晝夜節律基因在肺鱗狀上皮細胞癌的紊亂表現; Perturbed expression of the circadian genes in squamous cell lung carcinoma
49
0
0
全文
(2) PER1 和/或 CRY1 promoter 甲基化,但是在 29 個非癌組織中, 。 只有 6 個樣本有 PER1 或 CRY1 promoter 異常甲基化(P<0.05) 而免疫組織染色的結果則發現晝夜節律基因蛋白在癌組織中呈 現出紊亂的表現方式,顯示出在肺鱗狀上皮細胞癌細胞中,晝夜 節律生物時鐘是以非規律且不協調的方式運作。總結來說,本研 究結果顯示出在肺鱗狀上皮細胞癌中的晝夜節律基因的表現是 呈現出失調的現象,而本現象有部分是因晝夜節律基因中的. PER1 及 CRY1 的 promoter 中發生異常甲基化,這個結果促使肺 鱗狀上皮細胞癌的晝夜節律依照自己的版本運作,表現出異於正 常細胞的晝夜節奏。本研究應可為肺鱗狀上皮細胞癌的化學時間 治療法(Chronotherapy)提供一些分子學的基礎觀點。 關鍵字:晝夜節律基因;肺鱗狀上皮細胞癌;晝夜週期;促進子 甲基化.
(3) ABSTRACT Circadian cycle exerts influences on tumor initiation and progression in many human cancers. In order to clarify the role of the circadian genes in bronchial carcinogenesis, we assessed nine circadian genes including PER1, PER2, PER3, CRY1, CRY2, CLOCK, BMAL1, CKIε, and Timeless in 29 surgically resected squamous cell lung carcinoma and the paired non-cancerous tissues by real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR), methylation-specific polymerase chain reaction (MS-PCR), and immunohistochemical studies. Our results show that the expression levels of all nine circadian genes in cancerous tissues were significantly reduced compared with the paired non-cancerous tissues (p< 0.00001). Methylation of CpG islands was found in the promoter of PER1 (5 of 29 cancerous tissues and 1 of 29 non-cancerous tissues) and CRY1 (16 of 29 cancerous tissues and 5 of 29 non-cancerous tissues; p<0.05), but not in the promoters of other 7 circadian genes in the paired squamous cell lung cancer and normal tissues that we had investigated. In total, 18 of the 29 cancerous tissues showed CpG methylation in the promoter of PER1.
(4) and/or CRY1, but 6 of the 29 paired non-cancerous tissues revealed methylation of either PER1 or CRY1 promoter (p<0.05). Immunohistochemical staining revealed that the circadian gene protein patterns were perturbed in cancerous tissues indicating asynchronous operation of the circadian clock in squamous cell lung carcinoma. In conclusion, our results suggest that dysregulation of expression of the circadian genes in squamous cell lung cancer was partly due to the inactivation of the PER1 and/or CRY1 genes by promoter hypermethylation, which induce serious of effects on cancer-version circadian controlled proteins expression accordingly. Differential expression of circadian genes in cancerous and non-cancerous cells may provide a molecular foundation for chronotherapy of squamous cell lung cancer. Keywords: circadian genes; squamous cell lung carcinoma; circadian cycle; DNA hypermethylation..
(5) 論文目次 第一章 緒論………………………………………… p.1 1-1 日夜節奏的運作和生理現象………………………1 1-2 晝夜節律基因之表現及調控………………………1 1-3 晝夜節奏失調與癌症形成…………………………3 1-4 肺癌的現況…………………………………………3 1-5 抑癌基因 Promoter 異常甲基化現象與癌症的形成 ………………………………………………………4 1-6 實驗目的……………………………………………6. 第二章 材料與方法…………………………………… 7 2-1 實驗材料……………………………………… 7 2-2 萃取 DNA……………………………………… 7 2-3 萃取 RNA…………………………………………… 8 2-4 同步定量反轉錄聚合酶連鎖反應…………………8 2-5 甲基化特異性聚合酶連鎖反應……………………9 2-6 免疫組織化學研究…………………………………10.
(6) 2-7 統計方法………………………………………… 11. 第三章 結果……………………………………………12 第四章 討論……………………………………………24 附表一 ……………………………………… 32 附表二 ……………………………………… 34 參考文獻………………………………………36.
(7) 第一章 緒論. 1-1 日夜節奏(Circadian rhythm)的運作和生理現象 日夜節奏是指週期大約為 24 小時的節奏。日夜節奏為內生的計 時系統,而且日夜生理時鐘(circadian clock)可依照外在環境 律動的變化而設定並準時運轉[1]。一個準時運轉的日夜生理時 鐘可以確實的使生物體的生理機能,生物化學表現,及行為模式 準確地配合一天 24 小時而運作[2],例如體溫,睡眠節奏,許多 內分泌激素如腎上腺皮質素,生長激素,褪黑激素,尿中鉀離子 濃度等都呈現出日夜節奏變化。 當日夜生理時鐘受干擾使得節奏紊亂,在人類已觀察到會衍生 多種疾病的發展;例如心血管疾病,精神疾患,及癌症[3]。 1-2 晝夜節律基因之表現及調控 在哺乳動物中,日夜生理時鐘的控制中樞位於前下視丘腦的 前視交叉上核(suprachiasmatic nucleus 簡稱 SCN)。此神經 核內的神經元細胞本身可呈現自發性的週期活動,經由神經元細 胞內的分子生理時鐘的運作及調控,準確的配合生活環境的 24 小時日夜變化,並輸出不同的生化訊號至周邊組織;完成生理及. 1.
(8) 行為活動的調控。目前已知有多種晝夜節律基因參與日夜節奏的 運作,包括有 Period 1 (PER1),Period 2 (PER2),Period 3 (PER3),Cryptochrome 1 (CRY1),Cryptochrome 2 (CRY2),. CLOCK,BMAL1,Casein Kinase Iε(CKIε),及 Timeless (TIM)[4]。這群基因經由正向及負向的迴饋系統交互運作,完成 約 24 小時的日夜節奏運行[5]。主要正向迴饋機制包含兩個 helix-loop-helix 的轉錄因子 CLOCK 及 BMAL 1[6],當這兩個轉 因子偶合在一起,經由作用於 CACGTG 促處進區(E-box),可趨 使 3 種 Period 基因(Per 1,Per 2,Per 3)及 2 種 Cryptochrome 基因(Cry 1,Cry 2)基因表現[2,4,5,6],經轉錄成 m-RNA 並 轉譯為蛋白於細胞質內,在細胞質中 PER 蛋白及 CRY 蛋白則作為 負迴饋機制的要角,經由 PER 及 CRY 蛋白與蛋白間的作用, PER/CRY 複合體可自細胞質轉位至細胞核中,同樣地,也是經由 CACGTG 促進區,PER/CRY 複合體可關閉 CLOCK-BMAL 1 的轉錄作 用,使 CRY 及 PER 蛋白產量減少。除了 PER/CRY 複合體的蛋白間 作用之外,PER 蛋白為 Casein KinaseIε所磷酸化的程度及 PER 與 TIMELESS 之間的交互作用,亦會影響 PER 位於細胞質或細胞 核的能力而改變負向迴饋的作用[1,2,7,8]。 生物時鐘調控基因不僅位於 SCN 中,在週邊組織細胞,甚至 2.
(9) 於不朽細胞內(immortalized cells),亦可發現他們的運作。在 哺乳動物的組織培養細胞中,我們確實可發現日夜節奏的表現, 只是其節奏表現在數個細胞週期後便會鈍化掉,需要不時加入 SCN 訊號來維持其日夜節奏的表現,也就是說,SCN 對週邊組織 的日夜奏有其重要的控制角色[9,10,11]。 1-3 晝夜節奏失調與癌症形成 日夜節奏生理時鐘可調控許多的生物節奏律動,在正常的人 類生理方面,其可控制許多生理日夜表現,例如血壓,睡眠清醒 週期,荷爾蒙產量,體溫,免疫系統,消化道分泌,及肝臟解毒 系統等作用[12]。除此之外,不規律的日夜週期則會促使腫瘤的 產生,許多的流行病學的研究指出一些需日夜顛倒或有時差問題 的工作者,例如空服員或護士,得到乳癌的機率大增[13,14], 而且從動物及人類腫瘤組織的研究也發現組織內生性的日夜節 奏紊亂與癌症的發生及預後有其關聯性[15,16]。 1-4 肺癌的現況 肺癌在全世界的癌症中為致死原因的首位,因為早期診斷困 難及低存活率,肺癌目前已成為健康照顧系統的嚴肅課題 [17,18,19]。近數十年來歐美肺癌病人有顯著的增加,在台灣也. 3.
(10) 是如此,從 1982 年起,癌症死亡成為我國十大死因之首位。根 據衛生署的統計資料顯示過去數年來國人男性癌症死亡原因 中,肺癌僅次於肝癌,為第二位的癌症死亡原因;在女性,肺癌 則持續高居癌症死亡原因的第一位。最近幾年台灣每年有將近 6000 人死於肺癌。肺癌的死亡率極高,整體肺癌患者五年存活 率約為 10%。近年來儘管醫學已有相當進展,肺癌的活療成果還 是非常令人失望。 肺癌可以分為小細胞肺癌及非小細胞癌兩大類,其中後者又 可分為腺癌,鱗狀上皮細胞癌,大細胞癌等類型,在台灣非小細 胞癌佔約 85~88%,其分期簡單分為Ⅰa,Ⅰb,Ⅱa,Ⅱb,Ⅲa, Ⅲb 及Ⅳ期,Ⅰ及Ⅱ屬於局部早期發現,Ⅳ期為末期已轉移至其 他的器官。非小細胞肺癌的治療,第一,第二,第三 A 期以手術 治療為原則,但術後發生轉移或復發的機率甚高,而不能開刀的 第三 B 及第四期病例對化學藥物及放射線治療大多不敏感,目前 一些新的化學治療藥物或化學治療策略已被提出,例如時間治療 法(Chronotherapy)。 1-5 抑癌基因 Promoter 異常甲基化現象與癌症的形成 我們已知癌症的形成是一種多重步驟的過程,其中包含有多 種致癌基因的堆積組合,任何一種癌症均包含有複雜的基因改變; 4.
(11) 致癌基因的活化及抑癌基因的抑制都參與癌症活化的過程。其中 基因的變異包含有多種過程,例如基因突變(mutation),基因刪 除(deletion),及基因放大(amplification),上述情況都與腫 瘤的形成有關[20]。但是目前已經很清楚地瞭解單單基因異常並 不能代表整個癌化過程,表基因改變(Epigenetic alteration) 亦扮演著重要的角色,DNA 的 Epigenetic alteration 並不會改 變 DNA 的核苷酸序列,但經由核苷酸基的一些修飾過程也會促成 癌細胞形成[21,22]。哺乳動物細胞內,基因的 CpG islands 內 胞嘧啶的甲基化(methylation)與否與基因的表現有很顯著的關 聯性[23],而促進子(Promoter)的甲基化與否可由 Methylction-Specific PCR 來測定,這種技術是先以 Sodium bisulfite 先處理待測之 DNA,Sodium bisulfite 可使非甲基化 之胞嘧啶轉化為尿嘧啶,而甲基化之胞嘧啶仍維持原狀,再以設 計過之 Primer 行 PCR 放大基因片段,就可分辨出甲基化與非甲 基化的 DNA[24]。 CpG islands 通常位於基因的 Promoter 附近,長度大約在 200~1000b.p.富含有 Cytosine 及 Guanine 兩種核苷酸,很多的 報告已指出多種癌症的形成與一些基因(包括與細胞週期有關的. P16Ink4a,P15Ink4b,P14ARF,DNA 修復有關的 hMLH1,BRCA1,及與 5.
(12) apoptosis 有關的 DAPK,TMS1)之 Promoter 附近的 CpG islands 甲基化有關[25,26,27],甚至當病人之癌細胞內的腫瘤抑制基因 (tumor suppressor genes)發生甲基化會有較差的臨床預後 [28]。 1-6 實驗目的 人類肺癌的形成過程與晝夜節律基因表現之間的關係目前 尚未被探討過。在本篇研究,我們使用同步定量 RT-PCR 及 Methylation-Specific PCR 來探究晝夜節律基因在肺鱗狀上皮 細胞癌內的基因表現及 promoter 甲基化的情形,再以免疫組織 化學染色方法來研究其蛋白質表現狀況。. 6.
(13) 第二章 材料與方法. 2-1 實驗材料 29 個於彰化基督教醫院自 29 位肺鱗狀上皮細胞癌患者手術 下來的標本,均包含有癌症部分及鄰近非癌組織,此樣本群均由 組織學証實並經由病理醫師確定診斷。組織樣本取下後均立即冷 凍並眝存於液態氮中直到進行萃取 DNA 和 RNA 為止。 臨床上這 29 個病人中有 2 位女性,27 位男性,年齡分佈為 41 歲到 77 歲,平均年齡為 62 歲。臨床分期為:4 位為第一 B 期,3 位為第二 A 期,7 位為第二 B 期,第三 A 期則有 15 位病人。 2-2 萃取 DNA 病理組織取下後,放入 eppendorf tube 中,加 100ml extraction buffer 及 2μL proteinase K(10μg/ml),在 65 ℃作用 4 小時,加入 40μL potassium acetate,vortex 混合均 勻,置於冰上 1 小時後離心 12000rpm,10 分鐘,4℃,小心地取 上清液至新的 eppendorf tube,加入 0.7 倍體積的 isopropanol,緩和的搖晃數下後,離心 12000rpm,10 分鐘 4℃, 倒掉上清液,以 70% alcohol 清洗沈澱物,此清洗步驟重覆三次 晾乾後,加入 20μL 無菌的 ddH2O 溶解,存於 4℃備用[29]。 7.
(14) 2-3 萃取 RNA RNA 萃取為使用 Commercial kit(RNA-BeeTM,Tel-Test,Inc, Texas, USA) ,萃取步驟完成後將其貯存於 pellet 中並冷藏於-70 ℃的環境中備用。 2-4 同步定量反轉錄聚合酶連鎖反應(Real-time quantitative RT-PCR) 利用 9 個晝夜節律基因(PER1,PER2,PER3,CRY1,CRY2,. CLOCR,BMAL1,CKIε和 Timeless)的 mRNA 序列,我們使用 primer Express software(Applied Biosystems,Forster City,CA) 來設計 forward 和 reverse primers 及 TaqMan Probes,TaqMan Probes 合成時並標示以不同的螢光染料(Applied Biosystems)。forward primers,reverse primers 及 probes 的序列,均表列於附表 1 中。 在 Real-time qRT-PCR 的實驗裡,我們是以 glyceraldehydes-3-phosphate dehydrogenase(GAPDH)基因的 表現量來作為內部控制值(Internel Control),在癌組織及配 對非癌組織中,9 種晝夜節律基因均以 GAPDH 為內部控制值,並 以 PCR 回數閥值(CT)差分別得到相對回數閥值(△CT),接著 再比較癌組織中晝夜節律基因之△CT 和非癌組織中晝夜節律基 8.
(15) 因的△CT 得到△△CT,從而獲得晝夜節律基因於組織間的相對 。 表現量(2-△△CT) 所有的 Real-time qRT-PCR 反應均是在 ABI-7700 的 Sequence detector(Applied Biosystems)中執行,並使用 TaqMan EZ RT-PCR core kit(Applied Biosystems);在 25μL 的終容積中含有 40nM 的 primer,200nM 的 probe,300μM 的 deoxynucleoside triphosphate(dNTP) ,3mM 的乙酸錳,2.5U rTth DNA polymerase 和 1 倍的 PCR 緩衝液,整個 PCR 反應回數 設定為反轉錄反應,50℃,2 分鐘,接著為 60℃,30 分鐘,再 來是 95℃,5 分鐘,再行以 40 回的 PCR 反應(設定為 94℃,20 秒,62℃,1 分鐘)。 2-5 甲基化特異性聚合酶連鎖反應(Methylation-specific PCR,MS-PCR) 所有樣本的萃取 DNA 均先以 Sodium bisulfite 處理過,作 MS-PCR 所需的甲基化特異性及非甲基化特異性的 Primers 序列 列於附表 2,執行 MS-PCR 的方法如下:4μg 的 DNA 於 40μL 的 H2O 中並以 10μL 的 1M NaOH 在 37℃中,10 分鐘,進行 denature, 接著於 10mM hydroquinone 30μL 和 520μL 的 1.5M Sodium bisulfite 在 50℃下放置 16 小時進行修飾(Modification)。修 9.
(16) 飾後的 DNA 樣本置於 Wizard DNA Purification kit(Promega, Madison,WI,USA)並以 100μL H2O(預溫至 65-70℃)稀釋, 稀釋液中再加入 50μL 的 1M NaOH 並於室溫中置放 5 分鐘,接著 以 150μL 100% isopropanol 進行沈澱,並以 70% ethanol 清 洗後,再加入 45μL 的 H2O 中。此製備液接著行 PCR 反應,以製 備液加入 1 倍的 PCR 緩衝液,1m MgCl2,100μg primer,0.2mM dNTP,2.5U Taq Polymerase,於 Thermal cycler 上跑 35 回 PCR 反應,每一回包括有 94℃,1 分鐘行 denature,60℃,1 分鐘行 methylated 或 unmethylated primers anneal,72℃,1 分鐘行 extension,72℃,5 分鐘行 final extension。35 回後,PCR 產物置於 3.5%瓊膠上泳反應,並以 ethidium bromide 染色,並 在 UV 光下進行判讀。 CpG methylase(SssI)可以使所有的 Cytosine 甲基化,所 以拿來當作甲基化特異性 primer 的 positive control,從正常 人抽血的檢體因為在晝夜節律因中沒有異常甲基化,可作為非甲 基化特異性 primer 的 positive control,沒有經過 sodium bisulfite 處理過的組織樣本(包含癌組織及非癌組織)則作為 本反應的 Negative Control 組。 2-6 免疫組織化學研究(Immunohistochemistry study) 10.
(17) 先將置於 poly-1-lysine 包圍的 slide 中石蠟包埋的檢體行 脫臘步驟,玻片置於甲醛中及 3% H2O2 後,以階段性酒精及 PBS 來 re-hydrate,再置放於 10mM citrate buffer 中,並於 PBS 中加熱至 100℃,20 分鐘,加入晝夜節律基因蛋白(PER1,PER2, PER3,CRY1,CRY2,CLOCK,BMAL1,CKIε及 TIMELESS)的特殊 抗體(Santa Cruz Biotechnology,CA,USA)後於室溫置放 20 分鐘,再將玻片取出,以 PBS 仔細的清洗 3 次後,置入 HRP/Fab Polymer 中 30 分鐘,經此步驟後,我們可以 3, 3’-diamino-benzidine tetrahydrochloride 來觀察組織玻片 中 peroxidase 的 activity,另外再以 Hematoxylin 染色當作 Counterstain,完成這些步驟後,加阿拉伯膠封片,以斜角 45 度放上蓋玻片,於室溫待膠乾後,以顯微鏡觀察並拍照紀錄。 2-7 統計方法 以 Microsoft Excel 2000 進行資料建檔,SPSS for windows Release 9.0(SPSS,Chicago,IL)進行統計分析,以平均值及 標準差,描述癌組織及非癌組織之間,Promoter 甲基化個數的 差異,並以 t Test 檢定, P 值小於 0.05 為顯著性差異。. 11.
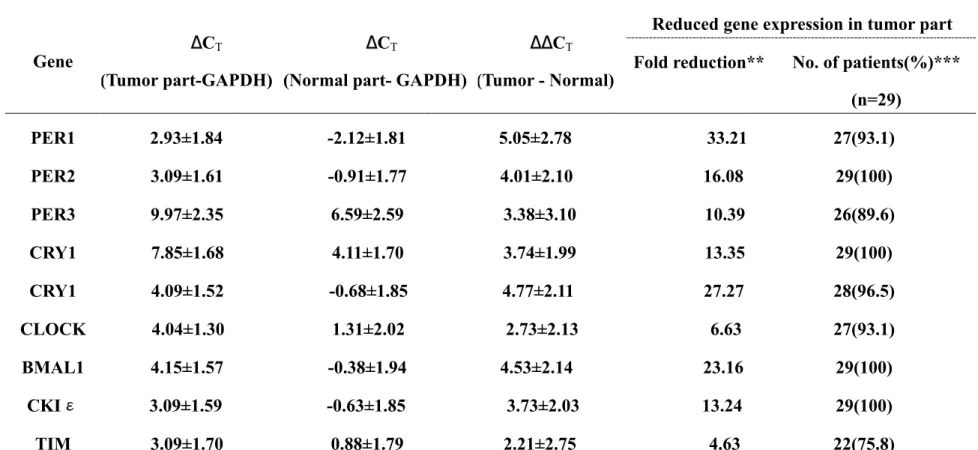
(18) 第三章 結果. 3-1 Determination of the mRNA levels of the nine circadian genes by real-time quantitative RT-PCR 為了瞭解在肺鱗狀上皮細胞癌組織中,九種晝夜節律基因的 轉錄狀況,我們是以定量同步 RT-PCR(Real time quantitative RT-PCR)的方法分別對 29 個癌組織及其配對非癌組織作 mRNA 量的測定。並且我們也以同樣的方法測量 GAPDH 的 mRNA 量來作 為內在的參考值(endogenous reference),並以其作為內部控制 值(internal control)來比較各晝夜節律基因的 mRNA 的量。以 比較 PCR 回數閥值差(ΔCT)的方式分別施行於癌組織及其配對 非癌組織,此兩者之間的差異(ΔΔCT)就可以決定這 29 組樣 本組織間的關係。表 3 綜合了以上的結果顯示出 29 個肺鱗狀上 皮細胞癌組織的晝夜節律基因 mRNA 的量均呈現表現低下的結 果。 我們觀察到在與配對的非癌組織比較之後,肺鱗狀上皮細胞 癌的九種晝夜節律基因 mRNA 表現的平均相對量均呈現低下的結 果,在分析 mRNA 相對表現量(2-ΔΔCT)後,發現其差異從 TIM 基. 12.
(19) 因的 4.63 倍到 PER1 基因的 33.21 倍之範圍(統計 P 值小於 105) , 且大部份的晝夜節律基因在癌組織的平均表現水準都比非癌組 織低十倍以上。以各別組織來分析的話,在 29 對組織樣本中, 大部分的癌組織其晝夜節律基因的 mRNA 表現也是呈現低下的狀 態(見表 3) ,除了 TIM 基因的 22/29 個樣本(佔 75.8%)外,其 他的畫夜節律基因 mRNA 量於 29 個癌組織樣本中大部分均是呈現 低下的現象(100%到 89.6%之間) 。其中有四種基因(PER2,CRY1,. BMAL1,CKIε)在所有癌組織中(100%)均呈現表現低下的結果, 總結來看,real time quantitative RT-PCR 的結果顯示在 29 個肺鱗狀上皮細胞癌樣本中,9 種晝夜節律基因的 mRNA 量在癌 組織中均低於其配對的非癌組織。 (見表 3). 13.
(20) 表 3.肺鱗狀上皮細胞癌組織中晝夜節律基因 mRNA 量的表現 ________________________________________________________________________________________________________. Gene. ∆CT. ∆CT. ∆∆CT. (Tumor part-GAPDH) (Normal part- GAPDH) (Tumor - Normal). Reduced gene expression in tumor part Fold reduction**. No. of patients(%)*** (n=29). PER1. 2.93±1.84. -2.12±1.81. 5.05±2.78. 33.21. 27(93.1). PER2. 3.09±1.61. -0.91±1.77. 4.01±2.10. 16.08. 29(100). PER3. 9.97±2.35. 6.59±2.59. 3.38±3.10. 10.39. 26(89.6). CRY1. 7.85±1.68. 4.11±1.70. 3.74±1.99. 13.35. 29(100). CRY1. 4.09±1.52. -0.68±1.85. 4.77±2.11. 27.27. 28(96.5). CLOCK. 4.04±1.30. 1.31±2.02. 2.73±2.13. 6.63. 27(93.1). BMAL1. 4.15±1.57. -0.38±1.94. 23.16. 29(100). CKΙε. 3.09±1.59. -0.63±1.85. 3.73±2.03. 13.24. 29(100). TIM. 3.09±1.70. 0.88±1.79. 2.21±2.75. 4.63. 22(75.8). 4.53±2.14. * ∆CT and ∆CT values were calculated after normalization with the GAPDH expression levels; ∆∆CT values were calculated as the difference between the ∆CT values of the paired cancer and normal tissues. All results shown are mean±SE values. ** P (t test) <0.00001 in all cases. ***Other patients in each gene analysis group analyzed showed reduced gene expression in the normal tissues.. - 14 -.
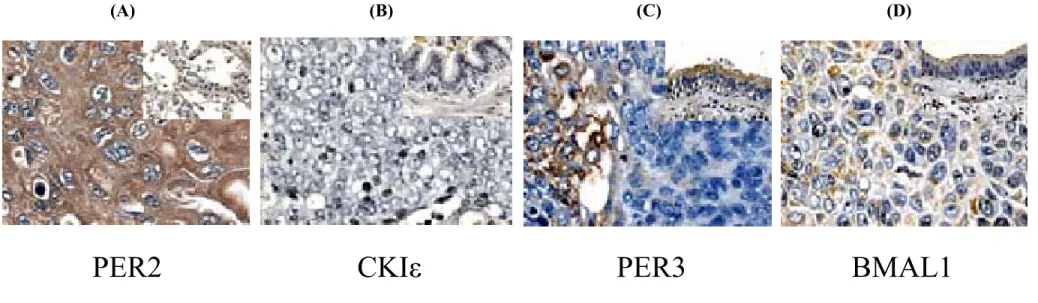
(21) 3-2 Immunohistochemical analysis of the circadian gene proteins 我們以免疫組織化學染色的方式來觀察 9 種晝夜節律基因 蛋白在 29 個癌組織及配對非癌組織的表現形態,我們發現在癌 組織中,這九種晝夜節侓基因蛋白的分佈狀況為異質性及均質 性並存(圖 1 及圖 2)。但在非癌組織中,大部分的畫夜節律基 因蛋白則是呈現均質性染色表現。在 29 個癌組織樣本中有一個 癌組織樣本為 9 種晝夜節律基因蛋白均為陽性染色且混雜著異 質及均質染色分佈狀態,有 2 個癌組織樣本則是 9 種晝夜節律 基因蛋白均呈現陰性染色反應,其他 26 個癌組織樣本則是部分 晝夜節律基因蛋白抗體染色為陽性,其他的則是陰性染色反 應,且多數為異質性染色分佈狀態(圖 3a) ,另一方面來看, 如果以晝夜節律基因蛋白的種類來看其於各癌組織樣本的陽性 染色反應率來看,我們得到以下結果: PER1, 24/29 (82.7%); PER2, 21/29 (72.4%); PER3, 24/29 (82.7%); CRY1, 13/29 (44.8%); CRY2, 9/29 (31%); CLOCK,15/29 (51.7%); BMAL1, 14/29 (48.2%); CKIε,7/29 (24.1%) and TIM, 26/29 (89.6%) (圖 3b) 。從這些結果看起來,肺鱗狀上皮細胞癌組織中的癌 細胞群內存在著不的日夜節律表現,也就是說,癌組織中的癌 細胞群有著不同步調的畫夜節奏調控方式。 - 15 -.
(22) (A). (B). PER2. CKIε. (C). (D). PER3. BMAL1. 圖 1.免疫組織化學染色結果,圖中為癌細胞染色結果,右上角為同一樣本之非癌部分。(A) 以 PER2 抗體行免疫組織化學染色,癌細 胞中呈現陽性且均質性分布。(B) 以 CKIε抗體行免疫組織化學染色,癌細胞中呈現完全陰性反應。(C) 以 PER3 抗體行免疫組 織化學染色,癌細胞中呈現部份陽性染色(左側)及部份陰性染色反應(右側)。(D) 以 BMAL1 抗體行免疫組織化學染色,癌細胞中 呈現陽性且異質性分布,一些癌細胞為弱陽性染色,另一些癌細胞為強陽性染色。400 倍放大(右上角為 100 倍放大). - 16 -.
(23) PER1. PER2. PER3. CRY1. CRY2. CLOCK. BMAL1. CKIε. TIM. (A). (B). (C). 圖 2.以九種晝夜節律基因蛋白抗體行免疫組織化學染色,圖中為癌細胞染色結果,右上角為同一樣本之非癌部分。(A) 選取的代表案 例為 PER1,PER2,PER3,CRY1,CLOCK 及 TIM 抗體呈現陽性且異質性染色。CRY2,CKIε為弱陽性。BMAL1 則呈陰性反 應。(B) 選取的代表案例為 CRY1 抗體呈現陽性且均質性染色。PER1,PER2,PER3,BMAL1,CKIε及 TIM 抗體呈現陽性且 異質性染色。CRY2 及 CLOCK 則呈陰性反應。(C) 選取的代表案例為所有九種晝夜節律基因蛋白抗體均呈現陰性染色反應。400 倍放大(右上角為 100 倍放大). - 17 -.
(24) Figure 3(a). heterogeneus*. homogeneous. No. of tissue. 30 20 10 0. PER1. PER2. PER3. CRY1. CRY2. CLOCK. BMAL1. CkIε. TIM. homogeneous. 6. 4. 1. 2. 0. 1. 1. 1. 3. heterogeneus*. 18. 17. 23. 11. 9. 14. 13. 6. 23. * p< 0.05 Figure 3(b). No. of tissue. 8 6 4 2 0 negative. one. two. three. four. five. six. seven. eight. nine. No. of CCG protein expression. 圖 3.以免疫組織染色法來分析九種晝夜節律基因蛋白於 29 個肺鱗狀上皮細胞癌組 織中的染色表現。條狀圖為陽性染色反應。(a)九種晝夜節律基因蛋白呈陽性 染色的頻率,大部分的癌組織為異質性呈色方式(P<0.05) (b)每個癌組織樣 本中,晝夜節律基因蛋白陽性染色的個數. - 18 -.
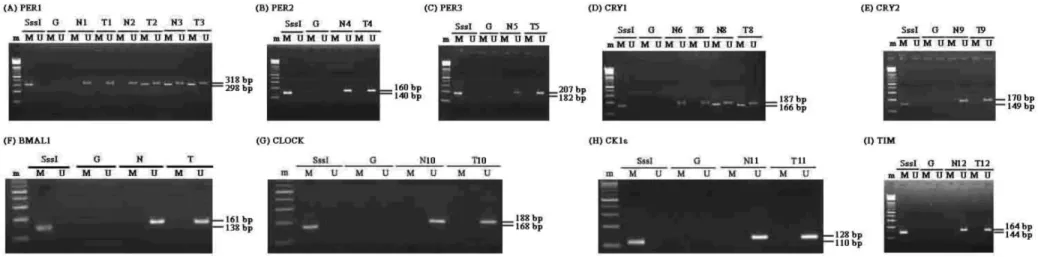
(25) 3-3 Circadian genes methylation in human squamous cell lung carcinoma 為了瞭解晝夜節律基因在癌組織中的 mRNA 的表現遠比非 癌組織較為低下的原因,我們考慮到是否在癌組織中晝夜節律 基因促進子的 CpG island 發生甲基化現象,使得該基因的轉錄 受到影響。我們以 MS-PCR 的方法來測定這 29 個肺鱗狀上皮細 胞癌樣本包括癌組織及非癌組織中的九種晝夜節律基因的 promoter 中 CpG island 的甲基化狀況,我們設計了甲基化及 非甲基化特異性 primers(Methylation- and unmethylation-Specific Primers)(見表 2),並以 PCR 的方 法來放大及分辨出各晝夜節律基因促進子的甲基化及非甲基化 狀態(見圖 4) 29 組肺鱗狀上皮細胞癌組織及其配對非癌組織的 MS-PCR 實驗結果顯出這 29 組樣本中,有 7 種晝夜節律基因(PER2,. PER3,CRY2,CLOCK,BMAL1,CKIε,TIM)在癌組織及配對非 癌組織中均沒有發現 promoter 中有甲基化的現象。有一組(包 括癌及非癌組織)樣本存在 PER1 promoter 甲基化(3.45%)。有 5 個樣本(17.24%)只有在癌組織發現 PER1 promoter 甲基化。 ,其癌及非 而在 CRY1 promoter 序列中,有 5 組樣本(17.24%) - 19 -.
(26) 癌組織均有 CRY1 promoter 甲基化現象。16 個(55.17%)呈現 出只有在癌組織中有 CRY1 promoter 甲基化存在。MS-PCR 的結 果見表 4。綜合來看,在 29 個癌組織樣本中,有 18 個存在著. PER1 或 CRY1 promoter 甲基化,其中 2 個為單一 PER1 promoter 甲基化,13 個為單一 CRY1 promoter 甲基化,其他 3 個則同時 存在 PER1 和 CRY1 promoter 甲基化。在 29 個配對非癌組織中, 則有 6 個存在著 PER1 或是 CRY1 promoter 甲基化,其中有 1 個是 PER1,另外 5 個則是 CRY1,而且這 6 個非癌組織樣本的配 對癌組織中也有同樣的 PER1 及 CRY1 promoter 甲基化發生。 表 4.肺鱗狀上皮細胞癌組織及配對非癌組織中晝夜節律基因 MS-PCR 的分析結果 DNA methylation. tissue. 2 genes Per1 + Cry1. 1 gene Per1 Cry1. Total. Cancerous (n=29). 3. 2. 13. 18*. Paired non-cancerous (n=29). 0. 1. 5. 6. *p<0.05. - 20 -.
(27) 圖 4.九種晝夜節律基因於肺上皮細胞癌中的甲基化分析。圖中的代表樣本為 MS-PCR 於 PER1(A)、PER2(B)、PER3(C)、CRY1 (D) 、CRY2(E) 、BMAL1(F)、CLOCK(G) 、CKIε(H) 、TIM(I)的結果。平行的放大反應分別以特殊的 Primer 作用於非 甲基化(U)及甲基化(M)之 DNA。m:100 b.p. ladder Markers; N:配對非癌組織;T:癌組織;SssI:CpG Methylase 作用後 之 DNA;G:未經 Sodium Bisulfite 修飾之 DNA。. - 21 -.
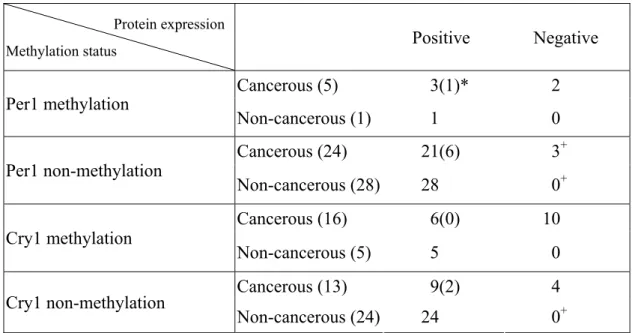
(28) 3-4 CpG methylation and expression of CRY1 and PER1 in squamous cell lung carcinoma 我們進一步的以前述的 MS-PCR 和蛋白質表現的結果來交 叉分析是否 promoter 甲基化會影響到基因的蛋白表現方式,我 們將焦點放在 PER1 和 CRY1 這兩個較常發生 promoter 甲基化的 基因進行比較。在 16 個 CRY1 promoter 存在著甲基化之癌組織 樣本中,有 10 個(62.5%) ,其免疫組織染色是呈現陰性反應, 而且其他 6 個陽性染色樣本則是異質性染色分佈狀態。另外, 13 個沒有 CRY1 promoter 甲基化的癌組織中,則只有 4 個(30%) 呈現 CRY 抗體陰性染色反應,另外 9 個陽性反應的組織中,有 7 個為異質性,2 個為均質性的染色分佈狀態。而 5 個存在著. CRY1 promoter 甲基化的非癌組織中,其 CRY1 抗體則均為陽性 染色且以異質分佈方式呈現。在 5 個存在著 PER1 promoter 甲 基化的癌組織中,有 2 個的 PER1 抗體染色為陰性反應,其他 3 個陽性染色反應的樣本,有 2 個為異質性,1 個為均質性蛋白 分佈方式,另外 24 個 PER1 promoter 無甲基化的癌組織中,有 3 個(12.5%)呈現 PER1 抗體陰性染色反應,15 個為陽性異質 性染色,6 個則為陽性同性質染色方式。由這些結果顯示,在 肺鱗狀上皮細胞癌組織中晝夜節律基因 promoter 甲基化的確 - 22 -.
(29) 會影到該基因蛋白的表現,從而對整個日夜節奏調控產生不正 常影響。. 表 5.肺鱗狀上皮細胞癌組織及配對非組織中 PER1 及 CRY1 Promoter 甲基化及其蛋 白表現的相互關聯 Protein expression. Positive. Methylation status. Per1 methylation. Per1 non-methylation. Cry1 methylation Cry1 non-methylation. Negative. Cancerous (5). 3(1)*. 2. Non-cancerous (1). 1. 0. Cancerous (24). 21(6). 3+. Non-cancerous (28). 28. 0+. Cancerous (16). 6(0). Non-cancerous (5). 5. 0. Cancerous (13). 9(2). 4. Non-cancerous (24). 24. * Number of positive staining (number of homogeneous presentation) + p< 0.05. - 23 -. 10. 0+.
(30) 第四章 討論. 於本研究中,我們發現所有的鱗狀上皮細胞肺癌組織的 9 種晝夜節律基因的 mRNA 的量均比鄰近正常組織的量為低。為了 瞭解造成此種不正常表現的原因,我們對其中五個樣本的 9 種 晝夜節律基因的基因碼進行定序,但是,在這些肺鱗狀上皮細 胞癌組織中,我們並沒有發現任何一種畫夜節律基因之序列有 特殊的異常,此現象表示基因序列突變並不是這些日夜節奏基 因不正常表現的原因。我們已知在促進子(Promoter)附近的 CpG islands 上的甲基化會抑制 promoter 的功能,進而抑制基 因之表現,因此,我們開始探討是否在肺鱗狀上皮細胞癌組織 中,promoter 的甲基化是促成日夜節奏調控基因 mRNA 低下的 因素。promoter 附近的 CpG islands 去甲基化可使此段基因開 始進行轉錄工作,相反的,在不朽細胞及轉型細胞(transformed cells)中則常可發現正常非甲基化的 CpG islands 發生甲基化 [30]。Virmani 等人已發現在肺癌細胞中,經常有 retinoid acid receptorβ-2 表現低下的現象,並証實是經由 CpG islands 甲 基化的結果[31],其他與肺癌有關聯的 promoter 甲基化的基因 包括有多種細胞表面醣蛋白 E-Cadherin 和 H-Cadherin, - 24 -.
(31) Cyclin-dependent protein kinase inhibitors, P16Ink4a 和. P15Ink4b,death-associated Protein kinase(DAPK)以及 adenomatous polyposis coli gene(APC)等等[32,33,34]。 在本研究中,為了探討日夜節奏基因的不正常表現的原因,我 們使用 Methylation Specific PCR 的方法並設計出與 promoter 甲基化和與非基化片段配對的 Primers。我們發現在鱗狀上皮 細胞肺癌組織中,晝夜節律基因之 promoter 的甲基化與其 mRNA 異常表現有所關聯。我們發現在 29 個癌組織中有 5 個組織其. Per1 promoter 甲基化(佔 17.2%) ,16 個組織其 Cry1 有 promoter 甲基化現象(佔 55.2%) ,此結果顯示有約半數的肺鱗狀上皮細 胞癌,其晝夜節律基因的表現低下的原因與其 promoter 的甲基 化有關。另外,我們也發現在 29 個配對非癌組織中也可發現 1 個組織有 Per1,5 個組織其有 Cry1 promoter 甲基化現象,但 是這 6 個組織均存在有其鄰近癌組織同樣的 Per1 及 Cry1 promoter 甲基化,此結果顯示在某些肺鱗狀上皮細胞癌案例, 即使鄰近癌的組織為病理學確定為正常細胞,其分子生物時鐘 其實已有所變異。再者,在 13 個癌組織其 Cry1 promoter 並沒 有甲基化現象中有 4 個(佔 30%) ,24 個癌組織其 PER1 promoter 並沒有甲基化現象中有 3 個(佔 12.5%)在免疫組織染色後, - 25 -.
(32) 並沒有相對應的蛋白質表現,顯示出應該還有其它未發現的機 轉控制這兩個晝夜節律基因的基因表現。 我們是以免疫組織化學染色的方法來研究這九種晝夜節律 基因的蛋白質表現。我們發現在所有 29 個肺鱗狀上皮細胞癌組 織的晝夜節律基因的蛋白質表現方式與其相配對的非癌組織均 完全不同。在癌組織中的晝夜節律基因蛋白質的呈現方式為多 樣化的,且各樣本間的差異性頗大,在這裡面有兩個癌組織其 九種晝夜節律基因蛋白的組織免疫抗體染色完全呈現陰性反 應,而其他 27 個癌組織則呈現出只有數種晝夜節律基因蛋白抗 體呈陽性反應,且其染色分佈也混雜著均質性及異質性的複雜 表現。而且,在同一個癌組織切片中,我們可以發現不同的晝 夜節律基因蛋白染色的呈現方式包括有不同的晝夜節律基因蛋 白種類及異質性的染色呈色現象。相反的,在正常組織中,我 們發現大部分的免疫組織染色呈現出相似且均質的染色表現。 所以,本研究可顯示在肺鱗狀上皮細胞癌中,其晝夜節律的表 現與正常非癌組織相比較是大不相同的。再者,我們也發現在 同一個癌樣本中,不同的癌細胞群也有不同的晝夜節律基因的 蛋白表現,顯示出肺鱗狀上皮細胞癌中的癌細胞群中也存在著 不同的生物時鐘表現方式。在 16 個 CRY1 promoter 甲基化的樣 - 26 -.
(33) 本中有 10 個(62.5%)其 CRY1 抗體染色呈現陰性,在 5 個 PER1 promoter 甲基化的樣本中有 1 個(20%)其 PER1 抗體染色呈陰 性,這個結果顯示 promoter 甲基化與基因表現受抑制之間有所 關聯,另一方面來看,我們並沒有發現除了 CRY1 及 PER1 以外 的晝夜節律基因的 promoter 有 CpG island 的甲基化現象。所 以,在肺鱗狀上皮細胞癌組織中,其晝夜節律基因的異常表現 及紊亂的蛋白質呈現方式應該與 CRY1 及或 PER1 的基因受抑制 有關。而這兩種晝夜節律基因(PER1 及 CRY1),則關係到整個 日夜節奏運作方式及影響到所有晝夜節律基因相互連鎖的正負 向迴饋機制的交互作用[4]。 在日夜節奏分子生物鐘的運作中,PER1 和 CRY1 均扮演著 重要的角色,哺乳動物的晝夜週期調控機制中,PER 和 CRY 基 因群各有其重要的地位[3,35]。在 PER 基因群中,PER2 蛋白可 作為 BMAL1 基因轉錄的正向調節者,PER1 咸認為其功能為經由 其與它種晝夜節律基因蛋白之間的交互作用,PER1 可以穩定其 他的日夜節律調控蛋白活性。而 PER3 蛋白的功能則是作為日夜 節奏的輸出訊號[36]。而 CRY 基因群則主要作用於負迴饋交互 作用環之中,CRY 蛋白可直接抑制 CLOCK-BMAL1(轉錄因子)媒 介的轉錄作用,且不需經由 PER 和 TIM 蛋白。除此之外,CRY - 27 -.
(34) 蛋白也可與 PER 蛋白形成雙合體或三合體,進而影響 PER 蛋白 在細胞質或細胞核的位置,從而影響晝夜節律基因的轉錄調控 [8,37]。從實驗動物的觀察研究中顯示,實驗老鼠的 PER1 或 CRY1 基因 knockout 的老鼠,將它們置於一個永晝或永夜的環 境中,這些動物的活動週期及運動能力均會失去節奏性 [35,38]。 很多的週邊組織細胞(包括舌上皮,小腸 Crypt 細胞,骨 髓細胞和心肌細胞) ,其細胞增生週期均表現出晝夜節律 [10,39,40,41]。以人類口腔上皮細胞和皮膚上皮細胞為例,晝 夜節律基因的週期表現和細胞分裂週期有關的蛋白(P53, Cyclin-E,Cyclin-A,Cyclin-B1,ki-67 )之間的相關性已有 所發現:hPER1 mRNA 量的表現尖峰約在細胞週期的 G1 期,而 且,hBMAL1 的尖峰期約略與 M 期同時[42]。在 Matsuo 的研究 中,當實驗老鼠行陪份肝切除 72 個小時後,CRY -deficiency 的老鼠其肝臟新生後重量遠低於野生型的老鼠。從這個觀察再 行進一步研究,他們更進一步發現晝夜節律和細胞週期間的關 係:在 CRY -deficiency 的老鼠行部分肝均除後,其肝細胞中 的 WEE1 呈現出穩定上升的狀態,從而抑制 CDC2 的活性,這個 結果使得新生肝細胞細胞週期中的 M 期慢下來[43]。從晝夜節 - 28 -.
(35) 律基因表現的時間點與細胞週期進行的時間點之間相互配合的 關聯,顯示出整個晝夜節律分子生物鐘的運作不僅影響到細胞 週期一期一期的漸進運作,而且晝夜節律基因也會影響到細胞 週期相關基因的表現,這個過程又牽涉到細胞週期的 Checkpoint 和細胞凋亡 (Apoptosis)的控制[44]。這個控制系 統又與整個龐大複雜的交互網路包括有 glucocorticoid,retinoids,cAMP,PKC,WNT,calcium 和 MAPK 訊息傳遞有關[45,46,47,48]。當晝夜節律基因發生突變時會導 致細胞週期過程產生變化而促使細胞癌化產生[44],PER2 基因 本身即可被視為抑癌基因,因為其可以經由影響其他晝夜節律 調控基因而調節部分細胞週期的過程並促成細胞修復。但是, 根據老鼠肝細胞新生的觀察,即使細胞週期被其它因素所抑 制,分子生物鐘仍持續按照節奏進行,顯示出晝夜節律基因可 以獨立行使功能[43]。根據上述的基礎陳述,我們建議在我們 所研究的肺鱗狀上皮細胞癌樣本中,PER1 及 CRY1 的失活會引 致其他的晝夜節律基因失調,進而影響組織細胞的細胞週期過 程,從而使細胞產生癌化的細胞轉型(transformation)。 總結來說,我們己經研究了肺鱗狀上皮細胞癌的九種晝夜 節律基因的表現方式,本研究的結果顯示出肺鱗狀上皮細胞癌 - 29 -.
(36) 的晝夜節律基因呈現出混亂的調節方式,而這種失調現象的原 因很可能是因為 CRY1 和/或 PER1 基因的 promoter 發生甲基化 所造成,抑或是其他未明的晝夜節奏調控基因所導玫的。我們 建議肺鱗狀上皮細胞癌有自己的晝夜節奏,且這種節奏呈現方 式與正常組織的節奏並不相同。再者,晝夜節律基因似平在腫 瘤形成的過程中扮演著重要的角色,晝夜節律基因失調會導致 癌細胞進行並促進腫瘤的生長及擴散。 目前已經有充份的証據顯示在癌細胞和正常細胞之間,其 細胞增生和對藥物代謝效率的時間點存在著差異性[49,50],而 這種差異性就構成了癌症化學治療中的時間治療法 (chronothera- py)的理論基礎。也就是說日夜生物時鐘節奏會 影響細胞動力學(cell kinetics),藥物時間動力學 (chronopharmacokinetics) 和藥物時間耐受性 (chronotolerance)以及藥物時間效率 (chronoefficacy)[51,52]。當時間治療法使用在癌症化學治療 時,這個治療方式可在癌細胞受到抑制的同時,正常細胞組織 受損傷的程度可以降至最小,進而使得癌症治療的效果改善。 基於肺鱗狀上皮細胞癌的晝夜節律基因表現異於正常組織 的結果,本研究或可提供一個分子生物方面的基礎觀點,以使 - 30 -.
(37) 在臨床上對於肺鱗狀上皮細胞癌患者設計出合適的時間化學治 療方式。. - 31 -.
(38) 附表 1.同步定量反轉錄聚合酶連鎖反應中用於分析肺鱗狀上皮細胞癌中九種晝夜節 律基因所用之 Primers 及 Probes. Gene. Amplicon (bp). Tm. PER1. 102. 59. F:. 70. Probe: CGA ATC GTC TAC ATT TC. 59. R:. AGG ACG GCT GCC TGC TC. 57. F:. TCC AGA TAC CTT TAG CCT GAT GA. 70. Probe: CCA TCT ACA AGT GGC TGC. 56. R:. TTT GTG TGT GTC CAC TTT CGA. 58. F:. ACA AAC AGA ACC ACA AGG CA. 68. Probe: CTT ACA AGC TGG TTT GC. 58. R:. CGT CCA TTT GTT GGC ATT T. 58. F:. AGA ACA GAT CCC AAT GGA GAC T. 69. Probe: TAT CAG GCG TTA TTT GCC TGT. 59. R:. GTG CAT TCC AGG GAT CAT AGA. 60. F:. CCT CCT CAG TCG GGA TCA A. 69. Probe: CGA TGG AGG TTC CTA C. 58. R:. GCG GGA GTT CAG TTT CCT TA. 59. F:. CAA CGC ACA CAT AGG CCA T. PER2. PER3. CRY1. CRY2. CLOCK. 98. 94. 91. 86. 107. Primer (5’→3’) CAT CAC GTC TGA GTA CAC ACT TCA GA. - 32 -.
(39) BMAL1. CK1ε. TIM. 97. 110. 91. 70. Probe: CAG GTT AGC TAC ACC TCA G. 59. R:. TTG GGT TCT TCA ACA GTG CA. 58. F:. CAT GCA ACG CAA TGT CCA G. 70. Probe: CTG TGC TAA GGA TGG CT. 58. R:. GTG TAT GGA TTG GTG GCA CCT. 60. F:. AGG TCC TCT GCA AAG GCT ATC C. 70. Probe: CGA ATT CTC AAC ATA CCT CAA. 60. R:. GCG GAA GAG CTG ACG TAG GTA A. 59. F:. CAG CAC CAC CAG GAC AAG C. 68. Probe: CTG ATG GTG AAC TTG ACA C. 60. R:. GCA GAT TGC CAA AAC AGA GCA. - 33 -.
(40) 附表 2.九種晝夜節律基因之 Promoter 甲基化分析所使用的 Primers. Gene Name. Methylation-specific (5’→3’). PER1. ATTTAGGTTTACGTGCGTTC. Amplicon (bp) 298. CGACTCAAAAACGAAAATCG PER2. GCGGTTTCGTTGCGGTTTA. CGGTTTTCGTTCGAGTTCGC. 140. TCGTTTTGTTTTTAGGGGTC. 182. GTTTATTTTCGGTATTTCGC. 166. 149. 168. GTGTTTTTTTATTTTTACGC. 138. AGCGTTGTGATTTTTAGATC CCTACGCGAAAAACGAAACG. TGGGTGGTTGGGTGGGAGTTTTGGGTATTTGT. 207. ATTTTGGGTGGGTTGTTTTGTTTTTAGGGGTT. 187. TTGAGATTTGGTTTATTTTTGGTATTTTGT. 170. GTTGGTTGGTTGTGTTTTTTATTTGATTAGGTTTT. 188. AGTAGAAAATTTATAGTGTTGTGATTTTTAGATT. 161. TCCCCACCTACACAAAAAACAAAACA 110. GTTAGTATTTGTGTGTTTTTTTATTTTTATGT. 128. CCACCCCAAAAAATTTAAAAAACCCA. CGAAAAATTTAAAAAACCCG TIM. 160. CAACTTACCCCATTAAACAACACAAAACCA. AAATACGCGAAATCGCGTCG CK1e. GTGGTGTGGTGTGGTTTTGTTGTGGTTTAT. TACCTTCCACCTAACAATTAACCCAAAAACA. CGTTAAACAACACGAAACCG BMAL1 GGAGGTGTTTGTTTATTCGC. 318. ACACCCACCACAACCACCTAAAAACAACA. TAACGATTAACCCAAAAACG CLOCK GTTTTTTATTCGATTAGGTTTC. TAGTATTAGTATTTAGGTTTATGTGTGTTT. AATCCAACACCAACAACCCAACTAACTAAAACA. GCAACCGCCTAAAAACGACG CRY2. (bp). ACACCCCCACACCAACACCATTTCAAACCA. ACGATTAATCGTCGAAACCG CRY1. Amplicon. AACAACAATCCAACTCAAAAACAAAAATCA. GCCGACGCCGTTTCAAACCG PER3. Unmethylation-specific (5’→3’). 144. GTACTGTTTGAACTCTTTC ACAGAAGAGAGTGAGTGAGG. - 34 -. 164.
(41) 參考文獻. 1. Rensing, L, Meyer-Grahl, U, Ruoff, P. Biology timing and the clock metaphor: oscillatory and hourglass mechanisms. Chronobiology international 2001; 18(3): 329-369. 2. Reppert, S.M., Weaver, D.R. Molecular analysis of mammalian circadian rhythms. Annu Rev Physol 2001; 63: 647-76. 3. Hastings, M.H., Reddy, A.B., Maywood, E.S. A clockwork web: circadian timing in brain and periphery, in health and disease. Nat Rev Neurosci 2003; 4: 649-661. 4. Reppert, S.M., Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002; 418: 935-941. 5. Shearman, L.P., Sriram, S, Weaver, D.R., et al. Interacting molecular loops in the mammalian circadian clock. Science 2000; 288: 1013-1019. 6. Gekakis, N., Staknis, D., Nguyen, H.B.,et al. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998; 280:1564-1569. - 35 -.
(42) 7. Panda, S., Antoch, M.P., Miller, B.H., et al. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell 2002; 109: 307-320. 8. Kume, K., Zklka, M.J., Sriram, S., et al. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 1999; 98: 193-205. 9. Balsalobre, A., Damiola, F., Schibler, U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 1998; 93: 929-937. 10. McNamara, P., Seo, S-B., Rudic, R.D., et al. Regulation of CLOCK and MOP4 by nuclear hormon receptors in the vasculature: ahumoral mechanism to reset a peripheral clock. Cell 2001; 105: 877-889. 11. Zylka, M.J., Shearman, L.P., Weaver, D.R., et al. Three period horologes in mammals: differential light responses in the suprachiasmatic circadian clock and oscillating transcripts outside of brain. Neuron 1998; 20: 1103-1110. 12. Moore-Ede, M.C., Czeisler, C.A., Richardson, G.S. Circadian. - 36 -.
(43) timekeeping in health and disease. N Engl J Med 1983; 309: 469-476. 13. Hansen, J. Light and night, shiftwork, and breast cancer risk. Jounal of the National Cancer Institute 2001; 93: 1513-1515. 14. Rafnsson, V., Tulinius, H., Jonasson, J.R., et al. Risk of breast cancer in female flight attendants: a population-based study (Iceland) Cancer Cause and Control 2001; 12: 95-101. 15. Mormont, M.C., Levi, F. Circadian-system alternations during cancer processes: A review. Int J Cancer 1997; 70: 241-247. 16. Sephton, S., Spiegel, D. circadian disruption in cancer: a neuroendocrine-immune pathway from stress to disease? Brain, Behavior, and Immunity 2003; 17: 321-328. 17. Cancer Statistics. 1999,CA-A Journal for clinicians. January 1999; 49(1): 8-64. 18. American Society of Clinical Oncology: treatment of un-resectable non-small cell lung cancer guideline: update 2003. Journal of Clinical Oncology. 2004, 22, 2: 330-353. 19. Greenwald, H.P., Polissar, N.L., et al. Social factors, treatment, - 37 -.
(44) and survival in early-stage non-small cell lung cancer. Am J Public health 1998; 88(11): 1681-1684. 20. Bishop, J.M. Molecular themes in oncogenesis. Cell 1991; 64: 235-248. 21. Jones, P.A., Baylin, S.B. The fundamental role of epigenetic events in cancer. Nat Rev Genet 2003; 3: 415-428. 22. Baylin, S.B., Esteller, M., Rountree, M.R., et al. Aberrant patterns of DNA methylation, chromatin formation and gene expression in cancer. Hum Mol Genet 2001; 10: 687-692. 23. Baylin, S.B., Herman, J.G. DNA hypermethylation in tumorigenesis. Trends Genet 2000;16: 168-174. 24. Herman, J.G., Graff, J.R., Myohanen, S., et al. Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci USA 1996; 93: 9821-9826. 25. Toyooka, S., Toyooka, K.O., maruyama, R., et al. DNA methylation profiles of lung tumors. Mol Cancer Therapeutics 2001; 1: 61-67. - 38 -.
(45) 26. Esteller, M., Corn, P.G., Baylin, S.B., et al. A gene hypermethylation profile of human cancer. Cancer Res 2001; 61: 3225-3229. 27. Virmani, A.K., Muller, C., Rathi, A., et al. Aberrant methylation during cervical carcinogenesis. Clin Cancer Res 2001; 7: 584-589. 28. Brabender, J., Usadel, H., Danenberg, K.D., et al. Adenomatous polyposis coli gene promoter hypermethylation in non-small cell lung cancer is associated with survival. Oncogene 2001; 20: 3528-3532. 29. Su, T.H., Wang, J.C., Tseng, H.H., et al. Analysis of FHIT transcripts in cervical and endometrial cancers. Int J Cancer 1998; 76: 216-222. 30. Antequera, F., Boyes, J., Bird, A. High levels of de novo methylation and altered chromatin structure at CpG islands in cell lines. Cell 1990; 62: 503-514. 31. Virmani, A.K., Rathi, A., Zochbauer-Muller, S., et al. Promoter methylation and silencing of the retinic acid receptor-beta gene in lung carcinomas. J Natl Cancer Inst 2000; 92: 1303-1307.. - 39 -.
(46) 32. Tsou, J.A., Hagen, J.A., Carpenter, C.L., et al. DNA methylation analysis: a powerful new tool for lung cancer diagnosis. Oncogene 2002; 21: 5450-5461. 33. Tang, X., Khuri, F.R., Lee, J.J., et al. hypermethylation of the death-associated protein (DAP) kinase promoter and aggressiveness in stage I non-small-cell lung cancer. L Natl Cancer Inst 2000; 92: 1511-1516. 34. Zochbauer-Muller, S., Fong K.M., Virmani, A.K., et al. Aberrant promoter methylation of multiple genes in non-small cell lung cancers. Cancer Res 2001; 61: 249-255. 35. Van Der Horst, G.T., Muijtjens, M., Kobayashi, K., et al. Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms, Nature 1999; 398: 627-630. 36. Bae, K., Jin, X., Maywood, E.S., et al. differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron 2001; 30: 525-536. 37. Griffin, E.A., Staknis, D., Weitz, C.J. Light-indepenent role of CRY1 and CRY2 in the mammalian circadian clock. Science 1999;. - 40 -.
(47) 286: 768-771. 38. Bae, K., Weaver, D.R. Light-induced phase shifts in mice lacking mPER1 or mPER2. J. Biol. Rhythms 2003; 16: 123-133. 39. Bjarnason, G.A., Jordon, R.C.K., Sothern, R.B. Circadian variation in the expression of cell-cycle proteins in human oral epithelium. Am J Path 1999; 154: 613-622. 40. Buchi, K.N., Moore, J.G., Hrushesky, W.J.M., et al. circadian rhythm of cellular proliferation in the human rectal mucosa. Gastroenterology 1991; 101: 410-415. 41. Smaaland, R., Laerum, D.D., Lote, K., et al. DNA synthesis in human bone marrow is circadian stage dependent. Blood 1991;77: 2603-2611. 42. Bjarnason, G.A., Jordon, R.C.K., Wood, P.A., et al. Circadian expression of clock genes in human oral mucosa and skinassociation with specific cell-cycle phases. Am J Path 2001; 158: 1793-1801. 43. Matsuo, T., Yamaguchi, S., Mitsui, S., et al. Control mechanism of the circadian clock for timing of cell division in vivo. Science 2003; - 41 -.
(48) 302: 255-259. 44. Fu, L., Pelicano, H., Liu, J., et al. The circadian gene Period2 plays an important role in tumor suppression and DNA damage response in vivo. Cell 2002: 111; 41-50. 45. Canaple, L., Kakizawa, T., Laudet, V. The days and nights of cancer cells. Cancer Res 2003; 63: 7545-7552. 46. Fu, L., Lee, C.C. The circadian clock: pacemaker and tumor suppressor. Nature Reviews 2003; 3: 350-361. 47. Williams, J.A., Su, H., Bernards, A., Field J., et al. A circadian output mediated by NF1 and the Ras/MAPK pathway. Science 2001; 293: 2251-2256. 48. Ko, G. Y-P., Ko, M.L., Dryer, S.E. Circadian regulation od cGMP-gated cationic channels of chick retinal cones: ErK MAP kinase amd Ca/calmodulin-depenent protein kinase II. Neuron 2001; 29: 255-266. 49. Mormont, M.C.& Levi, F. Circadian-system alternations during cancer processes: a review. Int J Cancer 1997; 70: 241-247. 50. Focan, C. Circadian rhythms and cancer chemotherapy. Pharmac - 42 -.
(49) Ther 1995; 67: 1-52. 51. Levi, F. Circadian chronotherapy for human cancers. The Lancet oncology 2001; 2: 307-315. 52. Mormont, M.C. & Levi, F. Cancer chronotherapy: principles, applications, and perspectives. American Cancer Society 2002; 155-169.. - 43 -.
(50)
數據

+2
相關文件
Normalization by the number of reads in the sample, or by calculating a Z score, should be performed on the reported read counts before comparisons among samples. For genes with
6 《中論·觀因緣品》,《佛藏要籍選刊》第 9 冊,上海古籍出版社 1994 年版,第 1
The first row shows the eyespot with white inner ring, black middle ring, and yellow outer ring in Bicyclus anynana.. The second row provides the eyespot with black inner ring
Robinson Crusoe is an Englishman from the 1) t_______ of York in the seventeenth century, the youngest son of a merchant of German origin. This trip is financially successful,
fostering independent application of reading strategies Strategy 7: Provide opportunities for students to track, reflect on, and share their learning progress (destination). •
Malins, “Squamous cell carcinoma arising in the lining of an epidermoid cyst within the sublingual gland—a case report,” British Journal of Oral and Maxillofacial Surgery, vol..
Persons with a phenotype mediated by one of these MC1R genetic variants are at greater risk of UV-induced skin cancers, because pheomelanin not only provides less effective
To evaluate the clinicopathologic features, prognostic factors, and management of patients in the North Chinese population with head and neck squamous cell carcinoma (HNSCC)
