行政院國家科學委員會專題研究計畫 成果報告
利用噬菌體表達人腦 cDNA 基因庫研究日本腦炎病毒蛋白與
宿主細胞因子之功能性交互作用
計畫類別: 個別型計畫 計畫編號: NSC92-2314-B-039-030- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 中國醫藥大學醫事技術學系 計畫主持人: 林振文 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 11 月 2 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※利用噬菌體表達人腦 cDNA 基因庫研究日本腦炎病毒蛋白與宿主
細胞因子之功能性交互作用
※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※※計畫類別:■個別型計畫
□整合型計畫
計畫編號:NSC92-2314-B-039-030
執行期間:92 年 8 月 1 日至 93 年 7 月 31 日
計畫主持人:林振文
執行單位:中國醫藥大學
中
華
民
國
93
年
10
月
29
日
行政院國家科學委員會專題研究計畫成果報告
計畫編號:NSC92-2314-B-039-030
執行期限:92 年 8 月 1 日至 93 年 7 月 31 日
主持人:
林振文 中國醫藥大學醫學檢驗生物技術系
Abstract
JEV NS2B-NS3 protease was associated with neurovirulence, while neurovirulence is associated with the apoptotic level of the brain cells. The apoptosis-induced ability and the cellular protein interaction of the JEV NS2B-NS3 protease could be important for elucidating the molecular level of the JEV neurovirulence. In this study, we investigated the ability of the JEV NS2B-NS3 to induce apoptosis in brain cell lines, and further identified the interaction of the JEV NS2B-NS3 with specific cellular proteins using phage display
technology. Cell necrosis was found in transfected Neuro-2a cell with the
NS2B-NS3-EGFP in-frame fusion gene, whereas Annexin V staining also indicated that the apoptotic cell was found in the transfected cells with the NS2B-NS3-EGFP gene. Nine cDNA sequences displayed NS2B-NS3 interacting phage clones were determined using direct nucleotide sequencing, and their deduced amino acid sequences could be grouped into the major family and the miscellaneous family. The major family was characterized by the conserved repeat sequence ERKK, and matched with five different proteins in the database of GenBank + EMBL + DDBJ sequences from EST division. Two of these five proteins, DRPLA protein and U1 small ribonucleoprotein 1 (SNRP) homolog were proposed to relate with the enhancement of apoptosis. Furthermore, the binding interaction of recombinant NS2B-NS3 interacting proteins with the JEV NS2B-NS3 protease was confirmed by coimmunoprecipitation and ELISA.
Introduction
Japanese encephalitis virus (JEV) is a number of the Flaviviridae family, including other important human pathogens such as dengue virus, yellow fever, St. Louis encephalitis virus, Murray valley encephalitis virus, Langat virus, West Nile virus, and tick-borne encephalitis
(TBE) virus (Burk and Monath, 2001; Lindenbach and Rice, 2001). JEV is a
mosquito-borne neurotropic flavivirus (Burk and Monath, 2001), causing severe central nerve system diseases such as poliomyelitis-like acute flaccid paralysis, aseptic meningitis and encephalitis (Solomon et al., 2000). Japanese encephalitis (JE) has a high fatality rate of 30% and around half of the JE survivors have severe neurological sequela (Solomon et al., 2000). Approximately 50000 JE cases with 10000 deaths are reported annually in Asian countries (Tsai, 1997; Burk and Monath, 2001).
The JEV genome is a single-stranded, positive-sense RNA with about 11,000 nucleotides in length (Lindenbach and Rice, 2001). The single, long ORF of the JEV genome encodes for three structural proteins (capsid (C), membrane (prM/M), and envelope (E)) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5). The mature viral proteins are produced through proteolytic processing of the single polyprotein with the viral serine protease (NS2B-NS3) and various cellular proteases (Lindenbach and Rice, 2001). NS3 (about 70 kDa) is involved with several enzyme activities, including serine protease, helicase, and RNA triphosphatase (Rice, 1996; Lindenbach and Rice, 2001). The N-terminal one third of NS3 contains His51, Asp75, and Ser135 as the protease active sites (Ryan et al., 1998), while NS2B is the cofactor for the viral serine protease NS3 (Chambers et al., 1993; Falgout et al., 1993). Sequence analysis of the JEV strains indicated that mutations at NS2B-NS3 correlated with the neurovirulence (Ni et al., 1995; McMinn, 1997; Monath, 2001), while neurovirulence is associated with the apoptotic level of the brain cells (Liao et al., 1997; Jan et al., 2000). Recent, NS2B-NS3 protease of other flaviviruses,
such as dengue viruses and Langat virus, has been demonstrated to induce apoptosis (Prikhod’ko et al., 2002; Shafee and Abubakar, 2003), being in line with the apoptosis induced by a number of other viral proteases (Li et al., 2002; Barco et al., 2000). Therefore, the apoptosis-induced ability and the cellular protein interaction of the JEV NS2B-NS3 protease could be important for elucidating the molecular level of the JEV neurovirulence.
Phage display provides a powerful tool for the study of the ligand-protein interaction from in vitro selection of cDNA libraries with the target protein (Smith, 1995; Johnsson and Ge, 1999; Benhar, 2001). Each unique protein in a cDNA library could be expressed and fused with the coat protein gene VI in M13 phage (Hufton et al., 1999), the head protein D in phage (Santini et al., 1998) and the capsid protein gene 10B in T7 phage (Yamamoto et al., 1999). Phage display cDNA libraries have been demonstrated to identify the binding proteins of the interest ligands, such as the 2-methylnorharman- interacting brain proteins (Gearhart et al., 2002) and the adaptor protein 2-assaociated kinase (Conner and Schmid, 2002). Screening cDNA libraries displayed on the phage surface appears to be convenient, rapid and effective for identification of the interest ligand-interacting proteins.
In this study, we investigated the ability of the JEV NS2B-NS3 to induce apoptosis in brain cell lines, and further identified the interaction of the JEV NS2B-NS3 with specific cellular proteins using phage display technology.
Materials and methods Viruses and cells
The JEV strain T1P1 was used in this study as previously described (Chiou and Chen, 2001). Neuro-2a cells were grown in minimum essential medium (MEM) with 2 mM L-glutamine, 1mM sodium pyruvate and 10 % fetal bovine serum (FBS).
Cloning and expression analysis of the JEV NS2B-NS3 protease in Neuro-2a cells
For monitoring the expression in Neuro-2a cells, the NS2B-NS3 protease was fused in-frame with at the N-terminus of the enhanced green fluorescent protein (EGFP). The
EGFP gene was generated by PCR with the primers
5'-CGGCCGCTCGAGATGGTGAGCAAGGGCGAGGAG-3' and
5'-GGGCCCTCTAGACTTGTACAGCTCGTCCATGCC-3'. This EGFP PCR product was
digested with XhoI and XbaI, and ligated to the XhoI-XbaI site of
pcDNA3.1-pcDNA3.1/HisC (Amersham) to generate the vector pcDNA3.1-EGFP. The
NS2B-NS3 gene derived from the nucleotides 4215 to 5147 of the JEV T1P1 genome (GenBank No. AF254453) was amplified using RT-PCR with the paired primers
5’-ATCCGAATTCGGGTGGCCAGCTACTGAGTTTTTG-3’ and
5’-GGTGCTCGAGGGTGTAAGCTTCTGGGACTGGTTC-3’. The forward primer
mentioned above contained an EcoRI restriction site, and the reverse primer included an XhoI restriction site. The NS2B-NS3 PCR product was digested with EcoRI and XhoI, and then cloned into the pcDNA3.1-EGFP to construct the plasmid pcDNA3.1-NS2B-NS3-EGFP. Neuro-2A cells at 60-90% confluency in 6-well plates were transfected with 5 g of
pcDNA3.1-EGFP or pcDNA3.1-NS2B-NS3-EGFP using the GenePorter reagent.
According to manufacturer’s direction (Gene Therapy Systems, San Diego,CA). The
5-hour incubation with the mixture of the plasmid DNA and GenePorter reagent, then were selected using the DMEM containing 10 % FBS and 200 µg/ml of G418. The expression of the JEV NS2B-NS3 in Neuro-2a was detected by the expression of the fusion partner protein EGFP using fluorescence microscopy and Western blotting analysis with anti-His tag antibodies. The apoptotic cells were detected using Annexin V staining.
Fluorimetric assay of caspase profiling
Transfected Neuro-2a cells were harvested after 5 days of the G418 selection for measuring the activity of caspase-2, 3, 8 and 9 using BD ApoAlert Caspase Fluorescent Assay Kits (BD Biosciences). 2 × 105cells per well were lysed in 50 μl of lysis buffer and centrifuged at 12,000 g for 5 min at 4 ℃. Supernatant was transferred into the well of the caspase profiling assay plate and incubated at 37 ℃ for 2 hr. Finally, the caspase profiling assay plate was analyzed in the fluorescent plate reader with excitation at 380 nm and emission at 460 nm.
Purification of recombinant NS2B-NS3 protease synthesized in E. coli
The NS2B-NS3 protease gene was amplified by RT-PCR and cloned into the
EcoRI-XhoI site of the bacterial expression vector pET24a (Novagen) to generate the plasmid
pET24a-NS2B-NS3. The E. coli strain BL21(DE3) was transformed with the plasmid
pET24a-NS2B-NS3, and grown overnight in LB medium in presence of 100 μ g of ampicillin per ml. The cells were then diluted 50-fold in LB medium containing ampicillin and grown at 37 ℃. Once the cultures reached an absorbance at 600nm of 0.6, they were induced by the addition of 0.5mM isopropyl-β-D-thiogalactopyranoside (IPTG) at 20 ℃ for 4 hr. Finally, the bacteria were harvested by centrifugation at 9,000 rpm for 15 min at 4 ℃, and resuspended in the denature buffer (10 mM imidazole, 8 M urea and 1mM
-mercaptoethanol) for sonication. The supernatant after centrifugation (10,000 x g for 20 min) was incubated with the refolding buffer (10 mM imidazole and 1mM-mercaptoethanol)
at 4 ℃ for 3 hr, and then purified with the His·Bind Resin (Novagen). The eluted
recombinant NS2B-NS3 protease was further dialyzed against phosphate-buffered saline.
Biopanning of phage display brain cDNA libraries with the NS2B-NS3 protease
A brain cDNA library fused to the C terminus of capsid protein 10B in T7 phages was
purchased from Novagen (Madison, WI). Five rounds of biopanning for screening
high-affinity phage clones were carried out with recombinant NS2B-NS3 protease coated onto microplates (5g per well). After blocking with 5 mg/ml BSA in TBST (Tris-buffered saline, pH 7.5, 0.1% Tween-20), the phage clones reacted with mAb E3.3 in TBST (Tris-buffered saline, pH 7.5, 0.1% Tween-20), and eluted with the soluble NS2B-NS3 protease. The nucleotide sequences of cDNA displayed by selected phages were directly amplified using the T7 Select UP primer 5'-GGAGCTGTCGTATTCCAGTCA-3' and determined using the sequencing with the ABI PRISM 377 DNA Sequencer (Perkin-Elmer, USA).
Direct ELISA binding assays
The selected phage clone (1010 p.f.u./well) were coated onto microplates for determining the binding reaction with the NS2B-NS3 protease for 1 h at room temperature. After washing with TBST, bound NS2B-NS3 protease was detected using the ELISA with the anti-His tag monoclonal antibody and anti-mouse IgG antibodies conjugated to
peroxidase (Pharmacia). ELISA product was developed using a chromogen solution
containing ABTS (2,2’-azino-di-(3-ethylbenzthiazoline-6-sulfonate) and hydrogen peroxide and then measured at A405 nm.
Expression of recombinant NS2B-NS3-inteacting proteins
The cDNA displayed by selected phages were amplified by PCR with the pair primer
of T7 Select UP primer and the following reverse primers:
5’-CCGCAAGCTTATGGACAGATATCATGAC -3’for the phage clone NO. 24 and
5’-CCGCAAGCTTGGCAGGGCTGTCTTTCTT -3’for phage clone NO. 46. The PCR
product was digested with EcoRI and Hind III, and then cloned into the EcoR I/Hind III site of the expression vector pET43.1b (Novagen). The resultant plasmid was transformed into the E. coli strain BL21(DE3). The NS2B-NS3 interacting protein was generated in E. coli and fused with the Nus tag and His tag at its N-terminus.
Co-immunoprecipitation and Wstern blotting
The mixture of the NS2B-NS3-ineracting protein and JEV NS2B-NS3 protease was first incubated with anti-Nus tag mAb for 4 h at 4 ℃, and then reacted with the protein A-Sepharose beads for the addition of 2-h incubation. After centrifugation, the pellet was washed four time with NET buffer (150 mM NaCl, 0.1 mM EDTA, 30 mM Tris-HCl, pH 7.4). The immunoprecipitate were dissolved in 2X SDS-PAGE sample buffer without 2-mercaptoethanol and boiled for 10 min. Proteins were resolved on 12% SDS-PAGE gels and electrophoretically transferred to nitrocellulose papers. The resultant blots were blocked with 5% skim milk, and then reacted with the anti-His tag mAb for a 1-h incubation. The blots were then washed with TBS three times and overlaid with a 1/1000 dilution of rabbit anti-mouse IgG antibodies conjugated with alkaline phosphatase (KPL). Following another1-h incubation at room temperature, the blots were developed with NBT/BCIP (Invitrogen).
Results and Discussion
JEV NS2B-NS3 protease induces apoptosis in Neuro-2a cells
The expression of the NS2B-NS3 protease that was fused at the N-terminus of EGFP was examined by direct fluorescent signal of the transfected cells (Fig 1, B and D). Cell necrosis was found in transfected Neuro-2a cell with the NS2B-NS3-EGFP in-frame fusion gene (Fig 1, D and F), but not in the tranfected cells with the EGFP gene only (Fig 1, A, C, and E). Annexin V staining also indicated that the apoptotic cell was found in the transfected cells with the NS2B-NS3-EGFP gene (Fig 2, B), but not in the tranfected cells with the EGFP gene only (Fig 2, B).
Purification of recombinant NS2B-NS3 protease
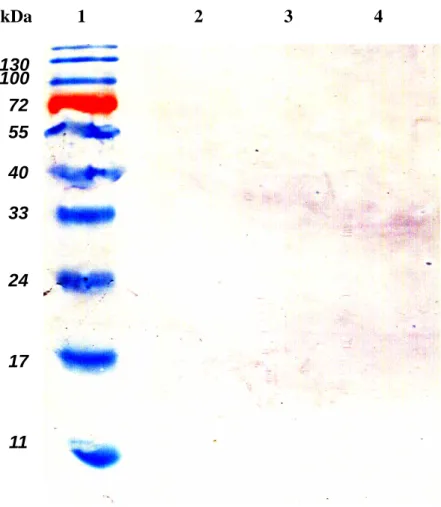
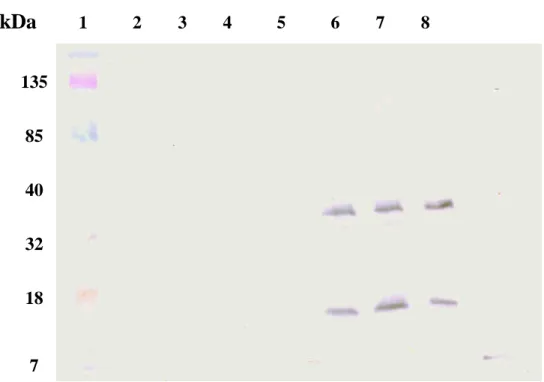
The NS2B-NS3 protease synthesized in E. coli was purified using the immobilized metal affinity chromatography. Western blotting with anti-His tag mAb demonstrated that a 34-kDa band for the NS2B-NS3 protease and a 19-kDa band for the NS3 protein E protein were observed at the eluted fraction with 400 mM imidazole (Fig. 4, lanes 6-8). The result also indicated the cis-cleavage activity of the purified NS2B-NS3 protease.
Selection of high affinity NS2B-NS3-interacting cDNA phage clones
To identify the NS2B-NS3-interacting cDNA clones, a human brain cDNA library displayed on the T7 phage surface was screened by the binding affinity to recombinant NS2B-NS3 protease. After six rounds of biopanning procedures, fifty phage clones were
selected for determining the reactivity of their corresponding phage-displayed
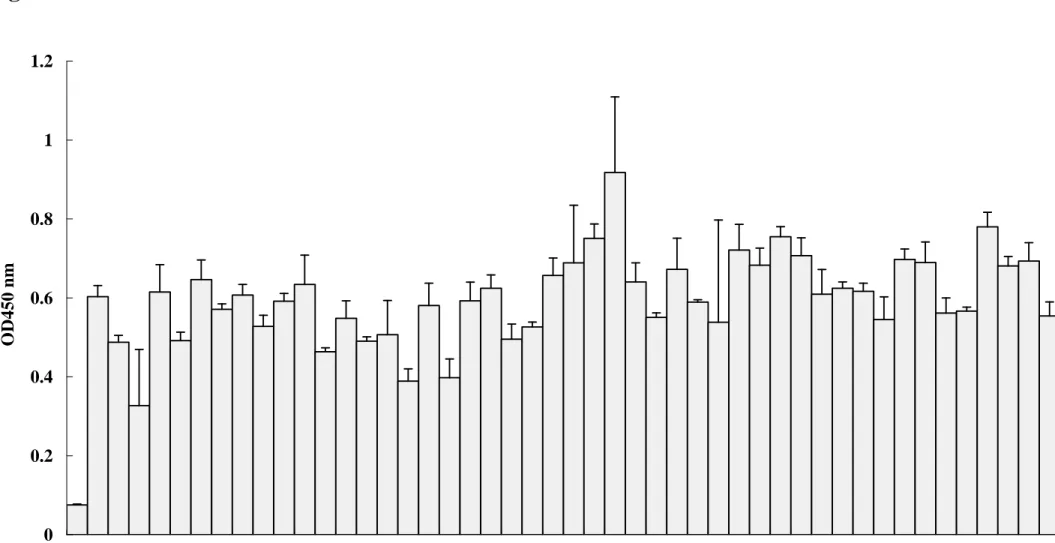
cDNA-encoded proteins with NS2B-NS3 protease. Nine phage clones with high affinity to NS2B-NS3 protease were identified using direct ELISA (Fig 5). The selected phage clones showed approximately 6- to 9-fold increases in direct ELISA binding compared to wild type
phage T7 as a negative control (Fig 5).
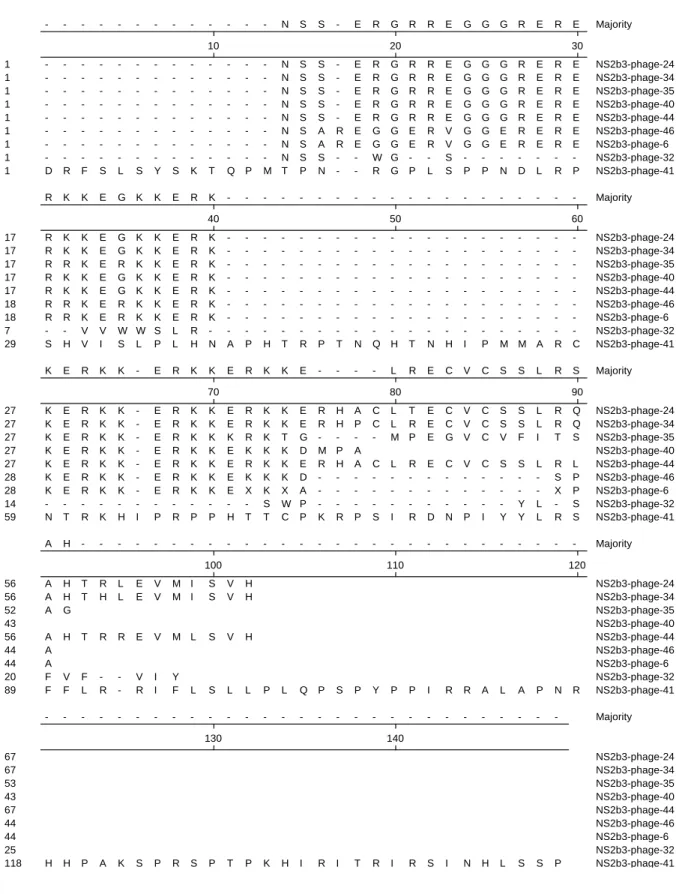
Sequence analysis of the brain cDNA displayed on the high-affinity phage clones
The cDNA sequences displayed on the C-terminus of phage coat protein 10B were determined using direct nucleotide sequencing. The deduced amino acid sequences of high-affinity cDNA clones were translated based on the indicated nucleotide sequences (Fig 6). Amino acid sequence alignment showed that sequences of these NS2B-NS3-interacting proteins could be grouped into the major family and the miscellaneous family. The major family contains 7 clones (phage NO. 6, 24, 34, 35, 40, 44, and 46), and has high homology at the N-terminus. The major family was characterized by the conserved repeat sequence ERKK. The miscellaneous family included two phage clones (NO. 32 and 41), showing no similarity to each other. The deduced amino acid sequence of the NS2B-NS3- interacting proteins was analyzed using the Protein-protein BLAST (blastp) based on the database of GenBank + EMBL + DDBJ sequences from EST division. Five different proteins with the ERKK repeat motif were found to show the similarity of the amino acid sequence to the major family of identified NS2B-NS3-interacting proteins (Table 1). The name, the NCBI accession number, and proposed or known functions were also listed in Table 1. The DRPLA protein was a candidate gene for dentatorubral-pallidoluysian atrophy (DRPLA) and has a high homology to the atrophin-1 protein. U1 small ribonucleoprotein 1 (SNRP) homolog showed a homology to the ACINUS (apoptotic chromatin condensation inducer in
the nucleus). These two proteins DRPLP and SNRP were proposed to relate with the
enhancement of apoptosis.
Binding of recombinant NS2B-NS3-interacting proteins to the JEV protease
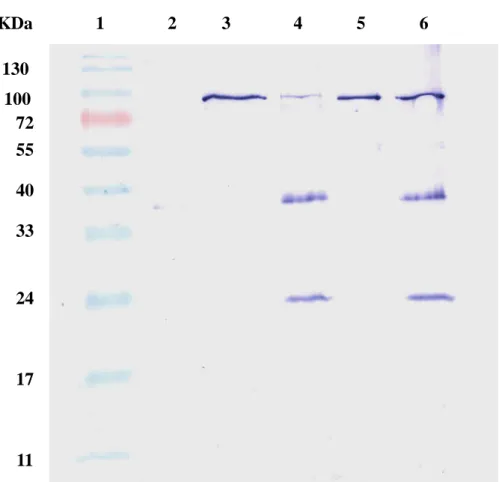
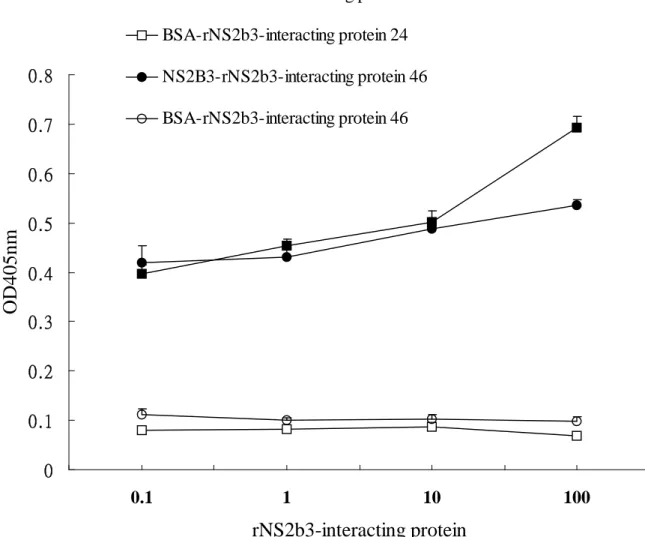
the JEV protease, recombinant NS2B-NS3-interacting protein were generated in E. coli and fused with the Nus tag and His tag at the N-terminus. After the incubation of the JEV protease and NS2B-NS3-interacting protein, recombinant NS2B-NS3-interacting protein was immunoprecipitated by using an anti-Nus tag antibody, and bound JEV protease was detected by immbunoblotting with an anti-His tag antibody (Fig 7). Coimmunoprecipitation assay indicated the JEV protease was found in the immunoprecipitates with NS2B-NS3-interacting proteins 24 and 46 (Fig 7, lanes 4 and 6), whereas on immunoprecipitating reaction was found in the interaction of the JEV protease with the protein A-sepharose beads (Fig 7, lane 2). Subsequently, the binding ELISA assay demonstrated a dose-dependent manner in the interaction of the JEV protease with recombinant NS2B-NS3 interacting proteins 24 and 46 (Fig 8).
Acknowledgements
We would like to thank China Medical University and National Science Council,
Taiwan for financial supports (CMU92-MT-03, CMU93-MT-04, and
References
Prikhod'ko GG, Prikhod'ko EA, Pletnev AG, Cohen JI. Langat flavivirus protease NS3 binds caspase-8 and induces apoptosis. J Virol. 2002 Jun;76(11):5701-10.
Shafee N, AbuBakar S. Dengue virus type 2 NS3 protease and NS2B-NS3 protease precursor induce apoptosis. J Gen Virol. 2003 Aug;84(Pt 8):2191-5.
Yamamoto M, Kominato Y, Yamamoto F. Phage display cDNA cloning of protein with carbohydrate affinity. Biochem Biophys Res Commun. 1999 Feb 16;255(2):194-9. Conner SD, Schmid SL. Identification of an adaptor-associated kinase, AAK1, as a
regulator of clathrin-mediated endocytosis. J Cell Biol. 2002 Mar 4;156(5):921-9. Danner S, Belasco JG. T7 phage display: a novel genetic selection system for cloning
RNA-binding proteins from cDNA libraries. Proc Natl Acad Sci U S A. 2001 Nov 6;98(23):12954-9.
Benhar I. Biotechnological applications of phage and cell display. Biotechnol Adv. 2001 Feb 1;19(1):1-33.
Hufton SE, Moerkerk PT, Meulemans EV, de Bruine A, Arends JW, Hoogenboom HR. Phage display of cDNA repertoires: the pVI display system and its applications for the selection of immunogenic ligands. J Immunol Methods. 1999 Dec 10;231(1-2):39-51. Santini C, Brennan D, Mennuni C, Hoess RH, Nicosia A, Cortese R, Luzzago A. Efficient
display of an HCV cDNA expression library as C-terminal fusion to the capsid protein D of bacteriophage lambda J Mol Biol. 1998 Sep 11;282(1):125-35.
Fig. 1
pcDNA3.1-EGFP
pcDNA3.1-NS2B-NS3-EGFP
A
B
C
D
E
F
Secondary day (200X) Fluorescence Fifth day (400X) Fluorescence Fifth day (400X) Phase contrastFig. 2
A
Fig. 3.
130 100 72 55 40 33 24 17 11 kDa 1 2 3 4Fig. 4.
135 85 40 32 18 7kDa
1 2 3 4 5 6 7 8Fig. 5.
0 0.2 0.4 0.6 0.8 1 1.2 blank1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 Phage NO. O D 4 5 0 n mFig. 6.
- - - N S S - E R G R R E G G G R E R E Majority 10 20 30 - - - N S S - E R G R R E G G G R E R E 1 NS2b3-phage-24 - - - N S S - E R G R R E G G G R E R E 1 NS2b3-phage-34 - - - N S S - E R G R R E G G G R E R E 1 NS2b3-phage-35 - - - N S S - E R G R R E G G G R E R E 1 NS2b3-phage-40 - - - N S S - E R G R R E G G G R E R E 1 NS2b3-phage-44 - - - N S A R E G G E R V G G E R E R E 1 NS2b3-phage-46 - - - N S A R E G G E R V G G E R E R E 1 NS2b3-phage-6 - - - N S S - - W G - - S - - - -1 NS2b3-phage-32 D R F S L S Y S K T Q P M T P N - - R G P L S P P N D L R P 1 NS2b3-phage-41 R K K E G K K E R K - - - Majority 40 50 60 R K K E G K K E R K - - - -17 NS2b3-phage-24 R K K E G K K E R K - - - -17 NS2b3-phage-34 R R K E R K K E R K - - - -17 NS2b3-phage-35 R K K E G K K E R K - - - -17 NS2b3-phage-40 R K K E G K K E R K - - - -17 NS2b3-phage-44 R R K E R K K E R K - - - -18 NS2b3-phage-46 R R K E R K K E R K - - - -18 NS2b3-phage-6 - - V V W W S L R - - - -7 NS2b3-phage-32 S H V I S L P L H N A P H T R P T N Q H T N H I P M M A R C 29 NS2b3-phage-41 K E R K K - E R K K E R K K E - - - - L R E C V C S S L R S Majority 70 80 90 K E R K K - E R K K E R K K E R H A C L T E C V C S S L R Q 27 NS2b3-phage-24 K E R K K - E R K K E R K K E R H P C L R E C V C S S L R Q 27 NS2b3-phage-34 K E R K K - E R K K K R K T G - - - - M P E G V C V F I T S 27 NS2b3-phage-35 K E R K K - E R K K E K K K D M P A 27 NS2b3-phage-40 K E R K K - E R K K E R K K E R H A C L R E C V C S S L R L 27 NS2b3-phage-44 K E R K K - E R K K E K K K D - - - S P 28 NS2b3-phage-46 K E R K K - E R K K E X K X A - - - X P 28 NS2b3-phage-6 - - - S W P - - - Y L - S 14 NS2b3-phage-32 N T R K H I P R P P H T T C P K R P S I R D N P I Y Y L R S 59 NS2b3-phage-41 A H - - - Majority 100 110 120 A H T R L E V M I S V H 56 NS2b3-phage-24 A H T H L E V M I S V H 56 NS2b3-phage-34 A G 52 NS2b3-phage-35 43 NS2b3-phage-40 A H T R R E V M L S V H 56 NS2b3-phage-44 A 44 NS2b3-phage-46 A 44 NS2b3-phage-6 F V F - - V I Y 20 NS2b3-phage-32 F F L R - R I F L S L L P L Q P S P Y P P I R R A L A P N R 89 NS2b3-phage-41 - - - Majority 130 140 67 NS2b3-phage-24 67 NS2b3-phage-34 53 NS2b3-phage-35 43 NS2b3-phage-40 67 NS2b3-phage-44 44 NS2b3-phage-46 44 NS2b3-phage-6 25 NS2b3-phage-32 H H P A K S P R S P T P K H I R I T R I R S I N H L S S P 118 NS2b3-phage-41Fig. 7.
130 100 72 55 40 33 24 17 11 KDa 1 2 3 4 5 6Fig. 8.
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.1 1 10 100rNS2b3-interacting protein
O
D
4
0
5
n
m
NS2B3-rNS2b3-interacting protein 24 BSA-rNS2b3-interacting protein 24 NS2B3-rNS2b3-interacting protein 46 BSA-rNS2b3-interacting protein 46Table 1.
Putative NS2BNS3-interacting protein (NCBI Accession )
NS2BNS3-interacting region within the indicated protein (Score)
Characteristics and/or functions of interacting protein
References
DRPLA protein (BAA07534.1) AREEKEREREREREKERE
REKER (103)
A candidate gene for dentatorubral-pallidoluysian atrophy (DRPLA); a high homology to the
atrophin-1 protein; found mainly in neuronal cytoplasm; enhances apoptosis
Yazawa et al., 1995; Waerner et al., 2001
HBV preS1 (21-47) binding protein (AAK58397.1) ERKKERKKERKKEKKER KKRKKKEKERKKE (138) Unknown U1 small ribonucleoprotein 1 SNRP homolog (AAF19255.1) EREREREREREREKEKER ERERERDRDRDRTKER (121)
A subunit of the spliceosome; a homology to the ACINUS (apoptotic chromatin condensation inducer in the nucleus)
Du and Rosbash, 2002; Ishikawa et al., 1998 S164 (AAC97961.1 ) EREREREREREREKEKER ERERERDRDRDRTKER (121)
Mapped at chromosome 14q24.3, the gene for presenilin 1
Voltage-gated sodium channel type XI alpha subunit (AAF17480.1)
WVIWWNLRKTCYQIVK HSWFESFIIFVI (67)
Voltage-dependent gating and conductance;
mediates brain-derived neurotrophic factor-evoked membrane depolarization
Hypothetical protein (XP_499069) GRKEGRKEGRKEGKKER
RKERKKERKKERKKER (180)