Interaction between NBS1 and the mTOR/Rictor/SIN1 complex
through specific domains
Jian-Qiu Wang1,2,5, Jian-Hong Chen1,5, Yen-Chung Chen1,4, Mei-Yu Chen1, Chia-Ying
Hsieh1, Shu-Chun Teng3, and Kou-Juey Wu1*
Authors’ Affiliations: 1Institute of Biochemistry and Molecular Biology, National
Yang-Ming University, Taipei 112, Taiwan; 2Institute of Aging Research, Department
of Basic Medical Science, School of Medicine, Hangzhou Normal University, Hangzhou, 310036, China; 3Institute of Microbiology, College of Medicine, National
Taiwan University, Taipei 100, Taiwan
4:Present address: Taiwan Advance Biopharm (TABP), Inc., Xizhi city, New Taipei City
221, Taiwan
5: These authors contributed equally to the work.
*:Correspondence should be addressed to:
Kou-Juey Wu, Institute of Biochemistry and Molecular Biology, National Yang-Ming University, No.155, Li-Nong St., Sec.2, Peitou, Taipei 112, Taiwan; Email: [email protected]; Tel: 886-228267328; Fax: 886-228264843
Abstract
Nijmegen breakage syndrome (NBS) is a chromosomal-instability syndrome. The NBS gene product, NBS1 (p95 or nibrin), is a part of the Mre11-Rad50-NBS1 complex. SIN1 is a component of the mTOR/Rictor/SIN1 complex mediating the activation of Akt. Here we show that NBS1 interacted with mTOR, Rictor, and SIN1. The specific domains of mTOR, Rictor, or SIN1 interacted with the internal domain (a.a. 221-402) of NBS1. Sucrose density gradient showed that NBS1 was located in the same fractions as the mTOR/Rictor/SIN1 complex. Knockdown of NBS1 decreased the levels of phosphorylated Akt and its downstream targets. Ionizing radiation (IR) increased the NBS1 levels and activated Akt activity. These results demonstrate that NBS1 interacts with the mTOR/Rictor/SIN1 complex through the a.a. 221-402 domain and contributes to the activation of Akt activity.
Introduction
Nijmegen breakage syndrome (NBS) is a chromosomal-instability syndrome with the manifestation of cancer predisposition, radiosensitivity, microcephaly, and growth retardation [1-3]. The NBS gene product, NBS1 (p95 or nibrin), is a part of the Mre11-Rad50-NBS1 complex that is essential for DNA double strand break repair [1,2]. NBS1 carries out its checkpoint functions through the phosphorylation by ataxia-telangiectasia mutated (ATM) protein after ionizing radiation [4-6]. We previously demonstrated that c-MYC, an oncoprotein, directly activates the expression of NBS1 [7]. The function of NBS1 related to proliferation is demonstrated by the phenotypes of diminished expansion of the inner cell mass of mutant blastocysts (Nbs1 null) and cellular proliferation defects in Nbs1m/m mouse
embryonic fibroblasts [8-10]. Overexpression of NBS1 induces transformation activity through interacting with the p110 subunits of phosphoinositide kinase (PI 3-kinase) to activate PI 3-kinase activity [11, 12], indicating that NBS1 overexpression is an oncogenic event. In head and neck squamous cell carcinoma patients, increased NBS1 expression is a prognostic factor of aggressive head and neck cancer [13]. All these results indicate that NBS1 overexpression may play an important role in tumorigenesis.
through phosphorylation at Thr-308 by phosphoinositide-dependent protein kinase 1 followed by phosphorylation at Ser-473 by the mTOR/Rictor (mTORC2) complex to achieve full activation [16,17]. Activated Akt then regulates a wide range of target proteins that control cell proliferation (e.g. GSK-3β, Foxo1/3a), survival (e.g. BAD), and cell growth (e.g. mTOR) [15]. Akt regulates critical processes of tumorigenesis and plays an important role in oncogenesis [16]. Altered expression or mutation of many components of the PI3K/Akt pathway has been implicated in human cancer [15,18]. Recently, a new member of the mTORC2 complex, SIN1, was identified [19-21]. SIN1 is critical to maintain the mTORC2 complex and regulate Akt kinase activity and substrate specificity [19-21]. Other example such as TSC1-TSC2 complex was shown to associate with the mTORC2 complex and activate Akt kianse activity (Akt Ser-473 phosphorylation)[22]. However, it remains to be explored whether other protein could participate in the mTORC2 complex to induce Akt kinase activity.
In this report, we demonstrate that NBS1 interacts with the mTOR/Rictor/SIN1 complex using a specific domain (a.a. 221-402). The domain in mTOR, Rictor, or SIN1 that interacts with NBS1 is also mapped. Knockdown of NBS1 decreases Akt kinase activity. IR increased the levels of NBS1 and phosphorylated Akt. These results indicate that NBS1 is a critical component of the mTOR/Rictor/SIN1 complex
Results
Interaction between NBS1 and the components of the mTOR/Rictor/SIN1 complex
We previously demonstrated that NBS1 interacted with the p110α subunit of the PI 3-kinase to induce PI 3-kinase/Akt activity [12]. However, it is possible that NBS1 may also interact with Akt to induce Akt activity. To test whether there is interaction between NBS1 and the mTOR/Rictor/SIN1 complex that is responsible for the activation of Akt activity, co-immunoprecipitation assays were performed between NBS1 and each component of the mTOR/Rictor/SIN1 complex. The results showed that the anti-NBS1 antibody pulled down mTOR in 293T cells overexpressing both NBS1 and mTOR (Fig. 1A). In addition, the anti-mTOR antibody also pulled down NBS1 (Fig. 1B), demonstrating their interaction when both proteins were overexpressed in 293T cells. Similar assays were performed to test the interaction between NBS1 and Rictor and the results showed that NBS1 interacted with Rictor in 293T cells overexpressing both proteins (Fig. 1C-D). Finally, similar assays were performed to test the interaction between NBS1 and SIN1β, which also showed the interaction between these two proteins (Fig. 1E-F). All the results demonstrated that NBS1 interacted with the mTOR/Rictor/SIN1 complex.
Mapping of domain in mTOR or Rictor interacting with NBS1
In order to map the domain in mTOR or Rictor that interacts with NBS1, different truncation mutants of mTOR or Rictor were generated. Different truncation mutants of mTOR (mTOR1-651, mTOR640-1418, and mTOR1401-2549) were co-expressed with NBS1 in 293T cells followed by co-immunoprecipitation. The results showed that only mTOR1-651 interacted with NBS1 (Fig. 2A-B). The full length and different truncation mutants of Rictor (Rictor1-1708, Rictor1-789, and Rictor664-1708) were co-expressed with NBS1 in 293T cells followed by co-immunoprecipitation. Co-immunoprecipitation experiments showed that the domain 1-789 a.a. of Rictor interacted with NBS1 (Fig. 2C-D). These results indicated that the N-terminal domains (1-651 a.a. domain of mTOR and 1-789 a.a. domain of Rictor) of both proteins interacted with NBS1.
Mapping of the NBS1 domain interacting with mTOR or Rictor
In order to map the domain in NBS1 that interacts with mTOR or Rictor, different truncation mutants of NBS1 (C-terminal truncation mutant NBS1-653 and N-terminal truncation mutant NBSp70) were expressed with mTOR or Rictor followed by co-immunoprecipitation [12]. The result showed that both the mutants interacted with mTOR and Rictor (Fig. 3A, C). Further fine mapping of the domain using two
different NBS1 truncation mutants (NBS221-402 and NBS402-653) together with mTOR or Rictor showed that only NBS221-402 interacted with mTOR and Rictor (Fig. 3B, D). All the results indicated that the internal domain (a.a. 221-402) of NBS1 interacted with both mTOR and Rictor.
Mapping of the interacting domain between NBS1 and SIN1β
To map the domain in SIN1β that interacts with NBS1, different SIN1β truncation mutants (SIN1β1-267, SIN1β266-487, SIN1β336-487) were co-expressed with NBS1 in 293T cells followed by co-immunoprecipitation. The result showed that only SIN1β1-267 interacted with NBS1 (Fig. 4A). To map the domain in NBS1 interacting with SIN1β, different NBS1 truncation mutants as mentioned above were co-expressed with SIN1β in 293T cells followed by co-immunoprecipitation. The result showed that the domain in NBS1 interacting with SIN1β was also mapped to the a.a. 221-402 domain of NBS1 (Fig. 4B).
In vivo interaction between NBS1 and the mTOR/Rictor/SIN1 complex
In order to test whether NBS1 indeed interacts with the mTOR/Rictor/SIN1 complex, co-immunoprecipitation assays using extracts from a lung cancer cell line H1299 were used. The result showed that the anti-NBS1 antibody pulled down mTOR, Rictor, and SIN1β (Fig. 5A, left panel), supporting their interaction in vivo.
Co-immunoprecipitation experiment using the anti-Raptor antibody did not pull down the whole mTOR/Rictor/SIN1 complex or NBS1 (Fig. 5A, right panel), indicating that NBS1 did not interact with the mTOR/Raptor complex. Finally, a sucrose density gradient experiment using extracts from H1299 cells was performed to test whether these proteins were localized in the same fraction. The result showed that indeed NBS1, mTOR, Rictor, and SIN1β were localized in the same fraction (fraction 4 is the major fraction) (Fig. 5B). All the results indicated that NBS1 interacted with the mTOR/Rictor/SIN1 complex in vivo.
Knockdown of NBS1 decreased the levels of phosphorylated Akt and its downstream targets
In order to test the correlation between NBS1 levels and its effect on the Akt activity, siRNA experiment to knockdown NBS1 was performed. The result showed that knockdown of NBS1 decreased the phosphorylated Akt levels (pAkt Ser-473) (Fig. 6), indicating that NBS1 contributed to the Akt activity. In addition, the phosphorylation levels of certain Akt downstream targets such as GSK-3β and Foxo1/3a were also decreased following NBS1 knockdown (Fig. 6A). These results supported the role of NBS1 in the activation of Akt activity.
Since IR was shown to activate Akt signaling [23,24], we tested the physiological relevance of NBS1 to activate Akt activity through IR. IR increased the levels of NBS1 and phosphorylated Akt (pAkt Ser-473) in two different cell lines (Fig. 6B). Knockdown of NBS1 in H1299 cells abolished the increase in the levels of NBS1 and phosphorylated Akt under IR treatment (Fig. 6C). These results supported the physiological role of NBS1 to activate Akt activity.
Discussion
Overexpression of NBS1 was previously shown to induce transformation through the PI 3-kinase/Akt pathway [11-13]. However, in this report, we demonstrated that NBS1 overexpression could also interact with the mTOR/Rictor/SIN1 complex to activate Akt activity. This result is the further extension of the ability of NBS1 to interact with both the upstream PI3K and downstream Akt to regulate the activity of PI3K/Akt pathway. It is also supported by recent literatures demonstrating the dual function of NBS1 in the repair of DNA breaks and proliferation and the requirement of NBS1 in IGF-1 induced cellular proliferation [25,26]. In addition, the ability of RNA regulon to promote Akt activity also goes through the upregulation of NBS1 levels [27]. Both results support the role of NBS1 in the activation of Akt activity.
SIN1 was the critical molecule recently demonstrated to be important for the activation of Akt activity through interacting with the mTOR/Rictor complex [19-21]. Other molecules that could interact with the mTOR/Rictor complex were also recently identified including PRR5 and the TSC1/TSC2 complex [22,28]. The ability of NBS1 to interact with each component of the mTOR/Rictor/SIN1 complex showed that NBS1 may be another critical component of the mTOR/Rictor/SIN1 complex, which was supported by the result that knockdown of NBS1 decreased the phosphorylation levels of Akt and its downstream targets (Fig. 6A). In addition, knockdown of NBS1
abolished the increase in NBS1 and phosphorylated Akt levels under IR, supporting the physiological relevance of NBS1 in the activation of Akt activity (Fig. 6B-C). This result also points to the role of a DNA double strand break repair protein to activate Akt activity. It is interesting that NBS1 uses the same internal domain (a.a. 221-402) to interact with each component of the mTOR/Rictor/SIN1 complex. This domain contains the ATM-dependent phosphorylation sites (Ser278, Ser343, Ser397). In contrast, NBS1 uses the conserved C-terminal domain (a.a. 653-669) to interact with the subunits of PI3K to achieve its activation [12]. Further structural analysis of co-crystalized NBS1 and the mTOR/Rictor/SIN1 complex will delineate the molecular interactions between these molecules. Our previous results showed that increased NBS1 expression was constantly observed in different tumor samples [11,13,29], suggesting that increased NBS1 levels in tumor tissues may contribute significantly to the increased Akt activity observed in different tumor samples. Our discovery points to the unique role of NBS1 in the activation of the PI3K/Akt pathway using a newly identified internal domain (a.a 221-402) to interact with the mTOR/Rictor/SIN1 complex to achieve activation.
Materials and Methods
Cell lines, plasmids, transfections, and stable clones
The human embryonic kidney 293T and H1299 (a non-small cell lung cancer cell line) were previously described [12, 30]. The OCEM-1 cell line is a head and neck cancer line obtained from ATCC. The Flag-NBS1 full length and its truncation mutants were described [12]. The pSUPER-NBS1 and pSUPER-scrambled-si constructs were described [12]. The full length HA-mTOR expression vector was constructed by assembling fragments containing the nucleotides 1-4254 and 4201-7650 of the mTOR gene that were amplified from the human Marathon-Ready brain cDNA (Clontech) by PCR and subcloned into the yTA (Yeastern) vector. The
sequences of primers used were
5’-CAATTGGTCGACATGCTTGGAACCGGACCTGC-3’ and
5’-GCTAGCGCGCTAGCTGATGAGAGATTCTAGAATGG-3’ for mTOR (1-4254),
and 5’-GAACTGGAGTTCCAGTTCCAGAAAGGC-3’ and
5’-CAATTGGCGCGCTTACCAGAAAGGGCACCAGC-3’ for mTOR (4201-7650). The yTA-mTOR(1-4254) and yTA-mTOR(4201-7650) plasmids were digested by
SalI and BssHII and by XbaI and BssHII, respectively. Each mTOR fragment was
ligated into the pDNR-CMV vector (Clontech). The SalI-XbaI mTOR fragment from the mTOR(1-4254) plasmid was ligated into the
pDNR-CMV-mTOR(4201-7650) plasmid to generate the full length pDNR-CMV-mTOR(1-7650) expression vector. The HA epitope-tagged full-length mTOR was generated following the BD Creator cloning protocol (Clontech) using the pLP-CMV-HA vector as the acceptor plasmid. The Myc-Rictor expression vector was generated by assembling PCR fragments amplified from human Marathon-Ready brain cDNA (Clontech) to make the full-length Rictor expression vector. The ATG-containing 5’ primer and the stop codon-containing 3’ primer (described in Table 1) introduced SalI and KpnI restriction sites, respectively. The final assembled fragment of Rictor (nucleotides 1-5191) was digested by the two endonucleases and ligated into the pCMV-Myc (Clontech) plasmid to generate the full length pCMV-pCMV-Myc-Rictor (1-5191) expression vector. The HA-mTOR and Myc-Rictor truncation mutants were constructed by using the appropriate primers and PCR to generate different truncation mutants [31,32] (Table 1). Since there are different SIN1 splice variants [33], only the SIN1β full length expression vector was generated by using RT-PCR of mRNA from 293T cells followed by inserting into the CMV-HA vector. The oligonucleotides used to generate the full length SIN1β and its truncation mutants were also described in Table 1.
Western blot analysis was performed as described [12]. For Western blot analysis, 50 µg protein extracts from each clone were loaded to 10% SDS-PAGE gels and transferred to nitrocellulose filters. The filters were probed with various antibodies (described in Table 2) and an anti-ß-actin antibody was selected as a loading control. Signals were developed using an ECL chemiluminescence kit (Amersham Biosciences, U.K.).
Co-immunoprecipitation assays were performed as described [12,34]. Briefly, the anti-NBS1, anti-mTOR, anti-Rictor, anti-SIN1, anti-HA, or anti-Myc antibody was incubated for 5 h with 500 l of whole cell extracts prepared by lysis in 150 mM NaCl, 1% Nonidet P-40, 1% deoxycholate, 0.1% SDS, 50 mM Tris.HCl-pH 7.5, and protease inhibitors, from 293T cells overexpressing proteins of interest or from H1299 cells to detect in vivo interaction. The immune complexes were incubated overnight with protein-A beads, preblocked with 10% BSA. The immunoprecipitates were washed 3 times with TNTG buffer (20 mM Tris.HCl-pH 7.5, 150 mM NaCl, 0.1% Triton-X 100, 10% glycerol), mixed with 1x Laemmli dye, boiled for 10 min and loaded on SDS- polyacrylamide (SDS-PAGE) gels [12]. After transfer, the filters were blocked with blocking buffer [12], probed with primary and secondary antibody sequentially and developed. The antibodies used are described in Table 2. Data shown are representative of at least 2 experimentsfrom independent cell cultures or different
transfections.
Sucrose density gradient analysis
A sucrose density gradient was generated as described [35] and the extracts of H1299 cells were loaded to the continuous gradient. After centrifugation, the proteins were collected from different fractions, run on SDS-PAGE, and transferred to membranes. The blot was probed with various antibodies to detect the presence of different components of mTOR/Rictor/SIN1 and NBS1.
Ionizing radiation (IR)
Exponentially growing cells were temporarily incubated with 2 ml of PBS in 10 cm dishes and were irradiated at room temperature using 60Co γ-ray source with a
dose of 5.0 Gy (dose rate 3 Gy/min). After irradiation, cells were washed twice with PBS and cultured in fresh DMEM medium containing 10% FBS for another 12 hours followed by harvesting for Western blot analysis.
Acknowledgments
References
[1] Czorna K, Chunghtai S, Chrzanowska KH (2008) Mystery of DNA repair: the role of MRN complex and ATM kinase in DNA damage repair. J Appl Genet 49: 383-396.
[2] Antoccia A, Kobayashi J, Tauchi H, Matsuura S, Komatsu K (2006) Nijmegen breakage syndrome and functions of the responsible protein, NBS1. Genome Dyn 1: 191-205.
[3] D’Amours D, Jackson SP (2002) The Mre11 complex: at the crossroads of DNA repair and checkpoint signaling. Nature Rev Mol Cell Biol 3: 317-327.
[4] Lim D S, Kim ST, Xu B, Maser RS, Lin J, et al. (2000) ATM phosphorylates p95/nbs1 in an S-phase checkpoint pathway. Nature 404: 613-617.
[5] Zhao S, Weng YC, Yuan SS, Lin YT, Hsu HC, et al. (2000) Functional link between ataxia-telangiectasia and Nijmegen breakage syndrome gene products. Nature 405: 474-477.
[6] Wu X, Ranganathan V, Weisman DS, Heine WF, Ciccone DN, et al. (2000) ATM phosphorylation of Nijmegen breakage syndrome protein is required in a DNA damage response. Nature 405: 477-482.
[7] Chiang YC, Teng SC, Su YN, Hsieh FJ, Wu KJ (2003) c-MYC directly regulates the transcription of NBS1 gene involved in DNA double-strand break repair. J Biol Chem 278: 19286-19291.
[8] Zhu J, Petersen S, Tessarollo L, Nussenzweig A (2001) Targeted disruption of the Nijmegen breakage syndrome gene NBS1 leads to early embryonic lethality in mice. Curr Biol 11: 105-109.
[9] Williams BR, Mirzoeva OK, Morgan WF, Lin J, Dunnick W, et al. (2002) Murine model of Nijmegen breakage syndrome. Curr Biol 12: 648-53.
[10] Kang J, Bronson RT, Xu Y (2002) Targeted disruption of NBS1 reveals its roles in mouse development and DNA repair. EMBO J 21: 1447-55.
[11] Chen YC, Su YN, Chou PC, Chiang WC, Chang MC, et al. (2005) Overexpression of NBS1 contributes to transformation through the activation of phosphatidylinositol 3-kinase/Akt. J Biol Chem 280: 32505-32511.
[12] Chen YC, Chiang HY, Yang MH, Chen PM, Chang SY, et al. (2008) Activation of phosphoinositide 3-kinase by the NBS1 DNA repair protein. J Mol Med 86: 401-412.
[13] Yang MH, Chiang WC, Chang SY, Chen PM, Teng SC, et al. (2006) Increased NBS1 expression is a marker of aggressive head and neck cancer and overepxression of NBS1 promotes transformation. Clin Cancer Res 12: 507-515. [14] Cantley LC (2002) The phosphoinositide 3-kinase pathway. Science 296:
1655-1657
[15] Vivanco I, Sawyers CL (2002) The phosphatidylinositol 3-kinase-Akt pathway in human cancer. Nature Rev Cancer 2: 489-501
[16] Bellacosa A, Chan TO, Ahmed NN, Datta K, Malstrom S, et al. (1998) Akt activation by growth factors is a multiple-step process: the role of the PH domain. Oncogene 17: 313-325
[17] Sarbassov DD, Guertin DA, Ali SM, Sabatini DM (2005) Phosphorylation and regulation of Akt/PKB by the Rictor-mTOR complex. Science 307: 1098-1101. [18] Testa JR, Bellacosa A (2001) AKT plays a central role in tumorigenesis. Proc
Natl Acad Sci USA 98: 10983-10985
[19] Jacinto E, Facchinetti V, Liu D, Soto N, Wei S, et al. (2006) SIN1/MIP1 maintains rictor-mTOR complex integrity and regulates Akt phosphorylation and substrate specificity. Cell 126: 1-13.
[20] Yang Q, Inoki K, Ikenoue T, Guan KL (2006) Identification of Sin1 as an essential TORC2 component required for complex formation and kinase activity. Genes & Dev 20: 2820-2832.
[21] Frias MA, Thoreen CC, Jaffe JD, Schroder W, Sculley T, et al. (2006) mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s. Curr Biol 16: 1865-1870.
complex is required for proper activation of mTOR complex 2. Mol Cell Biol 28: 4104-4115.
[23] Contessa JN, Hampton J, Lammering G, Mikkelsen RB, Dent P, Valerie K, Schmidt-Ullrich RK (2002) Ionizing radiation activates Erb-B receptor dependent Akt and p70 S6 kinase signaling in carcinoma cells. Oncogene 21: 4032-4041.
[24] Suman S, Johnson MD, Fornace AJ Jr, Datta K (2012) Exposure to ionizing radiation causes long-term increase in serum restradiol and activation of PI3K-Akt signaling in mouse mammary gland. Int J Radiat Oncol Biol Phys 84: 500-507.
[25] Saidi A, Li T, Weih F, Concannon P, Wang ZQ (2010) Dual function of Nbs1 in the repair of DNA breaks and proliferation ensure proper V(D)J recombination and T-cell development. Mol Cell Biol 30: 5572-5581.
[26] Hematulin A, Sagan D, Eckardt-Schupp F, Moertl S (2008) NBS1 is required for IGF-1 induced cellular proliferation through the Ras/Raf/MEK/ERK cascade. Cell Signal 20: 2276-2285.
[27] Culjkovic B, Tan K, Orolicki S, Amri A, Meloche S, et al. (2008) The eIF4E RNA regulon promotes the Akt signaling pathway. J Cell Biol 181: 51-63.
[28] Woo SY, Kim DH, Jun CB, Kim YM, Vander-Haar E, et al. (2007) PRR5, a novel component of mTOR complex 2, regulates PDGFβ expression and signaling. J Biol Chem 282: 25604-25612.
[29] Yang MH, Chang SY, Chiou SH, Liu CJ, Chi CW, et al. (2007) Overexpression of NBS1 induces epithelial-mesenchymal transition and co-expression of NBS1 and Snail predicts metastasis of head and neck cancer. Oncogene 26:1459-1467. [30] Yang MH, Wu MZ, Chiou SH, Chen PM, Chang SY, et al. (2008) Direct
regulation of TWIST by HIF-1α promotes metastasis. Nature Cell Biol 10: 295-305.
[31] Toral-Barza L, Zhang WG, Lamison C, LaRocque J, Gibbons J, et al. (2005) Characterization of the cloned full-length and a truncated human target of rapamycin: activity, specificity, and enzyme inhibition as studied by a high capacity assay. Biochem Biophy Res Comm 332: 304-310.
[32] Sarbassov DD, Ali SM, Kim DH, Guertin DA, Latek RR, et al. (2004) Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and Raptor-independent pathway that regulates the cytoskeleton. Curr Biol 14: 1296-1302. [33] Schroder W, Cloonan N, Bushell G, Sculley T (2004) Alternative
polyadenylation and splicing of mRNAs transcribed from the human Sin1 gene. Gene 339: 17-23.
[34] Tsai YP, Yang MH, Huang CH, Chang SY, Chen PM, et al. (2009) Interaction between HSP60 and β-catenin promotes metastasis. Carcinogenesis 30: 1049-1057.
[35] Damaraju S, Zhang N, Li N, Tao L, Damaraju VL, et al. (2010) Evidence for copurification of micronuclei in sucrose density gradient-enriched plasma membranes from cell lines. Anal Biochem 396: 69-75.
Figure Legends
Figure 1. Interaction between NBS1 and the mTOR/Rictor/SIN1 complex. A & B. Co-immunoprecipitation assays showed the interaction between NBS1 and mTOR in 293T cells overexpressing both proteins. C & D. Co-immunoprecipitation assays showed the interaction between NBS1 and Rictor in 293T cells overexpressing both proteins. E & F. Co-immunoprecipitation assays showed the interaction between NBS1 and SIN1β in 293T cells overexpressing both proteins.
Figure 2. Mapping of the domain in mTOR or Rictor interacting with NBS1. A & B. Co-immunoprecipitation assays showed the interaction between the N-terminal domain of mTOR and NBS1. Whole cell extracts were shown in A. C & D. Co-immunoprecipitation assays showed the interaction between the N-terminal domain of Rictor and NBS1. Whole cell extracts were shown in C.
Figure 3. Mapping of the domain in NBS1 interacting with mTOR or Rictor. A & B. Co-immunoprecipitation assays showed the interaction of the internal domain (a.a. 221-402) of NBS1 with mTOR. C & D. Co-immunoprecipitation assays showed the interaction of the internal domain of NBS1 with Rictor.
A. Co-immunoprecipitation assays showed the interaction of the N-terminal domain (a.a. 1-267) of SIN1β with NBS1. B. Co-immunoprecipitation assays showed the interaction of the internal domain (a.a. 221-402) of NBS1 with SIN1β.
Figure 5. Co-immunoprecipitation assays and sucrose density gradient analysis showed the interaction between NBS1 and the mTOR/Rictor/SIN1 complex in vivo. A. Co-immunoprecipitation assays showed the in vivo interaction of NBS1 with the mTOR/Rictor/SIN1 complex using extracts from H1299 cell line. B. Sucrose density gradient analysis showed the co-localization in the same fractions between NBS1 and the mTOR/Rictor/SIN1 complex.
Figure 6. siRNA mediated knockdown of NBS1 decreased the levels of phosphorylated Akt (pAkt Ser-473) and its downstream targets and IR increased the levels of NBS1 and phosphorylated Akt. A. Western blot analysis showed that knockdown of NBS1 decreased the phosphorylated levels of Akt, GSK-3β, and Foxo1/3a). B. Ionizing radiation of two different cell lines (H1299, OCEM-1) increased the levels of NBS1 and phosphorylated Akt. C. Knockdown of NBS1 in H1299 cells abolished the increase in NBS1 and phosphorylated Akt levels under IR.
Table 1. The oligonucleotide sequences used to generate various full length and truncation constructs of mTOR, Rictor, and SIN1β.
DNA constructs Sequence (5’ 3’) Restriction
Enzyme site Ampli -con (bp) mTOR constructs pCMV-HA-mTOR1-2549 (full length)
F:CAATTGGTCGACATGCTTGGAACCGGACCTGC MfeI, SalI 7674 R:CAATTGGCGCGCTTACCAGAAAGGGCACCAGC MfeI, BssHII
pCMV-HA-mTOR1-651 F: CAATTGGTCGACATGCTTGGAACCGGACCTGC MfeI, SalI 1977
R:GCTAGCGCGCTACACTTGCACTGCGGTCTGGC NheI, BssHII
pCMV-HA-mTOR640-1418 F: CAATTGGTCGACCATGCTCATGTGGTTAGCC MfeI, SalI 2361
R:GCTAGCGCGCTAGCTGATGAGAGATTCTAGAA TGG
NheI, BssHII
pCMV-HA-mTOR1401-2549 F: GAACTGGAGTTCCAGAAAGGC XbaI from
vector
3466 R: CAATTGGCGCGCTTACCAGAAAGGGCACCAGC MfeI, BssHII
Rictor constructs pCMV-Myc-Rictor1-1708 (full length) F:GATTGACGTCGACCGTCAATATGGCGGCGATC G SalI 5170 R:CTGGTACCATCATAAATATGAGGTCAGGATTC KpnI pCMV-Myc-Rictor1-789 F:GATTGACGTCGACCGTCAATATGGCGGCGATC G SalI 2407 R:CTGGTACCTACTGAATGAGAGCATGAAGATTG GC KpnI
pCMV-Myc-Rictor664-1708 F: TGCGTCGACTGGAACACTTTCTTGCCACCCTC SalI 3171
R: CTGGTACCATCATAAATATGAGGTCAGGATTC KpnI
SIN1β constructs
pCMV-HA-SIN1β F: CGGAATTCCGGCCTTCTTGGACA EcoRI 1461
(full length) R: GAAGATCTTCACTGCTGCCCGG BglII
pCMV-HA-SIN1β1-267 F: CCGGAATTCGGATGGCCTTCTTGGAC EcoRI 801
R: CGGGGTACCTCACTTTTCAACCAGGGC KpnI
pCMV-HA-SIN1β266-486 F: CCGGAATTCGGGAAAAGTACTCATCT EcoRI 666
R: CGGGGTACCTCACTGCTGCCCGGA KpnI
pCMV-HA-SIN1β334-486 F: CCGGAATTCGGGACATAGCCACAGTA EcoRI 462
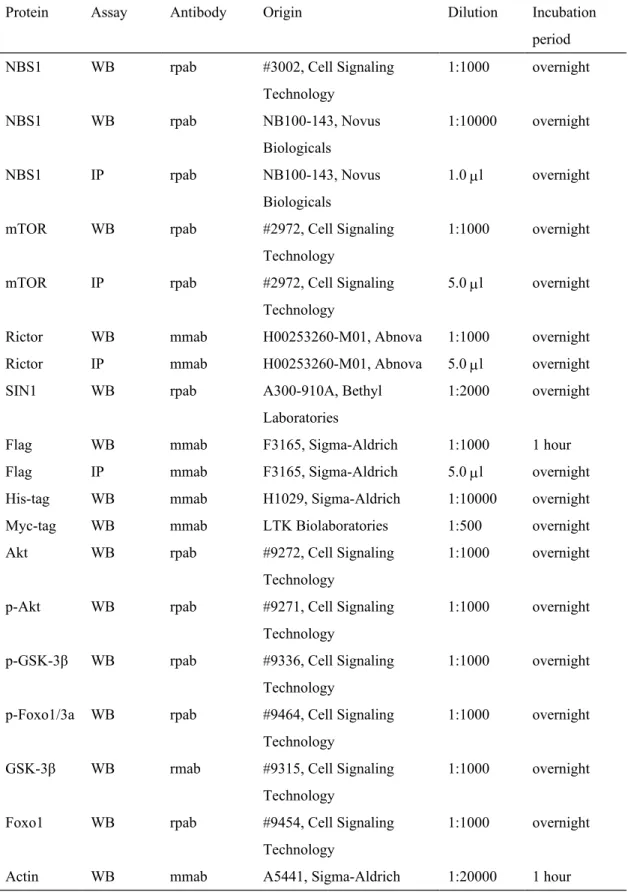
Table 2. List of proteins and characteristics of the corresponding antibodies used to detect the proteins
Protein Assay Antibody Origin Dilution Incubation period NBS1 WB rpab #3002, Cell Signaling
Technology 1:1000 overnight NBS1 WB rpab NB100-143, Novus Biologicals 1:10000 overnight NBS1 IP rpab NB100-143, Novus Biologicals 1.0 l overnight mTOR WB rpab #2972, Cell Signaling
Technology
1:1000 overnight mTOR IP rpab #2972, Cell Signaling
Technology
5.0 l overnight Rictor WB mmab H00253260-M01, Abnova 1:1000 overnight Rictor IP mmab H00253260-M01, Abnova 5.0 l overnight SIN1 WB rpab A300-910A, Bethyl
Laboratories
1:2000 overnight Flag WB mmab F3165, Sigma-Aldrich 1:1000 1 hour Flag IP mmab F3165, Sigma-Aldrich 5.0 l overnight His-tag WB mmab H1029, Sigma-Aldrich 1:10000 overnight Myc-tag WB mmab LTK Biolaboratories 1:500 overnight Akt WB rpab #9272, Cell Signaling
Technology
1:1000 overnight p-Akt WB rpab #9271, Cell Signaling
Technology
1:1000 overnight p-GSK-3β WB rpab #9336, Cell Signaling
Technology
1:1000 overnight p-Foxo1/3a WB rpab #9464, Cell Signaling
Technology
1:1000 overnight GSK-3β WB rmab #9315, Cell Signaling
Technology
1:1000 overnight Foxo1 WB rpab #9454, Cell Signaling
Technology
1:1000 overnight Actin WB mmab A5441, Sigma-Aldrich 1:20000 1 hour
Abbreviations: WB, Western blot; IP, Immunoprecipitation; mmab, mouse monoclonal antibody; rpab, rabbit polyclonal antibody