36 Cheung KL and Kong AN: Molecular targets of dietary
phenethyl isothiocyanate and sulforaphane for cancer chemoprevention. AAPS J 12: 87-97, 2010.
37 Xiao D and Singh SV: p66Shc is indispensable for phenethyl isothiocyanate-induced apoptosis in human prostate cancer cells. Cancer Res 70: 3150-3158, 2010.
38 Yang MD, Lai KC, Lai TY, Hsu SC, Kuo CL, Yu CS, Lin ML, Yang JS, Kuo HM, Wu SH and Chung JG: Phenethyl isothiocyanate inhibits migration and invasion of human gastric cancer AGS cells through suppressing MAPK and NF-kappaB signal pathways. Anticancer Res 30: 2135-2143, 2010. 39 Wang LG and Chiao JW: Prostate cancer chemopreventive
activity of phenethyl isothiocyanate through epigenetic regulation (review). Int J Oncol 37: 533-539, 2010.
40 Arends MJ, Morris RG and Wyllie AH: Apoptosis. The role of the endonuclease. Am J Pathol 136: 593-608, 1990.
41 Bortner CD, Oldenburg NB and Cidlowski JA: The role of DNA fragmentation in apoptosis. Trends Cell Biol 5: 21-26, 1995. 42 Hoeppner DJ, Hengartner MO and Fisher DE: Programmed cell
death: from development to disease. Meeting report. Biochim Biophys Acta 1242: 217-220, 1996.
43 Hong WK, Sporn MB: Recent advances in chemoprevention of cancer. Science 278: 1073-1077, 1997.
44 Kelloff GJ, Crowell JA, Steele VE, Lubet RA, Malone WA, Boone CW, Kopelovich L, Hawk ET, Lieberman R, Lawrence JA, Ali I, Viner JL and Sigman CC: Progress in cancer chemoprevention: development of diet-derived chemopreventive agents. J Nutr 130: 467S-471S, 2000.
45 Zhang R, Humphreys I, Sahu RP, Shi Y and Srivastava SK: In
vitro and in vivo induction of apoptosis by capsaicin in
pancreatic cancer cells is mediated through ROS generation and mitochondrial death pathway. Apoptosis 13: 1465-1478, 2008. 46 Stennicke HR and Salvesen GS: Properties of the caspases.
Biochim Biophys Acta 1387: 17-31, 1998.
47 Yong WP, Innocenti F and Ratain MJ: The role of pharmaco-genetics in cancer therapeutics. Br J Clin Pharmacol 62: 35-46, 2006.