台灣山櫻花種子於休眠至發芽階段種子內亞硫酸基水解酵素活性的變化
68
0
0
全文
(2) 台灣山櫻花種子於休眠至發芽階段 種子內亞硫酸基水解酵素活性的變化 The Dramatic Changes of Sulfatase Activities in Prunus campanulata Maxim. Seeds from Dormancy to Germination. 研 究 生:邱靜玟. Student:Ching-Wen Chiu. 指導教授:楊裕雄 博士. Advisor:Dr. Yuh-Shyong Yang. 國立交通大學 生物科技系 碩 士 論 文. A Thesis Submitted to Department of Biological Science and Technology College of Biological Science and Technology National Chiao Tung University in partial Fulfillment of the Requirements for the Degree of Master of Science in Biological Science and Technology July 2006 Hsinchu, Taiwan, Republic of China. 中華民國九十五年七月.
(3) 台灣山櫻花種子於休眠至發芽階段 種子內亞硫酸基水解酵素活性的變化 學生:邱靜玟. 指導教授:楊裕雄 博士 國立交通大學生物科技學系碩士班. 摘. 要. 種子休眠是普遍存在許多植物種子的生理現象,種子可以避開不適合發 芽的環境或是逆境,增加植株發芽散佈的優勢;但種子休眠的生化、生理 機制至今仍不清楚。台灣山櫻花種子具有深度休眠性,種子利用暖、低溫 混合層積處理之後可以解除休眠;但山櫻花種子僅經過暖溫層積處理後可 能逐漸成熟,但播種在適合的環境之下仍不能發芽。山櫻花種子經過暖溫 或冷溫層積處理後,種子內亞硫酸基水解酵素的活性有很顯著的改變。本 文說明亞硫酸基水解酵素的活性變化可能對於山櫻花種子的發育及從休眠 至解除休眠、發芽過程中扮演重要的調控功能。. i.
(4) The Dramatic Changes of Sulfatase Activities in Prunus campanulata Maxim. Seeds from Dormancy to Germination. Student:Ching-Wen Chiu. Advisors:Dr. Yuh-Shyong Yang. Department of Biological Science and Technology, College of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan. ABSTRACT Seed dormancy is an ecological adaptation for many plant species to avoid the stress and unfavorable environment of germination. However, there have been very few molecular or biochemical studies about the detail of seed dormancy. Prunus campanulata Maxim. (Taiwan Cherry) seeds are deeply dormant. Warm followed by cold stratification, which contains imbibition and warm following cold treatment, can release seeds from dormancy to germination. P. campanulata seeds become mature during warm stratification but will not germinate even in optimal condition. Dramatic changes of the sulfatase activities were observed following warm and cold stratification of the P. campanulata seeds. Desulfation might be important for the regulation of cherry seeds from dormancy to germination.. This report explored. desulfation and its effects on dormancy and development of P. campanulata seeds.. ii.
(5) Acknowledgement 光陰荏苒,即將離開生活了七年的交通大學。這段時光不但充實,也讓我獲 得許多有形及無形的收穫。 最感謝的,莫非於 楊裕雄教授當年願意收留我這個什麼都不懂的小毛頭, 讓我有機會接觸到有趣的生化領域,並授予我很多重要的觀念及態度。也非常感 謝林業試驗所 簡慶德博士在研究上對我的指導與勉勵,才能讓我順利的完成論 文。感謝口試委員 呂平江教授和 林彩雲教授對於此論文的指導,使學生在研究 的方向及態度上更臻成熟。 謝謝陳舜英小姐及簡博士實驗室的成員們在實驗上幫了非常多的忙,讓此論 文內容可以更加充實及完備。 謝謝LEPE這個溫暖的大家庭陪伴著我成長:莊博、恩仕學長、小木學長、 晟洋學長、Rich學長、郁吟學姐、大樹、阿棚學長、政哲學長、小米學姊、大晃 學長、惠菁學姊、淵仁學長、秉鈞學長、怡宏學長、阿斌、陳蘋果、小羿、僕僕 哥、蜜柑、程允、Peggy、淑婷、陸宜、小志、小樹、將平、韋汝、小丸、美春、 嘉蔚、瓊欣、曉薇…,如果有機會再選擇一次,我仍然會選擇LEPE。 感謝有非常多的好朋友,一直在我身邊給我鼓勵;感謝非常體諒我的家人 們、有你們的支持,讓我無後顧之憂的在外求學;謝謝何小何讓我知道我有堅強 跟力量可以完成很多事情;要感謝的人實在太多,衷心的感謝所有幫助過我的人。 謝謝天,讓我有機會在這個世界上活著。 I love you all. Leslie 來 2006.7 新竹. iii.
(6) CONTENTS. PAGE. Abstract (Chinese) …………………………………………………………… i Abstract (English) …………….…..……………………………………….… ii Acknowledgement ………….……..………………………………………… iii Contents …………………………………………………………………...… iv Contents of Figures and Tables ……...…………….………………...……… vii Abbreviations and Symbols ……..………………….……………………..… ix. The dramatic changes of sulfatase activities in Prunus campanulata Maxim. seeds from dormancy to germination. Chapter 1. Introduction ………………………………………………..…… 1. 1.1 Seed dormancy 1.2 Seed dormancy of cherry Prunus campanulata Maxim. 1.3 Biochemistry about seed dormancy 1.4 Biological functions of sulfation and desulfation in plants. Chapter 2. Materials and Methods………………………..………………… 11 iv.
(7) 2.1 Materials 2.2 Seed harvesting, the warm and/or cold stratification, and germination test of P. campanulata seeds 2.3 Extract and assay of plant sulfatases in P. campanulata seeds. Chapter 3. Result…………………………………………………..…..…… 14. 3.1 Germination test of P. campanulata seeds 3.2 The activity of sulfatases during the progress of dormancy breaking in of P. campanulata seeds. Chapter 4. Discussion………………………………………………....…… 18. 4.1 Germination of P. campanulata seeds 4.2 The activity of sulfatases/ST during the progress of dormancy breaking in P. campanulata seeds 4.3 Influence of endocarp and tests on P. campanulata seed dormancy 4.4 Dormancy releasing of P. campanulata seeds. Chapter 5. Conclusion………………………………………………....…… 25. v.
(8) Acknowledgement …….……………………………………..………....…… 26 References …………………..…………………………………………..…… 44 Appendix ………………………..…..…………………………………..…… 53 Autography ……...…………..…………………………………………..…… 56. vi.
(9) CONTENTS OF FIGURES AND TABLES. Table I. PAGE. Germination percentage of P. campanulata seeds following warm or/and cold stratification.……...………….………………..……… 27. Table II. Dormancy categories.…………………………………………....… 29. Figure 1 Seed dormancy is controlled by environmental factors and seed physiology………………………………………………………… 2. Figure 2 The life history of Prunus campanulata Maxim. in nature………....… 4 Figure 3 Flowers, fruits, and seeds of cherry Prunus campanulata Maxim.….… 4 Figure 4 The biological function of sulfotransferases………...…………….…..… 8 Figure 5 The biological function of sulfatases……………………………....…9 Figure 6 The reaction of enzyme assay with fluorescence photometry.…......… 30 Figure 7 The overview of experimental flow chart.…..…………………....…. 31 Figure 8 Germination percentages of P. campanulata seed following cold stratification in three different year.……..……………………..…... 32. Figure 9 Longitudinal section through a seed of P. campanulata.................….. 33 Figure 10 Sulfatase activity was detected in endocarp, testa , and embryo......… 34 Figure 11 The activities of sulfatase in P. campanulata seed extracts during the. vii.
(10) warm and/or cold stratification in 2004.……………………......… 35. Figure 12 The activities of sulfatase in P. campanulata desalted seed extracts during the warm and/or cold stratification in 2004……...............… 36. Figure 13 The tendency of sulfatase activities in P. campanulata seed extracts during the warm stratification…….............................................… 37. Figure 14 The tendency of sulfatase activities in P. campanulata seed extracts during the cold stratification…...…............................................… 38. Figure 15 P. campanulata sulfatases activity and percentage of germination during warm plus cold stratification.……………………… .....… 39. Figure 16 The activities of phenol sulfotransferase (Rat ST1A1) were inhibited by extract from P. campanulata seeds…………………………….… 40. Figure 17 Removal of testa and endocarp readily remove seed dormancy of P. campanulata ……………………………………………………………..... 41. Figure 18 The relation between water uptake and seed physiology…………..… 42 Figure 19 The hypothesis of desulfation regulation in seed dormancy and germination ……………………………………………………….…. 43. viii.
(11) ABBREVIATIONS AND SYMBOLS. ε. Absorption (extinction) coefficient. A280. Absorption at 280nm. A760. Absorption at 760nm. AMP. Adenosine -5’-monophosphate. C1. Seeds treated with 1 week cold stratification. DTT. Dithiothreitol, Cleland’s Reagent. EDTA. (Ethylenedinitrilo)tetraacetic acid. Ki. Inhibitory constant. Km. Michaelis constant. MU. 4-methylumbelliferone. MUS. 4-methylumbelliferyl sulfate. P. campanulata. Prunus campanulata Maxim.. PAP. Adenosine 3’,5’-bisphosphate 3’-phosphoadenosine 5’-phosphate. PAPS. Adenosine 3’-phosphate 5’-phosphosulfate 3’-phosphoadenosine 5’-phosphosulfate. PMSF. Phenylmethylsulfonyl fluoride. PST. Phenol sulfotransferase. SDS. Sodium Dodecyl Sulfate. TEMED. N,N,N’,N’,-tetramethylethylene diamine. Tris. 2-Amino-2-(hydroxymethyl)-1,3-propanediol. Vmax. Maximum velocity. W1. Seeds treated with 1 week warm stratification ix.
(12) ABBREVIATIONS AND SYMBOLS (continued) W1C4. Seeds treated with 1 week warm then 4 weeks cold stratification. W1C6. Seeds treated with 1 week warm then 6 weeks cold stratification. W6C4. Seeds treated with 6 weeks warm then 4 weeks cold stratification. W6C6. Seeds treated with 6 weeks warm then 6 weeks cold stratification. W6C8. Seeds treated with 6 weeks warm then 8 weeks cold stratification. x.
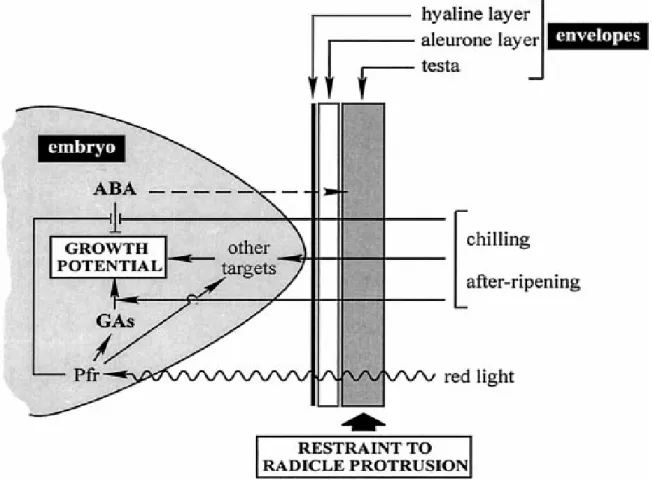
(13) Chapter 1. Introduction. 1.1 Seed dormancy Seed dormancy is defined as the failure of an intact viable seed to complete germination under favorable conditions (Bewley, 1997) as an ecological adaptation (Geneve, 2003).. The role of seed dormancy is across all species to spread. germination across time but in synchrony with seasonal cycle to avoid unfavorable weather to maximize competitive advantage and ensure the establishment of plants (Baskin and Baskin, 1998). Environmental stimuli can trigger dormancy release. Seed moisture content, light, and temperature are arguably the most important on primary dormancy and considerations for conducting and interrupting seed dormancy research (Geneve, 2003). Seed dormancy also depends on seed structures, especially those surrounding the embryo, and on factors, including several plant hormones, affecting the growth potential of the embryo (Koornneef et al., 2002) (Figure 1). Studies with model species that have comparatively shallow dormancy have identified a number of genes involved in the regulation of dormancy and germination (Bewley, 1997; Koornneef et al., 2002). However, there have been very few molecular studies in more deeply dormant species in which moist seeds require prolonged exposure to low temperature to release dormancy and induce germination (Stephen et al., 2003).. 1.
(14) There is little is known about the mechanism in detail of seed dormancy and germination (Bewley, 1997).. Figure 1 Seed dormancy is controlled by environmental factors and seed physiology (Debeaujon and Koornneef, 2000).. 1.2 Seed dormancy of cherry Prunus campanulata Maxim. Deeply dormant Taiwan cherry (Prunus campanulata Maxim.) seeds are used to study the seed dormancy.. P. campanulata seeds are known for their. morphological and physiological dormancy which can be broken by a combination of. 2.
(15) warm and cold stratification, however some ungerminated seeds were found when seeds were stratified at 5 ℃ alone (Chien et al., 2002). P. campanulata distributed from Japan’s Ryukyu Islands and Taiwan to southern China, is a deciduous and outstanding flowering tree with dense branching (Hiroyoshi, 1993). The trees grow on mountains at elevations of between 400 and 2,500 m. P. campanulata produces red and campalunate flowers from January to March and dark red fruits from April to May (Figure 2), make it a valuable ornamental plant. The mature fruits of P. campanulata exhibit a dark red or dark purple color (Chien et al., 1996; Chien et al., 2002) (Figure 3). The dormancy mechanism in Prunus, like those of other species of deeply dormant seeds, appears to be complex. Prunus seeds have embryo dormancy and require a period of after-ripening in the presence of moisture and oxygen to overcome it (Grisez et al., 2000).. The embryo is dormant and the surrounding structures. (endosperm, seed coat and endocarp) also impose some control (coat-enhanced dormancy). Embryo dormancy is deemed to be present when the embryo fails to grow after the removal of the tissue that encloses it (Stephen et al., 2003). In the other hand, because of their stony endocarps, Prunus seeds often been thought to have seedcoat dormancy.. The surrounding hard endocarp of embryo appeared to a. weakening to permit radicle protrusion after stratification in P. campanulata (Chien et. 3.
(16) al., 2006), but it is permeable to water.. Prunus is not thought truly hard-seeded. (Grisez et al., 2000).. Figure 2 The life history of Prunus campanulata Maxim. in nature.. Figure 3 Flowers, fruits, and seeds of cherry Prunus campanulata Maxim.. 4.
(17) Development, maturity and inhibition of germination may be involved in seed dormancy.. Lee et al. (2006) exposed that significant protein changes between. dormant and dormancy-broken seed of P. campanulata which were observed by 2D gel electrophoresis. There are some biochemical reactions activated to maintain or break seed dormancy.. 1.3 Biochemistry about seed dormancy The direct enzymetic effect or mechanism about seed dormancy remains to be answered. Little is known regarding their roles or interactions with other observed changes although many enzymes have been studied with respect to dormancy. Lang (1994) reviewed seed dormancy investigation of enzymology in seed physiology. Membrane lipid biosynthetic enzymes were thought to be prime targets for low-temperature regulation because biochemical and physical membrane restructuring usually occurs with exposure to low temperature.. Seeds are particularly rich in. proteases used to hydrolyze storage proteins during the last stages of germination and during embryonic development (Gegenheimer, 1990). Some biomolecules about seed dormancy, germination and plant development are reported.. Debeaujon et al. (2000) indicated that most of the pigmentation. mutants exhibited a reduced dormancy compared with corresponding wild-type of. 5.
(18) Arabidopsis. Dormancy of cereal caryopses might be at least partially controlled by the high level of free phenolic acid (Weidner and Paprocka, 1997). Some flavonoid conjugates, including the sulfate esters, may play a role in the regulation of plant growth by blocking the quercetin-stimulated accumulation of the auxin phytohormone (Faulkner and Rubery, 1992).. The sulfated peptide phytosulfokine (PSK,. Tyr(SO3H)-Ile-Tyr(SO3H)- Thr-Gln) was an intercellular signal that plays a key role in cellular differentiation and proliferation in plants (Matsubayashi et al., 2002). Desulfated PSK lost both mitogenic activity and competitive ability for ligand binding (Matsubayashi and Sakagamin, 1999). The activity of tyrosylprotein sulfotransferase (TPST) involved in PSK biosynthesis of higher plant was detected and an in vitro assay system of TPST was established (Hanai et al., 2000).. 1.4 Biological functions of sulfation and desulfation in plants Some biomolecules are activated by sulfation or desulfation in plants, sulfation and desulfation may play an important role in seed dormancy, germination and plant development. It was thought that sulfation and desulfation may play the part of the intermolecular recognition and signaling processes in plant growth, development, and adaptation to stress (Varin et al., 1997). The sulfation (also known as sulfonation and sulfuryation) of biomolecules have long been known to take place. 6.
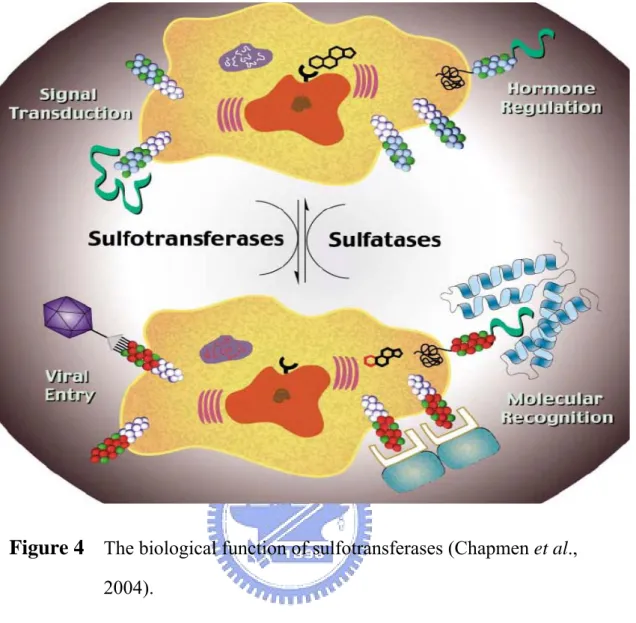
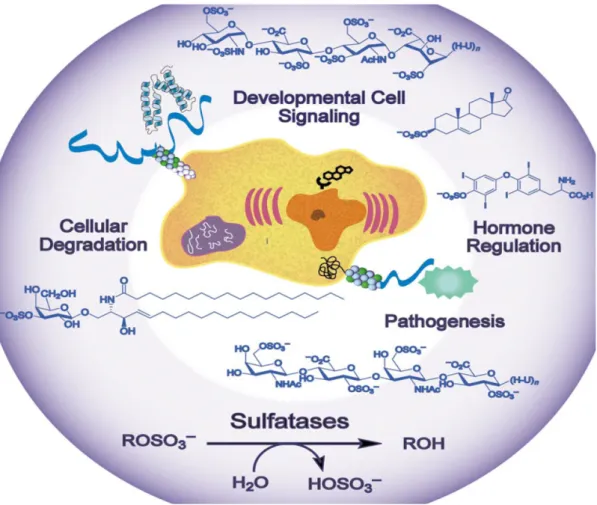
(19) in a variety of organisms, and new biological functions continue to be uncovered in connection with these important transformation (Chapmen et al., 2004). Sulfonate conjugation not only facilitates transport and excretion of hydrophobic molecules by increasing their water solubility, it abolishes the biological activity of hormones such as estrogens. The sulfonation reaction also can lead to the production of biologically active molecules (Gidda et al., 2003). Sulfotransferases (EC 2.8.2.*), the enzymes that catalyze sulfation, are involved in detoxification, hormone regulation, drug metabolism, molecular recognition, and biochemical signal pathway, and implicated in many pathophysiological processes (Chapmen et al., 2004, Figure 4). Sulfatases (EC 3.1.5.*) cleave sulfate esters in biological systems. Sulfatases are underlined by the presence of several genetic disorders characterized by the abnormal accumulation of intermediate sulfate compounds in human (Parenti et al., 1997). Sulfate conjugation constitutes an important reaction in the transformation of xenobiotics and in the modulation of the biological activity of steroid hormones and neurotransmitter in mammals (Varin et al., 1997).. Because they have been. implicated in the outset of hormone-dependent cancers, lysosomal storage disorders, developmental abnormalities, and bacterial pathogenesis, sulfatases were considered to be targets for therapeutic endeavors (Figure 5).. 7.
(20) Figure 4 The biological function of sulfotransferases (Chapmen et al., 2004).. Desulfation activity in plants has seldom been reported in the literatures. Arylsufatases in the unicellular green alga Chlamydomonas reinhardtii (de Hostos et al., 1988; Yildiz et al., 1994) and the multicellular green alga Volvox carteri (Hallmann and Sumper, 1994) were produced in response to sulfate deprivation in environment and to develop the capacity to transport sulfate more rapidly. Baum and Dodgson (1957) detected the arylsulfatase activity in the seeds of Sinapis niger (black mustard), but no further investigation for the biochemical properties and biological 8.
(21) function of arylsulfatase from Sinapis niger. Roots of sterile –grown crops contained arylsulfatase activity (Knauff et al., 2003). It was quantified within the rhizosphere and roots of different crop species. It was suggested that arylsulfatase contributed to the mineralization of organic sulfur to sulfate for plant uptake, and arylsulfatase activity was also induced under sulfate deficiency condition. They also assumed that the enzyme activity is derived from endophytic bacteria not by higher plants, or an alternative possibility is the existence in both plant and bacteria arylsulfatase.. Figure 5 The biological function of sulfatases (Hanson et al., 2004).. 9.
(22) This is the first research about desulfation in higher plant seed dormancy, and some biomolecules were activated by sulfation or desulfation affecting seed dormancy, germination, or seedling development of plants.. Using fluorescence. photometry assay, we investigated the variation of sulfatase activity in P. campanulata seeds following warm and cold stratification.. We also present a. hypothesis about the regulation of sulfation/desulfation in seeds from development to dormancy in order to explain the mechanism of seed dormancy and related events about sulfation/desulfation in seeds.. 10.
(23) Chapter 2. Materials & Methods. 2.1 Materials The RC DC protein assay kit was obtained from BioRad (Richmond, CA, USA). All the other chemicals were the highest purity grade, purchased from Sigma (St. Louis, MO), and the details were listed in Appendix B.. 2.2 Seed harvesting, the warm and/or cold stratification, and germination test of P. campanulata seeds Mature fruits of P. campanulata were harvested in early May every year. Fruits in 2004 were from Ali Mt. (23°32’N, 120°47’E) at an elevation of 2000 m, Chiayi County, central Taiwan. Fruits in 2005 were from Coaling Shihbi park in Yunlin County. Immature fruits of P. campanulata were harvested in the interval from March to early May in 2006, from Hsin-hsien nursery, Taipei county. The seeds were extracted by removing the pulp in water, and filled, sunken seeds were used for subsequent treatment.. Fresh P. campanulata seeds were mixed with moist. sphagnum in a polyethylene bag (0.04 mm in thickness) and warm-stratified at alternating temperatures of 30/20 ℃ with 12 hours of fluorescent light (80-100μE/m2/s), and cold-stratified at temperatures of 4℃ in dark. Excess water in. 11.
(24) the sphagnum was removed to obtain a water content of about 400% by mass (dry weight basis). For warm plus cold treatment, fresh seeds were stratified at 30/20 ℃ for a period followed by cold stratification. Germination test was performed at the same temperature as that of the warm stratifying temperature of 20/30 ℃ with 12 hours daily photoperiod. For each test, 3 replicates of 50 seeds each per treatment were used. Germination, judged by radicle protrusion of at least 5 mm, was recorded weekly. Results were expressed as the percentage germination (%). Fresh and treated seeds were lyophilized and stored at -80℃ for enzyme assay.. 2.3 Extract and assay of plant sulfatases in P. campanulata seeds The endocarp, testa or embryo of P. campanulata seeds was homogenized with PRO 250 homogenizer (PRO Scientific, Oxford, CT, USA) contained 125 mM sucrose, 1 mM dithiotheitol, 1 mM EDTA , 10%(v/v) glycerol, 1 mM PMSF, and 10 mM Tris-HCl at pH 7.4. The suspension was centrifuged for 30 minute at 22,000 g at 4℃. twice.. The. final. supernatant. liquid. was. used. in. assaying. of. sulfatase/sulfotransferase activity with fluorescence spectrometry as describe below. The sulfatase assaying reaction mixture (1000 μl) contained 100 mM. 12.
(25) Tris-base (pH 7.0) or 100 mM phosphate buffer (pH 7.0), 5 mM 2-mercaptoetnol, 500 μM MUS, and P. campanulata seed extract, incubated in 25 ℃. The difference of fluorescence 450 nm emission excited at 350 nm was detected to calculate the activity of sulfatases from P. campanulata seed extract. The sulfotransferase assay reaction condition is the same plus 10 μM PAP and 5 μM 2-naphthol (Figure 6). Protein concentrations of P. campanulata seed extract were estimated by RC DC protein assay kit from BioRad. The small molecules in seed extract were removed by passing extract through the HiTrap desalting column.. Desalted seed extract was concentrated by. ultrafiltering with Amicom Ultra-15 centrifugal filter devices PL-30(30000 nominal molecular weight limit).. Desalted seed extract was also used in assaying of. sulfatase/sulfotransferase activity. The experimental flow chart was shown in Figure 7.. 13.
(26) Chapter 3. Results. 3.1 Germination test of P. campanulata seeds Table 1 is germination percentage of P. campanulata freshly harvested seeds or seeds treated by warm or/and cold stratification in three years.. The period of every. germination test was 12 weeks. Untreated freshly harvested seeds germinated less than 1% each year. Seeds treated with warm stratification also germinated poorly. For example, seeds treated with warm stratification for 6 weeks germinated 0.7% (2003), 6.0 % (2004) and 0.7(2005) in three different year. Therefore, warm stratification did not affect germination of P. campanulata seeds directly. Seeds treated with the same cold stratification germinated incoherently. For example, seeds treated with cold stratification longer than 8 weeks germinated 89.3% (8 weeks, 2003), 12.0%(8 weeks, 2004) and 68.0%(9 weeks, 2005) in three different year. However, the longer cold stratification, the more germination of P. campanulata seeds in the same year (Figure 8). The mixed (warm plus cold) stratification was a better strategy of breaking dormancy of P. campanulata seeds. Seeds treated with 6 weeks warm and 8 weeks cold stratification germinated completely.. Seeds treated with cold stratification 8. weeks resulted only 12% germination, but seeds treated with 1 week warm and 8. 14.
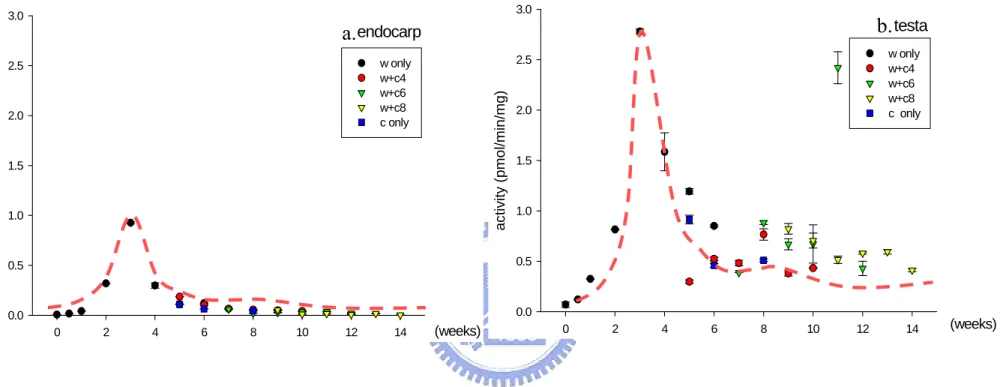
(27) weeks cold stratification germinated completely in 2004. Seeds treated with W1C4 resulted 57.3 % germination, and the germination of seeds treated with W1C6 increased to 94.0%. Familiarly, mixed stratified seeds treated with the same warm but longer cold stratification resulted higher germination percentage. For example, seeds treated with W6C4, W6C6 and W6C8 resulted germination to 52.7%, 72.0%, and 98.0% in 2004.. 3.2 The activity of sulfatases during the progress of dormancy breaking in of P. campanulata seeds The freshly-harvested or treated seeds were separated into endocarp, testa (including seed coat and endosperm), and embryo (including cotyledons, epicotyl, hypocotyl and radicle) (Figure 9), and sulfatase activity was assayed. Sulfatase activity was detected in endocarp and testa of treated seeds, but not in embryo (Figure 10). Sulfatase activity was not detected in freshly-harvested P. campanulata seeds. Different sulfatase activities were recorded in P. campanulata seeds that were stratified differently and harvested in different years. After the process of warm and/or cold stratification In P. campanulata seeds harvested in 2004 and assays of sulfatases, tendency of change of sulfatases activity in P. campanulata seeds was obtained. The sulfatase activity was detected in testa and. 15.
(28) endocarp shown in Figure 11 and Figure 12. There is less sulfatase activity assayed in freshly harvested P. campanulata seeds.. The freshly-harvested dry P.. campanulata seeds were imbibed and treated by changing temperature in 1 to 3 weeks, and then the activity of sulfatases began to be induced. The higher sulfatase activity exhibited in only warm-stratified seeds. Seeds treated with warm stratification for exact 3 weeks showed a maximum sulfatases activity in testa and endocarp. The sulfatase acitvity was reduced gradually and stable in testa and endocarp following the period of stratification. Figure 13 showed the sulfatase activities in seeds treated with warm stratification only in different years, 2002, 2003, and 2004. The maximum sulfatase activities were exposed in the first to third weeks during warm stratification. Figure 14 showed the sulfatase activities in seeds treated with cold stratification only in different years, 2004 and 2005. The sulfatase activities were induced following the period during cold stratification, but the sulfatase activities of cold-treated seeds were less than activities of warm-treated seeds. According to Figure 11, 12, 13 and 14, the sulfatase activity in testa were higher than that activity in endocarp per milligram of tissue at the same condition of stratification, but the higher total activity was assayed in endocarp of whole tissue generally.. When seed dormancy was. released by 6 weeks warm stratification following 8 weeks cold stratification, low sulfatase activity was detected in testa and endocarp. Figure 15 showed the activity. 16.
(29) of sulfatases of P. campanulata seeds exposed to continue warm then cold stratification, and the cold stratification started from the seventh week to the fourteenth weeks. The dash line indicates the germination percentage for the same treatment and time.. Seeds treated with 6 weeks warm plus 8 weeks cold stratification. germinated to 98%, and sulfatase activity was low but had stable tendency. However, seeds treated with warm stratification only germinated less than 10%, but exhibited maximum sulfatases activity.. 17.
(30) Chapter 4. Discussion. 4.1 Germination of P. campanulata seeds According to the germination experiments of P. campanulata seeds, most of seeds do not germinate under either warm stratification or cold stratification. Warm plus cold stratification could induce seed germination significantly.. The result. indicated that a combination of warm and cold stratification is necessary for releasing dormancy of P. campanulata seeds completely. Small amount of seeds germinated after cold stratification only, and sometimes the germination percentage was over ten percent depending upon the year and the location of the seed harvested (Chien et al., 2000), such as seed harvested from Coaling Shihbi park in 2005. Only cold-stratified seeds germinated differently possibly due to different maturity of freshly-harvested seeds.. In practice, a mixture of fruits at different degree of ripeness could be. harvested from different trees although the pericarp color of fruits appeared mature (Chien et al, 2006). Warm followed by cold stratification were capable of releasing seed dormancy completely and promoting germination. Sometimes cold stratification also stimulated seed germination, but retained about 10-30% ungerminated seeds in the stratification periods. The timing for stratification is different yearly especially at warm period. It is likely that some seeds need more experienced “warm treatment”. 18.
(31) before they were delivered from the field to the laboratory or it may just reflect the heterogeneity of the seeds.. 4.2 The activity of sulfatases/ST during the progress of dormancy breaking in P. campanulata seeds We found that significant change in the activity of desulfation following warm and/or cold stratification during the progress of dormancy breaking in P. campanulata seeds. When P. campanulata seeds were harvested freshly, there is less sulfatase activity exhibited in seed envelope. The activity of sulfatases was induced when seeds detected the signal of warmness plus moisture. The sulfatase activity exhibited maximum in only warm-stratified period. It is speculated that specific hydrolysis reactions may participate to initial stage of seed dormancy and dormancy breaking. A stable sulfatase activity results in the release of seed dormancy and promotion of germination. The activity of sulfotransferase was less detected within the period of stratification. However, the seed extract from testa and endocarp of freshly-harvested P. campanulata seeds inhibited phenol sulfotransferase (rat ST1A1) activity strongly. Stratified seed extract reduced the inhibition on rat ST1A1 (Tsai YY, 2005, unpublished data) (Figure 16). Significant variations in seed proteins were also observed by 2D gel electrophoresis before and after breaking of seed. 19.
(32) dormancy in P. campanulata (Lee et al., 2006). The warm and/or cold stratification may act as a signal to change germination potential of P. campanulata seeds (Chien et al., 2006). Sulfatase activity arose in P. campanulata seeds from warm stratification and the desulfation may participate to regulate the mechanism in the initial stage of P. campanulata seed dormancy.. 4.3 Influence of endocarp and tests on P. campanulata seed dormancy In P. campanulata seeds, the sulfatases activity was detected in seed envelope including endocarp and testa. Removing both endocarp and seeds coat promoted radicle protrusion promptly, thus a covering-imposed dormancy appeared in P. campanulata seeds (Chien et al., 2006). The sulfatases in testa and endocarp may be induced or remain for of physiological reactions for seed dormancy and germination. The testa in higher plant seeds is known a multifunctional tissue which may interfere with dormancy, and also protects the embryo against adverse environmental conditions (Debeaujon et al., 2000). Seed testa may contain some germination inhibitors, such as phenolic compounds and ABA, to cause chemical dormancy (Lin, 1996). An inhibitory nature was considered to locate mainly in the covering tissues of the freshly harvested P. campanulata seed (Chien et al., 2006). It was found that high ABA contents were detected in the testa but not in the embryo and cotyledon, and reduced. 20.
(33) 6~12 times during warm and/or cold stratification; a higher concentration of GA4 was detected in embryo and cotyledon of dormancy-breaking P. campanulata seed (Lee et al., 2006). Lee et al. also found that protein patterns of testa significantly differed from those of the cotyledon and embryo in dormant and dormancy-breaking P. campanulata seeds. Germination experiments also reveal that the removal of testa of P. campanulata seed eliminated its dormancy effect (Chien et al., 2006). However, Geneve (2003) indicated that embryos of deep dormant seeds isolated from these seeds either will not germinate or will grow into abnormal seedlings with a dwarf phenotype, termed physiological dwarf. It seems that not only germination inhibitors but also development promoters are contained in seed testa.. Furthermore, the higher total. sulfatase activity in P. campanulata seeds was assayed in endocarp than activity in testa. There may be some biomolecules hydrolyzed from woody endocarp and testa influx into embryo to regulate the seed dormancy.. Nevertheless, there was few. publication about influence of endocarp on seed dormancy, but endocarp was thought to prevent imbibition and form seed mechanical dormancy. Germination experiments reveal that the removal of endocarp of P. campanulata seed eliminated about 20% dormancy effect (Figure 17) (Chien et al., 2006). There was no report about the influence of endocarp on seed chemical dormancy. We detected sulfatase activity in endocarp and testa of treated P. campanulata seeds, but not in freshly-harvested seeds. 21.
(34) and treated embryo. Desulfation could affect some metabolites in testa and endocarp of P. campanulata seeds between dormancy maintenance and releasing.. 4.4 Dormancy releasing of P. campanulata seeds P. campanulata seeds are deeply dormant. They exhibit morphological as well as physiological dormancy due to the undeveloped embryo at the time of fruit collection (Chien et al., 2002). Freshly-harvested and intact seeds of P. campanulata required a 1-6 weeks warm followed by 8 weeks cold stratification for complete germination.. However, P. campanulata seeds treated by just 8 weeks cold. stratification explored different germination percentages in different harvesting years and places. Many scientists discussed that how temperatures affect the physiology to release seed dormancy. According to Geneve (2003), in nature, morphophysiological seeds are usually shed from the plant with an undeveloped embryo that requires a warm period to initiate growth inside of seed covering (morphological dormancy) (Table 2).. Once the embryo reaches a certain size, it can respond to chilling. temperature, which releases physiological dormancy, so seeds usually require warm (>15℃) followed by cold (1 to 10℃) conditions, i.e. embryo development during the warm temperature cycle and breaking physiological dormancy during the chilling cycle. This is not for P. campanulata seeds because removal of teata and endocarp. 22.
(35) readily release dormancy (Chien et al., 2006) (Figure 17). We could speculate that seeds were treated by warm stratification for embryo development and maturity, and treated by cold stratification for releasing physiological dormancy, but mechanisms are not understood. Lin (1996) considered that physiologically dormant seeds have the low metabolic ability in embryo and need to stratification to remove the germination inhibitor and produce germination promoter. The warm stratification may allow P. campanulata to continuous seed development to obtain more proportion of mature seeds in the seed lots (Chien et al., 2006). Many proteins in P. campanulata seed sections increasing or decreasing following warm stratification, and these proteins may be involved in the post-harvested maturation of seeds (Lee et al., 2006). In the other hand, temperature also impacts dormancy release for seeds with exogenous physical dormancy on seed covering. For instance, some seeds require high temperature or daily fluctuations (>15℃ change) in temperature to allow imbibitions (Baskin and Baskin, 1998; Geneve, 2003). Effects of warm temperature on seeds are not only permeability of seed envelope but also embryo full development. It is necessary for seed stratification to imbibe moisture. Factor affecting seed dormancy is the interaction between temperature and seed moisture content. Chilling stratification is not effective unless seeds are hydrated (Geneve, 2003). Some catabolic reactions catalyzed by enzymes occur during imbibition. According. 23.
(36) to Vertucci (1993), very few enzyme-mediated reactions occur when dry seeds first hydration level (type 1 water). Some catabolic reactions catalyzed by enzymes occur in second hydration level (type 2 water), but seed germination can only be completed if type 5 water is present, although some metabolism necessary for germination (i.e. protein synthesis) occurs in the fourth hydration level (type 4 water) (Figure 18). Lee et al. (2006) showed that changes in some group of proteins probably indicate biochemical activities of the P. campanulata seed after water uptake (Lee et al., 2006). Water uptake may improve enzyme activity of catabolism such as hydrolases and sulfatases. The brief hypothesis of the correlation between seed dormancy and desulfation regulation is shown in Figure 19. Seed testa and endocarp contain several metabolites to regulate dormancy and germination. Seeds germination in various treatments implied that germination inhibitors or dormancy inducers were probably increased or induced during warm stratification, and those were decreased or removed during cold stratification (Lee et al., 2006). Sulfatases may be the key to increase the concentrations of dormancy inducers and germination inhibitors.. When warm. stratification processed, seed germination was inhibited and embryo was getting fully mature. Mature seeds were treated by cold stratification and germination inhibitors in seeds were degraded, and then seeds germinate completely.. 24.
(37) Chapter 5. Conclusion. This is the first report of sulfatase activity in woody plant.. Significant. change of the sulfatase activity was observed following warm and cold stratification, the treatments for the dormancy breaking of Taiwan cherry (P. campanulata) seed. The sulfatase activity exhibited maximum in only warm-stratified period, and Seed germination followed sulfatase activity occurrence. Sulfatase activity was detected, but PST activity was not. However, the activity of rat phenol sulfotransferase was inhibited strongly by the seed extract. The effect of warm and cold stratification on the activities of sulfation/desulfation was first reported and analyzed. It speculated specific hydrolysis reactions which relate to embryo development and maturity may participate to initial stage of seed dormancy and germination. This research provides a novel view to investigate the biological mechanism of seed dormancy.. 25.
(38) Acknowledgement. We appreciate Dr. Ching Te Chien, Ms. Shun Ying Chen and coworkers of Dr. Chien’s team to provide seed materials and assistance and guidance about stratification and germination tests.. 26.
(39) Table 1. Germination percentage of P. campanulata seeds following warm or/and cold stratification.. 1-1 germination before 2003 Treatment (weeks) *Germination (%). Freshlyharvested seeds 0. Warm 6 + Cold 8 99.3. Warm 6. Cold 2. Cold 4. Cold 6. Cold 8. 0. 0.7. 22.7. 70.0. 89.3. 1-2 germination in 2004 Treatment (weeks). *Germination (%) Cold 4 Cold 5 Cold 6 Cold 8. Freshlyharvested (Warm 0) 4.0 5.3 8.7 12.0. Warm 1. Warm 2. Warm 3. Warm 4. Warm 5. Warm 6. 3.0 57.3 94.0 100. 2.7 68.0 87.3 100. 4.7 70.0 95.3 98.7. 5.3 33.3 87.3 98.7. 2.7 52.7 90.0 100. 6.0 52.7 72.0 98.0. 27.
(40) Table 1. continued. 1-3 germination in 2005 Treatment (weeks) *Germination (%). Freshlyharvested 0. Warm 6 + Cold 8 100. Warm 6 0.7. Cold 2 0.7. Cold 4 22.0. Cold 6 51.3. Cold 8 -. Cold 1 1.3. Cold 3 8.7. Cold 5 30.7. Cold 7 48.0. Cold 9 68.0. Treatment (weeks) Germination (%). * Germination, judged by radicle protrusion of at least 5 mm, was recorded weekly for a total 8 weeks. The abbreviation: Warm, seeds were treated by warm stratification; Cold, seeds were treated by cold stratification; -, no data.. 28.
(41) Table 2 Dormancy categories. (Geneve, 2003) Dormancy type 1. primary dormancy a. exogenous dormancy Physical Chemical b. endogenous dormancy Morphological Rudimentary Linear Physiological Nondeep. Intermediate Deep c. combinational Morphophysiological Epicotyl Epicotyl and radicle (double dormancy) Exo-endodormancy. 2. secondary dormancy a. thermodormancy b. conditional dormancy. Causes of dormancy Imposed by factors outside the embryo Impermeable seedcoat Inhibitors in seed coverings Imposed by factors in the embryo The embryo is not fully developed, at the time the seed shed from the plant Small undifferentiated embryo Small differentiated embryo < 1/2 size of the seed Factors within embryo inhibits germination Positively photodormant (requires light) Negatively photoodormant (inhibited by light) After-ripen Embryos germinates if separate from the seed coat, often responds to gibberellic acid Embryos do not germinate when remove from seedcoat or will form a physiological dwarf Combination of different dormancy, conditions that must be satisfied sequentially Combination of underdeveloped or rudimentary embryo and physiological dormancy Radicle begins growth when temperature and water permit, but epicotyl is dormant Radicle and epicotyl require chilling stratification, but radicle is released during first year and then Combination of exogenous and endogenous dormancy conditions. ex. Physical (hard seedcoat) + intermediate physiological dormancy After primary dormancy is relieved, high temperature induced dormancy Change in ability to germinate related to time of the year. Conditions to break dormancy. Scarification. Representative genera. Baptisia, Convolvulus, Gleditsia, Lupinus, Beta, Iris. Removal of seed covering (fruits) Leaching seeds Warm or cold stratification Cold stratification and potassium nitrate Warm stratification and gibberellic acid. Anemone, Ranunculus Daucus, Cyclamen, Viburnum. Red light Darkness Short period of dry storage Moderate periods (up to 8 weeks) of cold stratification. Lactuca, primula Cyclamen, Nigella Cucumis, Impatiens Aconitum, Cornus, Pinus. Long periods (> 8 weeks) of cold stratification. Dictamnus, Euonymus, Prunus, Rhodotypos. Cycle of warm and cold stratification. Asimina, Helleborus, Magnolia, Mertensia Asarum, Paeonia. Warm followed by cold stratification. Ilex,. Cold stratification followed by warm followed by a second cold stratification Sequential combinations of dormancy release treatments. Ex. Scarification followed by cold stratification. Convallaria, Trillium. Growth regulators or cold stratification. Apium, Lactuca, Viola. Chilling stratification. Many species with endogenous dormancy display conditional dormancy. 29. Cercis, Tilia.
(42) a.. CH3. N. HO O P O HO. O. N O. S. -. O. O O. O. O. HO. P O OH. N. HO. O O S O O HO P O HO. O. MU. OH. PAP. PST. O. N. O. MUS. HO. NH2. PST. N N O. N. NH2 N. oso3. O. ex. 350nm em. 450nm. 2-naphthol. P O HO OH. OH. 2-naphthyl-S. PAPS. b.. CH3. SULFATASES. O -. S. O. O O. O. O. HO. O. O. MU. MUS. ex. 350nm em. 450nm. Figure 6 The reaction of enzyme assay with fluorescence photometry by assaying MU in 350 nm excitation and 450 nm emission. a. The transfer reaction of phenol sulfotransferases (PST). sulfatases. 30. b. The hydolysis reaction of.
(43) 1. warm and cold stratification of Prunus campanulata seeds. 2. extract enzyme solution from treated seeds. 7. germination test. 3. enzyme assay of sulfatases and phenol sulfotransferases. 8. analysis of germination percentage. 4. separation of low MW molecules from seed extract (desalted extract). 5. enzyme assay of sulfatases and sulfotransferases. 6. analysis of enzyme activity. Figure 7 The overview of experimental flow chart.. 31.
(44) germination test of P. campanulata. 100. 2003 2004 2005. 90 80. germination(%). 70 60 50 40 30 20 10 0 0. 1. 2. 3. 4 5 6 cold-treated period(weeks). 7. 8. 9. Figure 8 Germination percentages of P. campanulata seed following cold stratification in three different year: before 2003, 2004, 2005.. 32.
(45) hilum endocarp seed coat testa. cotyledons epicotyl. embryo. hypocotyl radicle endosperm. Figure 9 Longitudinal section through a seed of P. campanulata. A small amount of degenerate endosperm can be seen to be adherent to the hypocotyl and radicle (modifying from Chien, et al., 2006).. 33.
(46) sulfatase activity in seed endocarp, testa and embryo. sulfatase activity (pmol/min/mg). 6. 5. endocarp testa embryo. 4. 3. 2. 1. 0 2005 fresh 2005 W3. 2005 W6. 2005 C8 2005 W6C8 2004 W3. treatment. Figure 10 Sulfatase activity was detected in endocarp, testa , and embryo. .. 34.
(47) 3.0. 3.0. activity (pmol/min/mg). 2.0. w only w+c4 w+c6 w+c8 c only. 2.5. w only w+c4 w+c6 w+c8 c only. 2.5. activity (pmol/min/mg). b. testa. a.endocarp. 1.5. 1.0. 2.0. 1.5. 1.0. 0.5. 0.5. 0.0. 0.0 0. 2. 4. 6. 8. 10. 12. 14. 0. (weeks). 2. 4. 6. 8. 10. 12. 14. (weeks). Figure 11 The activities of sulfatase in P. campanulata seed extracts during the warm and/or cold stratification in 2004. a, sulfatase activities in seed endocarp, b, sulfatase activities in seed testa. The x-axis indicates to the sum of the periods of warm and cold stratification (the total stratified time). The y-axis indicates to sulfatase activities detected at different stratifying conditions.. 35.
(48) 3.0. 3.0. a. endocarp (desalted) w only w+c4 w+c6 w+c8 c only. 2.0. w only w+c4 w+c6 w+c8 c only. 2.5. activity (pmol/min/mg). activity (pmol/min/mg). 2.5. b. testa (desalted). 1.5. 1.0. 0.5. 2.0. 1.5. 1.0. 0.5. 0.0. 0.0. 0. 2. 4. 6. 8. 10. 12. 14. (weeks). 0. 2. 4. 6. 8. 10. 12. 14. (weeks). Figure 12 The activities of sulfatase in P. campanulata desalted seed extracts during the warm and/or cold stratification in 2004. a, sulfatase activities in seed endocarp, b, sulfatase activities in seed testa. The x-axis indicates to the sum of the periods of warm and cold stratification (the total stratified time). The y-axis indicates to sulfatase activities detected at different stratifying conditions.. 36.
(49) 3.0. 3.0. B. testa. 2005 2004 2003. 2.5. sulfatase activity (pmol/min/mg). sulfatase activity (pmol/min/mg). A. endocarp. 2.0. 1.5. 1.0. 0.5. 2.5. 2005 2004 2003. 2.0. 1.5. 1.0. 0.5. 0.0. 0.0 0. 2. 4. 6. 8. 10. 12. 0. 14. 2. 4. 6. 8. 10. 12. 14. stratification time (weeks). stratification time (weeks). Figure 13 The tendency of sulfatase activities in P. campanulata seed extracts during the warm stratification in 2003(W0 to W12), 2004(W0 to W6) and 2005(W0, W3, W6). The points at the fourteenth week were W6C8 treated seeds. a, sulfatase activities in seed endocarp, b, sulfatase activities in seed testa.. 37.
(50) 3.0. 3.0. 2005 2004. 2.5. B. testa sulfatase activity (pmol/min/mg). sulfatase activity (pmol/min/mg). A. endocarp. 2.0. 1.5. 1.0. 0.5. 0.0. 2005 2004. 2.5. 2.0. 1.5. 1.0. 0.5. 0.0 0. 2. 4. 6. 8. 10. 12. 14. 0. 2. 4. 6. 8. 10. 12. 14. stratification time (weeks). stratification time (weeks). Figure 14 The tendency of sulfatase activities in P. campanulata seed extracts during the cold stratification in 2004(C5, C6 and C8), and 2005 (C0 to C8). The points at the fourteenth week were W6C8 treated seeds. a, sulfatase activities in seed endocarp, b, sulfatase activities in seed testa.. 38.
(51) Figure 15 P. campanulata sulfatases activity and percentage of germination during warm plus cold stratification.. 39.
(52) 100. relative activity of sulfotransferase (%). A. endocarp 2004 fresh 2004 W1 2004 W5 2004 W6 2004 W6C4 2004 C8. 80. 60. 40. 20. 0 0. 200. 400. 600. 800. endocarp weight (mg) 100. relative activity of sulfotransferase (%). B. testa 80. 60. 40. 20. 0 0. 20. 40. 60. 80. 100. 120. testa weight (mg). Figure 16 The activities of phenol sulfotransferase (Rat ST1A1) were inhibited by extract from P. campanulata seeds during warm/cold stratification. A, inhibition of endocarp extract. B, inhibition of testa extract (Tsai YY, 2005, unpublished data). 40.
(53) Germination (%). 100. 80. 60. 40. 20. 0 0. 1. 2. 3. 4. 5. 6. 7. 8. 9 10 11 12 13 14 15 16 17 18 19 20 21. Time of incubation (days). Figure 17 Removal of testa and endocarp readily remove seed dormancy of P. campanulata. Germination percentage of P. campanulata seeds with both the endocarp and seed coat removed (open circles), with the endocarp removed (seed coat retained) (closed circles), with the endocarp removed and treated with 100 ppm GA3 (closed triangles), and intact seed (closed squares) was plotted against time of incubation (Chien et al., 2006).. 41.
(54) Figure 18 The relation between water uptake and seed physiology. (Vertucci, 1993). 42.
(55) --OH : germination inhibitors or dormancy inducer SO3Warm stratification. Sulfatase activity. OSO3-. Cold stratification --OH (degradation?). Sulfotranferase activity ? induce or maintain dormancy. inhibit or active germination process?. inhibit germination process (embryo development). release dormancy. induce germination. Figure 19 The hypothesis of desulfation regulation in seed dormancy and germination. It speculated that seed testa and endocarp contain several metabolites to regulate dormancy and inhibit germination.. 43.
(56) References. Baskin CC, Baskin JM, (1998) Seeds: Ecology, Biography, and Evolution of Dormancy and Germination. San Diego, CA, USA: Academic press. Baum H, Dodgson KS (1957) Differentiation between myrosulphatase and the arylsulphatases. Nature 179:4553-4554. Bewley JD (1997) Seed germination and dormancy. The Plant Cell 9:1055-1066. Chien CT, Chen SY, Chang WL (2002) Stratification and Gibberellin Treatment for Seeds of Four Taiwanese Tree Species. Taiwan J For Sci 17 (1):51-57 (in Chinese with English summary). Chien CT, Chen SY, Chung JD, and Yang YS (2006) Association of seed covering and gibberellin contents with dormancy and germination in cherry seeds, Prunus campanulata., Seed Sci. Res., Submitted.. Chien CT, Chen SY, Yang JC (2002) Effect of stratification and drying on the. 44.
(57) germination and storage of Prunus campanulata seeds. Taiwan J For Sci 17 (4):413-20. Chien CT, Lin SH, Hsu PJ (1996) The strategies enhancing germination of Prunus campanulata seeds. Taiwan For. 22(2):10-11 (in Chinese). Chapmen E, Best MD, Hanson SR, Wong C-H (2004) Sulfotransferases: structure, mechanism, biological activity, inhibition, and synthetic utility. Agnew. Chem. Int. Ed. 43: 3526-348. Debeaujon I, Koornneef M, (2000) Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol. 122:415-424. Debeaujon I, Leon-Kloosterziel KM, Koornneef M, (2000) Influence of the testa on seed dormancy, germination, and longevity in arabidopsis. Plant physiology 122:403-413. de Hostos EL, Togasaki RK, Grossman A (1988) Purification and biosynthesis of a. 45.
(58) derepressible periplasmic arylsulfatase from Chlamydomonas reinhardtii. J. Cell Biol. 106:29-37. Faulkner IJ, Rubery PH (1992) Flavonoids and flavonoid sulphates as probes of auxin-transport regulation in Cucurbita pepo hypocotyl segments and vesicles. Planta 186: 618–625. Gegenheimer P, (1990) Preparation of extracts from plants. Meth. Enzymol. 182: 174-193. Geneve RL (2003) Impact of temperature on seed dormancy. Hort. Sci. 38: 336-341. Gidda SK, Miersch O, Levitin A, Schmidt J, Wasternack C, Varin L (2003) Biochemical. and. molecular. characterization. of. a. hydroxyjasmonate. sulfotransferase from Arabidopsis thaliana. J. Biol. Chem. 278(20):17895-17900. Grisez TJ, Barbour JR, Karrfalt RP (2000) Prunus L., IN: Woody Plant Seed Manual, F.T. Bonner (tech. coord.), R.G. Nisley (managing ed.), USDA-Forest Service, Washington , DC. (http://ntsl.fs.fed.us/wpsm/Prunus.pdf). 46.
(59) Hallmann A and Sumper M (1994) An inducible arylsulfatase of Volvox carteri with properties suitable for a reporter-gene system. Eur. J. Biochem. 221:143-150. Hanai H, Nakayama D, Yang H, Matsubayashi Y, Hirota Y, Sakagami Y (2000) Existence of a plant tyrosylprotein sulfotransferase: novel plant enzyme catalyzing tyrosine O-sulfation of preprophytosulfokine variants in vitro. FEBS Lett. 470:97-101. Hanson SR, Best MD, Wong C-H (2004) Sulfatases: structure, mechanism, biological activity, inhibition, and synthetic utility. Angew. Chem. Int. Ed. 43:2-29. Harris RM, Wood DM, Bottomley L, Blagg S, Owen K, Hughes PJ, Waring RH, Kirk CJ (2004) Phytoestrogens are potent inhibitors of estrogen sulfation: implications for breast cancer risk and treatment. JCEM 89(4):1779-1787. Hell R (1997) Molecular physiology of plant sulfur metabolism. Planta 202:138-148. Hiroyoshi O in: Editorial Committee of the Flora of Taiwan, Rosaceae, Flora of. 47.
(60) Taiwan, 2nd ed., 3, Taipei, Taiwan:Publisher. 1993, pp. 69-157. Jensen M, Eriksen EN (2001) Development of primary dormancy in seeds of Prunus avium during maturation. Seed Science and Technology 29: 307-320. Johnson DA, Wu T, Li P, Maher J (2000) The effect of steroid sulfatase inhibition on learning and spatial memory. Brain research 865:286-290. Knauff U, Schulz M, Scherer HR (2003) Arylsulfatase activity in the rhizosphere and roots of different crop species. Europ. J. Agronomy 19:215-223. Ko Y (2002) Plant Physiology. Taipei, Taiwan: Yihsient press (in Chinese). Koornneef M, Bentsink L, Hilhorst H (2002) Seed dormancy and germination. Curr. Opin. Plant Biol. 5:33-36. Lang GA (1994) Dormancy - the missing links: integrating molecular information. Hort Sci. 29:1255-1263. 48.
(61) Lee CS, Chien CT, Lin CH, Chiu YY, Yang YS (2006) Proteins changes between dormant and dormancy-broken seeds of Prunus campalunata Maxim. Proteomics 6(14). (accepted). Leustek T, Saito K (1999) Sulfate transport and assimilation in plants. Plant Physiol. 120:637-643. Lin TP (1996) Harvest, Treatment, Storage, Dormancy, and Germination in Seeds of Woody Plants. Taiwan: Taiwan Forestry Research Institute press (in Chinese). Matsubayashi Y, Ogawa M, Morita A, Sakagami Y (2002) An LRR receptor kinase involved in perception of a peptide plant hormone, phytosulfokine. Science 296:1470-1472. Matsubayashi Y, Sakagami Y (1999) Characterization of specific binding sites for a mitogenic sulfated peptide, phytosulfokine-a, in the plasma-membrane fraction derived from Oryza sativa L. Eur. J. Biochem. 262:666-671. Matsubayashi Y, Yang H, Sakagami Y (2001) Peptide signals and their receptors in. 49.
(62) higher plants. Trends in Plant Science 6(12):573-577. Ohkimoto K, Liu M, Suiko M, Sakakibara Y, Liu M (2004) Characterization of a zebrafish estrogen-sulfation cytosolic sulfotransferase: inhibitory effects and mechanism of action of phytoestrogens. Chemico-biological interaction 147:1-7. Pai TG, Suiko M, Sakakibara Y, Liu M (2001) Sulfation of flavonoids and other phenolic dietary compounds by the human cytosolic sulfotransferases. Biochem. Biophys. Res. Comm. 285:1175-1179. Parenti G, Meroni G, Ballabio A (1997) the sulfatase gene family. Curr Opin Genet Dev. 7:386-391. Saito K (2000) Regulation of sulfate transport and synthesis of sulfur-containing amino acids. Curr. Opin. Plant Biol. 3:188-195. Seeley SD (1997) Quantiication of endodormancy in seeds of woody plants. HortScience 32: 615-617. 50.
(63) Stephen JR, Dent KC, Finch-Savage WE (2003) Molecular responses of Prunus avium (wild cherry) embryonic axes to temperatures affecting dormancy. New Phytologist 161: 401-413. Varin L, Ibrahim RK (1992) Novel flavonol 3-sulfotransferase, purification, kinetic properties, and partial amino acid sequence. J. Biol. Chem. 267:1858-1863. Varin L, Marsolais F, Richard M, Rouleau M (1997) Biochemistry and molecular biology of plant sulfotransferase. FASEB J. 11:517-525. Vertucci CM (1993) Predicting the optimum storage conditions for seeds using thermodynamic principles. J. Seed Technol. 17:41-53. Weidner S, Paprocka J (1997) Preharvest sprouting as related to change in concentration of phenolic compounds in cereal grain and embryo sensitivity to phenolic acids during seed development. Proceedings of COST 828 Workgrou. p. 2. Meeting, Barcelona, November 10.. 51.
(64) Yildiz FH, Davies JP, Grossman AR (1994) Characterization of sulfate transport in Chlamydomonas reinhardtii during sulfur-limited and sulfur-sufficient growth. Plant Physiol. 104:981-987. 52.
(65) APPENDIX MATERIALS AND METHODS. 藥品與方法. A. Equipments equipment. Company. Hitrap desalting column. Pharmacia (Hong Kong). U3300 UV/VIS Spectrophotometer. Hitachi. F-4500 Fluorescence Spectrophotometer. Hitachi. Orbital Shaker Incubator. DEHG YNG. Uitrafiltration System. Amicon. Eppendrof centrifuge 5410. Eppendrof. Himac CR 22G 高速離心機. Hitachi. pH meter. Fisher Scientific. PRO 250 homogenizer. PRO Scientific. Mixer Mills MM 301. Retsch. 53.
(66) B. Chemicals and Reagents (1) Extract buffer reagent. final conc.. MW. Tris-HCl. 10mM. 121.14. Sucrose. 125mM. 342.30. Glycerol. 10%. DTT. 1mM. 154.2. (dithiothreitol). others pH=7.4 or pH=8.0. 使用前加入,加入後的 buffer 保存於 4℃。 置於-20℃冰箱,拿出後放入乾燥箱備用Æ 易吸水!. EDTA. 1mM. 372.24. reagent. stock conc.. MW. PMSF. 200mM. 174.2. others 以 isopropanol 做 solvent。 buffer 在使用前皆需要加入 1mM 的 PMSF。. 54.
(67) (2) The reagent of sulfatase/ST assay with fluorescence spectroscopy reagent. Stock. MW. others. 250mM. 78.13. Fluka,濃度≧99%,d=1.115,置於防爆箱。. Tris-base. 0.2M. 121.14. pH=7.0(視需求而定). β-naphthol. 0.5mM. 144.2. 怕光,置藥品櫃,分裝時用棕色的. conc. 2-mercaptoethanol (C2H6OS ). eppendoff;acetone 置防爆櫃 MUS (4-methyl-. 10mM. 294.3. 置-20℃冰箱,取出時先置於乾燥箱。. umbelliferyl sulfate) MU (4-methyl-. 溶液需避光。 1 μM. 置乾燥箱,store at R.T.,黃色粉末;350nm. 198.2. 吸收, 450nm 發出螢光. umbelliferone) (C10H7O3Na ) PAP (adenosine. 1mM. 427.2. 置-20℃冰箱,取出時先置於乾燥箱。 以A260定濃度(以ddH2O歸零,ε=15100. 3’,5’-diphosphate). M-1cm-1) AMP (adenosine. 1mM. 368.2. 置-20℃冰箱,取出時先置於乾燥箱。 以A260定濃度(以ddH2O歸零,ε=15400. -5’-monophosphate). M-1cm-1). C. 酵素活性計算公式 每 mg 種子組織萃取酵素活性. =. (E x 450 改變量. ( pmol / min/ mg ). × 萃取液總體積 (ε × assay 加入的萃取液體積. (mL ) × 10 6 ) ÷ 總組織重 (mg ) (μ L )). 55.
(68) AUTOGRAPHY Education 2003-now. Laboratory of Enzyme and Protein Engineering, Institute of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan. 1999-2003. Department of Applied Chemistry, National Chiao Tung University, Hsinchu, Taiwan. 1996-1999. National Pingtung Girls’ Senior High School, Pingtung, Taiwan. 1993-1996. Chung Cheng Junior High School, Pingtung, Taiwan. 1987-1993. Chung Hsiao Elementary School, Pingtung, Taiwan. Experience 2003-2006. Teaching assistant of General Chemistry Experiment, Department of Applied Chemistry, National Chiao Tung University, Hsinchu, Taiwan. 2002-2005. Project assistant of BioWeb, Department of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan (http://biotech.life.nctu.edu.tw/). Publication 1. 江秉潔,邱靜玟,陳羿蓉,陳韋宏,林志生。食藥用菇類專利趨勢分析 (The Patent Analysis of Edible or Medical Fungi)。食品工業,37:5,民 94.05,頁 51-60。. 56.
(69)
數據

+7





相關文件
The first row shows the eyespot with white inner ring, black middle ring, and yellow outer ring in Bicyclus anynana.. The second row provides the eyespot with black inner ring
After the Opium War, Britain occupied Hong Kong and began its colonial administration. Hong Kong has also developed into an important commercial and trading port. In a society
• helps teachers collect learning evidence to provide timely feedback & refine teaching strategies.. AaL • engages students in reflecting on & monitoring their progress
Robinson Crusoe is an Englishman from the 1) t_______ of York in the seventeenth century, the youngest son of a merchant of German origin. This trip is financially successful,
fostering independent application of reading strategies Strategy 7: Provide opportunities for students to track, reflect on, and share their learning progress (destination). •
Strategy 3: Offer descriptive feedback during the learning process (enabling strategy). Where the
How does drama help to develop English language skills.. In Forms 2-6, students develop their self-expression by participating in a wide range of activities
The IEC endeavours to ensure that the information contained in this presentation is accurate as of the date of its presentation, but the information is provided on an
