行政院國家科學委員會專題研究計畫 成果報告
克雷白氏肺炎桿菌第一型纖毛和第三型纖毛交互表現的調
控
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 98-3112-B-009-005- 執 行 期 間 : 98 年 08 月 01 日至 99 年 07 月 31 日 執 行 單 位 : 國立交通大學生物科技學系(所) 計 畫 主 持 人 : 彭慧玲 計畫參與人員: 碩士班研究生-兼任助理人員:曾品瑄 碩士班研究生-兼任助理人員:鄭崴云 碩士班研究生-兼任助理人員:徐豪君 博士班研究生-兼任助理人員:鄭新耀 報 告 附 件 : 出席國際會議研究心得報告及發表論文 處 理 方 式 : 本計畫可公開查詢中 華 民 國 99 年 08 月 05 日
行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
Regulation for the counter-expression of type 1 and type 3
fimbriae in Klebsiella pneumoniae
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 98-3112-B-009-005-
執行期間: 九十八 年 八 月 一 日 至 九十九 年 七 月 三十一 日
計畫主持人:彭慧玲
共同主持人:
計畫參與人員:黃盈蓉,吳健誠,王哲充,鄭崴云
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
▓赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
■出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,■一年□二年後可公開查詢
執行單位: 交通大學生物科技學系
中 華 民 國 九十九 年 七 月 三十 日
TABLE OF CONTENTS
I. Abstracts
Ia.
Abstract
in
Chinese page
II
Ib.
Abstract
in
English page
III,
VI
II. Background and Significance
page
1
III.
Material
and
Methods
page
5
IV. Results and Discussion
page 7
V. Literature Cited
page 14
VI. Self-Assessment
page 19
I. Abstract
Ia. Abstract in Chinese
關鍵詞:克雷白氏肺炎桿菌,第一型纖毛與第三型纖毛的對立表現、纖毛與莢膜多 醣表現的交互調控。 細菌纖毛藉由其黏附特異性與特定的細胞結合,為其造成感染的關鍵步驟。已 知,第一型或第三型纖毛是克雷白氏肺炎桿菌臨床分離株表現的主要黏附因子。我 們計劃探討調控第一型纖毛和第三型纖毛的對立表現(counter-expression)的機制, 並且建立負責纖毛和莢膜多醣表現之交互調控的作用關係。過去一年的計劃成果如 下: 目標一、建構註解為調控基因的特定基因缺損株,探討其對此二纖毛對立表現的 影響:利用抗MrkA的血清進行西方墨點法分析和酵母菌凝集試驗顯示:位於此二纖 毛基因組間的pecS或pecM基因缺損會提升第三型纖毛的表現;而可以轉譯成EAL功 能蛋白的mrkJ(KP4551)和PilZ功能蛋白的mrkH(KP4554)基因缺損對MrkA蛋白 的表現量或對克雷白氏肺炎桿菌凝集酵母菌的活性並沒有顯著的影響;然而,破壞 GerE家族的轉錄因子基因mrkI(KP4552)或csgD可以明顯提高其凝集酵母菌的能 力,mrkI的缺損更阻斷了MrkA的表現,此結果顯示PecS/M可能是第三型纖毛的負向 調控蛋白、MrkI可能是調控第一型纖毛和第三型纖毛對立表現的因子,CsgD可能負 向調控甘露糖以外的含糖黏附蛋白的表現。
目標二、建構跳躍子突變基因庫(transposon-insertion mutant library),篩選可 能的調控基因:之前的實驗結果顯示,破壞可以調控細菌夾膜產生的調控基因rcsB, 會增加第一型纖毛的表現,相反的會降低MrkA蛋白的表現。我們以定量PCR證實促 進莢膜多醣表現的調控蛋白RcsB負向影響FimB的轉錄,進而影響第一型纖毛和 FimK的表現,最後藉由FimK影響二次訊息分子c-di-GMP的濃度而調控第三型纖毛 的表現。為了篩選某個因子因而rcsB缺損而降低MrkA蛋白的表現,我們以帶有 PmrkA-LacZ報告系統的CG43S3Z01rcsB缺損株為帶有跳躍子質體的接受株,以X-Gal 培養盤篩選顏色變化的突變株。核酸定序分析結果顯示兩株增加LacZ活性的插入點 分別是glycosidase和lacI基因,而兩株降低LacZ活性的插入點分別是carbon starvation protein A和未知功能的yehZ基因,這些結果顯示此跳躍子突變效率仍需提高。
目標三、探討外膜逆境反應(Envelope stress responses)的調控在此二纖毛對立 表現中可能扮演的角色:我們以酵母菌凝集活性、MrkA蛋白表現、生物膜形成等測
試分析已取得的外膜孔蛋白OmpK35(OmpF)、OmpK36(OmpC)、OmpK37(OmpN)
或OmpA基因缺損株,結果顯示這些缺損株凝集酵母菌的活性差異並不明顯;而 OmpC或OmpA基因缺損會降低MrkA蛋白的表現;另外,這些外膜孔蛋白的缺失對 生物膜的形成也有不同的影響。
I. Abstract
Ib. Abstract in English
Keywords: Klebsiella pneumoniae, counter-expression of type 1 and type 3 fimbriae, interacting regulation with CPS biosynthesis
Fimbriae (also called pili) play a key role for bacteria to attach to specific host cell during the establishment of an infection. The expression of type 1 or type 3 fimbriae in the clinical isolates of Klebsiella pneumoniae has been commonly reported. We have proposed to explore the regulatory mechanisms involved in the control of
counter-expression between type 1 and type 3 fimbirae and interacting regulation between fimbriae and CPS biosynthesis. The accomplished experiments in a year are as following:
1) To investigate if the deletion of the specific regulatory genes affects the expression: The analysis of western hybridization against MrkA antibody and yeast agglutination revealed: Deletion of phgS or phgM increased the expression of MrkA; The deletion of mrkJ (KP4551), encoding EAL domain protein, or mrkH (KP4554), encoding PilZ domain protein had no apparent effect on the YA activity or MrkA expression. However, deletion of csgD or mrkI (KP4552), which encoding a
GerE-family transcription factor, apparently increased the YA activity. On the other hand, the expression of MrkA was abolished in mrkI mutant. These results suggest PecS/M is a negative regulatory system for the expression of type 3 fimbriae, MrkI is the regulator for the counter expression of the two fimbriae, and CsgD probably controls the expression of a mannose-resistant adhesin.
2) We have used qPCR analysis to show that RcsB, an activator for the CPS synthesis, negatively affect the expression of FimB and hence the expression of type 1 fimbriae and FimK. The expression of FimK appeared to modulate level of the second
messenger c-di-GMP which in turn affects the expression of MrkA. In the meantime,a transposon-insertion mutant library derived from CG43S3Z01rcsB- strain carrying
PmrkA-LacZ was generated, and 8 mutants with color changes on the X-gal plate had
been isolated. The sequences determined revealed that the disrupted genes include the gene encoding a putative glycosidase and lacI, and the gene encoding carbon starvation protein and an unknown function protein YehZ. The efficiency of Tn-mutagenesis has to be improved, however.
3) To investigate if envelope stress plays a role in the regulation of the
counter-expression: The mutants with deletion of either porin gene encoding Ompk35 (OmpF), Ompk36 (OmpC), Omp37 (OmpN) or OmpA were generated. As assessed using yeast agglutination activity, MrkA expression, and biofilm formation activity, no
apparent deletion effect on the yeast agglutination activity was observed for any of the mutants. However, either ompC or ompA deletion was found to reduce the expression of MrkA. Moreover, either of the gene deletion affected differently the biofilm formation activity.
II. Background and Significance
Klebsiella pneumoniae (KP) infections
As a common nosocomial pathogen, Klebsiella pneumoniae (KP) causes suppurative lesions, septicemia, and urinary and respiratory tract infections in immunocompromised patients (27, 41). In Taiwan, KP is the leading cause of liver abscess in patients with diabetes mellitus. A steady increase of the incidence of Klebsiella Liver Abscess (KLA) in the patients of diabetes, malignancy, renal disease, or pneumonia was reported (5, 18, 32, 58, 61). Reports of KLA in western countries have also been accumulating (15, 29). Virulence traits involved in the KLA pathogenesis including K1 capsule, magA, rmpA, and type 1 fimbriae of the hypermucoviscous KP strains have been implicated (4, 7, 34, 36, 64). KP adhesins
Fimbriae (also called pili) are the best understood structures for the attachment to host cells to establish a persistent infection. Among the five adhesins identified in KP, type 1 fimbriae are frequently present in the pyelonephritis isolates of (44, 53, 54). Type 3 fimbriae, referred to as the mannose-resistant Klebsiella hemagglutin (Mr/KH), have also been shown in some uropathogenic isolates of KP (56). The nonfimbrial adhesin CF29K, which is involved in the adherence to human intestinal cell line Caco-2 (9), and a fimbrial antigen KPF-28 that has been shown to be a determinant for the colonization on human gut (10), are both encoded by a β-lactamase-producing plasmid (8). The fifth nonfimbrial adhesin mediates an aggregative adhesion pattern in intestinal cell lines (13). The recent analysis of the KP NTUH-K2044 genome revealed, in addition to type 1 and type 3 fimbriae encoding operons, seven novel fimbriae kpa, kpb, kpc, kpd, kpe, kpf, and kpg (94IDP11). This is consistent with many findings that multiple fimbriae are required for a bacteria to establish an infection (59).
Expression of type 1 fimbriae
Type 1 fimbriae, which are characterized by their binding to mannosides, are expressed in most members of the family Enterobacteriaceae (12, 21, 25). Encoded by the
fimAICDFGH gene cluster, type 1 fimbriae is composed of the major subunit FimA and the
adhesin FimH. The expression of type 1 fimbriae is phase variable which is mediated by an invertible 414-bp cis element, fimS, that is located upstream of fimA. Flipping of fimS alternates E. coli between type 1 fimbriated and nonfimbriated states, namely ON and OFF phase. The site-specific recombination process requires either FimB (ON-to-OFF or OFF-to-ON switch) or FimE (ON-to-OFF switch only) (30). KP carries an extra gene fimK downstream to the fim operon. A negative role of FimK for the expression of type 1 fimbriae has been suggested to explain why KP is a less prevalent etiological agent of UTI (44).
The recent DNA microarraydemonstrated a coordinate expression betweentype 1 and P fimbriae in an inverse manner (20). This cross-regulationoccurred when PapB, a positive regulator for type P fimbriae expression, was expressed to inhibit the FimB-promoted
recombination, but enhance the expression of fimE (63). The deletion of ompA in E. coli K1 was shown to decrease the expression of type 1 fimbriae (40). Moreover, an iron-dependent regulation of FimE via IscR was reported to decrease expression of type 1 fimbriae (62). Most recently, Rcs phosphorelay has also been reported to affect the expression of type 1 fimbriae via positively regulating the FimB expression (47).
Expression of type 3 fimbriae
Type 3 fimbriae, which is encoded by mrk gene cluster, have been demonstrated to be responsible for the adherence of KP to uroepithelial cells, respiratory epithelia, intestinal and endothelial cells of (22). The mrk operon includes mrkA, encoding the major fimbrial subunit; mrkB and mrkC, respectively coding for chaperone and usher proteins for the assembly of the fimbriae; mrkD coding for the adhesin with a MR/K hemagglutination activity (1, 16); and mrkF, encoding a protein to affect the length and adhesion activity of the fimbriae (24). The bacterial growth on abiotic surfaces is facilitated, in part, by the major fimbrial subunit MrkA protein, whereas growth on surfaces coated with a human extracellular matrix (HECM) requires the presence of the MrkD adhesin (16, 55). We have shown in 94IDP11 that the type 3 fimbrial expression could be readily demonstrated in the clinical isolates. The expression of type 1 fimbriae, however, was only evident in the mrkA deletion mutant or in KP carrying a fimB-overexpression plasmid. This suggested the presence of a cross-talk regulation on the expression of the two fimbriae.
Cross-talk regulation on the expression of fimbriae and CPS
Down regulation of cps operon following the attachment of type 1 fimbriae to D-mannose receptor was observed in E. coli (46), which implying a counter-expression of the two surface components. However, the global regulator RcsB for the expression of CPS (60) was recently shown to affect positively the expression of type 1 fimbriae (47). Clinically isolated KP usually produces large amounts of capsular polysaccharide (CPS) as a major virulence factor to protect the bacteria from phagocytosis and from killing by serum factors (28, 42). Several reports in KP indicated that inhibition of capsule synthesis enhances type 1 fimbrial function (14, 37, 45, 48, 49). The deletion of rcsB reduced the expression of CPS and the expression of type 3 fimbriae, but increased the activity of type 1 fimbriae, indicating an interacting regulation plays a role for the differential expression of the two surface components.
C-di-GMP is involved in regulation of the expression of fimbriae
Cyclic-di-GMP, the most recently identified second messenger in bacteria, is synthesized from GTP by diguanylate cyclases (DGCs) and broken down by specific phosphodiesterases (PDEs) (43, 50). DGC activity is associated with the GGDEF domain protein, while c-di-GMP-specific PDE activity is associated with the EAL or HD-GYP domain proteins. The PilZ-domain proteins are one of the currently known c-di-GMP effector (19). In general, c-di-GMP stimulates the biosynthesis of adhesins and
FimK and KP4554 are both EAL-domain containing proteins, while KP4551 encodes the only c-di-GMP sensor (PilZ domain protein) in the genome. It would be of interest to investigate whether they play a role in controlling the level of c-di-GMP leading to the counter expression of type 1 and type 3 fimbriare.
Envelope stress responses
The regulatory systems involved in E. coli envelope stress responses include the CpxAR (Cpx) and BaeSR (Bae) two-component systems, the σE alternative sigma factor, the phage shock protein (Psp), and theRcs phosphorelay (17, 38). In Salmonella enterica, loss of mediated regulation might cause unfolded OMPs to accumulate in the periplasm leading to the triggering of the sigma E response (2, 3). The mutation of cpxA was able to increase ompC transcription but strongly decrease the transcription of ompF (2). In the absence of the Cpx pathway, the pap gene cluster synthesize aberrant P pili and DegP is induced to degrade the toxic, off-pathway aggregated subunits (26). Cpx envelope stress response also affects expression of bundle-forming pili of enteropathogenic E coli (38). On the other hand, adesion of type 1 fimbriae to abiotic surface leads to altered composition of the outer membrane proteins in E. coli (40). Conceivably, the regulatory systems involved in release of the envelope stress must play a modulation role for the expression, assembly and transportation of the envelope components, fimbriae and CPS. Our preliminary data indicated an envelope change affected differentially on the expression of type 1 and type 3 fimbriae. It is hence if the envelope stress responses exert a differential regulation, how or why the differentiation, would be of interest to be studied.
In general, two KP strains NTUH-K2044 of K1 serotype and CG43 of K2 serotype will be used in the following studies and their properties comparatively analyzed.
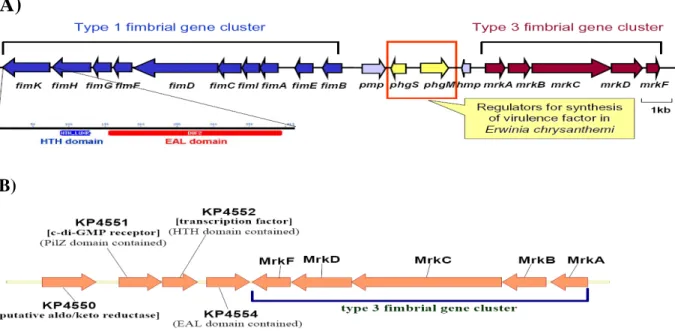
AIM 1. To investigate if the deletion the specific regulatory genes affects the expression: The genes include fimK, pecgS, pecM, KP4551(mrkJ), KP4552(mrkI), and KP4554 (mrkH) those are clustered with the operons encoding type 1 and type 3 fimbriae (Fig. 1). Sequence analysis revealed that (1) FimK contains in the N-terminal domain with an HTH motif and C-terminal EAL domain possibly providing a c-di-GMP phosphodiesterase (PDE) activity. (2) PecS and PecM show homology respectively with the virulence regulators PecS and PecM of Erwinia chrysanthemum. (3) MrkH, MrkI, and MrkJ respectively encode a
c-di-GMP sensor(PilZ protein), transcription factor with an HTH domain, and EAL protein. In addition to analyzing the deleting effects of the above mentioned regulatory genes on the expression of either fimbriae, the possibility of regulatory role of c-di-GMP in coordinate expression of the fimbriae and how are these proteins exert their regulatory function will also be investigated.
AIM 2. To generate transposon-insertion mutant libraries to screen for the regulatory genes: Transposon mutagenesis will be performed on CG43S3rcsB- strain, the mutants screened for decreasing activity of yeast agglutination, an assessment assay for type 1 fimbriae activity. Promoter reporter approach will also be used for the genome-wide screening of the transposon-insertion mutants on X-gal plate.
AIM 3. To investigate if envelope stress plays a role in the regulation of the
counter-expression: Deletion effect of OmpA, OmpC, OmpF and OmpN will be analyzed. Any of the genes showing apparent changes of the expression level will be selected for further study. The expression of CpxAR, σE and RcsB, which are activated in response to disruptions of the folding of envelope proteins and the integrity of the outer membrane, will be determined by quantitative PCR.
III. Material and Methods
1. Generation of type 1 fimbriae “lock ON” mutant and the mutants carrying either deletion of the involved regulators. Preliminary data indicated that the promoter of the type 1 fimbriae in either NTUH-K2044 or CG43 was generally in OFF phase. The fimK deletion in either strain had no apparent effect on either activity of type 1 or type 3 fimbriae. This is likely due to that fimK is resided in the fim operon structure and hence not expressed with an “OFF” phase promoter. RT-PCR will be firstly carried out the demonstrate the operon structure, and then fim “LOCK-ON” mutant derived from CG43 or NTUH K-2044 constructed before introducing with the secondary mutation of fimK deletion.
- Deletion mutant constructs of the genes involved in the regulation: The DNA fragments of 1 kb in length flanking both ends of the target gene will be amplified and the PCR fragments cloned into the suicide vector pKAS46 (51) or pCVD442. The plasmids will then be respectively transferred into E. coli S17-1λ-pir and then mobilized to K. pneumoniae NTUH K-2044 or CG43S3 (31) by conjugation. Occurrence of double recombination will be verified by PCR and confirmed with Southern blotting hybridization.
2.Assements of the fimbrial expression - To measure the fimbrial activity:
(1) Hemagglutination assay- Basically as described (16), the overnight grown bacteria suspended in PBS to approximately 109 CFU/ml and Human or animal erythrocytes in PBS will be collected. A series of four-fold dilution of the bacterial suspension will then be mixed with an equal volume of 3% (vol/vol) erythrocytes and the mixture incubated at room temperature for 30 min to allow erythrocytes settle to the bottom of the glass tube. (2) Yeast agglutination assay- Agglutination of yeast Saccharomyces cerevisiae AH109 will be
carried out as described (12). Briefly, the bacteria (2 x 109/ml) suspended in PBS or PBS with 2% mannose will be mixed with 10 mg/ml of yeast on a glass slide. The agglutination count is expressed as the highest dilution of the bacteria causing visible agglutination of yeast.
(3) ECM (extracellular metrix) binding assay- The wells of flatbottom microtiter plate (Nunc-ImmunoTM plate) will be coated with optimal concentrations of type IV collagen, type V collagen or fibronectin. Subsequently, each well will then be added with 100 μl bacteria (108cfu/ml) and the incubation continued for 2 h at 22°C with gentle shaking. After washing with 0.05% Tween-20 to remove the unattached bacteria, the attached bacteria will be washed off by 0.1% Triton X-100 and the adhesion determined by the recovery of the bacteria.
(4) Cell adherence assay- The tested cells will be seeded onto 24 well plate (TPP industries, France) and incubated to confluent growth in 5% CO2 for 48 h. Approximately 1x107 bacteria will then be added to each well containing about 105 cells, and the incubation continued for 1 h. After washing with phosphate-buffered saline (PBS), the cells will be lysed with 0.1% Triton X-100 and the cell-adhesive bacteria measured by recovery of the bacteria from the lysates. The cell lines include intestinal cells of Int-407 and HCT-8,
urinary bladder cancer cellsT24, hepatoma cells Hep-G2, colon adeno-carcinomaCaco-2. (5) Biofilm formation analysis- The bacteria diluted 1/100 in LB will be inoculated into each well of a 96-well microtiter dish and incubated at 37°C for 36 h. After the removal of the bacteria, 150 μl of 1% (w/v) crystal violet will be added to each well and the plate incubated for 30 min at room temperature. The plate will then be washed, the dye solubilized in 1% (w/v) SDS, and the absorbance at 595 nm determined as described (39). - To measure at the molecular level:
(1) Real time PCR or RT-PCR analysis: Total RNA will be isolated from K. pneumoniae cells by extraction with the TRI reagent (MolecularResearch Center, Cincinnati, Oh, USA) and the residual DNA eliminated with RQ1 RNase-free DNase (Promega, Madison,WI, USA). The cDNAs used for PCR amplification will be synthesized from 1 μg of total RNA using the random hexamer primer from a RT-PCR kit.
(2) Promoter activity measurement: The putative promoters will be PCR amplified using the specific primers and the PCR products subcloned in front of the promoterless lacZ gene on placZ15 (35). The bacteria carrying each of the reporter plasmids will be grown in suitable conditions, and the β-galactosidase activities measured.
(3) Western blotting hybridization: The assayed fimbrial proteins will be resolved by SDS-PAGE and then electrophoretically transferred onto a PVDF membrane (Millipore, Billerica, MA, USA). Polyclonal antibody will be prepared by immunizing five-week old female BALB/c mice and the antisera obtained by intra-cardiac puncture.
3. Transposon-mediated random mutagenesis
Transposon-mediated mutagenesis will be performed using a mini-Tn5 Km2 transposon (11). Briefly, the plasmids harboring either of the fimbrial promoter-lacZ transcriptional fusion will be mobilized into K. pneumoniae NTUH K2044lacZ- or CG43lacZ16 by conjugation. The mini-Tn5 containing plasmid will then be introduced by conjugation and the resulting transconjugants screened on LB agar plates supplemented with X-gal. Colonies displaying apparent alteration of color indicating increase or reduction of LacZ activity will be isolated. The transposon-inserted sequence will be determined using the specific primer.
IV. Results and Discussion
Specific aim 1- To investigate if the deletion the specific regulatory genes affects the expression:
As shown in Fig. 1A and B, the gene organizations which have been recently reported to be conserved in the genome of K. pneumoniae NTUH-K2044, MGH78578, and 342 (65) were also found in the contig sequences of K. pneumoniae CG43 (unpublished results from Dr. S.-F. Tsai, NHRI) determined using BLAST analysis. The cluster gene organization suggests a coordinate regulation on the expression of type 1 and 3 fimbriae.
(A)
(B)
Fig. 1. (A) Gene organization of the type 1 and 3 fimbriae and (B) the downstream genes of type 3 fimbrial operon in K. pneumoniae CG43.
Deletion of phgS (pecS) or phgM (pecM) increased the expression of MrkA
Dilution RT-PCR shown in Fig. 2(A) indicated that the deletion of either pecS (phgS) or pecM (phgM) increased the transcript of MrkA. The western blot further supported a negative regulatory role of PecSM on the expression of MrkA (Fig. 2B). On the other hand, neither deletion had apparent effect on the expression of type 1 fimbriae.
(A)
(B)
Fig. 2. (A) Limiting-dilution RT-PCR analysis of mrkA. (B) Western blotting analysis of the expression of type 3 fimbriae. M: marker. Lane 1: K. pneumoniae CG43S3; 2: K. pneumoniae CG43S3phgS-; 3: K. pneumoniae CG43S3phgM-.
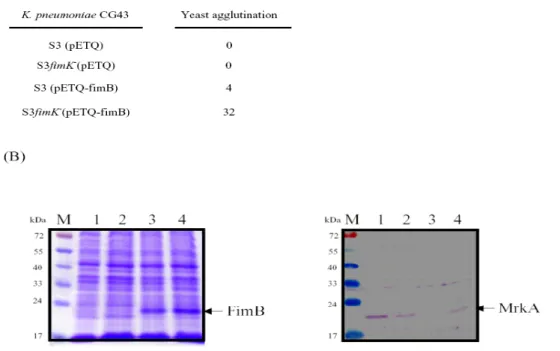
Deletion of fimK increased the expression of both fimbriae
As shown in Fig. 3A, deletion of the fimK increased the yeast agglutination activity. The gene deletion from the fimB overexpression bacteria also enhanced the expression of MrkA (Fig. 3B). These suggested a negative regulatory of FimK on the expression of the two fimbriae.
Fig. 3. Deletion effect of fimK on the expression of type 1 fimbriae (A) and type 3 fimbriae (B). Lanes 1, CG43S3(pETQ); 2, CG43S3fimK-(pETQ); 3, CG43S3(pETQ-fimB); 4, S3fimK
FimK exhibited a c-di-GMP phosphodiesterase (PDE) activity and c-di-GMP plays a positive role on the expression of type 3 fimbriae.
As shown in Fig. 4A, expression of YhjH, a c-di-GMP
PDE in E. coli W3110 apparently increased the bacterial swimming activity. Introducing in the bacteria with the FimK expression plasmid, pRK415-FimK, but not the one with single residue substitution (FimKE246A) also enlarged the swimming zone. This suggested that FimK is a c-di-GMP PDE and the residue E246 is essential for the activity. As shown in Fig. 4B,overexpression of the PDE domain of FimK abolished the expression of MrkA. On the other hand, overexpression of the GGDEF domain protein YdeH appeared to enhanced the expression of MrkA. In summary, FimK negatively regulates the expression of type 1 and type 3 fimbriae while
cyclic-di-GMP plays a positive role on type 3 fimbriae expression.
(A)
(B)
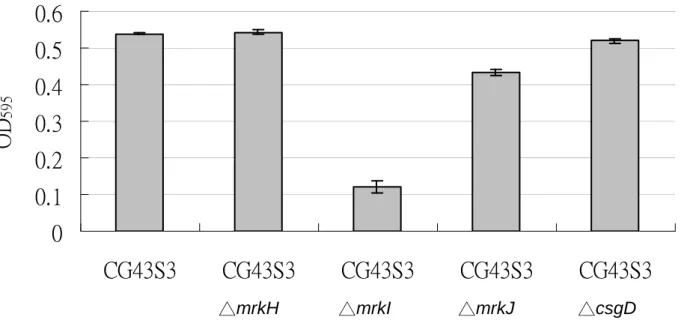
Deletion of mrkI reduced the expression MrkA but increased the expression of type 1 fimbriae As shown in Fig. 5A and 5B, deletion of KP4552 (mrkI) abolished the MrkA expression, while an increased activity of mannose sensitive yeast agglutination (MSYA) was noted. The deletion of mrkI appeared to decreased the biofilm formation implying a decreased expression of type 3 fimbriae. These revealed that KP4552 is probably the regulator for the counter-expression of the two fimbriae.
(A) (B) 1X PBS+ 1M mannose mrkH mrkJ csgD mrkI Wt 1X PBS Wt
mrkH
mrkI
mrkJ
csgD MrkA
(C)
Fig. 5. (A) Western blotting analysis for the expression of type 3 fimbriae using anti-MrkA antibody. M: pre-stained marker. Lanes 1: K. pneumoniae CG43S3; 2: K. pneumoniae CG43S3△ 4551; 3: K. pneumoniae CG43S3△4552; 4: K. pneumoniae CG43S3△4554; 5: K. pneumoniae CG43S3△csgD; 6: positive control for anti-MrkA. (B) Yeast agglutination activity assay for the expression of type 1 fimbriae (C) Biofilm formation activity.
Specific aim 2- To generate transposon-insertion mutant libraries to screen for the regulatory genes:
Deletion of RcsB decreased the MrkA expression but increased the activity of type 1 fimbriae The rcsB deletion, which decreased the production of the capsular polysaccharide (Fig. 6A), led to a slight reduction in MrkA expression (Fig. 6B) but an increased in type 1 fimbriae activity (Fig. 6C). This suggested a possible role of RcsB for the counter expression of the two fimbriae.
0
0.1
0.2
0.3
0.4
0.5
0.6
CG43S3
CG43S3
DKP4551
CG43S3
DKP4552
CG43S3
DKP4554
CG43S3
DcsgD
OD
59 5 △csgD △mrkJ △mrkI △mrkHFig. 6. Deletion effect of the cps regulators on the expression of cps (A), type 3 (B) and type 1 fimbriae (C). The cps regulators include RmpA (Regulator for mucoid phenotype) RmpA2, and RcsB.
RcsB negatively affected the expression of type 1 fimbriae
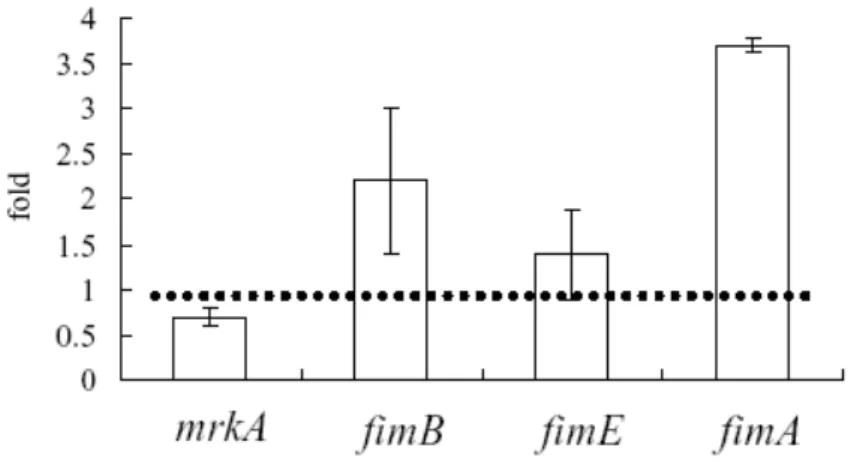
As shown in Fig. 7, Q-PCR analysis suggested RcsB negatively regulates the expression of fimB leading to a reduced activity of type 1 fimbriae. However, analysis of PmrkA revealed no consensus RcsB box which could be found within PfimB and PfimE. It implies an indirect control by RcsB on the expression of type 3 fimbriae. Transposon mutagenesis was hence carried out as shown in Fig. 7 to isolate the mutant displaying phenotype with a suppression of the rcsB deletion effect. Since all the isolated mutants appeared to carry defects in LacZ expression, more mutagenesis experiments have to be performed.
Fig. 6. Q-PCR analysis of the expression of mrkA, fimB, fimE, and fimA in CG43S3 and CG43S3rcsB-.
Fig.7. LacZ activity of K. pneumoniae CG43S3Z01rcsB-derived transposon-insertion mutants.
Specific aim 3- To investigate if envelope stress plays a role in the regulation of the counter-expression:
Deletion of OmpA or OmpC (OmpK36) affected the expression of MrkA and the biofilm formation
As shown in Fig. 8A, either ompA or ompK36 (ompC) deletion mutant exhibited reduced expression of MrkA under static growth suggesting a micraerobic control for the expression of type 3 fimbriae. Nevertheless, the ompC deletion had less effect on the biofilm formation compared to the deletion of
ompA. On the other hand, neither deletion had effect on the activity of MSYA.
(A)
(B) Fig.8. Analysis of the deletion effects of the omp genes on the expression of type 3 fimbriae (A) and biofilm formation (B).
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45
S3 △ompA △ompF △ompN △ompC
OD
V. References Cited
1. Allen, B.L., G. –F. Gerlach, and S. Clegg. 1991. Nucleotide sequence and functions of mrk determinants necessary for expression of type 3 fimbriae in Klebsiella pneumoniae. J. Bacteriol. 173:916-920.
2. Batchelor E, Walthers D, Kenney LJ, Goulian M. 2005. The Escherichia coli CpxA-CpxR envelope stress response system regulates expression of the porin proteins OmpF and OmpC. J Bacteriol 187:5723-5731.
3. Bossi L, Maloriol D, Fiqueroa-Bossi N. 2008. Porin biogenesis activates the sigma(E) response in Salmonella hfq mutants. Biochimie. 90: 1539-1544.
4. Brisse S, Fevre, Passet V, Issenhuth-Jeanjean S, Tournebize R, Diancourt L, Grimont P. 2009. Virulent clones of Klebsiella pneumoniae : Identification and evolutionary scenario based on genomic and phenotypic characterization. PLoS ONE 4 (3)
5. Chang FY, Chou MY. 1995. Comparison of pyogenic liver abscesses caused by Klebsiella
pneumoniae and non-Klebsiella pneumoniae pathogens. J. Formos. Med. Assoc. 94:232-237.
6. Chen YT, Chang HY, Lai YC, Pan CC, Tsai SF, Peng HL. 2004. Sequencing and analysis of the large virulence plasmid pLVPK of Klebsiella pneumoniae CG43. Gene 337: 189-198.
7. Chuang YP, Fang CT, Lai SY, Chang SC, Wang JT. 2006. Genetic determinants of capsular serotype K1 of Klebsiella pneumoniae causing primary pyogenic liver abscess. J Infect Dis 193:645-54.
8. Darfeuille-Michaud A, Jallat C, Aubel D, Sirot D, Rich C, Sirot J, and Joly B. 1992. R-plasmid-encoded adhesive factor in Klebsiella pneumoniae strains responsible for human nosocomial infections. Infect. Immun. 60:44-45.
9. Di Martino P, Bertin Y, Girardeau JP, Livrelli V, Joly B, Darfeuille-Michaud A. 1995. Molecular characterization and adhesive properties of CF29K, an adhesin of Klebsiella
pneumoniae strains involve in nosocomial infections. Infect. Immun. 63:4336-4344.
10.Di Martino P, Livrelli V, Sirot D, Joly B, and Darfeuille-Michaud A. 1996. A new fimbrial antigen harbored by CAZ-5/SHV-4-producing Klebsiella pneumoniae strains involved in nosocomial infections. Infect. Immun. 64:2266-2273.
11.de Lorenzo V, Timmis KN. 1994. Analysis and construction of stable phenotypes in gram-negative bacteria with Tn5- and Tn10-derived minitransposons. Methods Enzymol 235, 386-405.
12.Fader RC, Gondesen K, Tolley B, Ritchie DG, Moller P. 1988. Evidence that in vitro adherence of Klebsiella pneumoniae to ciliated hamster tracheal cells is mediated by type 1 fimbriae. Infect. Immun. 56:3011-3013.
13.Favre-Bonte S, Darfeuille-Michaud A, Forestier C. 1995. Aggregative adherence of Klebsiella
pneumoniae to human intestine-407 cells. Infect. Immun. 63:1318-1328.
14.Favre-Bonte S, Joly B, Forestier C. 1999. Consequences of reduction of Klebsiella
pneumoniae capsule expression on interactions of this bacterium with epithelial cells. Infect.
15.Fung CP, Chang FY, Lee SC, Hu BS, Kuo BI, Liu CY, Ho M, Siu LK. 2002. A global emerging disease of Klebsiella pneumoniae liver abscess: is serotype K1 an important factor for complicated endophthalimitis? GUT 50:420-424.
16.Gerlach GB, Allen L, Clegg S. 1989. Type 3 fimbriae among Enterobacteriaceae and the ability of spermidine to inhibit MR/K hemagglutination. Infect. Immun. 57:219-224.
17.Guisbert E, Rhodius VA, Ahuja N, Witkin K, Gross CA. 2007. Hfq modulates the σE-mediated envelope stress response and σ32-mediated cytoplasmic stress response in Escherichia coli. J Bacteriol 189:1963-73.
18.Han SHB. 1995. Review of hepatic abscess from Klebsiella pneumoniae. An association with diabetes mellitus and septic endophthalmitis. West. J. Med.162:220-224.
19.Hengge R. 2009. Principles of c-di-GMP signaling in bacteria. Nature Microbiology Rev 7:263-
20.Holden, N.J., B.E. Uhlin, and D.L. Gally. 2001. PapB paralogues and their effect on the phase variation of type 1 fimbriae in Escherichia coli. Mol Microbiol 42: 319–330.
21.Hornick DB, Allen BL, Horn MA, Clegg S. 1991. Fimbrial types among respiratory isolates belonging to the family of Enterobacteriaceae. J Clin Microbiol 29:1795-1800.
22.Hornick DB, Allen BL, Horn MA, Clegg S. 1992. Adherence to respiratory epithelia by recombinant Escherichia coli expressing Klebsiella pneumoniae type 3 fimbrial gene products. Infect. Immun. 60:1577-1588.
23.Huang YH, Ferrieres L, Clark DJ. 2006. The role of Rcs phosphorelay in Enterobacteriaceae. Res Microbiol 157: 206-12.
24.Huang YJ, Liao HW, Wu CC, Peng HL (2009) MrkF is a component of type 3 fimbriae in
Klebsiella pneumponiae. Res Microbiol 160:71-9.
25.Jones CH, Pinkner JS, Roth R, Heuser J, Nicholes AV, Abraham SN, Hultgren SJ. 1995. FimH adhesin of type 1 pili is assembled into a fibrillar tip structure in the Enterobacteriaceae. Proc Natl Acad Sci USA 92: 2081–2085.
26.Jones CH, Dexter P, Evans AK, Liu C, Hultgren SJ, Hruby DE. 2002. Escherichia coli DegP
protease cleaves between paired hydrophobic residues in a natural substrate: the PapA pilin. J Bacteriol 184:5762-5771.
27.Jong GM, Hsiue TR, Chen CR, Chang HY, Chen CW. 1995. Rapidly fatal outcome of bacteremic Klebsiella pneumoniae pneumonia in alcoholics. Chest 107:214-217.
28.Kabha K, Nissimov L, Athamna A, Keisari Y, Parolis H, Parolis LAS, Grue RM, Schlepper-Shafer J, Ezekowitz ARB, Ohman DE, Ofek I. 1995. Relationships among capsular structure, phagocytosis, and mouse virulence in Klebsiella pneumoniae. Infect. Immun. 63:847–852.
29.Keynan Y, Karlowsky JA, Walus T, Rubinstein E. 2007. Pyogenic liver abscess caused by hypermucoviscous Klebsiella pneumoniae. Scan J Infect Dis 39:828-830.
30.Klemm P. 1986. Two regulatory fim genes, fimB and fimE, control the phase variation of type 1 fimbriae in Escherichia coli. EMBO J. 5:1389-1393.
31.Lai YC, Peng HL, Chang HY. 2003. RmpA2, an activator of capsule biosynthesis in
Klebsiella pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level. J
Bacteriol 185:788-800.
32.Lau YJ., Hu BS, Wu WL, Lin YH, Chang HY, Shi ZY. 2000. Identification of a major cluster of Klebsiella pneumoniae isolates from patients with liver abscess in Taiwan. J. Clin. Microbiol. 38:412-4.
33.Laubacher ME, Ades SE. 2008. The Rcs phosphorelay is a cell envelope stress response activated by peptidoglycan stress and contributes to intrinsic antibiotic resistance. J Bacteriol. 191: 2065-2074.
34.Lee HC, Chuang YC, Yu WL, Lee NY, Chang CM, Ko NY, Wang LR. Ko WC. 2006. Clinical implication of hypermucoviscosity phenotype in Klebsiella pneumoniae isolates: association with invasive syndrome in patients with community-acquired bacteremia. J Internal Med 259:606-614.
35.Lin CT, Huang TY, Liang WC, Peng HL. 2006. Homologous response regulators KvgA, KvhA and KvhR regulate the synthesis of capsular polysaccharide in Klebsiella pneumoniae CG43 in a coordinated manner. J. Biochem 140:429-438.
36.Lin JC, Chang FY, Fung CP, Xu JZ, Cheng HP, Wang JJ, Huang LY, Siu LK. 2004. High prevalence of phagocytic-resistant capsular serotypes of Klebsiella pneumoniae in liver abscess. Micorbes Infect 6:1191-8.
37.Matatov R, Goldhar J, Skutelsky E, Sechter I, Perry R, Podschun R, Sahly H, Thankavel K, Abraham SN, Ofek I. 1999. Inability of encapsulated Klebsiella pneumoniae to assemble functional type 1 fimbriae on their surface. FEMS Microbiol Lett. 179:123-30.
38.Nevesinjac AZ, Raivio TL. 2005. Cpx envelope stress response affects expression of bundle-forming pili of enteropathogenic Escherichia coli. J Bacteriol 187:672-686.
39.O'Toole G, Kaplan HB, Kolter R. 2000. Biofilm formation as microbial development. Annu Rev Microbiol.; 54:49-79.
40.Otto K, Norbeck J, Larsson T, Karlosson K-A, Hermansson M. 2001. Adesion of type 1-fimbriated Escherichia coli to abiotic surface leads to altered composition of outer membrane proteins. J Bacteriol 183:2445-2453.
41.Podschun R, Ullmann U. 1998. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 11:589-603.
42.Regueiro V, Campos MA, Pons J, Alberti S, Bengoechea JA. 2006. The uptake of a Klebsiella
pneumoniae capsule polysaccharide mutant triggers an inflammatory response by human airway
epithelial cells. Microbiology 152:555-66.
43.Romling U., and D. Amikam. 2006. Cyclic di-GMP as a second messenger. Curr Opin Microbiol. 9:218-228.
44.Rosen DA, Pinkner JS, Jones JM, Walker JN, Clegg S, Hultgren SC. 2008. Utilization of an intracellular bacterial community pathway in Klebsiella pneumoniae urinary tract infection and the effects of FimK on type 1 pilus expression. Infect Immun 76:3337-3345.
45.Sahly H, Podschun R, Oelschlaeger TA, Greiwe M, Parolis H, Hasty D, Kekow J, Ullmann U, Ofek I, Sela S. 2000. Capsule impedes adhesion to and invasion of epithelial cells by Klebsiella
pneumoniae. Infect Immun. 68:6744-9.
46.Schwan WR, Beck MT, Hultgren SJ, Pinkner J, Woolever NJ, Larson T. 2005. Down regulation of the kps region 1 capsular assembly operon following attachment of Escherichia coli type 1 fimbriae. Infect Immun 73:1226-1231.
47.Schwan WR, Shibata S, Aizawa SI, Wolfe AJ. 2007. The two component response regulator RcsB regulate type 1 piliation in Escherichia coli. J Bacteriol 189:7159-63.
48.Schembri MA, Dalsgaard D, Klemm P. 2004. Capsule shields the function of short bacterial adhesins. J Bacteriol. 186:1249-57.
49.Schembri MA, Blom J, Krogfelt KA, Klemm P. 2005. Capsule and fimbria interaction in
Klebsiella pneumoniae. Infect Immun 73:4626-4633.
50.Schmidt AJ, Ryjenkov DA, Gomelsky M. 2005. The ubiquitous protein domain EAL is a cyclic diguanylate-specific phosphodiesterase: enzymatically active and inactive EAL domains. J Bacteriol 187: 4774-4781.
51.Skorupski K, Taylor RK. 1996. Positive selection vectors for allelic exchange. Gene 169:47-52.
52.Snyder JA, Haugen BJ, Lockatell CV, Maroncle N, Hagan EC, Johnson DE, Welch RA, Mobley HLT. 2005. Coordinate expression of fimbriae in uropathogenic Escherichia coli. Infect Immun 73:7588-96.
53.Stahlhut SG, Chattopadhyay S, Struve C, Weissman SJ, Aprikian P, Libby SJ, Fang FC, Angeliki Krogfelt K, Sokurenko EV. 2009. Population variability of the FimH type 1 fimbrial adhesion in Klebsiella pneumoniae. J Bacteriol 191:1941-1950.
54.Struve C, Bojer, Angeliki Krogfelt K. 2008. Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect Immun 76:4055-65.
55.Tarkkanen AM, Allen BL, Westerlund B, Holthofer H, Kuusela P, Risteli L, Clegg S, Korhonen TK. 1990. Type V collagen as the target for type-3 fimbriae, enterobacterial adherence organelles. Mol. Microbiol. 4:1353-1361.
56.Tarkkanen AM, R Virkola R, Clegg S, Korhonen TK. 1997. Binding of the type 3 fimbriae of
Klebsiella pneumoniae to human endothelial and urinary bladder cells. Infect. Immun.
65:1546-1549.
57.Teng C-H, Xie Y, Shin S, Di Cello F, Paul-Satyaseela M, Cai M, Kim KS. 2006. Effects of
ompA deletion on expression of type 1 fimbriae in Escherichia coli K1 strain RS218 and on the
association of E. coli with human brain microvascular endothelial cells. Infect Immun 74:5609-16.
58.Tsai F-C, Huang Y-T, Chang L-Y, Wang J-T. 2008. Pyogenic liver abscess as endemic disease, Taiwan. Emerging Infectious Diseases 14:1592-
59.van der Velden AW, Baumler AJ, Tsolis RM, Heffron F. 1998. Multiple fimbrial adhesins are required for full virulence of Salmonella typhimurium in mice. Infect Immun. 66:2803-8.
60.Wacharotayankun R, Arakawa Y, Ohta M, Hasegawa T, Mori M, Horri T, Kato N. 1992. Involvement of rcsB in Klebsiella K2 capsule synthesis in Escherichia coli K-12. J Bacterial 174:1063-7.
61.Wang JH, Liu YC, Lee SS, Yen MY, Chen YS, Wang JH, Wann SR, Lin HH. 1998. Primary liver abscess due to Klebsiella pneumoniae in Taiwan. Clin. Infect. Dis. 26:1434-1438.
62.Wu Y, Outten FY. 2009. IscR controls iron-dependent biofilm formation in Escherichia coli by regulation type 1 fimbria expression. J Bacteriol. 191:1248-1257.
63.Xia Y, Gally D, Forsman-Semb K, Uhlin BE. 2000. Regulatory cross-talk between adhesin operons in Escherichia coli: inhibition of type 1 fimbriae expression by the Pap B protein. EMBO. 19:1450-1457.
64.Yu WL, Ko WC, Cheng KC, Lee HC, Ke DS, Lee CC, Fung CP, Chuang YC. 2006. Association between rmpA and magA and clinical syndromes caused by Klebsiella pneumoniae in Taiwan. Clin Infect Dis 42:1351-8.
65.Wu, C.C., Huang, Y.J., Fung, C.P., Peng, H.L., 2010. Regulation of the Klebsiella
pneumoniae Kpc fimbriae by the site-specific recombinase KpcI. Microbiology
國科會補助專題研究計畫成果報告自評表
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)
、是否適
合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
▓達成目標
□
未達成目標(請說明,以 100 字為限)
□
實驗失敗
□
因故實驗中斷
□
其他原因
說明:
2. 研究成果在學術期刊發表或申請專利等情形:
論文:□已發表 □未發表之文稿 ▓撰寫中 □無
專利:□已獲得 □申請中 □無
技轉:□已技轉 □洽談中 □無
其他:(以 100 字為限)
3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以 500
字為限)
We have carried out the experiments in a year as planned, three manuscripts (1~3) are in the final stage to be submitted for publication. Many interesting findings including (a) FimK negatively regulated the expression of type 1 and type 3 fimbriae, (b) the second messenger c-di-GMP plays a positive role on the expression of both fimbriae, (c) PecSM affected negatively on the expression of type 3 fimbriae, however, had no apparent effect on the expression of type 1 fimbriae, (d) MrkI is likely the regulator for the counter- expression of the two types of fimbriae, (e) q-PCR analysis suggested RcsB negatively regulates the expression of fimB leading to a reduced activity of type 1 fimbriae, and (f) deletion of ompA or ompC reduced the MrkA expression and biofilm formation, allow unravel a cross-talk regulatory circuit on the expression of type 1 and type 3 fimbriae, which may provide intervening targets for antimicrobial agents.
1. Huang YJ, Wu CC, Cheng HY, and Peng HL. (2010) Regulation on the expression of type 1 and type 3 fimbriae in Klebsiella pneumoniae CG43.
2. Wu CC, Lin CT, Cheng WY, and HL Peng (2010) Identification of MrkI-mediated counter expression of type 1 and type 3 fimbriae in Klebsiella pneumoniae CG43.
3. Wu CC, Lin CT, and HL Peng (2010) Cyclic-di-GMP affects the expression of type 3 fimbriae via Fur-dependentregulation in Klebsiella pneumoniae CG43.
國科會補助專題研究計畫項下出席國際學術會議心得報告
日期:99 年月 日一、參加會議經過:
這個兩年一次的專題會議約有一百五十與會人數。五天的會議,有兩個 主要議題(一)細菌鞭毛運轉和趨化性機制;(二)細菌和酵母菌的訊 息傳遞。會議於早上九點開始至晚上九點半左右結束,平均每天有十二 個講題,分別有二十鐘、十分鐘和五分鐘的演講;下午四點至六點為壁 報討論時段,是個議程緊湊但內容極為豐富的學術研究會議。二、與會心得:
如附件三、考察參觀活動(無是項活動者略):
無四、建議:
鼓勵推動專題性的小型會議五、攜回資料名稱及內容:
此會議鼓勵最新發現及尚未發表的研究結果,所以,除了會議 流程外,並沒有摘要集也嚴禁照相。計畫編號
NSC98-3112-B-009-005-
計畫名稱 克雷白氏肺炎桿菌第一型纖毛和第三型纖毛交互表現的調控
出國人員
姓名
彭慧玲
服務機構
及職稱
交通大學生物科技學系
會議時間
99 年 1 月 24 日至
99 年 1 月 29 日
會議地點 美國洛杉磯- Ventura
會議名稱
(中文)微生物的訊息傳導
(英文)
Gordon Conference on “Sensory Transduction in Microorganisms”發表論文
題目
(中文)克雷白氏肺炎桿菌
RcsB 傳導的抗酸反應
(英文)
Characterization of the RcsB-mediated acid response in Klebsiella行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
Regulation for the counter-expression of type 1 and type 3
fimbriae in Klebsiella pneumoniae
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 98-3112-B-009-005-
執行期間: 九十八 年 八 月 一 日 至 九十九 年 七 月 三十一 日
計畫主持人:彭慧玲
共同主持人:
計畫參與人員:黃盈蓉,吳健誠,王哲充,鄭崴云
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
▓赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
■出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,■一年□二年後可公開查詢
執行單位: 交通大學生物科技學系
中 華 民 國 九十九 年 七 月 三十 日
國科會補助專題研究計畫項下出席國際學術會議心得報告
日期:99 年月 日一、參加會議經過:
這個兩年一次的專題會議約有一百五十與會人數。五天的會議,有兩個 主要議題(一)細菌鞭毛運轉和趨化性機制;(二)細菌和酵母菌的訊 息傳遞。會議於早上九點開始至晚上九點半左右結束,平均每天有十二 個講題,分別有二十鐘、十分鐘和五分鐘的演講;下午四點至六點為壁 報討論時段,是個議程緊湊但內容極為豐富的學術研究會議。二、與會心得:
如附件三、考察參觀活動(無是項活動者略):
無四、建議:
鼓勵推動專題性的小型會議五、攜回資料名稱及內容:
此會議鼓勵最新發現及尚未發表的研究結果,所以,除了會議 流程外,並沒有摘要集也嚴禁照相。計畫編號 NSC98-3112-B-009-005-
計畫名稱 克雷白氏肺炎桿菌第一型纖毛和第三型纖毛交互表現的調控
出國人員
姓名
彭慧玲
服務機構
及職稱
交通大學生物科技學系
會議時間
99 年 1 月 24 日至
99 年 1 月 29 日
會議地點 美國洛杉磯- Ventura
會議名稱
(中文)微生物的訊息傳導
(英文)
Gordon Conference on “Sensory Transduction in Microorganisms”發表論文
題目
(中文)克雷白氏肺炎桿菌 RcsB 傳導的抗酸反應
(英文)
Characterization of the RcsB-mediated acid response in Klebsiella克雷白氏肺炎桿菌第一型纖毛和第三型纖毛交互表現的調控
(NSC 98-3112-B-009-005 ) 九十八年度出國開會報告
The Gordon Conference on “Sensory Transduction in Microorganisms”, January
24-29, 2010.
Hwei-Ling Peng (彭慧玲), Department of Biological Science and Technology, National Chiao Tung University
這個兩年一次的專題會議於一月二十四號至二十九號在美國南加州Ventura舉行,主題為 Sensory transduction in Microorganism,約有一百五十與會人數。五天的會議,有兩個主要議 題(一)細菌鞭毛運轉和趨化性機制;(二)細菌和酵母菌的訊息傳遞。會議於早上九點開始 至晚上九點半左右結束,平均每天有十二個講題,分別有二十鐘、十分鐘和五分鐘的演講;下 午四點至六點為壁報討論時段,是個議程緊湊但內容極為豐富的學術研究會議。擇錄內容和心 得如下:
I. Diversity in Microbial Chemotaxis
- The first section covers the properties of Rhodobacter, E. coli, and P. aeruginosa. “ Rewiring the chemosensory pathways of Rhodobacter sphaeroides” given by Dr J. Armitage from Oxford University UK focused on why 4 CheA and 6 MCP are encoded by 3 operons in the genome and how do they play roles for the motility. He has shown that the different chemosory machineries play similar role but at different parts of the cell. Control mechanism for the location determination has also been studied.
- Dr R. Parales from UC Davis USA reported “Bacterial chemotaxis to environmental pollutants” mainly on the Pseudomonas chemotactic receptor proteins. In E. coli, 4 sets of MCP were identified. On the other hand, a total of 26 chemotactic receptor proteins were identified in Pseudomonas. The equipped with large numbers of chemosensory machineries may ensure the bacteria survive the diverse environmental challenges.
II. Intracellular Signaling in Bacteria
- Using microarray approach, Dr Ann Stock from UMDNJ-Robert Wood Johnson Medical Schooll described their investigation of “ Regulating response regulators”. In general, response regulator (RR) contains in the regulatory domain the conserved residues including Ser, Thr, and Tyr next to the Asp residue for the reception of the phosphate signaling molecule. Approximately 14% of RR contain only the regulatory domain, 12% have enzymatic domain at the C-terminus, while most RR act as transcription factor carrying with a DNA domain.
- M Laub (MIT) introduced in “Specificity and evolution of two-component signaling pathways” his theory about new pathways arise in order to ensure the specificity to prevent from unwanted cross-talk between 2CSs. By comparative analysis of the phosphor-relaying rate at different time-intervals of approximately 4500 2CS pairs, he showed that the 2CS with strong kinetics preference ensure the specificity of the phosphor-relay.
system” mentioned how they measured the feedback regulation using YFP to report the target gene expression and CFP to report the activity of the reference promoter. He concluded that a small molecule of 47 aa is involved in the feed-back regulation for the PhoQ/PhoP signaling pathway. They also demonstrated using 2-hybrid analysis that the small protein MgrB exerted its regulatory role by direct binding to PhoQ. The mgrB deletion conferred the bacteria an abnormal filamentous cell morphology implying an essential role of the MgrB-mediated control.
- L McCarter (U Iowa) reported in “Surface specific gene expression” of Vibrio
parahaemolyticus. They used microarray comparison to differentiate the swarmmer cells, 200~ 400
μm filaments with peri-flagella, and the swimmer cells with polar flagella. The analysis revealed, in addition to the LafK regulon, several groups of the LafK-independent genes including virulence genes, the genes involved in modulation of the c-di-GMP level were also identified in the swarmmer cells. Most interestingly, the secreted collagenase increased production of ~550 fold higher than the swimmer cells.
- U Jenal from U Basel, Switzerland in the talk “Coordination of Caulobacter cresentus cell cycle and pole morphogenesis” described the involvement of second messenger c-di-GMP in the control of the single cell to community stage. The interacting regulation YhjH, YcgR, YedQ, YfiN, YddV, and YegE were demonstrated.
III. Motility Apparatus and Regulation
- Dr. J Liu (U Texas Medical School) in the talk “Molecular architecture of spiral flagellar motor in situ” presented the use of Cryo-EM tomography to build up the model for the assembly of the spiral flagella motor; - D Blair (U Utah) in “Organization and mechanism of the flagellar switch” described the use of EM analysis revealed that the switching machinery is composed of 3 protein complex including 25 copies of FliG, 35 copies of FliM, and 140 copies of FliN.
- R Harshey (U Texas Austin) described in “The cyclic-di-GMP receptor protein YcgR localizes to the flagellar basal body and inhibits chemotaxis in Salmonella“ the use of co-IP analysis
demonstrated an interaction between YcgR and the rotor protein. The in vivo analysis for the
expression of YcgR-GFP in either fliM, fliG, or fliN deletion mutant further supported the presence of an interaction between YcgR and FliM and FliG. In contrast to the interaction between B. subtilis EpsE and FliG, of which EpsE acts as clutch to interact with FliG at late stage to inhibit the flagellar switch leading to the biofilm formation, the Salmonella YcgR interacts with FliM in the early stage to inhibit dhemotaxis.
IV. Intracellular Signaling in Eukaryotic Microorganisms
The 2CS of eukaryotic microorganism are mainly hybrid type and the phosphate group transferred from histidine kinase to Hpt module and then to response regulator (RR). Two types of RR are
transcription factors and MAPK pathway effectors involved in stress response.
- In the talk “ Osmosensory signal transduction in the budding yeast”, Dr H. Saito (U Tokyo) introduced the signal transduction pathway in order for the budding yeast growing in ripen fruit in
which the sugar concentration could reach to 5 M. The osmostress in the hyperosmotic environment was received by the sensor kinase and the signal relay eventually led to glycerol accumulation.
- In the talk “HKs, cell wall biosynthesis, and relationships to Candida albicans and
candidiasis”, R. Calderone (Georgetown UMC) introduced the fungal HK could be classified into 11 families on the basis of phylogenic analysis. Among them, ChKI, SinI, and NiKI are conserved and required for virulence. In the study, they have shown that ChkI is required for invasive candidiasis, QS, biofilm formation, adherence of Candida to host cells via regulating the enzymes for reconstitution of the cell wall system (rich in mannoprotein~ 30-40%).
- K. Shiozaki (UC Davis) reported in “ Stress sensing and response in fission yeast S. pombe” the use of 2-hybrid analysis to identify the SinI (stress MapK-interacting protein I)- dependent stress response pathway; - K. Borkovich (UCR) in “Two-component systems in Neurospora” introduced that11 hybrid HK, 1 Hpt, and 2 RR have been identified in Neurospora. She also described in details the signal relayed following the path from HK to Hpt then to RR and finally to the MAPK cascade.
V. Intercellular Communication
In ”What’s new with QS”, E. P. Greenberg (UW) introduced the term “Sociomicrobiology” of which QS allows coordination of group activities which induced by the signals of LuxI family. He described the discovery of a new class Aryl-HSL from Rhodopseudomonas palustris for the synthesis of QS products including light, exoenzymes, toxins, and aggregation factors. The bioassay excluded the signals of LuxN- 3-OH-C4, LuxR-C5-C8, RhlR-C4-C6, LasR C8-14, GtaR C14-16, or TraR C4-14. Finally, p-coumarate p-coumaroyl-HSL was shown to be the QS inducer for the RpaI-RpaR mediated regulon.
- K.L. Visick described in “The regulatory network controlling biofilm formation and
colonization by Vibrio fisheri” the identification of a RscS hybrid HK. He has shown the sensor kinase regulates the signal transduction control for the synthesis of polysaccharide (Syp), which is an
essential step for the bacterial colonization and biofilm formation. He also showed the experiments used including knock out the sypG gene, overexpression of rscS or sypE, interaction of SypE and SypA (the anti-sigma antagonist) to establish the phosphorelay from RscS to SypG-P then to SypA-P, and finally affects the biosynthesis of the polysaccharide, biofilm formation and the development of a symbiosis lifestyle.
- F Yildiz (UC Santa Cruz) in the talk “Regulation of biofilm formation in Vibrio cholerae” the role of c-di-GMP signaling potential effectors including PilZ (PlzA, PlzB, PlzC, PlzD, and PlzE) and RXXD, CdgG riboswitch in the control of the expression of the 62 GGDEF, EAL, HD-GYP domain proteins. He showed that the deletion of PilZ protein, CdgG had no effect on the colony rugosity or the biofilm formation. He has also shown that VpsT dimmer functions as a c-di-GMP effector to
determine the colony conversion from smooth to rugose.
- D. Kearns (Indiana U) described in “Bistable expression of motility genes in Bacillus subtilis” the heterogenous population phenotype: some are motile, while some are in chains and non-motile. To investigate how the gene expression differs in the different cell types, the reporters including
P-flagellin-GFP and P-autolysin-CFP were used. The monitoring in short cells and long chain cells revealed that Sigma D dependent gene expression is bi-stable.
- D. Zusman (UC Berkeley) also reported the use of reporter systems in “Coupling the FrzCD receptor to the A motility”. He has shown that using the A motility complex GST-FrzCD has pulled down from the Myxococcus lysate AlgZ and AgmU. The interaction with FrzCD was further
demonstrated using AglZ-YFP or AgmU-mcherry. He has shown that both proteins co-localized and the clusters appeared stationary as cells move forward. In addition, AgmU decorates a double helix showing a same periodicity of MreB which supports the previous finding that the A motility is MreB dependent. Finally, he showed the use of AgmU-GST in the pull down assay that in addition to FrzCD, AglZ, AglT, AglW and the MotAB homologues are required for the A motility.
VI. Pathogenesis and Virulence mechanisms
- Dr E. Groisman from Washington University gave an overview of “Controlling the activity of two-component regulatory systems”. He introduced genetic basis differences for the different control between related species, which include the presence or absence of the gene, differential regulation, allelic difference, and different genetic context. In Salmonella, PmrD-dependent pathway is involved in the regulation of the polymyxin resistance. While in E. coli, the drug resistance is mediated by a PmrD-independent control. However, although sharing less than 50% sequence identity, Ec-PmrD in
Salmonella works as Salmonella PmrD. On the other hand, Ec-PmrBc which shares much higher
sequence identity with Salmonella PmrBc exerts different dephosphorylation activity.
- “Inter-kingdom chemical signaling in bacterial pathogenesis” by Dr V. Sperandio from UTS Medical Center introduced the identified signaling molecules such as AI3, Epinepharine,
Norepinepharine, and Tyrosine in GI tract.
- “When is a predator not a predator? Prey dependent versus axenic growth in Bdellovibrio” given by Dr. L. Sockett from U Nottingham UK introduced the life styles of the 3.8 Mbp genome G(-) bacteria. They are host independent (HI) growth, which takes 2 wk on peptone-rich medium and host dependent (HD) growth. The microarray analysis has identified predation specific genes namely predatosome, which include genes for the expression of type IV pilus and flagella, 15 PilZ domain proteins, 1 EAL and 5 GGDEF domain proteins, and MreB, MreC, and MreD proteins.
Overall, I was impressed and overwhelmed with the stimulations of most of the reports. Most
importantly, I find the meeting extremely informative and inspiring for the future study of the sensory transduction in bacteria.
國科會補助專題研究計畫成果報告自評表
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)
、是否適
合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
■達成目標
□未達成目標(請說明,以 100 字為限)
□實驗失敗
□因故實驗中斷
□其他原因
說明:
2. 研究成果在學術期刊發表或申請專利等情形:
論文:□已發表 ■未發表之文稿 □撰寫中 □無
專利:□已獲得 □申請中 ■無
技轉:□已技轉 □洽談中 ■無
其他:
(以 100 字為限)
3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價
值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以
500 字為限)
Three manuscripts are to be submitted for publication. Many findings include FimK negatively but c-di-GMP positively regulated both fimbriae expressions; PecSM affected negatively the type 3 fimbriae expression; MrkI regulates the counter-expression; RcsB negatively regulates fimB expression; OmpA and OmpC affected MrkA expression and biofilm formation. These allow unravel a regulatory circuit on both fimbriae expressions and intervening targets for antimicrobial agents.