JSPP © 1991
Senescence of Rice Leaves XXX. Levels of Endogenous Poly amines
and Dark-Induced Senescence of Rice Leaves
Chien Teh Chen and Ching Huei Kao
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China The role of endogenous polyamines in the control of dark-induced senescence of detached rice leaves was investigated by quantitating levels of various polyamines by HPLC. Putrescine, spermidine and spermine were all present throughout senescence. Neither cadaverine nor 1,3-diaminopropane was detected. During dark-induced senescence, there was a marked decrease in levels of putrescine and an increase in those of spermidine and spermine. The rate of produc-tion of ethylene increased markedly upon excision of leaves. a-Difluoromethylarginine (DFMA) and a-difluoromethylornithine (DFMO) caused a reduction in levels of putrescine, yet had no effect on levels of spermidine and spermine. Neither DFMA nor DFMO had any effect on senes-cence or on the production of ethylene. Treatment with dicyclohexylamine (DCH) and methylglyoxal Ws-(guanylhydrazone) (MGBG) reduced levels of spermine and increased those of putrescine in detached leaves. After treatment with DCH or MGBG, both senescence and the production of ethylene were significantly promoted. The current results suggest that endogenous polyamines may not play a significant role in the control of dark-induced senescence of rice leaves. This conclusion is supported by the further observations that (a) benzyladenine, which is known to retard senescence, decreased levels of putrescine but had no effect on those of sper-midine and spermine; and (b) ABA, which promoted senescence, increased levels of putrescine and had no effect on those of spermidine and spermine.
Key words: S-adenosylmethionine decarboxylase — Arginine decarboxylase — Ethylene — Or-nithine decarboxylase — Polyamines — Rice leaf senescence.
In recent years, increasing attention has been paid to the question of whether or not polyamines have a func-the possible role of polyamines in growth and differentia- tional role in func-the control of senescence involves func-the use of tion (Evans and Malmberg 1989). Polyamines are effective inhibitors of polyamine synthesis. Using this approach, in retarding senescence of excised leaves and protoplasts Birecka et al. (1990) showed that dark-induced leaf senes-(Altman 1982, Cheng and Kao 1983, Galston et al. 1978, cence of barley and oats was not correlated with the levels Kaur-Sawhney and Galston 1979). Since decreases in of endogenous polyamines. The present investigation was levels of polyamines have been observed in aging and conducted to determine the role of endogenous polyamines senescing leaves, it is reasonable to suppose that they play a in the control of senescence of detached rice leaves. The role in the regulation of leaf senescence (Kaur-Sawhney et mechanism by which exogenous polyamines retard leaf se-al. 1982, Srivastava 1981). However, a decrease in levels nescence may be related to the possibility that they inhibit of polyamines is not always correlated with the onset of the biosynthesis of ethylene (Evans and Malmberg 1989). leaf senescence (Birecka etal. 1984). One way to approach Both ethylene and polyamines share SAM as a common precursor (Miyazaki and Yang 1987). Thus, changes in Abbreviations: ADC, arginine decarboxylase; BA, benzyl- rates of production of ethylene during dark-induced senes-adenine; Cad, cadaverine; Dap, 1,3-diaminopropane; DCH, cence of detached rice were also investigated,
dicyclohexylamine; DFMA, difluoromethylarginine; DFMO,
a-difluoromethylornithine; MGBG, methylglyoxal bis-(guanyl- Materials and Methods hydrazone); ODC, oraithine decarboxylase; Put, putrescine;
SAM, S-adenosylmethionine; SAMDC, S-adenosylmethionine Plant material—Rice (Oryza sativa L. cv. Taichung decarboxylase; Spd, spermidine; Spm, spermine. Native 1) seedlings were grown as previously described
(Chen et al. 1990). The apical 3 cm of the third leaves of 10-day-old seedlings were used for experiments. Ten seg-ments of rice leaves were floated on 10 ml of distilled water or of a test solution in a Petri dish. All samples were in-cubated at 21°C in the dark.
Chi determination—Chi was determined by the
method of Wintermans and De Mots (1965) after extrac-tion in 96% ethanol. Chi content is expressed as mg per g fresh weight.
Protein determination—Leaf segments were
homoge-nized in 25 mM sodium phosphate buffer (pH 7.5). The ex-tracts were centrifuged at 17,000 xg or 20min, and the supernatants were used for determination of protein by the method of Lowry et al. (1951).
Polyamine determinations—Leaf segments weighing
about 125 mg were homogenized in 5 ml of cold 5% perchloric acid, and the homogenates were kept at 4°C for 24 h. Each homogenate was centrifuged for 20min at 17,000 x g and the clear supernatant was used for benzoyla-tion according to Flores and Galston (1982). In brief, 1 ml of NaOH (2 N) was mixed with the 1 ml of the supernat-ant. After addition of 15 fA of benzoyl chloride, vortexing for 10 s, and incubation for 20 min at room temperature (about 25°C), 2.5 ml of saturated NaCl were added. Ben-zoyl-polyamines were extracted in 2.5 ml of diethylether. After centrifugation at 3,000 xg for 15 min, 1.5 ml of the ether phase were collected, evaporated to dryness in a vacuum concentrator and redissolved in 100 fil of metha-nol. Polyamine standards were benzoylated in a similar way. Ten fA aliquots of each redissolved sample were in-jected into a Waters M-6 UK Universal Liquid Chromato-graph. They were eluted through a 4 x 250 mm, 5 fim parti-cle size C18 reverse-phase column at a flow rate of 1 ml min"1 using a water (solvent A)/methanol (solvent B) step-ped gradient program (50 to 65% B in 7 min/65 to 80% B in 10.5 min/80 to 100% B in 2 min) followed by a column cleaning/regeneration cycle. Detection was accomplished by a Waters M481 absorbance detector at 254 nm. Results were recorded on a Waters M740 data module and plotted as nmol per g fresh weight.
Ethylene determination—Leaf segments were placed
vertically in test tubes which were closed with rubber stop-pers. After 1 h of incubation at 27°C in the dark, a 1-ml sample of gas was withdrawn from the headspace of each test tube. Ethylene was assayed using a gas chromato-graph equipped with an alumina column and a flame ioniza-tion detector.
Results
The senescence of detached rice leaves was followed by measuring the decrease in levels of Chi and the protein. Figure 1 shows that a decrease in levels of Chi and protein was evident at 2 and 3 days, respectively, after leaf
detach-% vf/30 ui
5
K a i o 0 10 UJ m z I -IU O PROTEIN • CHLOROPHYLL AC2H4 4.5 3.0vE-I
1.5 g3
OFig. 1 Changes in levels of chlorophyll and protein and in rates of production of ethylene in detached rice leaves incubated in darkness. Means ± SE, 4 replicates.
ment. This indicates that proteolysis precedes Chi degrada-tion during senescence of detached rice leaves in the dark, in good agreement with the results of Shibaoka and Thimann (1970) and our earlier work (Kao 1978).
In freshly excised rice leaf segments, ethylene produc-tion was low (Fig. 1). After incubaproduc-tion, the rate of ethylene production increased, reaching a maximum in 2 days, and subsequently declined (Fig. 1). The rise of ethylene production occurred prior to senescence.
A typical chromatogram of benzoyl derivatives obtaed from freshly excisobtaed leaves, with authentic standards in-cluded for comparison is shown in Figure 2. The major peaks corresponded to Put, Spd and Spm. Peaks in leaf ex-tracts corresponding to authentic Cad and Dap were not ob-served. The level of Put in detached rice leaves decreased progressively with increasing duration of incubation in darkness (Fig. 3). However, the level of Spd increased im-mediately after leaf detachment, reached a maximum level at days 1 and 2, and then decreased to the basal level (Fig. 3). The level of Spm also increased immediately after excision of leaves, reached maximum level at day 2, and remained unchanged thereafter (Fig. 3).
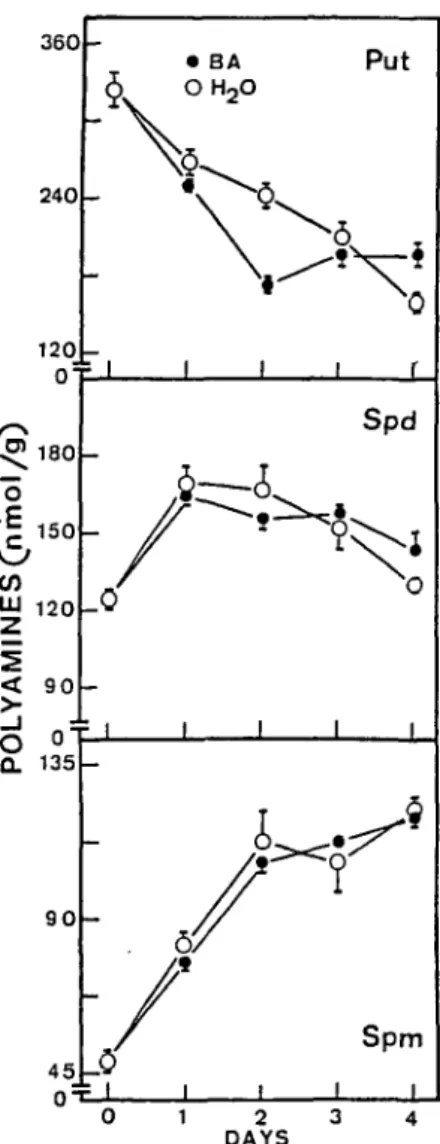
The time courses of changes in levels of Chi and pro-tein in detached leaves floating on water and on a solution of BA, a synthetic cytokinin, in darkness, are shown in Figure 4. It is clear that BA retards senescence of detached
100- 75- 50-X
A"
A ^ / V-i Pu t B _ J | / h^ c a a c_ \ L _ A J
r
3 JLJU-Sp m1
uU
r-Fig. 2 Separation of benzoylated diamines and polyamines by HPLC. A, Extract of leaves of 10-day-old seedlings; B, leaf ex-tract with authentic Put; C, leaf exex-tract with authentic Cad and Dap; D, leaf extract with authentic Spd and Spm. All authentic standards were present at 1.5 nmol. Numbers below trace A are retention times.
leaves. However, BA had no effect on levels of Spd and Spm, but it did decrease the level of Put over the first two days (Fig. 5). Figure 6 shows the effects of ABA on the se-nescence of detached leaves. ABA significantly promoted senescence. The effect of ABA on levels of polyamines is shown in Figure 7. ABA significantly increased the level of Put. However, no significant difference in levels of Spm in detached leaves was observed between those treated with ABA and controls in water. The decrease in level of Spd only occurred four days after the start of treatment with ABA. The observed effects of BA and ABA on levels of Put in detached rice leaves are in sharp contrast to those reported by other investigators (Mukhopadhyay et al. 1983, Suresh and Adiga 1978, Suresh et al. 1978), who reported that cytokinin and ABA increased and decreased levels of Put, respectively.
To characterize further the role of endogenous poly-amines in dark-induced senescence, inhibitors of the biosyn-thesis of polyamines, such as DFMO, DFMA, MGBG and
3 6 0 240 120
-
o-/^
5. 180 o E & 150 <n Hi | 120>-o
0. 90r-0 135 90 -4 5 -/ ^ : I 1 1 • /-0
r 1 1 1 Put 1 1 Spd i i S p m i i 2 3 DAYSFig. 3 Changes in levels of polyamines in detached rice leaves in-cubated in darkness. Mean ± SE, 3 replicates.
DCH, were used to change level of intracellular polyamines in detached rice leaves. Both DFMO, an irreversible inhibi-tor of ODC, and DFMA, an irreversible inhibiinhibi-tor of ADC, significantly reduced levels of Put, yet neither had any effect on levels of Spd and Spm (Table 1). The biosynthesis of Put in the aleurone layers of barley seeds was not inhibited by DFMO or DFMA (Lin 1984). However, our results are consistent with those reported by Roberts et al. (1984). DFMA was more effective in reducing the level of Put than was DFMO, indicating that the biosynthesis of Put in rice leaves occurs mainly via the reaction catalyzed by ADC. However, neither DFMA nor DFMO had any effect on the levels of Chi and protein or on the rate of production of ethylene (Tables 1 and 2). DCH and MGBG reduced levels of Spm in detached rice leaves (Table 1). There was also an increase in levels of Put in the DCH- and MGBG-treat-ed detachMGBG-treat-ed rice leaves. Neither DCH nor MGBG affectMGBG-treat-ed
4.5 3.0 5>40 3 0 2 0 10 -CHLOROPHYLL I I I I
Fig. 4 Effects of BA on levels of Chi and protein in detached
rice leaves incubated in darkness. Means ± SE, 4 replicates.
levels of Spd (Table 1) but they both caused significant decreases in levels of Chi and protein and increased rates of production of ethylene (Tables 1 and 2).
Discussion
The present investigation showed that Put, Spd and Spm were present in detached rice leaves throughout senes-cence. However, neither Cad nor Dap was detected in detached rice leaves during dark-induced senescence, in-dicating that Cad and Dap may not be involved in the con-trol of dark-induced senescence of detached rice leaves.
360 240 120 0=r I 180 150 en .. UJ 1 2 OL 0 • BA Put OH2O
-5
j r_ SpdFig. 5 Effects of BA on levels of polyamine in detached rice
leaves incubated in darkness. Means ± SE, 3 replicates.
The absence of Cad and Dap has also been reported in pea ovaries during senescence (Carbonell and Navarro 1989).
Table 1 Effects of DFMO, DFMA, DCH and MGBG on levels of polyamines and the rate of production of ethylene in detached rice leaves incubated in darkness
Treatment Control DFMO, 0.5 HIM DFMA, 0.5 HIM Control DCH, 2.5 TOM MGBG, 5.0 mu Ethylene (nlg-'h-1) 6.4±0.2 6.0 + 0.3 6.2±0.4 6.4±0.2 8.0±0.2 8.8±0.2 Put 250 ± 3 240± 4 126±10 176±9 234 ± 6 240±l Spd (nmolg-') 153±2 148±2 155±9 130±4 137±2 123 ±1 Spm 50±2 59±5 53±3 98±1 86±3 64±2 Rates of production of ethylene and levels of polyamines were determined 24 h after the start of incubation.
o^ o o> E 40 2 0 CHLOROPHYLL J I I I
Fig. 6 Effects of ABA on levels of Chi and protein in detached rice leaves incubated in darkness. Means ± SE, 4 replicates.
Exogenous applications of Spd and Spm retard senes-cence of detached rice leaves (Cheng and Kao 1983). If Spd and Spm play a role in the regulation of senescence of leaves, a decrease in levels of Spd and/or Spm would be ex-pected. However, such a decrease was not observed. The increase in levels of Spd and Spm observed in the present study simply reflects the changing metabolic requirements after detachment of leaves.
Our results shows that levels of Put in detached rice leaves decrease with increasing duration of dark incuba-tion. Moreover, the decrease in levels of Put occurs prior to loss of Chi and degradation of protein. Although it seems possible that Put might be involved in the control of senescence of detached rice leaves, this possibility was not confirmed by the results of the application of inhibitors of the biosynthesis of polyamines to detached rice leaves. DFMA and DFMO significantly decreased levels of Put without affecting senescence of detached rice leaves. By contrast, treatments with DCH and MGBG resulted in elevated levels of Put in detached rice leaves but also pro-moted senescence. The evidence presented here leads to the conclusion that a lowering of the level of Put is unlikely to be the factor responsible for the senescence of detached rice leaves in darkness. This conclusion is further sup-ported by the observations that (a) BA, which retards senes-cence of detached rice leaves in the dark, decreased levels of Put; and (b) ABA, which promots senescence, increased
4 0 0
3 0 0
Fig. 7 Effects of ABA on levels of polyamines in detached rice leaves incubated in darkness. Means ± SE, 3 replicates.
Table 2 Effects of DFMO, DFMA, DCH and MGBG on levels of Chi and protein in detached rice leaves incubated in darkness Treatment Chl_ (mgg"1) Protein (mgg"1) Control DFMO, 0.5 HIM DFMA, 0.5 HIM Control DCH, 2.5 mM MGBG, 5.0 mM 2.6±0.1 2.4±0.1 2.8±0.2 2.6±0.1 0.9±0.1 1.5±0.1 2 0 ± l 22±1 19±1 2 0 ± l 13±1 16±1 Levels of Chi and protein levels were determined 3 days after the start of incubation.
the level of Put. Birecka et al. (1984, 1990) also reported that a decline in levels of polyamines was not correlated with the onset of senescence. The present investigation provides support for the suggestion of Birecka et al. (1984) that endogenous polyamines may have effects that differ from the possibly nonspecific effects frequently reported for exogenous polyamines.
MGBG, an inhibitor of SAMDC in bacterial and mam-malian cells (Willams-Ashman and Schenone 1972), has recently been used in a number of investigations on poly-amine metabolism in plants. This compound inhibits SAMDC activity and reduces levels of Spd in wild carrot and maize (Feirer et al. 1985, Hirawasa and Suzuki 1983). Although we have demonstrated that SAMDC activity from rice leaves is strongly inhibited by MGBG (data not shown), it was the level of Spm rather than that of Spd that was reduced by the treatment with MGBG. DCH is a com-petitive inhibitor of Spd synthase (Hibasami et al. 1980). The inhibition of Spd synthase by DCH causes a reduction in levels of both Spd and Spm in cultured cotyledons of
Pinus radiata and germinating seeds of Acer (Biondi et al.
1988, Walker et al. 1985). In other systems no changes or increases in levels of Spm have been observed (Mitchell et al. 1985, Torrigiani et al. 1987). In the current study we found, however, that only levels of Spm were reduced by treatment of rice leaves with DCH. There is one report that DCH is not an inhibitor of Spd synthase (Batchelor et al. 1986).
Since ethylene and polyamines share SAM as a com-mon precursor (Miyazaki and Yang 1987), the changes in levels of polyamines should be inversely correlated with changes in rates of production of ethylene during senes-cence of detached rice leaves. However, this corrleation was not observed, and it appears that biosynthesis of poly-amines and ethylene does not involve active competition for the substrate, SAM, during the course of senescence.
A dramatic increase in the rate of production of ethylene was observed prior to senescence of detached rice leaves, an observation that is in good agreement with our earlier conclusion that senescence of detached rice leaves is directly linked to the production of ethylene (Kao and Yang 1983). This conclusion is further supported by the observations that (a) neither production of ethylene nor se-nescence of detached rice leaves was affected by DFMO and DFMA, and (b) treatments with DCH and MGBG, which resulted in an increase in the production of ethylene, pro-moted senescence of detached rice leaves. Since DCH and MGBG reduced levels of Spm and increased those of Put in rice leaves, the stimulatory effect of DCH and MGBG on the production of ethylene is most likely mediated by the redirection of a portion of the pool of SAM into the ethylene pathway. Characterization of the DCH- and MGBG-induced production of ethylene will be published elsewhere.
This work was supported financially by the National Science Council of the Republic of China (NSC 79-0409-B002-04). We thank Dr. Peter McCann and the Merrell Dow Research Institute, incinnati, Ohio, for the gift samples of DFMO and DFMA.
References
Alt man, A. (1982) Retardation of radish leaf senescence by poly-amines. Physiol. Plant. 54: 189-193.
Batchelor, K. W., Smith, R. A. and Watson, N. S. (1986) Dicyclohexylamine is not an inhibitor of spermidine synthase. Biochem. J. 233: 307-308.
Biondi, S., Torrigiani, P., Sansovini, A. and Bagni, N. (1988) Inhibition of polyamine biosynthesis by dicyclohexylamine in cultured cotyledons of Pinus radiata. Physiol. Plant. 72: 471— 476.
Birecka, H., Birecki, M. and Ireton, K. (1990) Endogenous poly-amine levels and dark-induced leaf senescence. Plant Physiol. 93': S-757.
Birecka, H., DiNolfo, T. E., Martin, W. B. and Frohlich, M. W. (1984) Polyamines and leaf senescence in pyrolizidine alkaloid-bearing Heliotropium plants. Phytochemistry 23: 991-997. Carbonell, J. and Navarro, J. C. (1989) Correlation of spermine
levels with ovary senescence and with fruit set and development in Pisum sativum L. Planta 178: 482-487.
Chen, C. T., Chou, I. T. and Kao, C. H. (1990) Senescence of rice leaves XX. Changes of proton secretion during senes-cence. Plant Sci. 66: 29-34.
Cheng, S. H. and Kao, C. H. (1983) Localized effect of poly-amines on chlorophyll loss. Plant Cell Physiol. 24: 1463-1467. Evans, P. T. and Malmberg, R. L. (1989) Do polyamines have roles in plant development? Annu. Rev. Plant Physiol. Plant Mol. Biol. 40: 235-269.
Feirer, R. P., Wann, S.R. and Einspahr, D. W. (1985) The effects of spermidine synthesis inhibitors on in-vitro plant devel-opment. Plant Growth Regul. 3: 319-327.
Flores, H. E. and Galston, A. W. (1982) Analysis of polyamines in higher plants by high performance liquid chromatography. Plant Physiol. 69: 701-706.
Galston, A. W., Altman, A. and Kaur-Sawhney, R. (1978) Poly-amines, ribonuclease and the improvement of oat leaf proto-plasts. Plant Sci. Lett. 11: 69-79.
Hemantaranjan, A. and Garg, O. K. (1984) Effects of cytokinins and several inorganic cations on the polyamine content of (Hordeum vulgare L.) endosperms. Indian J. Exp. Biol. 22: 379-381.
Hibasami, H., Tanaka, M., Nagai, J. and Ikeda, T. (1980) Dicyclohexylamine, a potent inhibitor of spermidine synthetase in mammalian cells. FEBS Lett. 116: 99-101.
Hirawasa, E. and Suzuki, Y. (1983) Biosynthesis of spermidine in maize seedlings. Phytochemistry 22: 103-106.
Kao, C. H. (1978) Senescence of rice leaves II. Antisenescent ac-tion of cytokinins. Proc. Natl. Sci. Counc. ROC 2: 391-398. Kao, C. H. and Yang, S. F. (1983) Role of ethylene in the
senes-cence of detached rice leaves. Plant Physiol. 73: 881-885. Kaur-Sawhney, R. and Galston, A. W. (1979) Interaction of
poly-amines and light on biochemical processes involved in leaf senes-cence. Plant Cell Environ. 2: 189-196.
Kaur-Sawhney, R., Shih, L. M., Flores, H. E. and Galston, A. W. (1981) Relation of polyamine synthesis and titer to age-ing and senescence in oat leaves. Plant Physiol. 69: 405-410. Lin, P. P. C. (1984) Polyamine metabolism and its relation to
response of the aleurone layers of barley seeds to gibberellic acid. Plant Physiol. 74: 975-983.
Lowry, O. H., Rosebrough, N. J., Farr, A. L. and Randall, R. J. (1951) Protein measurement with the Folin-phenol reagent. J. Biol. Chem. 193: 265-275.
Mitchell, J. L. A., Wahan, D. W., McCann, P. P. and Qasba, P. (1985) Dicyclohexylamine effect on HTC cell polyamine con-tent and ornithine decarboxylase activity. Biochim. Biophys. Acta 840: 309-315.
Miyazaki, J. and. Yang, S. F. (1987) The methionine salvage path-way in relation to polyamine biosynthesis. Physiol. Plant. 69: 366-370.
Roberts, D. R., Walker, M. A., Thompson, J. E. and Dumbroff, E. B. (1984) The effects of inhibitors of polyamines and ethylene biosynthesis on senescence, ethylene production and polyamine levels in cut carnation flowers. Plant Cell Physiol. 25: 315-322.
Shibaoka, H. and Thimann, K. V. (1970) Antagonism between kinetin and amino acids: experiments on the mode of action of cytokinins. Plant Physiol. 46: 212-220.
Srivastava, S. K., Raj, D. S. and Naik, B. I. (1981) Polyamine
metabolism during ageing & senescence of pea leaves. Indian J. Exp. Biol. 19: 437-440.
Suresh, M. R. and Adiga, P. R. (1978) Absence of parallelism be-tween polyamine and nucleic acid contents during induced growth of cucumber cotyledongs. Biochem. J. 172: 185-188. Suresh, M. R., Ramakrishna, S. and Adiga, P. R. (1978)
Regula-tion of arginine decarboxylase and putrescine levels in Cucumis sativus cotyledons. Phytochemistry 17: 57-63.
Torrigiani, P., Serafini-Fracassini, D. and Bagni, N. (1987) Poly-amine biosynthesis and effect of dicyclohexylPoly-amine during the cell cycle of Helianthus tuberosus tuber. Plant Physiol. 84:
148-152.
Walker, M.A., Roberts, D. R. and Dumbroff, E. B. (1988) Effects of cytokinin and light on polyamines during the greening response of cucumber cotyledons. Plant Cell Physiol. 29: 201-205.
Walker, M. A., Roberts, D. R., Shih, C. Y. and Dumbroff, E. B. (1985) A requirement for polyamines during the cell-division phase of radicle emergence in seeds of Acer saccharurri. Plant Cell Physiol. 26: 967-971.
Williams-Ashman, H. and Schenone, A. (1972) Methylglyoxal 6is-(guanylhydrazone) as a potent inhibitor of S-adenosyl-methionine decarboxylases. Biochem. Biophys. Res. Com-mun. 46: 288-295.
Wintermans, J. F. G. M. and De Mots, A. (1965) Spectrophoto-metric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim. Biophys. Acta 109: 448-453.