行政院國家科學委員會專題研究計畫 成果報告
SARS 冠狀病毒分離株之繼代培養、生長特性及中和分析與
P3 實驗室之運作
計畫類別: 個別型計畫 計畫編號: NSC93-2751-B-002-002-Y 執行期間: 93 年 07 月 01 日至 94 年 06 月 30 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 高全良 共同主持人: 李君男 計畫參與人員: 王維恭 楊惠茹 陳思齊 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 11 月 24 日
行政院國家科學委員會補助專題研究計畫
█ 成 果 報 告
□期中進度報告
(計畫名稱)
Continuous passage, characterization and neutralization assay of SARS-CoV
isolates and running of P3 laboratory
SARS 冠狀病毒分離株之繼代培養、生長特性及中和分析與 P3 實
驗室之運作
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC
93
-
2751
-
B
-
002
-
Y
執行期間:93 年 7 月 1 日至 94 年 6 月 30 日
計畫主持人:
高全良
共同主持人:
李君男
計畫參與人員: 王維恭、楊惠茹、陳思齊
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立台灣大學醫事技術系
中 華 民 國 94 年 9 月 30 日
中文摘要
SARS(急性嚴重呼吸道症候群) 為 2003 年爆發之新興感染症,包括大陸、香港、越 南、加拿大為主要流行區,台灣也受到明顯的衝擊。國外研究顯示新型冠狀病毒為引起 SARS之重要病原。新加波之研究報告更進一步針對病毒分離株進行全長病毒基因之解碼 發現至少有兩種之基因型之SARS冠狀病毒存在。不同基因型可能與不同地區流行SARS 有關。台灣地區自2003 年 3 月中旬發生首例之SARS病例以來,一共造成三波之流行。 此些不同之流行其病毒是否相同是值得探討之主題。本計畫,針對2003 年之SARS病例, 進行病毒之分離與鑑定。共有10 株病毒被分離成功。由其中選擇TW1 及TW6 兩株病毒做為研究對象。首先將TW1 分別感染不同之細胞株(Vero E6, FRHK-4, MK-2, HEp-2, A549, MDCK, BEAS-2B, Hep-G2, QT, Molt-4, U937, Jurkat),結果發現來自猴子之細胞株 (Vero E6, FRHK-4, MK-2),可以讓SARS-CoV(TW1)生長。其中又以Vero E6,可得最佳 之結果。將TW1 連續在Vero E6 細胞中繼代培養 10 代後,仍然生長良好。病毒產量仍維
持在 1X109,顯示此病株可能可做為疫苗選擇之參考。本計畫亦同時建立一SARS病毒中
和抗體的參考實驗室,以檢測免疫動物之血清及單株抗體中是否存在SARS病毒之中和抗 體。本實驗使用SARS冠狀病毒TW-1 株及猴腎細胞 Vero E6,使用傳統的 96 孔微量滴定 盤TCID50 抑制法,建立病毒中和試驗分析系統。利用此系統,檢測了 13 個SARS病患之 血清,結果發現此13 件檢體,同時對台灣地區前後流行期分離之病毒株(TW1 及TW6), 所產生之中和抗體無明顯之差異,顯示如選擇其中任 1 株病毒,當疫苗所產生之抗體, 應可保護此段期間SARS-CoV之感染。同時為因應疫苗的發展具有時效性評估之需求, 本研究亦同時發展一個快速的中和實驗。結合反轉錄反應與即時聚合脢連鎖反應,用來 偵測在中和反應之判定,初步結果顯示約 8 小時後,即可藉由測定在細胞中病毒所表達 出的正向及反向RNA量以判定是否有中和。此種方法不僅快速,且能減少在顯微鏡下的 觀察時間及減少在第三級實驗室中工作的時間。此快速法也作了初步測試,顯示此為可 行之方法。 本計畫亦提供P3 實驗室之維持。使得台大醫學校區之 SARS 相關研究得以進行。而利用 P3 實驗室,加上所建立之中和實驗。也替來自台大醫學校區、中研院、國衛院及生技產 業等各PI 提供多次之檢測服務。 關鍵詞:嚴重呼吸道症候群、冠狀病毒、病毒分離與鑑定、中和試驗 I
Abstract
By using Vero E6 cell culture, 10 strains of SARS coronavirus (SARS-CoV) were isolated. Continuous passages of two strains (TW1 and TW6) are kept. One strain (TW1) can keep high titer yield of virus after continuous passages in Vero E6 cells for ten times. Among different cell lines (Vero E6, FRHK-4, MK-2, HEp-2, A549, MDCK, BEAS-2B, Hep-G2, QT, Molt-4, U937, Jurkat) tested, the Vero E6 is the most susceptible cell line for growing SARS-CoV.
The traditional micro-neutralization test has been setup and optimized. Convalescent human sera were collected from 13 patients with SARS and examined for their neutralization activities. Results indicated that all of the patient sera possess neutralizing activity against both homologous and heterologous strains of SARS-CoV. The cross neutralization activities of the patient sera to the heterologous SARS viruses were mostly equal or only at a 2-fold difference to the homologous strain.
In this study, we also set up a reference laboratory for screening of the neutralizing activities against SARS-CoV present in the serum of patients, immunized animals, and in monoclonal antibodies and to develop a rapid test for this purpose. In addition to traditional neutralization method, combination of reverse transcription (RT) and real-time PCR is used to detect positive-sense or negative-sense RNA a few hours after the step of neutralization. In order to find the appropriate time to detect the RNA species, the kinetics for positive-sense and negative-sense RNA transcription will be quantitatively analyzed first. SARS-CoV TW-1 strain and Vero E6 cell line are used throughout the experiments. The preliminary data showed that both positive-sense and negative-sense RNA could be detected early after infection. Using RT real-time PCR, neutralization of SARS-CoV could be determined as early as eight hours after inoculation of cells with the virus-serum mixture. This method could reduce the waiting time at least 48 hours as compared to the traditional TCID50 method. In addition to the advantage of rapidity, this method could reduce the observation time under the microscope and the working time in the P3 laboratory. The preliminary data showed this rapid method of neutralization assay can work and practice well.
The P3 laboratory facility was also kept in whole function. It provides all SARS related study in National Taiwan University. This project also help to do neutralization assay of mouse hyper-imimune with Co60 inactivated SARS CoV or SARS VLP and some chemical agents for antiviral activity. A lot of samples requested for neutralization assays were from several PIs from NHRI, Academica Sinica , NTU and other research institutes.
Key words: SARS-CoV, Neutralization test, RT Real time PCR
Content
Introduction 1
Materials and Methods 3
Results 6
Discussion 8
Reference 8
Tables and Figures 10
Introduction
Since November 2002, severe acute respiratory syndrome (SARS) has been spreading in the world for 8 months. China, Hong Kong and Vietnam, Canada are the most affected areas. Taiwan also has been hit by this epidemic. There were 346 probable cases were found in Tawan, 2003. The mortality rate was 10.7% (WHO). New coronavirus (SARS-CoV ) has been recognized as the etiology of SARS. After comparing full length of SARS CoV genome, various genotypes were found circulated in different geographic areas (Ruan et. al, 2003). The first SARS-CoV (TW1) was isolated in our laboratory from the patient with probable SARS in first outbreak in Taiwan. The genomic full length of this virus isolate has been done and filed in GenBank (Huseh et al). After the first episode of SARS in Taiwan, several cluster of outbreak has occurred. By using Vero E6 cell culture, 10 strains of SARS coronavirus (CoV) were isolated in our laboratory from patients with probable SARS during the period from March to May 2003 that covered the first and second stage of SARS outbreak in Taiwan. Four strains were isolated from fatal cases. Most of the strains were isolated from clinical specimens such as sputum, nasopharyngeal aspirates or throat swabs. Only one strain was from urine sample. Among 10 strains, whole genomes of 4 strains were completely sequenced and put on GeneBank. Two strains, TW1 (from NTUH) and TW6 (from Ho-Ping Hosp) were passage in Vero E6 cells for 10 and 5 times respectively. Various genotypes of SARS-CoV were also found in different period of outbreaks in Taiwan (Yeh et al). However, the biological properties of SARS-CoV were still not well characterized. Ng et al has been reported SARS-CoV can grow rapidly in Vero E6 cells. Extracellular SARS-CoV virus particles were presented by 5 hours post inoculation in about 5 % of infected cells. The other properties such as plaque morphology, sensitivities in different growth temperature etc have been not known. Is there any difference of biological properties exited in these different clinical isolates? And are the biological properties of parent strains still kept in those virus strains after continuous passage for several times? For control and prevention of reemerging of SARS, a safe and efficacious vaccine against SARS is urgently needed. Neutralization of virus infectivity by antibodies is an important component of immunity to several virus infections (Klasse and Sattentau, 2002). Production of neutralizing antibodies is crucial for evaluation of the effectiveness of a candidate vaccine. Thus detection of neutralizing antibodies is an essential part of SARS vaccine development .In order to answer these questions and to provide neutralization assay, we proposed this research proposal. The project consists three parts.
(1) In order to understand the biological properties of SARS-CoV isolated in Taiwan. We use the first SARS-CoV strain isolated in our laboratory as reference. Some clinical isolates of SARS-CoV in the later time of outbreaks were chosen for study. In addition to the low passage number of virus isolates, the higher passage numbers of some strains were also used for comparing. By using the established neutralization assay, we will compare the neutralization activity of different clinical isolates against various SARS patient sera.
(2) We also intend to continue set up a reference laboratory for screening of the neutralizing activities against SARS coronavirus present in the serum of immunized animals and in
monoclonal antibodies. SARS coronavirus. TW-1 strain and Vero E6 cell line will be used throughout the experiments. Micro-neutralization test for SARS-CoV will be used in general. The inhibition of 100 TCID50 was used as the criteria for neutralization assay. The conditions for performance of micro-neutralization assays (TCID50) will be optimized and standardized further. In addition to conventional neutralization assay, rapid virus neutralization test is performed by quantitatively detection of viral RNA using reverse transcription and real-time PCR. Real-time PCR has been extremely useful for studying viral agents of infectious disease and helping to clarify disputed infectious disease processes. Real-time PCR is very sensitive and rapid. It has been used to detect viral transcripts with high sensitivity (Wang, et al. 2000, 2002).
Using reverse transcription (RT) and real-time PCR, it is possible to detect changes of the quantities of viral RNA. Our preliminary data showed that viral RNA could be detected early after infection. Using RT real-time PCR, neutralization of SARS CoV could be determined as early as eight hours after inoculation of cells with the virus-serum mixture. This rapid method will be optimized, standardized and compared with traditional methods. In order to find the appropriate time to detect the RNA species, the kinetics for RNA transcription were quantitatively analyzed. This method could reduce the waiting time at least 48 hours as compared to the traditional TCID50 method. In addition to the advantage of rapidity, this method could reduce the observation time under the microscope and the working time in the P3 laboratory.
(3) Since the virological work of the SARS-CoV needs to be carried out in the P3 facility, maintenance of a standard P3 core facility is very important for the SARS research. In last year’s project, the P3 facility at the College of Medicine, National Taiwan University has gone through a complete check up and maintenance as well as renewal of several equipments. In this year, the P3 facility requests expenses for the basic maintenance, consumables and the laboratory manager.
With the global efforts to combat this highly contagious and potentially life-threatening infectious disease, it is essential to maintain a standard P3 core facility for many studies involving the SARS-CoV. A good P3 facility at the College of Medicine, National Taiwan University will contribute greatly to our fights against this important infectious disease.
The results of this study can provide virus strains for molecular epidemiological study of SARS- CoV in Taiwan. It will provide important information for the vaccine development.
Materials and Methods
Continuous passage of SARS-CoV isolates. 3 to 5 strains of SARS-CoV isolated in NTUH
will be selected for continuous passage. Vero E6 will be used as the host cells. After confluent growth of cells in 25 cm2 tissue culture flask, virus will be added into the flask and incubate at 37 ℃ 5% CO2 incubator for absorption.1 hour. Eagle’s MEM supplemented with 2 % fetal calf serum will be added as maintenance media. After 3-5 days, when the viral cytopathic effect reaching above 75%, the culture supernatants are collected and centrifuged at 1800 rpm for 10 min. The clarified supernatants are collected and aliquoted into small quantities and stored at temperatures below –75℃. The passages were all performed in P3 laboratory located in College of Medicine, National Taiwan University (MC, NTU).
Titration of SARS virus infectivity. The virus stock will be titrated using standard TCID50 method and plaque assay. Virus suspension will be serially 10-fold diluted. For TCID50 method, the diluted virus will be seeded onto 96-well plate, then Vero E6 cells will be added. In order to get the best results, the length of incubation time, from 2 to 5 days, will be determined first. TCID50, 50% tissue culture infective dose, will be calculated according to the Reed-Muench method. For plaque assay, the diluted virus will be inoculated onto monolayer cultures of Vero E6 in 6-well or 24-well plate. After adsorption at 37℃ for 1h, the cells will be overlaid with EMEM containing 2% FCS and 0.3-0.4% agarose. The plaques will be visualized by staining with crystal violet. Before counting the plaques, the virus and cells in the plates will be treated with 10% formalin for at least 1 h. Plaque forming unit per ml (PFU/ml) will be calculated. .
Virus neutralization test. Neutralization test utilizing TCID50 method will be performed in 96-well plate. Virus suspension diluted to contain 100 TCID50 (in the volume loaded into each well) will be mixed with equal volume of diluted serum or monoclonal antibodies. When screening of large numbers of immunized animal serum, the serum will be diluted at 1:10 and tested by using TCID50 method. After the virus-serum mixture is incubated at 37℃ for one hour, 100μl of mixture was added to each micro-well with confluent growth of Vero E6 cells. T Two to 5 days later, the viral CPE will be examined under the inverted microscope. The serum with neutralizing activity will be further diluted, and the endpoint of the neutralizing activity will be determined.
Extraction of viral RNA. After Vero E6 cell suspension has been added onto the virus-serum
mixture in 96-well plate, total RNA will be extracted from the virus-infected cells at certain time point. A semi-automatic RNA extractor (ABI 6100) employed the RNA isolation kit (ABI) will be used.
Primer and probe design. Appropriate primer and probe sequences for real-time PCR are
selected through screening by the software PrimerExpress (ABI). The sequences in ORF1a and N gene of the genome of TW-1 strain (AY291451) have been screened. Fragments of less than 100 bp in length will be generated when subjected into the RT-real-time PCR. Primers for amplification of larger fragments of 200-300 bp in length containing the above sequences will also be designed using software OLIGO 6. These larger fragments are cloned and used as the constructs for production of RNA.
Generation of constructs and RNA. The 200-300 bp fragments generated by PCR, using the
plasmid containing TW-1 fragment as the template, are cloned into the vector pCRII-TOPO (Invitrogen). The clones are screened by restriction enzyme digestion and nucleotide sequence analysis. The in vitro-transcribed positive-sense RNA generated by T7 transcription (Promega) of the HindIII-linearized plasmid are purified by phenol-chloroform extraction and quantified by spectrophometry (Wang et al. 2000, Wang et al. 2002). The copy number of the RNA is calculated based on the concentration measured and its molecular weight.
RT-real-time PCR. RT-real-time PCR will be performed in a separate room from that used
for RNA isolation, and precautions for PCR will be followed to avoid contamination (Kwok and Higuchi 1989). An RT-real-time PCR will be used to quantify intracellular SARS virus RNA by
using different primers in the RT step. For quantification of positive-sense SARS virus RNA, an aliquot of total RNA isolated from SARS virus-infected or uninfected cells and known amounts of positive-sense RNA (ex. 1, 10, 100, 1,000, 10,000, 100,000, and 1,000,000 copies) are subjected to RT using an antisense primer, and a cDNA synthesis kit (Life Technologies). An aliquot of the cDNA is then subjected to real-time PCR using the designed primers and Probe, the TaqMan universal PCR master mix reagent kit (ABI). The amplification conditions will follow that recommended by the manufacturer. The ABI Prism 7000 sequence detector will be used to analyze the emitted fluorescence during amplification. A positive result is defined by the cycle number (CT value) required to reach the threshold, which is 10 times the standard deviation of the mean baseline emission calculated for PCR cycles 3 to 15. The amounts of intracellular SARS virus RNA will be determined.
Neutralization Test for Real-Time RT-PCR
Vero E6 cells were seeded in a 96-well plate in MEM plus 10% FBS at 37°C overnight until confluence. 100 TCID50SARS-CoV was incubated with the sera from patients in the convalescent phase or from mice immunized with synthetic peptides of SARS-CoV at 37°C for 1 h. Subsequently the cells were inoculated with the mixture of virus and sera at 37°C for 1 h. After the wells were washed with PBS three times, and then MEM plus 2% FBS was added to each well in a total volume of 100 μL. The contents of wells were collected on 8hr post-infection and titrated with real-time RT-PCR described as above.
Optimization and standardization of neutralization assay for SARS-CoV. We intend to
continue the task to establish a reference laboratory for screening of SARS neutralizing antibodies, not only by the traditional methods but also by a rapid method. In addition conventional neutralization assay, rapid virus neutralization test is performed by quantitatively detection of viral RNA using reverse transcription and real-time PCR. This method will be standardized and compared with traditional methods. The efficiency of real-time PCR using different sequences of SARS CoV genome as the target will be evaluated. It will be useful for improving the rapid assay. The neutralization will be collaborated with other principal investigators for antiviral activity assay and vaccine effectiveness evaluation.
Maintenace of P3 facility. In last year’s project, our P3 facility has gone through a complete
check up and maintenance as well as renewal of several equipments. In this year, the P3 facility requests expenses for the basic maintenance, consumables and the laboratory manager for support all the SARS study..
Results
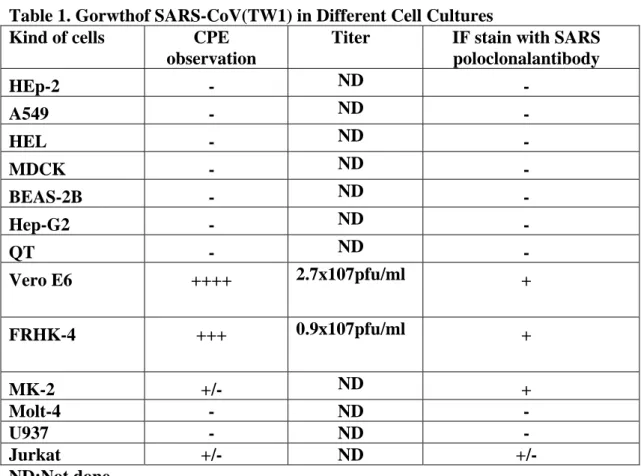
Susceptibility of cell lines for SARS-CoV growth
The cellular tropism of SARS-CoV was analyzed by passage TW1 strain in the different cell lines. The results indicated that cell lines (Vero E6 and FRHK-4) established from monkey were susceptible for SARS-CoV (Table 1). Among them, Vero E6 was more sensitive than FRHK-4 for virus growing. All cell lines from human origin were not support for SARS-CoV growth.
Optimization and standardization of neutralization assay for SARS-CoV
At original developed neutralization test, the cell suspension was added after the end of incubation of serum-virus interaction, the pre-cultured Vero E6 cells on 96 well plates were used in the latter stage due to the consistence of reading of the final results of neutralization. The titrations of SARS-CoV on the Vero E6 cells were also compared between culture medium with 2% fetal calf serum and serum free culture medium. The results indicated that the higher titer of SARS-CoV was found in serum medium than it did in the serum free medium (1 log10 ). So the monolayer and the culture medium with 2% fetal calf serum was used for neutralization test in the whole study.
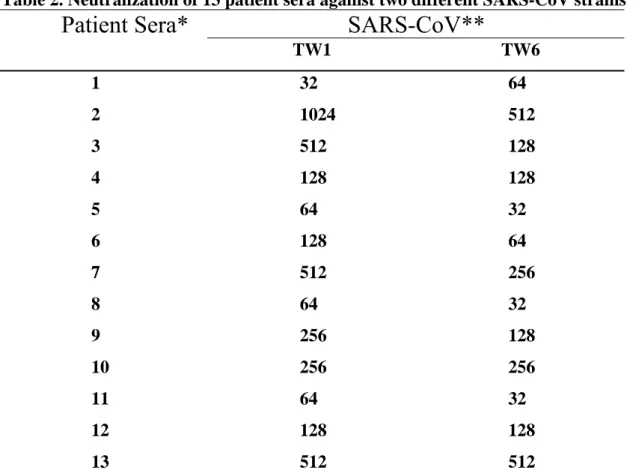
Neutralization activity of SARS-patients sera against SARS-CoV
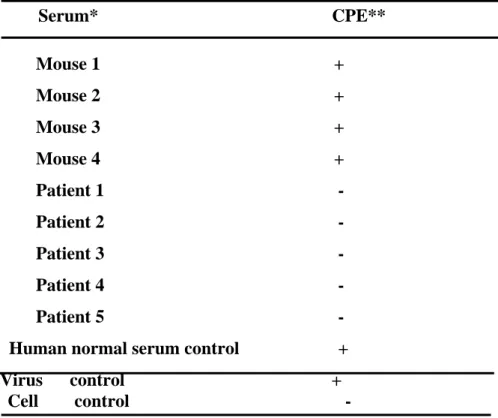
Convalescent human sera were collected from 13 patients with probable SARS and examined for their neutralization activities. Among the 13 specimen, three (sera 1-3) were collected from patients infected at the first stage of the SARS outbreak in Taiwan, ten (sera 4-13) were collected from patients infected at the second stage of the outbreak. The TW1 and TW6 strains were originally isolated from the patients 1 and 4, respectively. Results in Table 2 indicated that all of the patient sera possess neutralizing activity against both homologous and heterologous strains of SARS-CoV. The cross neutralization activities of the patient sera to the heterologous SARS viruses were mostly equal or only at a 2-fold difference to the homologous strain. The established neutralization test can also applied for the detection of mouse sera against synthetic peptides of SARS-CoV(Table 3).
Continue passage of SARS-CoV
SARS CoV can grow very after the continuous passage of TW1 in VeroE6 ten times. The virus yield sill keeps in the high titer even at the end of ten passagers(Table 4).
Establishment of a real-time PCR assay
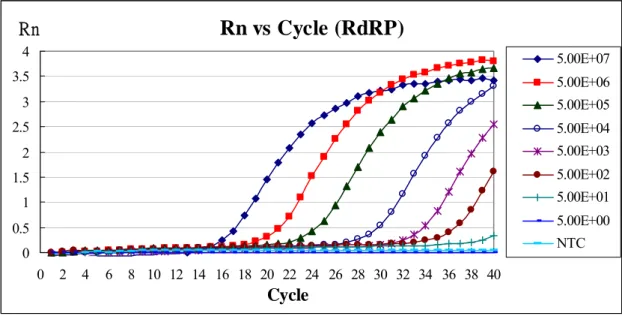
To establish a real-time RT-PCR assay for SARS-CoV, a primer pair, real-pol-F and real-pol-R, and a fluorescent probe (FAM), probe-pol, in SARS-CoV RdRP region were designed. In addition, a primer pair, real-3’-F and real-3’-R, and a fluorescent probe (VIC), probe-3’, in SARS-CoV NC region were designed. The feasibility of the real-time PCR assay was examined first. A standard curve was generated from serial 10-fold dilutions of known amounts of the SARS-CoV RdRP RNA transcript or NC RNA transcript (Fig.1,2)). The assay
had a linear response over at least a 7-log10 concentration range, from 50 to 50,000,000 copies/reaction (R2 = 0.997 or 0.999).
Time course of SARS-CoV RNA in virus-infected Vero E6 cell
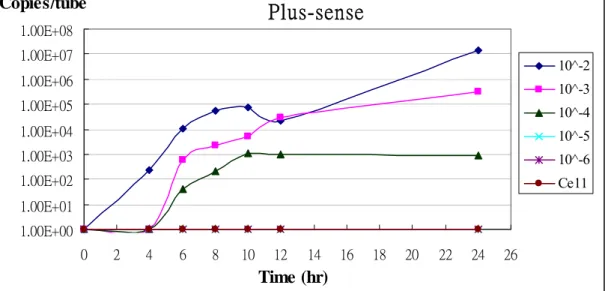
The SARS-CoV stock titer was determined by observation of cytopathic effect, which was 105.5 TCID50/25 μL (1.64 × 107 TCID50/ mL). Vero E6 cells were inoculated with serial 10-fold dilutions of SARS-CoV stock and subsequently the amounts of SARS-CoV RNA were detected at different time points, 0, 4, 6, 8, 10, 12, and 24 hours. Quantification of SARS-CoV by the real-time PCR assay during the course of infection is shown. The amounts of viral RNA increased gradually with time and could be detected at significant level after 8 hours. Even viral RNA could be detected at 4 hours after cells were infected with 10-2 virus stock (Fig. 3). These results indicated the applicability of this assay in monitoring viral replication during the course of infection.
Neutralization test by real-time RT-PCR
To determine whether the antibodies in the sera had neutralizing activity against SARS-CoV, the neutralization tests were necessary. The sera from patients in the convalescent phase or from mice immunized with synthetic peptides of SARS-CoV were examined by the neutralization tests. Thereafter the cytopathic effects (CPE) were observed by microscopy; the viral RNA, meanwhile, was measured by real-time PCR(Fig 4,5). The results obtained by traditional method demonstrated that the sera from healthy individuals had no neutralizing activities against SARS-CoV as CPE could be observed. However, the sera from patients in the convalescent phase could neutralize SARS-CoV because of no CPE. Unfortunately, the sera from mice immunized with synthetic peptides of SARS-CoV were unable to neutralize SARS-CoV.
The results of real-time PCR were consistent with those of traditional methods as above. The SARS-CoV RNA could be detected after the SARS-CoV was pretreated with sera from normal individuals and from mice. Nevertheless it could not be detected after virus was pretreated with sera from patients in the convalescent phase.
Neutralization activity assay
The P3 laboratory facility was kept in whole function. It provide all SARS related study in National Taiwan University. This project also help to do neutralization assay of mouse hyper-imimune with Co60 inactivated SARS CoV or SARS VLP and some chemical agents. The samples asked for neutralization assays were from several PIs
from NHRI, NTU other research institutes.
Discussion
The different susceptibility of various cell lines suggested that the expression of receptors for SARS CoV on the cells are different. For clinical diagnosis use the VeroE6 is still the best chose for culture of SARS CoV.
Various phyologenetic clusters of SARS-CoV have been reported during different outbreaks occurred in Taiwan. However the hyper-immune mouse of one cluster of SARS-CoV isolated from the first stage of outbreak could also effectively neutralize another cluster of SARS-CoV isolated from the second stage of outbreak. And the convalescent sera from SARS patients in two outbreaks also could cross neutralize both kinds of clusters of SARS-CoV efficiently. The results imply that choosing of SARS-CoV virus strain from first stage of outbreak in Taiwan as a vaccine candidate may provide the protection of different clusters virus infection and vice versa. It is also suggested that to get re-infection in human population recovered from the first stage outbreak of SARS with different clusters of SARS-CoV occurred in the late stage of outbreak in Taiwan is not easily anticipated. The biological properties of ten SARS-CoV isolates will be characterized in the future.
This study revealed that utilizing RT real-time PCR could indeed detect the neutralizing activity against SARS-CoV more rapidly. The parameters of this new method need be optimized further and more serum samples are required for further evaluation.
Reference
1. Berger, A., C. H. Drosten, H. W. Doerr, D. M. Stirmer, and W. Preiser. 2004.Severe acute respiratory syndrome (SARS)-paradigm of an emerging viralinfection. J. Clin. Virol. 29:13-22.
2. CDC. 2003. Update: severe acute respiratory syndrome-worldwide and United States, MMWR Mor. Mortal. Wkly. Rep. 52:664-665.
3. Poutanen, S. M., D. E. Low, B. Henry, S. Finkelstein, D. Rose, K. Green, R.Tellier, R. Draker, D. Adachi, M. Ayers, A. K. Chan, D. M. Skowronski, I.Salit, A. E. Simor, A. S. Slutsky, P. W. Doyle, M. Krajden, M. Petric, R. C.Brunham, A. J. McGeer, National Microbiology Laboratory, Canada, and Canadian SARS Study Team. 2003. Identification of severe acute respiratory syndrome in Canada. N. Engl. J. Med. 348:1995-2005.
4. WHO. www.who.int/csr/sars/country/table2003_09-23/en (Accessed Jan 26, 2004).
5. Ksiazek TG, Erdman D et al. 2003. A novel coronavirus associated with acute respiratory syndrome. N Engl J Med 348: 1948-51.
6. Drosten C et al. 2003. Identification of a novel in patients with severe acute respiratory syndrome. N Engl J Med 348: 1967-76.
7. Poutanen SM et al. 2003.Identification of severe acute respiratory syndrome in Canada. The New Engl J Med Apr. 10.
8. Tsang KW et al. 2003. A cluster of cases of severe acute respiratory syndrome in Hong Kong. The New Engl J Med. March 31
9. Enserink M and Vogel G. 2003. Deferring competition, global net closes in on SARA. Science 300: 224-225.
10. McIntosh, K. (2001). Fields Virology. New York, Lippincott Williams and Wilkins.
11. Leparc-Goffart, I., S. T. Hingley, et al. (1998). "Targeted recombination within the spike gene of murine coronavirus mouse hepatitis virus-A59: Q159 is a determinant of hepatotropism." J Virol 72(12): 9628-36.
12. Sanchez, C. M., A. Izeta, et al. (1999). "Targeted recombination demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a determinant of its enteric tropism and virulence." J Virol 73(9): 7607-1.
13. Escors, D., J. Ortego, et al. (2001). "The membrane M protein of the transmissible gastroenteritis coronavirus binds to the internal core through the carboxy-terminus." Adv Exp Med Biol 494: 589-93.
14. Lai, M. M. C. and K. V. Holmes (2001). Fields Virology. New York, Lippincott Williams and Wilkins.
15. Marra MA et al. 2003. The genome sequence of the SARS-associated coronavirus. Science 300:1399-04.
16. Rota, P. A., M. S. Oberste, et al. (2003). "Characterization of a novel coronavirus associated with severe acute respiratory syndrome." Science 300(5624): 1394-9.
17. Ruan Y et al. 2003. Comparative full-length genome sequence analysis of 14 SARS coronavirus isolates and common mutations associated with putative origins of infection. Lancet Published on line May 9, 2003.
18. Hsueh P. R., C. H. Hsiao, S. H. Yeh, W. K. Wang , P. J. Chen, J. T. Wang, S. C.Chang, C. L. Kao, P. C. Yang, SARS Research Group of NTU and NTUH.2003. Microbiologic characteristics, serologic responses, and clinical manifestations in acute respiratory syndrome, Taiwan. Emerg. Infect. Dis.9:1163-1167.
19. Klasse PJ, Sattentau. 2002. Occupancy and mechanism in antibody-mediated neutralization of animal viruses. J Gen Virol 83:2091-2108.
20. Mackay IM, Arden KE, Nitsche A. 2002. Survey and summary – Real-time PCR in virology. Nucleic Acids Research 30:1292-1305.
21. Wang WK., Lee CN., Kao CL., Lin YL. and King CC. 2000. Quantitative competitive reverse transcription-PCR for quantification of dengue virus RNA. J. Clin. Microbiol. 38:3306-3310.
22. Wang WK., Lee CN., Kao CL., Lin YL. and King CC. 2000. Quantitative competitive reverse transcription-PCR for quantification of dengue virus RNA. J. Clin. Microbiol. 38:3306-3310.
Table 1. Gorwthof SARS-CoV(TW1) in Different Cell Cultures
Kind of cells CPE
observation
Titer IF stain with SARS
poloclonalantibody HEp-2 - ND - A549 - ND - HEL - ND - MDCK - ND - BEAS-2B - ND - Hep-G2 - ND - QT - ND - Vero E6 ++++ 2.7x107pfu/ml + FRHK-4 +++ 0.9x107pfu/ml + MK-2 +/- ND + Molt-4 - ND - U937 - ND - Jurkat +/- ND +/- ND:Not done 10
Table 2. Neutralization of 13 patient sera against two different SARS-CoV strains
SARS-CoV**
Patient Sera*
TW1 TW6 1 32 64 2 3 4 1024 512 128 512 128 128 5 64 32 6 128 64 7 512 256 8 64 32 9 256 128 10 256 256 11 64 32 12 128 128 13 512 512* Sera 1-3 were collected from patients infected at stage 1 of SARS outbreak in Taiwan and sera 4-13 were collected from patients infected at stage 2 of SARS outbreak.
** TW1 and TW6 were isolated from patients 1 and 4, respectively.
Table 3. Detection of neutralizing antibodies against SARS-CoV
Serum* CPE** Mouse 1 + Mouse 2 + Mouse 3 + Mouse 4 + Patient 1 - Patient 2 - Patient 3 - Patient 4 - Patient 5 -
Human normal serum control + Virus control +
Cell control -
* Mouse 1~4: sera from mice immunized with synthetic peptides of SARS-CoV Patient 1~5: sera from patients in the convalescent phase
Human control: sera from normal individuals
** CPE observation at 72 hours after SARS-CoV infection + : CPE > 50% - : No CPE
Table 4. Virus titers in different passages of TW 1
Number of Passage Copies
5 1.133X109 6 5.469X109 7 4.504X109 8 1.687X109 9 1.931X109 10 1.288X109
12
Rn vs Cycle (RdRP)
0 0.5 1 1.5 2 2.5 3 3.5 4 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 Cycle Rn 5.00E+07 5.00E+06 5.00E+05 5.00E+04 5.00E+03 5.00E+02 5.00E+01 5.00E+00 NTCStandard Curve (RdRP)
R
2= 0.9966
0 5 10 15 20 25 30 35 401.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 1.00E+07 1.00E+08
Copy No. CT
Fig. 1 Standard curve for real-time PCR with the RdRP RNA
transcripts
The total volume of reactants is 25 μL, including 1× TaqMan Universal PCR Master Mix, 8 pmol primers(Real-pol-F and Real-pol-R), 8 pmole Taqman-MGB probe (Probe pol) and 5 μL RdRP positive-sense RNA standards (5 ~ 5 × 107 molecules). Real-time PCR was used to quantify the amounts of RNA and standard curve was plotted. R2 = 0.9966。NTC: no template control。
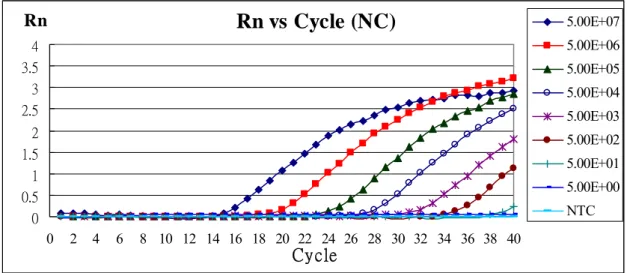
Rn vs Cycle (NC)
0 0.5 1 1.5 2 2.5 3 3.5 4 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 Cycle Rn 5.00E+07 5.00E+06 5.00E+05 5.00E+04 5.00E+03 5.00E+02 5.00E+01 5.00E+00 NTCStandard Curve (NC)
R
2= 0.9987
0 5 10 15 20 25 30 35 401.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 1.00E+07 1.00E+08
Copy No. CT
Fig. 2 Standard curve for real-time PCR with the NC RNA transcripts
The total volume of reactants is 25 μL, including 1× TaqMan Universal PCR Master Mix, 8 pmol primers(Real-3’-F and Real-3’-R), 8 pmole Taqman-MGB probe (Probe 3’) and 5 μL NC positive-sense RNA standards (5 ~ 5 × 107 molecules). Real-time PCR was used to quantify the amounts of RNA and standard curve was plotted. R2 = 0.9987。NTC: no template control。
Plus-sense
1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 1.00E+07 1.00E+08 0 2 4 6 8 10 12 14 16 18 20 22 24 26 Time (hr) Copies/tube 10^-2 10^-3 10^-4 10^-5 10^-6 Ce11Minus-sense
1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 1.00E+07 1.00E+08 0 2 4 6 8 10 12 14 16 18 20 22 24 26 Time (hr) Copies/tube 10^-2 10^-3 10^-4 10^-5 10^-6 Ce11Fig. 3 Time course of SARS-CoV RNA in virus-infected Vero E6 cells suspension was
10-fold serially diluted and then inoculated Vero E6 cells. The contents of wells were collected on 0, 4, 6, 8, 10, 12, and 24 hr postinfection. The templates, positive-sense NC gene RNA were reverse transcribed with clone-3’-R primer. The negative-sense NC gene RNA were reverse transcribed with clone-3’-F primer. Real-time PCR was then used to quantify the amounts of cDNA. Cell: uninfected Vero E6 cells。
Neutralization Test (RdRP)
1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 Mou se 1 Seru m Mouse 2 Se rum Mouse 3 Seru m Mouse 4 Serum Patien t1 Seru m Patien t2 Seru m Patie nt3 Ser um Patie nt4 Ser um Patient5 Serum Huma n con trol Seru m Virus contr ol Cell contr ol Copies/tube 0 hr 8 hrFig 4. Detection of neutralizing antibodies in sera by real-time PCR (RdRP
gene)
100 TCID50 SARS-CoV was incubated with the 10-fold diluted sera from patients in the convalescent phase or from mice immunized with synthetic peptides of SARS-CoV at 37°C for 1 h. Subsequently the cells were inoculated with the mixture of virus and sera. Then the wells were washed with PBS three times. The contents of wells were collected on 0 and 8hr postinfection. The templates, positive-sense RdRP gene RNA were reverse transcribed with clone-pol-R primer. Real-time PCR was then used to quantify the amounts of cDNA.
Neurtalization Test (NC)
1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 Mouse 1 Se rum Mouse 2 Se rum Mouse 3 Se rum Mouse 4 Se rum Patien t1 Seru m Patien t2 Seru m Patien t3 Seru m Patien t4 Seru m Patien t5 Seru m Huma n con trol Se rum Viru s contr ol Cell contr ol Copies/tube 0 hr 8 hrFig. 5 Detection of neutralizing antibodies in sera by real-time PCR (NC
gene)
100 TCID50 SARS-CoV was incubated with the 10-fold diluted sera from patients in the convalescent phase or from mice immunized with synthetic peptides of SARS-CoV at 37°C for 1 h. Subsequently the cells were inoculated with the mixture of virus and sera. Then the wells were washed with PBS three times. The contents of wells were collected on 0 and 8hr postinfection. The templates, positive-sense NC gene RNA were reverse transcribed with clone-NC-R primer. Real-time PCR was then used to quantify the amounts of cDNA.