以獲取運動速度的最大衝量原理探討垂直跳動作的擺臂作用效果
16
0
0
全文
(2) 國立臺灣體育學院學報第十四•十五期. APPLICATION OF MAXIMUM IMPULSIVE PRINCIPLE IN THE STUDY OF THE EFFECT OF ARM ACTIONS ON THE VERTICAL JUMP Chung-Yu Chen Abstract The larger area of effective impulse during a vertical jump corresponds a greater jump height. However, the principle of maximum impulse to achieve movement velocity indicated that there are three methods to increase the impulse area: to increase the peak force, to lengthen the time of applied force, and to raise the slope of force curve. The present study aimed to investigate the effect of the arms swing and no arms swing on countermovement vertical jump (CMJ) that acquired by force platform (1200Hz) and to compare the difference of 15 male college athletes from 12 female college athletes performing CMJ by the principle of maximum impulse to achieve movement velocity. This study also compared the reaction characters of EMG signals (1200Hz) and kinetics (120Hz) in order to interpret the mechanism of arms swing during CMJ. The results showed that lengthening the force applied time was the main factor to increase jumping height for arms swing versus no arms swing (p< .05), and female jumpers due to lesser strength factors their jumping height were lower than male jumpers height (p< .05), but the effect of arms swing showed no any gender difference (p>.05). EMG data indicated that the hamstrings might produce the action of plantar flexion during the arms swing of CMJ in order to postpone the gastrocnemius and soleus accelerating for contraction.. KEY WORDS: countermovement vertical jump, arms swing, electromyography, kinetics.. 2.
(3) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. 壹、緒論 一、問題背景 人體運動的過程中,經常有其他肢體伴隨共同作用的現象,例如在田徑的 跨欄時,運動員為了平衡下肢的抬高跨步動作,對側上肢就會動作幅度加大地 產生與下肢運動方向相反的運動,而通常生物力學研究者會以角動量守恆 (conservation of angular momentum)的觀點來加以解釋(Hay,1993;Hall,2003)。 而肢體除了產生平衡的效果以外,許多起跳動作常常有許多生物力學研究者或 運動指導者指出擺動動作可以增加起跳的蹬地力量與提高重心離地瞬間的高度 (Dapena,1999;Hall,2003),所以,跳高時起跳腳對地面蹬伸也結合了手臂的 上抬與非起跳腳的上踢或上擺工作。在體操運動或跳水運動中,肢體的擺動或 帶動也可以增加人體翻轉的角動量,而提升騰空階段旋轉的角速度(Miller, Henning, Pizzimenti, Jones, & Nelson,1989)。 上述文獻顯示肢體擺動帶來的效應不外乎是平衡身體的姿勢與增加對地面 的蹬地力量,特別是增加地面反作用力,Dapena (1999)則根據簡化的力量傳遞假 說(force transmission hypothesis),並通過 Alexander 的人體模型檢證擺臂速度相 對於人體垂直跳離地速度的關係,說明擺臂動作除了可以具有力量傳遞的功能 增加離地瞬間的速度外,擺臂也會減少髖關節伸肌向心工作的速度而加大了向 下的地面反作用力。若以原地垂直跳而言,動作者經常性透過手臂的擺動或帶 動來獲取較高的離地速度,俾使離地高度大幅增加。從生物力學的角度,垂直 跳的高度要越高,人體的離地速度就要越快,而若是要要求人體離地瞬間速度 要快,根本上就需要求人體在動作開始到離地前的下肢推蹬產生較大的衝量。 由圖 1A 的地面反作用力-時間曲線若減去動作者的體重 mg,就可得到動作者的 合力作用(ΣF=F(t)-mg),而此一合力的作用又等於 ma,所以,人體所受到的加 速度作用為:. 3.
(4) 國立臺灣體育學院學報第十四•十五期. a=. F (t ) − mg F (t ) = −g m m. 若人體重心加速度 a 對時間積分,就可獲得重心的速度(圖 1B),若重心速 度對時間積分,就可獲得人體重心動作過程的位移曲線(圖 1C),而通常力學上 定義功率(Power,也定義為爆發力)為速度與力量的乘積,因此,獲得了功率對 時間的曲線(圖 1D)。 事實上,上述的重心速度改變與地面反作用力的關係,也可以根據動量-衝 量定理(impulse-momentum relationship)加以解釋。動量-衝量的關係可表達為:. ∫. t2 t1. F ( t ) dt. = m ( v 2 − v1 ). m 為人體質量;v1 為 t1 時的重心速度;v2 為 t2 時的重心速度; F(t)為力量曲線函數。 上式等號的左邊代表著力量-時間曲線與時間軸所夾的面積,即為衝量,等 於動量的變化量(等式的右邊)。因為垂直跳過程的動作者合外力作用為 F(t)-mg, 此時若是合外力對時間積分(也就是 F(t)對體重線的面積),就會等於人體重心的 動量變化量,所以,動量-衝量的關係就應表達為:. ∫. t2. t1. ( F ( t ) − mg ) dt = m ( v 2 − v 1 ). 也就是說,F(t)對體重線下方的面積就為負衝量(為時間 1 到時間 2 的力量時間面積),F(t)對體重線上方的面積則為正衝量(為時間 2 到時間 4 的力量-時間 面積)。可見在垂直跳的動作中,若減少這負衝量的面積或是加大正衝量的面積, 均可以使人體獲得離地瞬間有較大的速度。. 4.
(5) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. 圖 1:原地垂直跳之生物力學曲線特徵。A 為地面反作用力-時間曲線;B 為重心 速度-時間曲線;C 為重心位移-時間曲線;D 為功率-時間曲線。1 為動作 開始;2 為下蹲速度最大值;3 為下蹲最低點;4 為速度最大值;5 為離地 瞬間;6 為騰空最高點;7 為著地瞬間。 由上述的生物力學原理,若是擺臂的效果提昇了原地垂直跳的離地高度, 那麼,就代表了離地瞬間速度的明顯增加,而這離地速度增加則源自於力量對 時間曲線的總面積(也就是總衝量)的增加。過去探討擺臂作用對於垂直跳效果的 相關研究,不外乎總衝量和離地速度的比較,或僅止於最大地面反作用力的探 討,Harman、Rosenstein、Frykman 與 Rosenstein (1990) 和 Payne、Slater 與 Telford. (1968)等研究也都指出擺臂垂直跳的最大地面反作用力顯著高於叉腰垂直跳的 最大地面反作用力(p< .05),研究者 Dapena (1999)更是直接認定擺臂垂直跳的地 面反作用力必大於沒有擺臂垂直跳動作而進行擺臂速度與離地速度關係的模 擬。然而,劉宇(1998)則總結了衝量-動量關係,並觀察體育運動追求跳得高、 跑得快、跳或投擲得遠的高競技水準特性,提出了「獲取運動速度的最大衝量 原理(the principle of maximum impulse to achieve movement velocity)」指出運動員 或動作表現者為了使人體或器械達到最大的運動速度,就必須使力量作用產生. 5.
(6) 國立臺灣體育學院學報第十四•十五期. 最大的衝量,其增加衝量的策略,也就是增加力量-時間曲線面積的方法有三: 增加最大力量、增加作用力的時間與增加力量-時間曲線的斜率。過去關於擺臂 效果的研究,從劉宇提出「獲取運動速度的最大衝量原理」的增加衝量策略上, 只是侷限於最大力量的探討與比較,是否有其他因為擺臂作用增加衝量的因 素,則亟待研究進一步證實了。. 二、研究目的 本研究的主要目的係以「獲取運動速度的最大衝量原理」推論的運動速度 增加可能因素為理論基礎,比較擺臂與否對垂直跳的有效衝量提昇機制。本研 究並結合肌肉電訊號與下肢關節力矩的同步計算,定性探討擺臂動作帶來之可 能效應。. 貳、研究方法 一、實驗參加者 本研究以 27 名大專校隊運動員(男生 15 名、女生 12 名)為志願的實驗參加 者,實驗參加者的基本資料如表 1,經研究者說明實驗目的與流程後簽署同意書 與基本資料調查表。 表 1:實驗參加者基本資料表。 組別 男生 平均數 標準差 女生 平均數 標準差. 6. 人數 15. 身高(cm). 體重(kg). 年齡(歲). 169.8 6.6. 61.2 6.6. 20.8 1.8. 164.6 4.3. 56.3 6.6. 20.4 1.0. 12.
(7) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. 二、實驗設備與程序 本研究以 Kistler (model 9287)測力板(1200Hz)、Biovision 多通道生物訊號收 集系統(8 頻道、1200Hz)與 Peak 高速攝影機(120Hz)等,透過同步訊號產生器進 行三機同步的訊號採集。其中,高速攝影機主要是收集實驗參加者右側矢狀面 的下肢運動學參數,並以 Peak Performance 的 Motus 二度空間影像分析系統數位 化點取實驗參加者原地垂直跳過程的肩峰、髖、膝、踝與腳尖等關節點,關節 標誌點經零相位移四階低通濾波的修勻處理後,結合 Winter (1990) 整理後的. Dempster 人體肢段參數與測力板的地面反作用力資料進行動力學逆過程(inverse dynamics)的下肢關節力矩計算;測力板收集得的原地垂直跳地面反作用力則經 BioWare Performance 1.0 版之分析軟體計算重心的壓力中心點、合力、加速度、 速度與位移等參數;肌肉電訊號係收集右側下肢的臀大肌(gluteus maximus)、半 膜肌(semimembranosus)、股二頭肌長頭(biceps femoris)、股直肌(rectus femoris)、 股內側肌(vastus medius)、脛骨前肌(tibialis anterior)、腓腸肌(gastrocnemius)與比 目魚肌(soleus)等肌肉電訊號,原始肌肉電訊號(EMG)經 Dasy Lab 6.0 版系統的. 50Hz-450Hz 濾波處理後,再取絕對值上翻處理(full-wave rectification),方進行 30Hz 的線性封波(linear envelope)處理。 實驗參加者以隨機的方式進行叉腰垂直跳與擺臂垂直跳測驗各 5 次,研究以 每一種跳躍的最佳表現(以離地速度最大值判定)作為統計分析的參數與進行曲 線特徵的比較。叉腰垂直跳動作要求實驗參加者在動作全程將手掌置於兩腰際 髖骨上緣;而擺臂垂直跳動作則要求實驗參加者在站立時雙手自兩側懸垂的自 然角度姿勢開始,動作時,下蹲並同時先手臂後擺(肩關節產生過度伸展動作) 再手臂前擺(肩關節屈)上跳離地,實驗參加者若先出現手臂前擺的肩關節屈動作 則視為失敗動作。. 7.
(8) 國立臺灣體育學院學報第十四•十五期. 三、統計分析與資料處理 測力板的動力學參數經 BioWare Performance 1.0 版分析軟體計算動作過程 的負衝量(圖 1A 的線 1 至線 2 面積)、正衝量(圖 1A 的線 2 至線 4 面積)、最大地 面反作用力(圖 1A 的線 2 至線 4 之曲線最大值)、最大發力率(圖 1A 的線 2 至線. 4 之曲線斜率最大值)與正負衝量產生的作用力時間等參數。再以 SPSS 11.0 版統 計軟體進行 2(性別)×2(跳法)的混合設計二因子變異數分析,統計的顯著水準α設 定為.05。研究的 EMG 訊號與動力學逆過程計算得的下肢關節力矩則僅作為動作 各階段之辨別與進行曲線變化特徵的定性分析。. 參、結果與討論 一、地面反作用力-時間曲線參數分析 表 2 顯示男子運動員與女子運動員完成擺臂與叉腰垂直跳的地面反作用力曲 線參數特徵。從騰空高度的比較性別與跳法的交互作用並沒有顯著差異存在(F(1, 25)=1.37,p=.25,η. (F(1,. 2. =.05,Power=.20),而在性別的主要效果則顯示男生高於女生. 25)=121.81,p=.00,η. 跳也高於叉腰垂直跳(F(1,. 2. =.83,Power=1.00)、在跳法的主要效果顯示擺臂垂直. 25)=115.39,p=.00,η. 2. =.82,Power=1.00),這一結果或. 許是可想而知,也與過去相關文獻的結果相同(Boudolos,1998;Harman 等人,. 1990)。當然,這一結果從力學原理推論也必然反映在垂直跳的擺臂跳離地速度 高於叉腰跳離地速度,且這離地速度來自於總衝量對身體質量的商,所以,由 表 2 的 總 衝 量 也 就 顯 示 性 別 與 跳 法 的 交 互 作 用 並 沒 有 顯 著 差 異 存 在 (F(1, 25)=0.72,p=.40,η. (F(1,. 2. =.03,Power=.13),而在性別的主要效果則顯示男生高於女生. 25)=100.70,p=.00,η. 2. =.80,Power=1.00)、在跳法的主要效果顯示擺臂垂直. 跳也高於叉腰垂直跳(F(1, 25)=70.60,p=.00,η2=.74,Power=1.00)。 由於垂直跳動作的地面反作用力-時間曲線的總衝量來自於正衝量與負衝量 的總和,所以,當總衝量在不同跳法與性別的主要效果達到統計的顯著差異後,. 8.
(9) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. 本研究則需進一步針對負衝量與正衝量各別分析。如表 2 所示,負衝量在性別與 跳法的交互作用有顯著差異存在(F(1, 25)=4.36,p=.05,η2=.15,Power=.52),而在 性別的主要效果則顯示男生高於女生(F(1, 25)=30.76,p=.00,η2=.55,Power=1.00)、 在跳法的主要效果則顯示擺臂垂直跳與叉腰垂直跳並沒有顯著差異存在 (F(1, 25)=0.329,p=.57,η. 2. =.01,Power=.09),這一性別差異顯示男生向下深蹲的整體. 作用均高於女生組,而有無擺臂對於原地垂直跳的差異並沒有影響負衝量的大 小,所以,擺臂與否之於總衝量的差異應就是正衝量的大小所產生了,這一結 果也確實反映在正衝量的變異數分析上。正衝量顯示性別與跳法的交互作用並 沒有顯著差異存在(F(1, 25)=0.01,p=.91,η2=.00,Power=.05),而在性別的主要效 果則顯示男生高於女生(F(1,. 25)=107.53,p=.00,η. 2. =.81,Power=1.00)、在跳法的. 主要效果顯示擺臂垂直跳也高於叉腰垂直跳 (F(1,. 25)=8.15 , p=.01 , η. 2. =.25 ,. Power=.78),可見擺臂跳的動作在騰空高度提升主要在正衝量的作用增加。 表 2:不同性別實驗參加者完成擺臂與不擺臂垂直跳動作之生物力學參數。 性別. 騰空 總衝量 負衝量 正衝量 高度*+ *+ + +* 跳 法 (m) (BW.s) (BW.s) (BW.s) 男生 叉腰跳 平均數 0.565 2.95 1.58 4.62 標準差 0.066 0.20 0.24 0.32 擺臂跳 平均數 0.657 3.14 1.50 4.78 標準差 0.062 0.19 0.27 0.37 女生 叉腰跳 平均數 0.358 2.29 1.10 3.57 標準差 0.033 0.18 0.08 0.20 擺臂跳 平均數 0.432 2.45 1.15 3.72 標準差 0.045 0.14 0.15 0.25. 最大 力量+ (BW). 最大 發力率+ (BW/s). 2.908 0.488. 17.304 6.929. 0.398 0.053. 0.439 0.040. 2.844 0.423. 18.171 6.863. 0.491 0.117. 0.459 0.032. 2.404 0.249. 12.727 4.327. 0.351 0.066. 0.389 0.050. 2.347 0.126. 12.385 3.055. 0.362 0.095. 0.460 0.111. 跳法:*p< .05. 負衝量 正衝量 時間 時間* (s) (s). 性別:+p< .05 BW 為單位體重. 9.
(10) 國立臺灣體育學院學報第十四•十五期. 依「獲取運動速度的最大衝量原理」指出衝量的增加來自於增加最大力量、 增加作用力的時間與增加力量-時間曲線的斜率等三種原因。因此,本研究則針 對貢獻正衝量的最大力量、作用力時間與最大發力率(力量-時間曲線的斜率)等 進行比較。結果發現最大力量在性別與跳法的交互作用並沒有顯著差異存在(F(1, 25)=0.01,p=.91,η. 2. =.00,Power=.05),而在性別的主要效果則顯示男生高於女生. (F(1, 25)=30.76,p=.00,η2=.55,Power=1.00)、在跳法的主要效果則顯示擺臂垂直 跳與叉腰垂直跳並沒有顯著差異存在(F(1, 25)=3.01,p=.10,η2=.11,Power=.39); 最大發力率在性別與跳法的交互作用也沒有顯著差異存在(F(1,. 25)=1.07,p=.31,. η2=.04,Power=.17),而在性別的主要效果則顯示男生明顯高於女生(F(1, 25)=5.86, p=.02,η2=.19,Power=.64),但是此一項參數差異比較的統計考驗力則較為低, 在跳法的主要效果則顯示擺臂垂直跳與叉腰垂直跳並沒有顯著差異存在 (F(1, 25)=0.20,p=.66,η. 2. =.01,Power=.07)。此一結果是與過去不管是 Harman 等人(1990). 的實證性研究或是 Dapena (1999)理論推導與模擬的基本假定等文獻完全不同, 顯示擺臂致使正衝量的增加在本研究中並沒有支持是力量值的大小或是發力率 的多寡使然。在進一步分析正衝量產生的力量作用時間,本研究發現性別與跳 法的交互作用並沒有顯著差異存在(F(1, 25)=3.41,p=.08,η2=.12,Power=.43),而 在性別的主要效果則顯示男生與女生也沒有顯著差異 (F(1,. 25)=1.46 , p=.24 ,. η =.06,Power=.21)、在跳法的主要效果顯示擺臂垂直跳也高於叉腰垂直跳(F(1, 2. 25)=10.41,p=.00,η. 2. =.29,Power=.87),此一結果顯示擺臂大幅增加了作用力的. 時間將近 50 毫秒。如果研究者假定作用力的時間係屬於垂直跳耀技術方面的指 標,那麼,從男生與女生的作用力時間沒有差異的此一結果,垂直跳的動作技 術應是沒有性別差異存在。所以,本研究也仿造過去比較垂直跳牽張-縮短循環. (stretch-shortening cycle)效果的文獻(Harrison & Gaffney,2001;Enoka,2002), 比較擺臂效果的高度提升率((擺臂跳高度-叉腰跳高度). 叉腰跳高度),結果發. 現男生(.17 ± .08)與女生(.21 ± .12)沒有顯著差異存在(F(1, 25)=1.17,p=.29,η2=.05,. Power=.18),這說明了若排除了肌力的因素,性別的差異在擺臂的效果上應是不. 10.
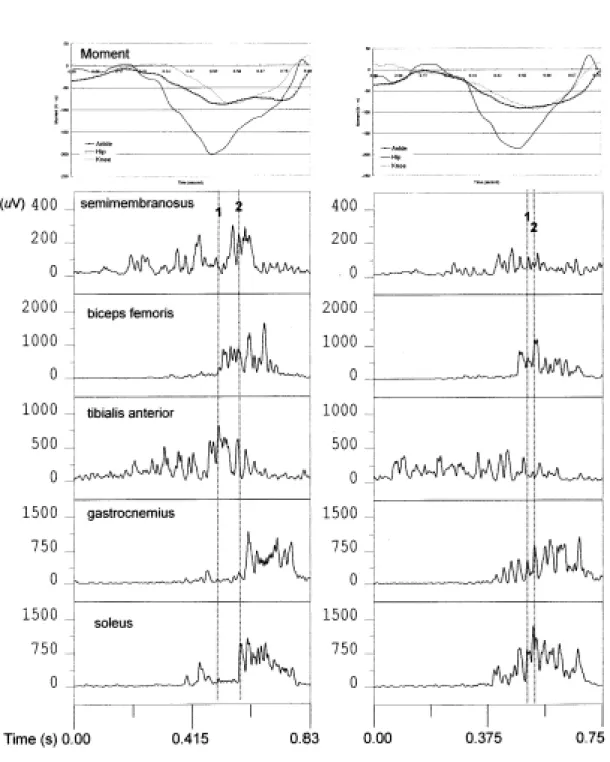
(11) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. 存在,這一結果也可以與前述作用力的時間無差異一因素相互呼應。. 二、關節力矩與肌肉電訊號分析 由於上述擺臂與叉腰垂直跳的動作時間有所不同,且在力矩的計算有使用 人體肢段參數的偏差與肌肉電訊號收集時的電極黏貼位置的一致性等研究的絕 對數值化限制,所以,在此一部分僅就動作之生物力學曲線特徵進行定性分析。 如圖 2 所示,男性實驗參加者 Q 叉腰垂直跳的動作時間略短於擺臂垂直跳約 80 毫秒,而從下肢的髖、膝、踝關節依序開始伸展的順序與時間差進行比較,研 究發現叉腰垂直跳的伸髖、伸膝、伸踝開始時間均非常接近,甚至有實驗參加 者的三個時間相同;而擺臂垂直跳的伸髖開始時間至伸踝開始時間就相差了約. 80 毫秒,且影響踝關節伸展的主要肌肉群包括腓腸肌與比目魚肌,都會在此一 伸髖開始至伸踝開始的階段出現肌肉電訊號抑制,並再從伸踝關節開始時又產 生肌肉電訊號。反觀叉腰垂直跳的此一階段,不僅幾乎沒有依序產生下肢關節 伸展的時間差,影響踝關節伸展的腓腸肌與比目魚肌也沒有明顯出現如同擺臂 垂直跳的肌肉電訊號抑制現象。因為,線 1 與線 2 分別為髖關節與踝關節開始 伸展的瞬間,若是結合此階段前後的關節淨力矩均是伸肌工作的階段,顯示在 伸髖、伸膝與伸踝關節前,下肢各關節的伸肌群應都是產生作負功的離心工作。 在這開始伸髖至開始伸踝關節的階段,擺臂垂直跳動作因為也處於上肢向 上帶動,對髖關節的作用就肢段間互動的動力學(intersegmental dynamics)概念, 將會對髖關節產生伸髖關節之依運動產生的力矩(motion dependent torque),而對 於髖關的主要伸展肌肉(屬於單關節的臀大肌與屬於雙關節的腿後肌腱群)正因 面臨由離心工作進入向心工作階段,特別是向心工作階段,本是伸展髖關節的 主要工作可能有擺臂工作產生了依運動產生的力矩介入,而轉移其作用至膝關 節或是對向前移動的小腿產生減速的作用。. 11.
(12) 國立臺灣體育學院學報第十四•十五期. 圖 2:實驗參加者 Q 在擺臂垂直跳(左側)與叉腰垂直跳(右側)的下肢關節淨力矩 (moment)與肌肉電訊號曲線。力矩負值表示關節伸展力矩,線 1 為髖關節 最大屈曲瞬間,線 2 為踝關節最大屈曲瞬間。. 12.
(13) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. van Ingen Schenau (1989)、Zajac (1993)與劉宇(1998)等人分別就力學模式或 實證現象說明雙關節肌肉或是單關節肌肉可以產生能量傳遞的功能,也可以對 所跨過的關節產生與力矩相反的作用(Lombard’s paradox 現象)或是對沒有跨過 的關節產生力矩的作用。就上述的推論,腿後肌腱群在擺臂的作用下,由於原 本應伸髖關節的主要工作為手臂擺動所取代或瓜分,而可能轉變為牽動了小腿 阻止其向前移動的趨勢,此一趨勢就致使腿後肌腱群對膝關節產生伸展的力矩. (Lombard’s paradox 現象),或是對沒有跨過的踝關節產生伸展的工作(協助減速 踝關節的繼續屈曲)。因為擺臂垂直跳在膝關節進入開始伸展階段前後,膝關節 伸肌(股直肌與股內側肌)的肌肉電訊號與叉腰垂直跳動作比較並沒有減弱之特 徵,所以,上述二種腿後肌腱群產生工作因素,對沒有跨過的踝關節產生伸展 的工作,以協助減速踝關節的繼續屈曲之可能性就大幅增加了。 圖 2 擺臂垂直跳動作的比目魚肌與腓腸肌在開始伸展踝關節前出現肌肉電訊 號抑制的現象,就恰好可以與上述的推測相互印證。比目魚肌與腓腸肌不同時 參予產生伸踝關節的工作,正可以減緩踝關節伸展的作用,以避免下肢三關節 同時工作的速度增加過快,以免提早出現人體重心速度高過下肢關節速度,而 造成踝關節尚未伸展蹬地就離開地面的情況。所以,Kibele (1998)與 Bobbert 和. van Ingen Schenau (1988)也都指出踝關節有效的產生蹠屈(dorsiflexion)工作或是 減慢蹠屈的速度,將均可以大幅增加人體重心的離地速度;擺臂垂直跳可以提 昇騰空高度的主要原因,就是影響下肢肌肉間的協調工作,進而使下肢產生最 佳化的動力鏈工作形式。. 肆、結論 本研究根據「獲取運動速度的最大衝量原理」層層抽離解析擺臂垂直跳可 以提昇重心騰空高度的原委,結果發現擺臂的作用對於垂直跳的主要貢獻為增 加作用力量的時間,進而增加離地的正衝量,研究實驗並不支持過去研究者認. 13.
(14) 國立臺灣體育學院學報第十四•十五期. 為擺臂的效應可以提昇蹬地力量的說法。也由於擺臂動作可以增進下肢有順序 的動力鏈形式,使得完成垂直跳工作的肌肉產生協調配合的共同作用效應,致 使蹠屈肌肉有效率地產生蹬地作用。當然,此一說法僅為研究者根據動力學與 肌肉電訊號的訊息提出之定性解釋,後續研究仍須就此肌肉協調配合的機制進 行正過程模擬分析。此外,關於性別差異問題,擺臂對於騰空高度的效應並無 統計的顯著差異,且正衝量的作用力時間也無性別差異存在,顯示若是排除肌 力問題,擺臂垂直跳的技術在本研究結果則無性別差異的情況。. 參考文獻 劉宇(1998)。生物力學原理。於許樹淵編,運動力學(pp. 69-78)。臺北市:中華 民國體育學會。. Bobbert, M. F., & van Ingen Schenau, G. J. (1988). Coordination of vertical jumping. Journal of Biomechanics, 21, 241-262. Boudolos, K. D. (1998). Jumping performance of young girls in track and field. In H. J. Riehle, & M. M. Vieten (Eds)., The Proceedings II of XVI International Symposium on Biomechanics in Sports (pp. 116-119). Konstanz, Germany: Universitätsverlag Konstanz. Dapena, J. (1999). A biomechanical explanation of the effect of arm actions on the vertical velocity of a standing vertical jump. Book of Abstracts for International Society of Biomechanics XVIIth Congress (p. 100). Calgary, Canada: 1999 XVIIth Congress of the International Society of Biomechanics. Enoka, R. M. (2002). Neuromechanics of human movement(3rd ed.). Champaign, IL: Human Kinetics. Hall, S. J. (2003). Basic biomechanics (4th ed.). New York: McGraw Hill.. 14.
(15) ˙以獲取運動速度的最大衡量原理探討垂直跳動作的擺臂作用效果. Harman, E. A., Rosenstein, M. T., Frykman, P. N., & Rosenstein, R. M. (1990). The effects of arms and countermovement on vertical jumping. Medicine and Science in Sports and Exercise, 22(6), 825-833. Harrison, A. J., & Gaffney, S. (2001). Motor development and gender effects on stretch-shortening cycle performance. Journal of Science and Medicine in Sport, 4(4), 406-415. Hay, J. G. (1993). The biomechanics of sports techniques (4th ed.). Englewood Cliffs, NJ: Prentice Hall. Kibele, A. (1998). Possibilities and limitations in the biomechanical analysis of counter-movement jump: A methodological study. Journal of Applied Biomechanics, 14, 105-117. Miller, D. I., Henning, E., Pizzimenti, M. A., Jones, I. C., & Nelson, R. C. (1989). Kinetic and kinematic characteristics of 10-m platform performances of elite divers: 1. Back takeoffs. International Journal of Sport Biomechanics, 5, 60-88. Payne, A. H., Slater, W. J., & Telford, T. (1968). The use of a force platform in the study of athletic activities. Ergonomics, 11, 123-143. van Ingen Schenau, G. J. (1989). From rotation to translation: Constraints on muti-joint movements and the unique action of bi-articular muscles. Human Movement Science, 8, 301-337. Winter, D. A. (1990). Biomechanics and motor control of human movement. New York: John Wiley & Sons. Zajac, F. E. (1993). Muscle coordination of movement: A perspective. Journal of Biomechanics, 26, 109-124.. 15.
(16)
(17)
數據

相關文件
Speakers on a team must work together effectively to present arguments and examples that support the team’s stance, and rebut the opposing team’s arguments to the greatest
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
Monopolies in synchronous distributed systems (Peleg 1998; Peleg
This kind of algorithm has also been a powerful tool for solving many other optimization problems, including symmetric cone complementarity problems [15, 16, 20–22], symmetric
Corollary 13.3. For, if C is simple and lies in D, the function f is analytic at each point interior to and on C; so we apply the Cauchy-Goursat theorem directly. On the other hand,
Corollary 13.3. For, if C is simple and lies in D, the function f is analytic at each point interior to and on C; so we apply the Cauchy-Goursat theorem directly. On the other hand,
õT¤_ .â·<íËju, Data Access Component Module 2FíŠ?. âÀÓ“, ©ø_ method úk’eé query v,
