JOURNAL OF EXPERIMENTAL MARtNE BIOLOGY
ELSEVIER
Journal of Experimental Marine Biology and Ecology, 199 (1996) Ill-122
AND ECOLOGY
Effects of salinity and ontogenetic movements on
strontium:calcium
ratios in the otoliths of the Japanese eel,
Anguilla japonica Temminck and Schlegel
Wann-Nian Tzeng
Departmenr of Zoology, College of Science, National Taiwan Univerdy, Taipei, 10617, Taiwan, ROC Received 6 June 1995; revised 27 November 1995; accepted 28 November 1995
Abstract
To study the mechanism of Sr incorporation into otoliths of the Japanese eel, Anguillajaponica, a total of 100 elvers collected from an estuary were reared in the laboratory at salinities of 0, 10, 25 and 35%0 for approximately seven months. The elvers grew from 56 mm TL to 100-300 mm TL. Twenty elvers were randomly selected and the Ca and Sr concentrations of their otoliths were analyzed from the primordium to the edge, using an electron microprobe equipped with a four-channel wavelength-dispersive spectrometer. Sr/Ca ratios in the otoliths of eels reared in various salinities were much lower than the ratio of 15 X IO-’ observed in elvers about one month before they arrived at the estuary. The irreversibility of Sr/Ca ratios at 35%0 salinity in this experiment indicated that the drastic change of the SrlCa ratios in otoliths of elvers was not due to the reduction of salinity in the coastal waters, but more likely to the development from leptocephalus to glass eel. The mean Sr/Ca ratios in the new increments of the otoliths of eels during the rearing period were highly correlated with salinity (S), [Sr/Ca] X lo7 = 3.797 + 0.14s (n = 20, r = 0.77), which can be used to predict elver movements and habitat utilization rates. Keywords: Japanese eel; Otolith; Migration; Strontium; Salinity; Development stage
1. Introduction
As in other teleost fishes, the otoliths of eels are composed of calcium carbonate in the crystal form of aragonite (Carlstrom, 1963; Degens et al., 1969; Morales-Nin, 1987; Lecomte-Finiger, 1992) and are deposited daily in alternating protein-rich and carbon- ate-rich layers as the fish grows (Pannella, 1971; Campana and Neilson, 1985; Lee and Lee, 1989; Tsukamoto, 1989). This variable deposition enables the daily age of the fish to be determined and their growth history to be studied (Tabeta et al., 1987; Tsukamoto, 0022-0981/96/$15.00 0 1996 Elsevier Science B.V All rights reserved
112 W. Tzeng I .I. Exp. Mar. Bid. Ed. 199 (1996) 111-122
1989, 1990; Tzeng, 1990; Jones, 1992; Tzeng and Tsai, 1992). In the formation of aragonite crystal a few trace elements co-precipitate with calcium carbonate (Gunn et al., 1992; Ophel and Judd, 1967; Rosenthal, 1956, 1963; Smith et al., 1979; Toole and Nielsen, 1992). Sr is one of the most common elements to replace Ca because it has a similar ionic radius and valence (Amiel et al., 1973). Recently, Sr/Ca ratios in otoliths, in relation to ambient temperature and together with daily ages, have been used to reconstruct the past environmental histories of fishes (Radtke et al., 1990; Townsend et al., 1992). Because sea water contains more Sr than freshwater (Tzeng and Tsai, 1994), the change in the Sr/Ca ratios in otoliths has also been used to study the migratory history of diadromous fishes such as American and European eels (Casselman, 1982), Hawaiian gobies (Radtke et al., 1988), salmonids (Kalish, 1990) and striped bass (Secor, 1992) in their movements between sea water and freshwater environments. These studies indicated that Sr/Ca ratios in otoliths have the potential for retrieving the past environmental history of the fish. However, the incorporation of Sr in otoliths is a complicated biogeochemical process influenced by environmental factors and physiolog- ical conditions, and Sr/Ca ratios as environmental predictors in fishes have only rarely been experimentally validated (Kalish, 1989, 199 1; Gallahar and Kingsford, 1992; Radtke and Shafer, 1992; Sadovy and Severin, 1992).
The Japanese eel, Anguilla japonica Temminck and Schlegel, is a catadromous fish. Its life cycle is similar to the European eel, Anguilla anguillu, and includes five stages: leptocephalus, glass eel, elver, yellow eel and silver eel (Bertin, 1956). The Japanese eel spawns in the waters west of Mariana Island (Tsukamoto, 1992) and the leptocephalus larvae drift with the North Equatorial Current and Kuroshio Current (Kimura et al.,
1994). As they approach the continental shelf, leptocephali metamorphose into glass eels and leave the Kuroshio for coastal waters (Tsukamoto and Umezawa, 1994). The glass eels become elvers in the estuaries (Bertin, 1956). During this long journey, the eels experience a salinity gradient from oceanic to coastal waters and a developmental transition from leptocephali to glass eels. Otolith microchemistry analyses indicate that the Sr/Ca ratios in otoliths of elvers change drastically during their migration from spawning ground to the river estuary (Otake et al., 1994; Tzeng and Tsai, 1994). It has been suggested that this drastic change in Sr/Ca ratios in otoliths is the result of the metamorphosis of the leptocephalus to a glass eel, or the result of a reduction of salinity in coastal waters (Otake et al., 1994; Tzeng and Tsai, 1994).
This study attempted to elucidate the factors affecting the Sr/Ca ratios in otoliths of the Japanese eel during its migration from ocean to river by examination of Sr and Ca concentration in the otoliths of elvers reared in waters of various salinity and to determine the reliability of Sr/Ca ratios as an environmental indicator for wild eels.
2. Materials and methods
Due to difficulty in collection of leptocephali and keeping them alive, elvers collected from the Tanshui River estuary in northern Taiwan on 8th March 1992 were used for the study. The elvers were at pigmentation stage VA with no pigment except caudal spots (Strubberg, 1913; Bertin, 1956). Their total lengths (TL) ranged from 52.2 mm to 58.9
W Tzeng I J. Exp. Mar. Bid. Ecol. 199 (1996) Ill-122 113
Table 1
Mean (5S.D.) total length and otolith diameter of the eels reared from elvers for 7 months in growth chambers at temperatures of 22-23°C and 27-28°C and salinities of 0, 10, 2.5 and 35%0
Treatment Temperature” (“C)
Salinity (%o)
No. fish survived Total length (mm) Otolith diameter (pm) I 28 2 28 3 28 4 28 5 28 6 23 7 23 8 23 9 23 IO 23 35 10 25 10 10 9 0 IO 0 6 35 10 25 9 IO 10 0 IO 35 9 143.45523.39 163.60228.52 184.11?35.04 164.95527.87 184.67t58.19 126.75129.22 129.33-+21.30 144.9Ok23.98 145.65220.52 148.44528.87 85 I .67 + I 12.48 961.632138.82 1000.10-t184.62 945.692 125.86 990.522213.78 761.24-+119.99 765.55~ 96.49 820.362 104.34 805.691- 97. IO 832.11-+141.95
The stocking density is 1 I-’ except the treatments of Nos. 5 and IO which were 0.5 I ’ “Temperature during daytime. Night temperature is 1°C lower.
mm with a mean of 55.3 mm. After an acclimation period of 2 days in brackish water of salinity 11..57%0 under natural photoperiod, 100 randomly selected elvers were divided into ten treatment groups, each containing ten eels and reared in growth chambers at two temperature regimes of 22-23°C and 27-28°C and salinities of 0, 10, 25 and 35%0. The stocking density was one fish 1-l except treatment Nos. 5 and 10 where the density was 0.5 1-l (Table 1). The temperature and salinity were set by referring to those where the leptocephali and glass eels were sampled (Kimura et al., 1994; Otake et al., 1994; Tzeng and Tsai, 1994). The salinities of the rearing water were prepared from an approximately 32%0 natural sea water adjusted with tap water and crude salt (dried natural sea salt obtained from Tainan table-salt processing plant). The fish were fed to satiation once a day with formulated larval eel food supplemented with live tubifex worms. Faeces were siphoned daily and the rearing water was changed at intervals of approximately 2 weeks. The water level was maintained constant by adding water periodically. The Sr and Ca concentrations and salinity of the rearing water (Table 2) were measured with an atomic absorption spectrophotometry (Perkin-Elmer 703) and salinometer (WTW micropro- cessor conductivity meter LF196), respectively.
The eels were reared for approximately 7 months and the experiment ended on 12 October 1992. The surviving eels were sacrificed, their total lengths were measured to
Table 2
Relationship between strontium (Sr) and calcium (Ca) contents, Sr/Ca ratios and the salinity of the rearing water Salinity (%G) Sr (ppm) Ca (ppm) Sr/Ca(XlO ‘) 0 0.17 27.67 6.14 10 1.9 181.0 10.50 25 3.95 3 14.5 12.56 35 5.4 47 1.67 I I .45
114 W. Tzeng I J. Exp. Mar. Biol. Ecol. 199 (1996) III-122
nearest 0.01 mm with calipers and they were weighed to 0.01 g with an electronic balance. Sagittal otoliths were removed 1 week after fixation in 95% alcohol.
The maximum diameter of each of the otoliths was measured with a computer-aided image processing system (VIPRO 512) and the otoliths were then embedded in resin. The procedures for embedding, polishing and etching of the otolith for Sr and Ca content measurements followed those described by Tzeng (1990) and Tzeng and Tsai (1992, 1994). The Sr and Ca contents in otoliths of the 20 randomly selected eels were analyzed using an electron microprobe equipped with a four-channel wavelength dispersive spectrometer (WDS; Shimadzu-ARL EMX-SM). In the analyses of Sr and Ca, SrTiO, and CaSO, were used as standards. The weight percentage composition of CaSO, equals 41.19% CaO plus 58.81% SO, and SrTiO, equals 56.46% SrO plus 43.54% TiO,. The electron beam was focused on an area approximately 5 ,um in diameter at intervals of approximately 20 pm along the frontal section of the otolith maximum radius from primordium to edge. The accelerating voltage of electron was 15 kV with specimen current of 0.01 ,uA. The counts for spectrometers of the elements were evaluated using five 4 s integration periods and at the wavelengths of 3.3685A (Ca-Ka) and 6.8628A (Sr-La), respectively. In the ZAF correction procedures (Chen and Tung, 1984), the weight percentages of SrO and CaO in otolith were calculated using a constant of CO, at 43.97%. The relative contents of Sr and Ca in a percentage of otolith weight were then estimated.
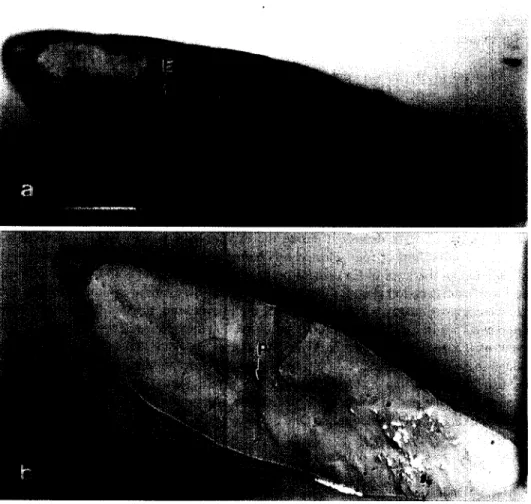
In order to establish the positions of the primordium, elver checks and microprobe spots on the otoliths before and after the WDS analysis, each otolith was photographed with transmitted and reflected light under a light microscope (Fig. la, b). The otolith was then etched with 5% EDTA (ethylene diamine tetra-acetate; pH adjusted with NaOH to 7.4) and photographed with transmitted light to reconfirm the primordium and elver checks (Fig. lc) and to discriminate the leptocephalus metamorphosis check from the transition of otolith microstructures and Sr/Ca ratios (Otake et al., 1994; Tzeng and Tsai, 1994). With the aid of the microphotographs, the distances from the primordium to each of the microprobe spots and the checks were measured precisely. The Sr/Ca ratios in the new increments of otolith, which encompassed the rearing period were then examined.
The differences between treatments for fish growth and otolith Sr/Ca ratios were examined using an analysis of variance (ANOVA). A forward stepwise regression analysis was used to test the relationships between otolith Sr/Ca ratios and temperature, salinity, ambient Sr/Ca ratios, fish length and weight and otolith diameter. The significance of differences in otolith Sr/Ca ratios between salinities was further tested by Scheffe’s multiple range analysis (Sokal and Rohlf, 1969).
3. Results 3. I. Growth
After 7 months of rearing, 93 eels had survived. The elvers grew from approximately 56 mm TL to 100-300 mm TL. Mortality rates of elvers did not differ among treatment groups (P>O.O5). The somatic and otolith growth rates were significantly different
W Tzeng / J. Exp. Mar. Bid. Ed. 199 (1996) III-122
Fig. 1. Microphotographs of the otolith from a 162 mm TL eel showing (a, c): primordium (P); leptocephalus metamorphosis check (M); elver check (E) and (b) 17 microprobe spots. Scale bar= 150 pm. Photos a, b and c were taken for different purposes, as described in the text.
116 W. Tzeng / J. Exp. Mar. Bid. Ed. 199 (1996) III-122
between different temperatures (P < O.Ol), salinities (P < 0.05) and rearing densities (PcO.05) and there was no interaction among these three treatments (P>O.O5). Both mean somatic and otolith growth rates were greater at 27-28°C than at 22-23°C at lower than at higher salinities and at lower than at higher rearing densities (Table 1). 3.2. Changes in Sr and Ca contents and SrlCa ratios in otoliths during development
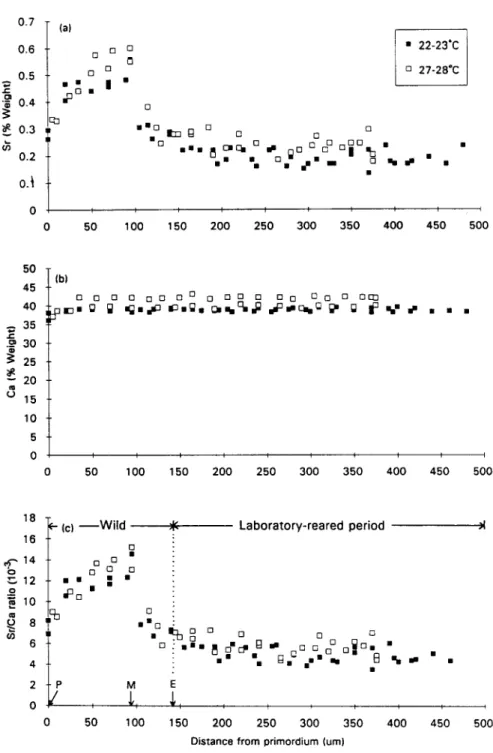
The otolith of a 162 mm TL eel reared from 56 mm TL at the salinity 10%0 and the temperature 27-28°C shows both leptocephalus metamorphosis and elver checks at approximately 89 pm and 145 ,um from the primordium, respectively, which were discernible from the change of microstructural pattern of their growth increments (Fig. 1). The elver check was deposited when the glass eel arrived at the estuary (Tzeng and Tsai, 1994; Tzeng et al., 1994) and provides a convenient wp to distinguish otolith increments deposited before and during the experimental period. The new increments outside the check were deposited during the experimental period. Both Sr content and Sr/Ca ratio in otoliths of four selected eels before the experimental period show a wide fluctuation; but the level of Ca content in the otoliths was stable both before and during the experimental period (Fig. 2). The Sr content and Sr/Ca ratio in otoliths of the four eels were approximately 0.3 and 8 X lop3 at the primordium, reached the highest values of 0.6 and 15 X lo-’ at the metamorphosis check and then dramatically decreased to low levels of 0.3 and 8X lo-’ at the elver check (Fig. 2). This indicated that the drastic change in Sr content and Sr/Ca ratio in elver otoliths occurred about one month before the fish arrived at the estuary. The highest Sr/Ca ratios in the otoliths probably correspond to the period when the leptocephalus metamorphosed to the glass eel stage before moving into coastal waters (Otake et al., 1994; Tzeng and Tsai, 1994). 3.3. Effects of salinity on otolith SrlCa ratios
ANOVA indicated that Sr/Ca ratios in new increments of the otolith deposited during the experimental period were not significantly different between temperatures (P>O.5). They were, however, significantly different between salinities (PcO.01) but there were no interactions between temperature and salinity (P>O.9). Forward stepwise regression analysis also indicated that of the six variables temperature, salinity, ambient Sr/Ca ratio, fish length and weight and otolith diameter, only salinity influenced the Sr/Ca ratios in otoliths.
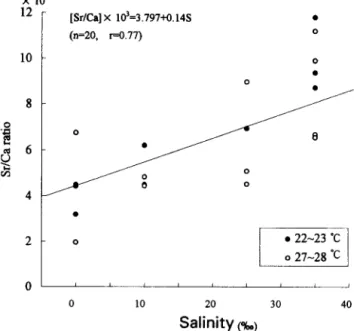
The relationship between otolith Sr/Ca ratio and salinity (S) was estimated to be [Sr/Ca] X lo3 = 3.797+0.14S (n =20, t-=0.77) (Fig. 3). Mean Sr/Ca ratios in new increments of otoliths increased significantly from 4.20X 10m3 in the freshwater to 9.27x 1O-3 at 35%0 salinity (Table 3) and this result indicated that Sr/Ca ratios in otoliths could be used as an environmental indicator for studying the migration of eels between freshwater and sea water. On the other hand, the Sr/Ca ratios of the new increments in otoliths of eels reared at the various salinities and both 22-23°C and 27-28°C were much lower than the highest value of 15 X lo-’ in the otoliths of the elvers 1 month before they arrived at the estuary (Figs. 2,3, Table 3). The irreversibility of Sr/Ca ratios in otolith of the eel at the salinity of 35%~ in the rearing experiment indicated that the drastic change of the Sr/Ca ratios in otoliths of elvers may be not due
W Tzeng I J. Exp. Mar. Biol. Ecol. 199 (1996) 1 I I - 122 117 0.7 - (al 0.6 -_ 0 no a 0.5 -- q z , .$ 0= . ; . 0.4 -. P 3 0 el z 35 .$ 30 -- 3 25 -- * ;;; 20 -- v 15 -- 10 -- 5 -- 0 1 1 0 50 100 150 200 250 300 350 400 450 500 18 16 % l4 G 12 0 f 10 0” 8 ZJ 6
-Wild __)(c___ Laboratory-reared period d
M E
1: L: ,
0 50 100 150 200 250 300 350 400 450 500 Distance from primordium (urn)
Fig. 2. Changes in Sr (a), Ca (b) contents and Sr/Ca ratios (c) in otoliths during development. Sr and Ca contents were measured from the primordium to the edge of the otoliths of four selected eels (143, 162, 165 and 173 mm in total length) reared from elvers at 10% salinity and two temperature regimes of 22-23°C and 27-28°C. E denotes the mean position of elver check; M, leptocephalus metamorphosis check and P,
118 W Tzeng I .I. Exp. Mar. Bid. Ed. 199 (1996) Ill-122
0 10 20 30 40
Salinity(s)
Fig. 3. Relationship between otolith Sr/Ca ratio and salinity.
to the reduction of salinity in the coastal waters, but more likely to the transition from leptocephalus to glass eel.
4. Discussion and conclusions
The relationship between Sr/Ca ratios in the otoliths of reared eels and ambient salinity (Fig. 3) is consistent with the change of Sr/Ca ratios in otoliths of the wild eels that experience changes in salinity during their migration from the estuary to the river (Tzeng and Tsai, 1994). These data also validate the use of Sr/Ca ratios in otoliths as environmental indicators to reconstruct the migratory history of the eel between freshwater and sea water. This change in Sr/Ca ratios in otolith has also been found in other diadromous fishes e.g., American and European eels (Casselman, 1982), salmonids (Kalish, 1990), Hawaiian gobies (Radtke et al., 1988) and striped bass (Secor, 1992) Table 3
Scheffe’s multiple range analysis of mean Sr/Ca ratios in new increments of the otoliths of the Japanese eel reared in different salinities. Groups of salinities sharing the same column of asterisks are homogeneous
Salinity (o/00) Sample size Sr/Ca ratio (X lo-‘) Homogeneous group
0 5 4.20 *
10 4 4.99 *
25 4 6.57 **
W. Tzeng I .I. Exp. Mar. Biol. Ecol. 199 (1996) III-122 I19
during their migration between freshwater and sea water. These results indicate that the relationship between Sr/Ca ratios in fish otoliths and ambient salinity may be an universal rule in diadromous fishes.
Incorporation of Sr into otoliths is a complicated biogeochemical process influenced by physical factors such as temperature, salinity and water chemistry, as well as by biological factors such as genetics, developmental stage, growth rate, food and physiological condition of the fish (Dodd, 1967; Gallahar and Kingsford, 1992; Kalish, 1989; Radtke and Shafer, 1992; Sadovy and Severin, 1992; Yamada et al., 1979). The Sr/Ca ratios in otoliths of elvers drastically decreased from a high value of 15 X IO-’ to a low level of approximately 8 X lop3 about 1 month before the fish arrived at the estuary (Fig. 2). It was suggested that this decrease corresponded to the period when the leptocephali metamorphose to glass eels and leave the warm Kuroshio Current for cold coastal waters (Otake et al., 1994; Tzeng and Tsai, 1994). During this period, salinity, temperature and the change in developmental stage may all have the potential to influence the Sr/Ca ratios of the otoliths.
The experimental eels were exposed to temperatures of 27-28°C and a salinity of 35%0, which were very close to the 27.0-29.O”C and 34.534.7%0 of the natural waters where the leptocephali were collected (Otake et al., 1994). The mean Sr/Ca ratios in the new increments of otoliths of the eels reared in 35%0 sea water (Fig. 3 and Table 3) were much lower than the highest value of 15 X lo-’ observed in otoliths of elvers before the experimental period (Fig. 2). The temperature and salinity in the natural environment gradually decrease during the elver migration from the Kuroshio area to coastal waters. It has been reported elsewhere that the Sr/Ca ratio in the otolith is inversely related to ambient temperature (Radtke et al., 1990; Townsend et al., 1992, 1995; Tzeng, 1994). Hence if the change in the Sr/Ca ratios in otoliths of elvers is due to temperature, the Sr/Ca ratios in the otoliths from metamorphosis to elver check (Fig. 2) should increase but not decrease because during this period the fish migrate toward a low temperature. On the other hand, if the change in Sr/Ca ratios in otoliths of elvers resulted from the salinity gradient, when the salinity of the rearing water was elevated to the salinity of 35%0, the Sr/Ca ratios in otoliths of the eel during the experimental period should increase to the highest value observed in the otoliths of elvers before the experimental period. This was not the case in the present study. Therefore, temperature and salinity are probably not the primary factors determining the drastic change of Sr/Ca ratios in otoliths of elvers during their migration from the open ocean to the river. An alternative possibility is the saltatory developmental change from leptocephalus to glass eel.
Until now, the leptocephali of Japanese eel or other species of Genus Anguillu have only rarely been collected and no experiments have been done to validate the mechanism of Sr incorporation in otoliths at the time of metamorphosis. Strontium metabolism in the leptocephalus stage has been considered to be different from that in the elver stage (Hulet, 1978; Keys and Willmer, 1932; Ophel and Judd, 1967; Utida and Hirano, 1973). The leptocephalus is known to contain extensive amounts of gelatinous extracellular matrix composed of sulfated glycosaminoglycans (GAGS) which are broken down during metamorphosis (Pfeiler, 1984, 1986). Since GAGS have an affinity for alkali earth elements, particularly Sr*+ (Comper and Laurent, 1978; Nishizawa, 1978; Hascall and Hascall, 1981; Toole, 198 l), GAGS breakdown may reduce the absorption of Sr’ ’
120 W. Tzeng / J. Exp. Mar. Biol. Ecol. 199 (1996) Ill-122
from sea water and result in a drastic decrease in otolith Sr content and, consequently, in Sr/Ca ratio (Otake et al., 1994). In this study, Sr/Ca ratios in otoliths of elvers increased gradually from the primordium, reached a peak prior to the metamorphosis check and then dropped suddenly thereafter (Fig. 2). Apparently, the drastic change of Sr/Ca ratios in elver otoliths corresponds closely to the metamorphosis from leptocephalus to glass eel.
In conclusion, the drastic change of the Sr/Ca ratios in otoliths of elvers is most likely due to the transition from leptocephalus to glass eel. Otolith Sr/Ca ratios at the elver stage are directly related to the ambient salinity and they can be used to predict elver movements and habitat utilization rates.
Acknowledgments
This study was financially supported by the National Science Foundation, Republic of China (Project NSC 82-0211-B002-044). The author expresses his gratitude to Miss H.F. Wu, Mr. Y.T. Wang, Miss C.H. Wang, Miss C.E. Wu and other colleagues at the laboratory of Fisheries Biology, Department of Zoology, National Taiwan University (NTU) for their assistance in the laboratory, to Dr. C.H. Chen, Department of Geology, NTU for assistance on WDS microprobe analysis and to Dr. C.F. Tsai and the anonymous referees for reviewing the manuscript.
References
Amiel, A.J., G.M. Friedman and D.S. Miller, 1973. Distribution and nature of incorporation of trace elements in modem aragonite corals. Sedimentology, Vol. 20, pp. 47-64.
Bertin, L., 1956. Eels - a biological study. Cleaver-Hume Press, London, 192 pp.
Campana, S.E. and J.D. Neilson, 1985. Microstructure of fish otoliths. Can. 1. Fish. Aquar. Sci.,Vol. 42, pp. 1014-1032.
Carlstrom, D., 1963. A crystallographic study of vertebrate otoliths. Biol. Bull., Vol. 25, pp. 441-463. Casselman, J.M., 1982. Chemical analyses of the optically different zones in eel otoliths. In, Proceedings of
rhe 1980 Noah American Eel Conference, edited by K.H. Loftus. Ont. Minist. Nat. Resour., Ont. Fish. Tech. Rep. Ser. 4, pp. 74-82.
Chen, C.H. and T.C. Tung, 1984. On-line data reduction for electron microprobe analysis. Acra Geol. Taiwan,
Vol. 22, pp. 196-200.
Comper, W.D. and T.C. Laurent, 1978. Physiological function of connective tissue polysaccharides. Physiol. Rev., Vol. 58, pp. 255-315.
Degens, E.T., W.G. Deuser and R.L. Haedrich, 1969. Molecular structure and composition of fish otoliths.
Mar. Biol., Vol. 2, pp. 105-l 13.
Dodd, R.J., 1967. Magnesium and strontium in calcareous skeletons: a review. J. Palaeonfol., Vol. 41, pp. 1313-1329.
Gallahar, N.K. and M.J. Kingsford, 1992. Patterns of increment width and strontium:calcium ratios in otoliths of juvenile rock blackfish, Girella elevara (M.). J. Fish Biol., Vol. 41, pp. 749-763.
Gunn, J.S., JR. Harrowfield, C.H. Proctor and R.E. Thresher, 1992. Electron probe microanalysis of fish otoliths: evaluation of techniques for studying age and stock discrimination. J. Enp. Mar. Biol. Ecol., Vol. 158., pp. l-31.
W. Tzeng I J. Exp. Mar. Biol. Ecol. 199 (1996) Ill-122 121
Hascall, V.C. and G.K. Hascall, 1981. Proteoglycans. In, Cell biology of exrracellular marrix, edited by E.D.
Hay. Plenum Press, New York, pp. 39-63.
Hulet, W.H., 1978. Structure and functional development of the eel leptocephalus Ariosoma balearicum (De La
Roche, 1809). Phil. Trans. R. Sot. Land. (Ser. B), Vol. 282, pp. 107-138.
Jones, CM., 1992. Development and application of the otohth increment technique. In, Orolirh microstructurr examination and analysis, edited by D.K. Stevenson and SE. Campana. Can. Spec. Publ. Fish. Aquat. Sci.,
Vol. 117, pp. I-11.
Kalish, J.M., 1989. Otolith microchemistry: validation of the effects of physiology, age and environment on otolith composition. J. Exp. Mar. Biol. Ecol., Vol. 132, pp. 151-178.
Kalish, J.M. 1990. Use of otolith microchemistry to distinguish the progeny of sympatric anadromous and non-anadromous salmonids. U.S. Fish. Bull., Vol. 88, pp. 657-666.
Kalish, J.M., 1991. Determinants of otolith chemistry: seasonal variation in composition of blood plasma, endolymph and otolith of bearded rock cod Pseudophysis barbarus. Mar. Ecol. Prog. Ser., Vol. 74, pp.
137-159.
Keys, A.B. and E.N. Willmer, 1932. Chloride secreting cells in the gills of fishes, with special reference to the common eel. J. Physiol. Lond., Vol. 76, pp. 368-378.
Kimura, S., K. Tsukamoto and T. Sugimoto, 1994. A model for the larval migration of the Japanese eel: roles of the trade winds and a salinity front. Mar. Biol., Vol. 119, pp. 185-190.
Lecomte-Finiger, R., 1992. Growth history and age at recruitment of European glass eels (Anguilla anguilla)
as revealed by otolith microstructure. Mar. Biol., Vol. 114, pp. 205-210.
Lee, T.W. and K.S. Lee, 1989. Daily growth increments and lunar pattern in otolith of the eel, Anguillu
japonica, in freshwater. Bull. Korea Fish Sot., Vol. 22, pp. 36-40.
Morales-Nin, B., 1987. Ultrastructure of the organic and inorganic constituents of the otoliths of the sea bass. In, Age and growfh of jish, edited by R.C. Summerfelt and G.E. Hall, Iowa State University Press, Ames,
IA, pp. 33 I-343.
Nishizawa, K., 1978. Marine algae from a viewpoint of pharmaceutical studies. Jup. J. Phycol., Vol. 26. 73-78.
Ophel, I.L. and J.M. Judd, 1967. Skeletal distribution of strontium and calcium and strontium/calcium ratios m various species of fish. In, Strontium Merubolism, edited by J.M.A. Lenihan, J.F. Loutit and J.H. Martin. Academic Press, London, pp. 101-109.
Otake, T., T. Ishii, M. Nakahara and R. Nakamura, 1994. Drastic changes in otolith strontium/calcium ratios in leptocephali and glass eels of Japanese eel Anguilla juponica. Mar. Ecol. frog. Ser., Vol. 113, pp.
1899193.
Pannella, G., 1971. Fish otoliths: daily growth layers and periodical patterns. Science, Vol. 173, pp.
1124-I 127.
Pfeiler, E., 1984. Glycosaminoglycan breakdown during metamorphosis of larval bonefish Albula. Mar. Biol. Lerr., Vol. 5, pp. 241-249.
Pfeiler, E., 1986. Towards an explanation of the developmental strategy in leptocephalus larvae of marine teleost fishes. Environ. Biol. Fish, Vol. 15, pp. 3-13.
Radtke, R.L., R.A. Kinzie III and S.D. Folsom, 1988. Age at recruitment of Hawaiian freshwater gobies.
Environ. Biol. Fish.,Vol. 23, pp. 205-213.
Radtke, R.L., D.W. Townsend, S.D. Folsom and M.A. Morrison, 1990. Strontium: Calcium ratios in larval
herring otohths as indicators of environmental histories. Environ. Biol. Fish., Vol. 27, pp. 51-61. Radtke, R.L. and D.J. Shafer, 1992. Environmental sensitivity of fish otolith microchemistry. Ausr. J. Mar.
Freshwater Rrs., Vol. 43, pp. 935-951.
Rosenthal, H.L., 1956. Uptake of Ca4’ and Sr”’ from water by freshwater fishes. Science, Vol. 126, pp. 699-700.
Rosenthal, H.L., 1963. Uptake, turnover and transport of bone seeking elements in fishes. Ann. N.Y. Acad. Sci..
Vol. 109, pp. 278-293.
Sadovy, Y. and K.P. Severin, 1992. Trace elements in biogenic aragonite: correlation of body growth rate and strontium levels in the otoliths of the white grunt, Haemulon plumieri (Pisces: Haemulidae). Bull. Mar. Sci..
Vol. 50, pp. 237-257.
Secor, D.H., 1992. Application of otolith microchemistry analysis to investigate anadromy in Chesapeake Bay striped bass Morone suxarilis. U.S. Fish. Bull., Vol. 90, pp. 798-806.
122 W. Tzeng I J. Exp. Mar. Biol. Ecol. 199 (1996) I1 I - 122
Smith, S.V., R.W. Buddemeier, R.C. Redalje and J.E. Houck, 1979. Strontium-calcium thermometry in coral skeletons. Science, Vol. 204, pp. 404-406.
Sokal, R.R. and F.J. Rohlf, 1969. Biometry. W.H. Freeman, San Francisco, CA.
Strubberg. A.C., 1913. The metamorphosis of elvers as influenced by outward conditions. Meddr. Kommn. Huvunders. Ser. Fisk., Vol. 4, pp. l-l I.
Tabeta, O., K. Tanaka, J. Yamada and W.N. Tzeng, 1987. Aspects of the early life history of the Japanese eel
Anguilla japonica determined from otolith microstructure. Nippon Suisun Gakkaishi, Vol. 53, pp. 17277 1734.
Toole, B.P., 1981. Glycosaminoglycans in morphogenesis. In, Cell Biology of Exrracellular Matrix, edited by E.D. Hay. Plenum Press, New York, pp. 259-294.
Toole, C.L. and R.L. Nielsen, 1992. Effects of microprobe precision of hypotheses related to otolith Sr/Ca ratios. U.S. Fish. Bull., Vol. 90, pp. 421-427.
Townsend, D.W., R.L. Radtke, S. Corwin and A.L. David, 1992. Strontium: calcium ratios in juvenile Atlantic herring Clupeu harengus L. otolith as a function of water temperature. J. Exp. Mar. Biol. Ecol., Vol. 160, pp. 131-140.
Townsend, D.W., R.L. Radtke, D.P. Malone and J.P. Wallinga, 1995. Use of otolith strontium:calcium ratios for hindcasting larval cod Gadus morhua distributions relative to water masses on Georges Bank. Mar. Ecol. Prog. Ser., Vol. 119, pp. 37-44.
Tsukamoto, K., 1989. Otolith daily growth increments in the Japanese eel. Nippon Suisan Gakkaishi, Vol. 55, pp. 1017-1021.
Tsukamoto, K., 1990. Recruitment mechanism of the eel, Anguifla japonica to the Japanese coast. J. Fish Biol., Vol. 36, pp. 659-671.
Tsukamoto, K., 1992. Discovery of the spawning area for Japanese eel. Nature, Vol. 356, pp. 789-791. Tsukamoto, K. and A. Umezawa, 1994. Metamorphosis: a key factor of larval migration determining
geographic distribution and speciation of eels. In, Proceedings of 4th Indo-fuci$c$sh conference, Bangkok, Thailand, 28 Nov.-4 Dec. 1993. pp. 231-248.
Tzeng, W.N., 1990. Relationship between growth rate and age at recruitment of Anguilla juponica elvers in a Taiwan estuary as inferred from otolith growth increments, Mar. Biol., Vol. 107, pp. 75-81.
Tzeng, W.N., 1994. Temperature effects on the incorporation of strontium in otolith of Japanese eel, Anguilla juponica Termminck and Schlegel. J. Fish Biol., Vol. 45, pp. 1055-1066.
Tzeng, W.N. and Y.C. Tsai, 1992. Otolith microstructure and daily age of Anguilla juponica elvers from the estuaries of Taiwan with reference to unit stock and larval migration. J. Fish Biol., Vol. 40, pp. 845-857. Tzeng, W.N., H.F. Wu and H. Wickstrom, 1994. Scanning electron microscopic analysis of annulus
microstructure in otolith of European eel, Anguillu unguilla. J. Fish Biol., Vol. 45, pp. 479-492. Tzeng, W.N. and Y.C. Tsai, 1994. Changes in otolith microchemistry of the Japanese eel, Anguilla juponica,
during its migration from the ocean to the rivers of Taiwan. J. Fish Biol., Vol. 45, pp. 671-684. Utida, S. and T. Hirano, 1973. Effects of changes in environmental salinity on salt and water movement in the
intestine and gills of the eel, Anguilla japonica. In, Responses offish 10 mvironmenral changes, edited by W. Chavin. Thomas, Springfield, pp. 240-269.
Yamada, S.B., T.J. Mulligan and S.J. Fairchild, 1979. Strontium marking of hatchery reared coho salmon