ELSEVIER
Biochimica et Biophysica Acta 1306 (1996) 133-136BB.
Biochi~ic~a
et Biophysica P~ta
Short sequence-paper
Genomic structure of carp mitogen-activated protein kinase kinase 1
gene
Jiann-Horng Leu a, Ming-Shyue Lee b, Kuan-Tien Chen c, Geen-Dong Chang b,
Chen-Kung Chou d, Chang-Jen Huang a,*
a Institute of Biological Chemistry., Academia Sinica, Taipei, Taiwan
b Graduate Institute of Biochemical Sciences, and National Taiwan Uniuersi~, Taipei, Taiwan c Department of Surgery, Universi~.' Hospital, National Taiwan University, Taipei, Taiwan
o Department of Medical Research, Veterans General Hospital, Taipei, Taiwan Received 30 November 1995; accepted 16 January 1996
Abstract
Carp mitogen-activated protein kinase kinase 1 (cMKKI) gene was isolated from a liver genomic library. The sequence around the
exon-intron boundaries and 2 kb of the promoter region were determined. Our data indicate that this gene is composed of 11 exons and 10 introns spanning about 9 kb. Multiple potential transcription initiation sites were located by primer extension analysis. Examination of 2 kb of 5'-flanking sequence revealed potential binding sites for a variety of transcription factors such as E2F, Ets-1, GATA-1, Myb, NF-IL6, Spl, and NF-kB.
Keywords: Genomic DNA sequence; Mitogen-activated protein kinase kinase; (Cyprinus carpio)
Mitogen-activated protein (MAP) kinases, also known as extracellular signal-regulated kinases (ERKs), are a family of serine/threonme kinases that mediate intra- cellular protein phosphorylation events. They have been shown to participate in a variety of signal transduction pathways and are rapidly activated in response to various extracellular stimuli [1-4]. Several subgroups of MAP kinase have been identified in vertebrates including the
ERK1 and ERK2 [5,6], c-jun amino-terminal kinase (JNK)
[7,8], p38 [9-11], and ERK5 [12].
The activation of MAP kinase is triggered by an up- stream activator, called MAP kinase kinase (MKK) or M A P K / E R K kinase (MEK) [13]. MEK is a dual speci- ficity kinase that phosphorylates MAP kinase on both tyrosine and threonine re:ddues [14,15], whereas it is acti- vated and phosphorylated by a further upstream activator on its serine/threonine residues [16]. Recently, there are also a growing number of MEKs [12,17,18]. Specific MEKs have been shown to phosphorylate specific MAP kinases in a given pathway. For example, MEK1 and MEK2 phosphorylate ERK1 and ERK2 in the cellular
* Corresponding author. Fax: + 886 2 3635038.
0 1 6 7 - 4 7 8 1 / 9 6 / $ 1 5 . 0 0 © 1996 Elsevier Science B.V. All rights reserved PII S 0 1 6 7 - 4 7 8 1 ( 9 6 ) 0 0 0 2 3 - 1
processes such as proliferation, differentiation, and devel- opment [4,13]. MEK3 and p38 are involved in the cytokine response [18,19], whereas MEK4 and JNK are shown to participate in the stress response [18]. More recently, MEK5 and ERK5 have been identified to constitute a new signal transduction pathway [12].
As an initial step to study signal transduction in fish, we have cloned a full-length cDNA encoding the carp MAP kinase kinase 1 ( c M K K I ) [20]. In this study, we further determined the genomic structure and the promoter region of this gene.
A commercially available carp liver lambda FIX II genomic library (Stratagene, La Jolla, CA, USA) was screened using the carp cDNA coding for MKK1 [20] as a probe. The probe was labeled using a DIG DNA Labeling Kit (Boehringer Mannheim, Mannheim, Germany). Two genomic clones (M1 and M2) were isolated from the
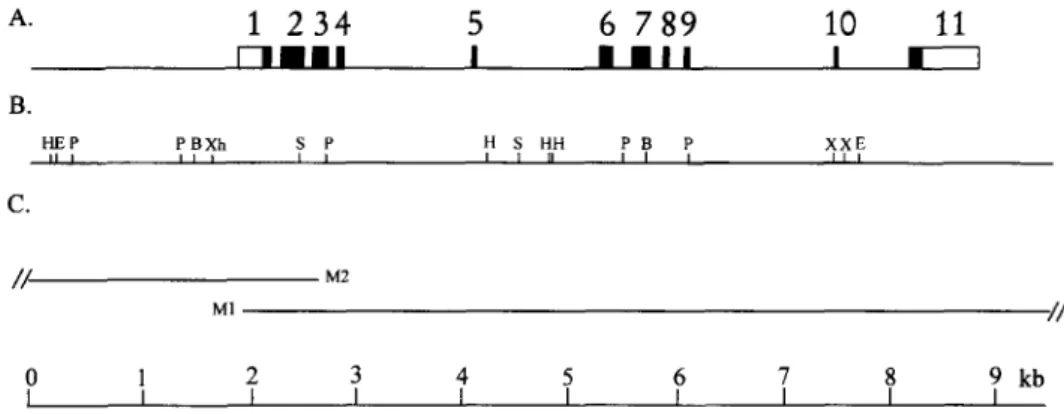
library. As shown in Fig. 1, the carp MKK1 gene spanned
about 9 kb and was composed of 11 exons and 10 introns. The sequence around the exon-intron boundaries was de- termined and shown in Table 1. All exon/intron bound-
aries identified conformed to the G T / A G splice
donor/acceptor rule [23]. Some exons were relatively small (46 bp to 78 bp), whereas the first and the last exons
134 J.-H. Leu et al. / Biochimica et Biophysica Acta 1306 (1996) 133-136 A. B. C.
1 2 34
5
6 7 89
10
11
r u n n n n I I • I I n H E P P B X h S P II I II t l I H S HH P B P X X E I I II I I I III / / M2 M1 - / / 0 1 2 3 4 5 6 7 8 9 kb I I I I I I r I I IFig. 1. Physical map of the carp MKK1 gene. (A) The structure of the gene is represented with regard to the organization of the exons and introns. Exons are indicated to scale by boxes 1-11. Solid boxes indicate the carp MKK1 coding region, whereas open boxes represent the 5'- and 3'-untranslated region. Introns and the 5'- and 3'-flanking regions regions are indicated by the solid lines. The entire gene spans about 9 kb in length and contains 11 exons. (B) The restriction map shows cleavage sites for endonucleases BgllI (B), EcoRI (E), HindlII (H), PstI (P), SacI (S), XbaI (X), and XhoI (Xh). (C) The two overlapping phage clones, M1, and M2, isolated from a Lambda FIX II carp genomic library, are shown to scale.
were large (341 bp and 612 bp). The size of introns varied considerably, ranging from 83 bp (intron 3) to 1.4 kb (intron 9). The first exon contained the Y-untranslated region and the first 28 amino acids and the last exon contained the last 36 amino acids plus the 3'-untranslated region. The other 9 exons (exon 2 to exon 10) encoded the kinase catalytic domain.
To identify sequence elements that might be involved in the transcriptional regulation of the carp MKK1 gene, the nucleotide sequence of 2 kb of the 5'-upstream region of the MKK1 gene relative to the ATG translation start codon was determined and shown in Fig. 2. Computer analysis of the sequence revealed numerous potential binding sites for
transcription factors. One T A T A box [21] and a C C A A T motif [22] was observed at the position o f nucleotides
- 7 5 3 to - 7 4 8 and - 1331 to - 1327, respectively. How- ever, these elements were located too far upstream from the transcription initiation site. Two putative binding sites for E 2 F [23] were found at the position o f nucleotides
- 607 to - 599 and - 364 to - 356. Another potential binding site for N F - I L 6 [24] was also observed at the position o f nucleotides - 1 4 4 4 to - 1 4 3 6 . It has been shown that genes containing the N F - I L 6 binding site are induced during inflammation [25]. Two proto-oncogene products Ets [26] and M y b [27] binding sites were also found at the position o f nucleotides - 1230 to - 1225, and
Table 1
Exon-intron sizes and junction sequences of the carp MKK1 gene
Exon Exon size 3'end of 5'end of Approximate 3'end of 5'end of
number (bp) the exon the intron size (bp) the intron the exon
1 341 GCA GCA TCA GA gtacgtatagagtt 84 ttccctgtctttag
A A S E
2 211 ATG GCC AGG AAG gtaagcgatcgacg 93 tgtattttccccag
M A R K
3 147 ATG GAC CAC ATG gtggggccagtggt 83 tgtatttgtcgcag
M E H M
4 78 GTT AGC ATA GCT gtaagtattatcaa 1200 ctctctttccacag
V S I A
5 52 ATG CAC AGA G gtacactcagagca 1160 aataatgcacagag
M H R D
6 125 TCG TAC ATG TCG gtgagtcacccacc ! 94 ctctgccttgacag
S Y M S
7 214 CCA GTT AGC G gtaagaacttgtta 120 ttgttttcccccag
P V S G
8 65 ATT GTC A A T GAG gtcagagaatttgc 131 tctacatcctgcag
I V N E
9 62 GTG ATG A A A TG gtgagctcctgttt 1400 gtgttttttcacag
V M K C
10 46 AAG ATG CTA ATG gtacgtacctttcc 380 atgttaatttacag
K M L M
11 612 AAAATGGATAGCTTGGTTCAAAGGCAAACATTTAATTGAGAATCAACTGGCGTTGCATCAAG
(the end of the cDNA)
G GCC AAC CTG A N L CTC ATT CAT L I H GAT GGA GGC D G G GTA CTC AGA V L R AC G'IT AAG CCC V K P CCG GAG AGA P E R
GA CAC GGA ATG H G M CCA CCA CCC P P P C CT ATG AAG L M K GGC CAC ACG G H T
J. -H. Leu et al. / Biochimica et Biophysica Acta 1306 (1996) 133-136 135
- 1289 to - 1284 and - 1676 to - 1671, respectively. A site for NF-kB [28], a ubiquitous gene activator that is induced in response to various extracellular signals, was observed at the position of nucleotides - 1851 to - 1842. NF-kB has been shown to involve the physical interaction of Spl [29] which was ~Llso found at the position of nucleotides - 1 2 7 6 to - 1 2 7 1 . In addition, a consensus GATA-1 [30] binding sequence was observed at the posi- tion of nucleotides - 1523 to - 1518.
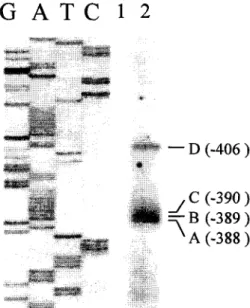
The transcription initiation site of the carp MKK1 gene was determined by primer extension of the 5' ends of the carp ovary poly (A) + RNA. Four major extended products were revealed (Fig. 3). The exact position of the extended products was determined by aligning the sequencing ladder obtained with the same primer. The bands A, B, C, and D
G A T C
1 2
__~_ ~ -- D ( - 4 0 6 ) . . / C ( - 3 9 0 ) -- B ( - 3 8 9 ) ~ - - ~ A ( - 3 8 8 ) -1980 A T C A C T G T A C T A A G C T T A C T A G T G G T A A T T T A A A A G G T G T T T T T T T T T T T T T T T T C T T C T -1920 C C A T C A T C C T T T T T T C A A C T A A T C C T G A A T T C T A A C T T G A A T A C T C A G T A C T G T A T C C T A NF-kI3 -1860 T C T G T G G C T G G G A A T T T C T C A A A A T A A G A A T A C C A C C A T G A A G G G T T A A A T T A C T T G T A A -1800 T C T T A G G T T T A T C T T T G A G T C G G C % T G G T C T T C C A T C T T C T C T T C A T G A C A G C C C A A T T T M~,b -1740 T T T C C C C C C C T T T T C T T T A T A A T T T G A A A G C G T A A A C A G T G C T G C A G T T G A C A G T T T C A C -1680 A T A A T A A G C C C T T A A A G T A G T A T G T T C C C C T A A A C T T G A C C A C A C T T T T G T G A T T T T T C A -1620 CATTTAAGGTAAGGAACTTCTTA/t%TAGACTTCCATCTTTAGACACCTAGAATGATCTAT G A T A - I -1560 TAGATGTGGATAAAATGTACATTT,3CTTACAATGAAAAGATAAATGGATAGTGATTGACG N F - I L 6 -1500 A C T A T A T A T T T C C A G C T C A G A G G T G A A G C C A G G T G A G G T T G A A G T G G G G T T T T T T T T T T G -1440 G G A A T G G A G G C A T C C C C T C T T A A T A C A G T G C T T A A G A C A G C A T G G G C A G T G G C T T G C A G T C C A A T -1380 T C T T G A T G T A A A T G T T T T G T T T G T G T C T T T T A T A T G T T C A T A T T A A A A G C C A A T A A A A A T M y b Spl -1320 A A T A A T A T T T T A T C A A G T T T T G A A r T G C A T C C A G T T G C T C C A G T G G G C G G G C A C A G A T T C E t s - i -1260 A T T G T T A T T G G C T A C T C T C A A C C C r T C T T G A G G A A A A T G T A A A T T G G T G A C C T A A A G T G G -1200 G A T T T G A C A A C A G G A G C G A A C C A G Y T G T C C C T G T C T G A A C T G A A A T C G A A C T G T A A A A G C -1140 T C T A A C A C C T G T C A A T T A T G T G T G ~ A A T T A A A C T G A T T T A A A A T A A A T G T C T T T C C A G T G -1080 A T T T T T A A A T C T T G G A T T T A A C G C C A T T G A T T G C T T A A A G A A T A T A C T T C T A T A C A C G T A -1020 T T C C T T A G A A A T G T A C A A T A A C A T ~ T T T T T T T T C C A G G A C A G C A T A C C A T T A C T A G A G T T 960 A A T T T T A C T A C A A T A A A A C T G G T T A C A A G T A C A T A A A A A G T A C T G A T G A T T T A T T T A C T T 900 T G A C A C C A A C C A C A T T A G T G C A C G ~ A G G T A T T T G C G T G G G G T A A A A A A C T A A A C A A A C A A 840 C A A A A T T A A A C C G T T T T G C T C C T T T T T G T T A A A G A T G G T A A C G A A T T A T C A C T A A G T T A C T A T A - b o x 780 C T T T A C T C A T A T G T A T A T T T T A C T G T T T A T A A A T A T A T A T A T A T A T A C G G T T A T T T G T C A 720 A C T G C A C A A C T T A T C A G G G T T T A C T G A C T G C A G A A A A T T A A T A G T A C G T A T C A T A T T A C A E 2 F 660 A T A A A T A T G A C A T G A T G T G C T A A A A T A T T T C A G A A A A T T G T A T T C T T A C G T T T T T T T G G C 600 G G C G A C T T C C T T G T G A C A G A T C T T T A A A T G T G A C C G T T A C T T C G T T C A C C G C T A G A T G C C 540 G T T C A T A T T C A A A T C A T C A C T C A T C C T T T T G T C A A A G C T C G C G A T A A C T C G A G C G T C C T G 480 G C G A C A G T T C T C G C G A G A G C T T G G G A A C G C T T C C T C T T G G T T G G A T A G C G C T T C C T G G C C E 2 P 360 C G C G G G T C T C C G G G C C A A A C A A T T T C G G A C A G C A T C T C G C G G T C G G T C C T G T G G C T G C G A V 300 A C G A C A G G C C T G A C C C G T G T T T T G T C T G A G G G A G C G C T C T C G A G T C G G A G G C C G T C T C G A 240 A G A G A C A T C C A C G G A T G A A C G C G C T G A A A T A A A C C G C A T T T C T C G C A C G T T T G C A C T C G C 180 T C T T T C C A C C T G A C A C C T A A C T A T T A T T T G G T A C A G C G T C T G G A G G C A T G A C G G G G T G T T 120 G A A A T T A G G G G C T T A C G A A T C G C A G A A A C T T C T A T A G A C A C A A G T G T A G C A A T T T G A A G G 60 ATTGAATGTCAGCAGAGTCAACC~GATTGTTCTCTTTGAAATAGATAACAAGCAACTAT___GGFig. 2. Nucleotide sequence of the 5'-flanking region of the carp MKK1 gene. The nucleotide number ( - - 1) stag at the G of the putative A T G
codon [20]. Potential binding sites for a variety of transcription factors are underlined. The 28-mer antisense oligonucleotide used for primer exten- sion analysis is also underlined. Candidate transcription stag sites by primer extension (see Fig. 3) are indicated by bent arrows. The stag of the carp MKK1 c D N A clone M K K 3 0 [20] is marked by open triangle at position - 262.
Fig. 3. Determination of the transcription initiation site of the carp MKK1
gene. [32P]-labeled primer (see Fig. 2) was annealed to 10 /.zg yeast
t R N A (lane 1, as a negative control) or 5 /xg poly (A) + R N A from carp ovary (lane 2) and extended with reverse transcriptase. The sequencing ladder of the carp MKKI gene labeled G, A, T and C was obtained by using the same primer and electrophoresed on the same gel. The extended products A - D and their positions relative to the initiator ATG codon are indicated.
corresponded to the sites at - 3 8 8 , - 3 8 9 , - 3 9 0 , and - 4 0 6 , respectively, relative to the initiator methionine codon (Fig. 2).
The significance of the presence of some transcription factor binding motifs in the putative promoter region are discussed as follows. The transcription factor E2F plays an important role in regulating genes involved in cell cycle control. E2F binding sites are found in the promoter region of human cdc2 gene [31], human cyclin D1 gene [32], and human cyclin A gene [33]. In this study, we demonstrated that E2F binding sites are present in the 5' flanking region of the carp MKK1 gene. The involvement of E2F regula- tory elements in regulating carp MKK1 promoter activity is currently being investigated.
GATA-1 is an abundant protein in cells of the erythroid lineage and its binding sites are found in the promoters of many hematopoietic-specific genes [30]. Recent evidence reveals that GATA-1 can activate transcription in a syner- gistic manner with Sp 1. In addition, the spacing of GATA-1 and Spl elements markedly influences synergistic activa- tion of transcription. When both binding sites are well separated (about 300 bp) and GATA-1 and Spl are coex- pressed, maximal transcription is achieved [34]. It is inter- esting to notice that there are also two potential binding sites for GATA-1 and Spl (separated about 250 bp) in the 5' upstream region of the carp MKK1 gene. Further inves- tigation is required to test whether such a superactivation of GATA-1 with Spl may occur in the fish cell line.
The Ets superfamily is a structural class of trans-acting phosphoproteins which have important roles in the control
136 J. -H. Leu et al. / Biochimica et Biophysica Acta 1306 (1996) 133-136
of growth and development [26]. c-Ets-1 has a consensus MAP kinase phosphorylation site (PLL 82 TP) and its tran- scriptional activity might be modulated by the R a s / M A P K pathway [35]. In fact, the activation of M K K / M E K leads to dual phosphorylation of MAP kinase and this activated MAP kinase in turn may modulate the activity of Ets- 1. At present, the significance of potential Ets-1 binding site in the carp MKK1 gene transcription needs further investiga- tion.
In conclusion, we have isolated and characterized the
carp MKK1 gene. This gene is composed of 11 exons and
10 introns spanning about 9 kb. In addition, the putative promoter region of the carp MKK1 gene has several potential binding sites for transcription factors which are implicated in the control of cell cycle and proliferation. To our knowledge, this is the first report to characterize the genomic structure of the M K K / M E K gene.
This research was supported by grants from the Na- tional Science Council and from the Health Administra- tion, Taiwan, Republic of China.
References
[1] Sturgill, T.W. and Wu, J. (1991) Biochim. Biophys. Acta 1092, 350-357.
[2] Pelech, S.L. and Sanghera, J.S. (1992) Trends Biochem. Sci. 17, 233-238.
[3] Davis, R.J. (1994) Trends Biochem. Sci. 19, 470-473. [4] Marshall, C.J. (1995) Cell 80, 179-185.
[5] Boulton, T.G., Nye, S.H., Robbins, D.J., Ip, Y.N., Radziejewska, E., Morgenbesser, S., Depinho, R., Panayotatos, N., Cobb, M.H. and Yancopoulos, G.D. (1991) Cell 65, 663-675.
[6] Sturgill, T.W. and Ray, L.B. (1986) Biochem. Biophys. Res. Com- mun. 134, 565-571.
[7] Derijard, B., Hibi, M., Wu, I.-H., Barrett, T., Su, B., Deng, T., Karin, M. and Davis, R.J. (1994) Cell 76, 1025-1037.
[8] Kyriakis, J.M., Banerjee, P., Nikolakaki, E., Dai, T., Rubie, E.A., Ahmad, M.F., Avrnch, J. and Woodgett, J.R. (1994) Nature 369, 156-160.
[9] Galcheva-Gargova, Z., Derijard, B., Wu, I.-H. and Davis, R.J. (1994) Science 265, 806-808.
[10] Han, J., Lee, J.-D., Bibbs, L. and Ulevitch, R.J. (1994) Science 265, 808-811.
[11] Rouse, J., Cohen, P., Trigon, S., Morange, M., Alonso-Llamazares, A., Zamanillo, D., Hunt, T. and Nebreda, A.R. (1994) Cell 78, 1027-1037.
[12] Zhou, G., Bao, Z.Q. and Dixon, J.E. (1995) J. Biol. Chem. 270, 12665-12669.
[13] Seger, R. and Krebs, E.G. (1995) FASEB J. 9, 726-735.
[14] Nakielny, S., Cohen, P., Wu, J. and Sturgill, T.W. (1992) EMBO J. 11, 2123-2129.
[15] Matsuda, S., Kosako, H., Takenaka, K., Moriyama, K., Sakai, H., Akiyama, T., Gotoh, Y. and Nishida, E. (1992) EMBO J. 11, 973-982.
[16] Lange-Carter, C.A., Pleiman, C.M., Gardner, A.M., Blumer, K.J. and Johnson, G.L. (1993) Science 260, 315-319.
[17] Sanchez, I., Hughes, R.T., Mayer, B.J., Yee, K., Woodgett, J.R., Avruch, J., Kyriakis, J.M. and Zon, L.I. (1994) Nature 372, 794-798. [18] Derijard, B., Raingeaud, J., Barrett, T., Wu, I.-H., Han, J., Ulevitch,
R.J. and Davis, R.J. (1995) Science 267, 682-685.
[19] Lee, J.C., Laydon, J.T., McDonnell, P.C., Gallagher, T.F., Kumar, S., Green, D., McNulty, D., Blumenthal, M.J., Heys, J.R., Landvat- ter, S.W., Strickler, J.E., McLaughlin, M.M., Siemens, I.R., Fisher, S.M., Livi, G.P., White, J.R., Adams, J.L. and Young, P.R. (1994) Nature 372, 739-746.
[20] Huang, C.J., Lee, M.S., Chang, G.D., Huang, F.L. and Lo, T.B. (1994) Biochim. Biophys. Acta 1220. 223-225.
[21] Breathnach, R. and Chambon, P. (1981) Annu. Rev. Biochem. 50, 349-383.
[22] McKnight, S.L. and Kingsbury, R. (1982) Science 217, 321-324. [23] Mudryj, M., Hiebert, S.W. and Nevins, J.R. (1990) EMBO J. 9,
2179-2184.
[24] Akira, S., Isshiki, H., Sugita, T., Tanabe, O., Kinoshita, S., Nishio, Y., Nakajima, T., Hirano, T. and Kishimoto, T. (1990) EMBO J. 9, 1897-1906.
[25] Akira, S., Nishio, M., Wanf, X., Wei, S., Matsusaka, T., Yoshida, K., Sudo, T., Naruto, M. and Kishimoto, T. (1994) Cell 77, 63-71. [26] Wasylyk, B., Hahn, S.L. and Giovane, A. (1993) Eur. J. Biochem.
211, 7-18.
[27] Faisst, S. and Meyer, S. (1992) Nucleic Acids Res. 20, 3-26. [28] Grilli, M., Chiu, J.J.S. and Lenardo, M.J. (1993) Int. Rev. Cytol.
143, 1-62.
[29] Jackson, S.P., MacDonald, J.J., Lees-Miller, S. and Tjian, R. (1990) Cell 63, 155-165.
[30] Orkin, S.H. (1992) Blood 80, 575-581. [31] Dalton, S. (1992)EMBL J. 11, 1797-1804.
[32] Motokura, T. and Arnold, A. (1993) Genes Chromosomes Cancer 7, 89-95.
[33] Henglein, B., Chenivesse, X., Wang, J., Eick, D. and Brechot, C. (1994) Proc. Natl. Acad. Sci. USA 91, 5490-5494.
[34] Merika, M. and Orkin, S.H. (1995) Mol. Cell. Biol. 15, 2437-2447. [35] Brunner, D., Ducker, K., Oellers, N., Hafen, E., Scholz, H. and