行政院國家科學委員會專題研究計畫 成果報告
利用阿拉伯芥 T-DNA Knockout 突變株 , 研究調控葉部形
態發育之功能性基因(3/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2311-B-002-003- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學植物科學研究所 計畫主持人: 靳宗洛 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 94 年 12 月 25 日
Abstract
Leaves are the most noticeable plant organs and play important roles in photosynthesis, respiration and gas exchange. However, the details of leaf development remain unclear. Therefore, our laboratory used forward genetics to identify the mutants with leaf development defect from T-DNA tagging pools in Arabidopsis. We use a T-DNA tagging vector, pPZP202: BAR: SK, to generate mutants and to screen for morphological mutants. In this study, we have identified a mutant, R1-2, which were selected in the T2 generation, with curled leaves and delayed flowering phenotypes. After R1-2 was crossed to the WT, none of the R1-2 phenotypes was observed in the F1 but were found in the F2 population. Therefore, it was suggested that R1-2 was a recessive mutant. Southern blotting analysis and inversed PCR identified two T-DNA linked together and inserted in the 2.5kb upstream of the At3g22970. Using RT-PCR and Northern blotting assay, we showed that the transcripts of At3g22970 were down-regulated in mutant, implicating that the phenotypes of R1-2 may be resulted from down-regulation of At3g22970.
At3g22970 was predicted as a protein containing DUF506 domain and located in the chloroplast. There are 14 genes that contain DUF506 domain in Arabidopsis thaliana and all with unknown function. Using RT-PCR and Northern blotting assay, it was detected that the expression of At3g22970 appeared rhythmic, and expressed in the whole plant, especially in flowers. However, the function of At3g22970 is still unknown. We observe the phenotype of the transgenic plants by over-expressing or knock-outing the
At3g22970 to address its function in Arabidopsis. Among these
over-expression lines, some lines with delayed flowering phenotype had reduced expression of At3g22970, but others were the same as WT. The correlation between delayed flowering time phenotype and reduced
expression of At3g22970, indicating that the phenotype was caused by co-suppression of the over-expressed At3g22970. Additionally, one of the SALK knock-out lines, SALK-085910, also exhibited a delayed flowering phenotype. Therefore, At3g22970 might play a role to control flowering time.
Introduction
Leaf development
Leaves are the most noticeable plant organs. The early events in leaf development have been divided into three stages, i.e. organogenesis (leaf initiation), setup of the morphological domains of the leaf, and completion of the tissue differentiation. At the organogenesis stage, stage 1, cells on the flank of the shoot apical meristem are set aside as the founder cells of the initiating leaf. The region that increased cell division rates characterize will give rise to the leaf primordium. Stage 2, delimits the basic morphological domains for the growth and development of leaf parts. Cell and tissue differentiation occurs during the final stage in stage 3, which leaf development by coordinated processes of cell division, expansion, and differentiation (Sylvester et al., 1996).
The details of leaf development remain unclear. However, many mutants are isolated with alterations in the leaf morphology, and it can be accumulated information about the role of individual genes in the control of leaf development.
SAM and organogenesis: leaf initiation
The shoot apical meristem (SAM) is a population of cells located at the tip of the shoot axis and self-renewing stem cell populations that generate all of the above-ground organs of higher plants. In Arabidopsis, the SAM is divided into three clonally distinct layers. Cells in the outermost layer (L1) produce epidermal tissues, whereas cells of the sub-epidermal layer (L2) and the internal layers (L3) differentiate into vascular and internal tissues, respectively. Superimposed across these cell layers are distinct zones of differential
meristem activity. A central zone (CZ) at the very apex harbors the unspecialized stem cells, which maintain themselves and also replenish cells in the adjacent peripheral zone (PZ) as they are lost during the formation of lateral organ primordia on the meristem flanks.
Meanwhile, maintenance of a functional meristem between stem cell divisions and cell differentiation requires many genes to regulate. In
Arabidopsis, two genes have been implicated in the maintenance of
undifferentiated cells in the meristem: SHOOT MERISTEMLESS (STM) and
WUSCHEL (WUS), both of which encode homeodomain proteins (Long et al.,
1996; Mayer et al., 1998). In a strong stm mutant, the meristem was absent at the end of embryogenesis (Barton and Poethig, 1993), however, in the wus mutant, the meristem was not established during embryogenesis.
On the other hand, recent reports had shown that keeping the correct number of stem cells is achieved by a negative feedback regulation of WUS-CLV pathway in Arabidopsis, i.e., the WUS mediated signal from the organizing center (OC) that specifies stem cell identity in the outermost layers, which signal back via the CLV pathway to limit the size of the WUS-expressing OC (Fletcher et al., 1999; Schoof et al. 2000). Therefore, clv mutants that failed to repress WUS activity and caused an increase in the size of both shoot and floral meristems (Clark et al., 1993; Clark et al., 1995; Laufs et al., 1998).
CLV and WUS genes are necessary for regulating meristem size, but
class-1 KNOTTED-like homeobox (KNOX) genes, such as SHOOTMERISTEMLESS (STM), BREVIPEDICELLUS (BP, formerly KNAT1), KNAT2, and KNAT6 genes, are required for the maintenance of meristem
identity and be down-regulated at the position of the next primordium (Vollbrecht et al., 1991; Long et al., 1996; Byrne et al., 2002). Mutations at the Class I KNOX gene STM1 led to absence of shoot meristems in Arabidopsis
(Barton and Poethig, 1993).
Establishment of leaf polarity
Plant lateral organs exhibit central-peripheral axis, proximal-distal, and adaxial-abaxial polarity. The central-peripheral axis extends from the apical-basal axis at the center of the organ outward to the organ epidermis. The proximal-distal axis runs from the base of the organ to the tip. The adaxial-abaxial axis runs from the surface of the lateral organ closest to the meristem to the surface of the organ furthest from the meristem.
Genetic studies in Arabidopsis have identified several families of genes that play a role in promoting proper adaxial-abaxial development. The class III HD-Zip genes, in particular of the PHABULOSA (PHB), PHAVOLUTA (PHV), and REVOLUTA (REV) genes promote adaxial identity and meristem maintenance (McConnell and Barton, 1998; McConnell et al., 2001; Emery et al., 2003). The dominant gain-of-function mutants had strongly adaxialized organs and formed trumpet-shaped leaves (McConnell and Barton, 1998). In contrast, mutations in all three genes results in radial abaxial leaves, a phenotype that is complementary to the dominant mutant phenotypes (Emery et al., 2003). The other two family members, ATHB8 and ATHB15, are also likely to direct vascular development, although their precise roles are yet to be uncovered (Baima et al., 2001; Ohashi-Ito and Fukuda, 2003).
The members of the YABBY gene family and KANADI1-3 (KAN1-3) genes promote abaxial identity (Eshed et al., 2001; Sawa et al., 1999; Siegfried et al., 1999). The YABBY genes appear to be plant specific and at least with six family members, including FILAMENTOUS FLOWER (FIL), CRABS CLAW (CRC), and INNER NO OUTER, found in Arabidopsis. The YABBY genes encode a zinc finger and a helix-loop-helix motif and are predicted to be
transcriptional regulators (Siegfried et al., 1999). The fil yab3 double mutants exhibited partial loss of abaxial characteristics (Siegfried et al., 1999; Kumaran et al., 2002). KAN gene encodes a protein that includes a conserved GARP domain, which binds to a specific region of DNA (Kerstetter et al., 2001). The leaves of kan1 kan2 mutants were partially adaxialised, and kan1 kan2 kan3 triple mutants displayed more complete adaxialisation. Genetic studies support that the roles of KAN genes in abaxial specification and YABBY genes in both abaxial fate and lamina outgrowth (Eshed et al., 2004).
The AS1/AS2 and YABBY genes can repress expression of class I KNOX homeobox genes was reported. AS2 (ASYMMETRIC LEAVES2, a
LOB-domain family transcription factor) is predicted to be expressed in the
adaxial organ domain where AS2 forms a complex with AS1 (ASYMMETRIC LEAVES1, a MYB family transcription factor), to promote repression of
KANADI and differentiation of adaxial cell fates (Lin et al., 2003). Therefore,
overexpression of AS2 results in adaxial-abaxial polarity defects, and implicates that AS2 also play a role in polarity establishment.
Cell division and cell expansion
Organ shape is the result of a coordination of cell division and cell expansion. For example, the cincinnata (cin) mutant in Antirrhinum reflected a change in the shape and the progression of a cell-cycle arrest front moving from the leaf tip toward the base. CIN encodes a TCP protein and is expressed downstream of the arrest front. (Nath et al., 2003). In contrast, the
struwwelpeter (swp) mutant had fewer cells number but larger cell size than
WT leaves and did not alter the final size of the organs in Arabidopsis. It has been proposed that organ size might be controlled by a compensation to achieve a more normal morphology between the amount of cells and cell size
(Autran et al., 2002).
Leaf expansion is dependent on both the division and the elongation of cells. The angustifolia (an), a mutant with narrower and thicker leaves than the WT, had altered cortical microtubule arrangements and reduced polar cell expansion in the leaf width dimension in Arabidopsis (Tsuge et al., 1996). AN encodes a CtBP/BARS-related (carboxy-terminal binding protein/brefeldin A ADP-ribosylated substrates-related) protein (Folkers et al., 2002; Kim et al., 2002) and is thought to be a key gene in regulation of the polar elongation of leaf cells in the leaf-width direction specifically (Tsuge et al., 1996). By contrast, the rotundifolia3 (rot3) mutant had shorter and wider leaves, because of defecting in a novel cytochrome P450 gene (Kim et al., 1999). The ROT3 gene appears to be the key gene that regulates the elongation of leaf cells in the leaf-length direction. In the an rot3-1 double mutant had an additive phenotype, suggesting that the two genes act independently (Tsuge et al., 1996).
Hormone and leaf development
Although the actions of plant hormones are not specific to leaves, it is also important to regulate of leaf shape. Hormones like gibberellic acid (GA), auxin and brassinosteroid (BR) appear to be involved in leaf development.
When ectopically expressed, rice and tobacco homeobox genes cause decreased GA levels (Kusaba et al., 1998; Sakamoto et al., 2001). The decreased levels of GA correlate with the severity of leaf shape and plant stature, and the application of exogenous GA could rescue these phenotypes (Kusaba et al., 1998; Hay et al., 2002).
Mutations in the polar auxin transport (PAT) efflux carrier PIN-FORMED1 (PIN1) and treatment of Arabidopsis or tomato with PAT inhibitors both led to a failure to initiate leaves. This phenotype implicates auxin transport in leaf
initiation and possibly as a negative regulator of KNOX gene expression in meristems (Abe et al., 2003).
The phenotype of the mutants that were related to the actions of brassinosteroid (BR) had smaller leaves than the WT in Arabidopsis. It is believed that the main role of BR is to stimulate the elongation of cells (Altmann et al., 1998).
miRNA and leaf devlopment
MicroRNAs (miRNAs) are important controllers of development in plants. miRNAs are 22-nucleotide regulatory RNAs that derive from stem-loop regions of endogenous precursor transcripts (Ambros, 2004; Bartel, 2004). Many miRNAs are known to pair to the messages of protein-coding genes to target these mRNAs for posttranscriptional repression.
For example, two miRNAs from Arabidopsis, miR165 and miR166, match
PHB, PHV and REV mRNA, and direct their cleavage (Emery et al, 2003).
Dominant mutations in PHB/PHV/REV genes all disrupted the miRNA-binding site and transcripts of the dominant mutants could not be cleaved in vitro (McConnell and Barton, 1998; McConnell et al., 2001). Mutations in
ARGONAUTE1 (AGO1), a key component in RNA interference (RNAi)-
mediated posttranscriptional gene regulation, result in adaxialisation of the leaf. The adaxial gene PHB and its miRNA regulator were misregulated in ago1 mutants, demonstrating regulation of the miR165, and consequently the corresponding target, by AGO1 (Kidner and Martienssen, 2004).
A class of TCP genes is also shown to be regulated by MiR-jaw. jaw-D dominant mutants caused overexpression of MiR-jaw, which targets miRNA-binding sites in the TCP transcripts for degradation, resulting in an uneven leaf surface. Therefore, miRNA-dependent regulation of TCP gene
expression may modulate the spatiotemporal characteristics of cell division arrest during leaf development. This regulation is likely to play an important role in determining final leaf shape in Arabidopsis (Palatnik et al., 2003).
Other factors that affect Leaf development
There are many factors that affect leaf development, such as light, water and temperature. Heterophylly is attributable to changes in leaf index because of the environmental adaptation. For example, plants that thrive at the waterside often exhibit heterophylly. Round-shaped and thick leaves with stomata are observed in dry upland conditions, while thin and elongated leaves with no or few stomata are formed under submerged conditions. Such dimorphic changes of leaves are interpreted as a kind of adaptive strategy of these aquatic and semi-aquatic plants to environmental changes, since they have to withstand two different environments of terrestrial conditions and submergence (Kuwabara, 2003).
The existence of two or more leaf types on the same plant is called heteroblastic (Sculthorpe, 1967). For example, Arabidopsis exhibits typical heteroblasty and leaf shape changes from the juvenile, through the early adult, to the late adult phase (Tsuge et al., 1996), with concomitant changes in the patterns of distribution of trichomes on the leaf surfaces (Chien et al., 1996; Telfer et al., 1997). The cauline leaves lack petioles (Tsukaya, 1995; Tsuge et al., 1996). The contours of the first and second true leave resemble those of cotyledons but the leaves have trichomes and a complex vascular system. Thus, the cotyledons might be considered to be as a particular type of foliage leaf and it might be appropriate to examine the cotyledon in the context of heteroblasty.
Transition from vegetative to reproductive phase
Floral initiation is a major developmental transition from vegetative to reproductive phase in plants and it is also regulated by various environmental and physiological cues. However, recent studies have revealed that misexpression of the PI, AP3, AG, and SEP3 genes, caused the curled leaf phenotype was shown in the tfl2 mutants (Kotakeet al., 2003). The curly leaf (clf) mutant also shows a curled leaf phenotype when ectopic expression of the
AG and AP3 was observed (Goodrich et al. 1997). Therefore, ectopic
expression of these genes in the leaves would disrupt the leaf developmet. Models of genetic control of flowering time in Arabidopsis reveal four independent floral promotion pathways: photoperiod, autonomous, gibberellin dependent, and vernalization (Koornneef et al., 1998; Simpson et al., 1999). (1) Photoperiod
Flowering of Arabidopsis is regulated by daylength. Flowering occurs much earlier under long days of 16 hours light than under short days of 8 hours.
CRY2 and PHYA, candidates for photoreceptors that perceive the photoperiod
under long days, entrain the circadian clock to oscillate within a period of 24 h (Mockler et al., 2003). Mutations affecting the photoperiod pathway delay flowering under long-day conditions, but are identical to WT under short-day conditions. For example, CONSTANS (CO) is a key player in the regulation of flowering by photoperiod. WT Arabidopsis plants flower earlier in long days than in short days, whereas co mutants flower late in both short and long days in Arabidopsis (Putterill et al., 1995).
(2) Autonomous
Mutations that affect the autonomous pathway delay flowering under all photoperiods. The autonomous pathway is defined by one group of late flowering mutants, such as fca, fpa, fve, fld, ld, and fy, which flower late under
both long-day and short-day conditions. However, this lateness can be suppressed by either a vernalization treatment or by growth in low red/far red light conditions (Koornneef et al., 1994). The characteristic feature of these mutants is that they all contain much higher levels of FLC transcript than the wild-type plant or late-flowering mutants associated with the photoperiod flowering pathway or the GA pathway (Sheldon et al., 2000).
(3) Gibberellin (GA) dependent
Mutations that strongly reduce GA biosynthesis delay flowering under long-days and almost abolish flowering under short-days. The GA pathway promotes flowering in SDs via a GAMYB-dependent (primarily MYB33) activation of LFY (Blázquez et al., 1998, 2001; Gocal et al., 2001). GA promotes flowering and is absolutely required for flowering in non-inductive short days. Mutations in genes involved in GA biosynthesis and signaling result in delayed flowering. A growing number of genes associated with this pathway have been identified, including SPY, PHOR1, RGLs, RGA, GAI, GA4, GA5, and FPF1 (Mouradov et al., 2002; Olszewski et al., 2002).
(4) Vernalization
An exposure to cold that stimulates flowering is called vernalization. The vernalization pathway promotes flowering in response to low temperature. Many naturally occurring mutants flower very late but flower early if exposed to low temperatures. The vernalization response is mediated by dominant alleles of two genes, FRI and FLC, through reducing the expression of FLC (Michaels et al., 2001). Other genes involved in vernalization response include VRN1 and VRN2 (Gendall et al., 2001), HOS1 (Lee et al., 2001) and VIP1-7 (Zhang et al., 2002).
Arabidopsis thaliana that belongs to the Brassicaceae family is widely
used as a model organism in plant biology. Although not of major agronomic significance, Arabidopsis offers important advantages for basic research in genetics and molecular biology: a small genome size, a rapid life cycle, and easy cultivation in restricted space, prolific seed production, and a large number of mutant stocks that are available for researchers from a variety of stock centers.
In fact, the identification of genes that act as developmental controls in leaf ontogeny is problematic for two reasons. On one hand, the inexistence of analogous phenomena in animals forces us to discard strategies based on gene cloning by homology. On the other hand, two important processes that take place in the leaf, photosynthesis and the exchange of gases with the environment both require the participation of a large number of gene products, primarily expressing in leaves. Therefore, the better experimental approach is forward genetics to screen the mutants that affect leaf morphogenesis. Forward genetics is slow and laborious, but it is also effective and guaranteed to provide the result that is “gene and its function”.
The T-DNA can be considered a type of transposon because it moves from the Ti plasmid to the plant DNA. T-DNA tagging is a method to generate mutations in plants by random insertion of a T-DNA, which can cause knock-out, down-regulation, and over-expression of flanking plant genes. In this study, we use a T-DNA tagging vector, pPZP202: BAR: SK, to generate mutants and screened for morphological mutants. We screened a R1-2 mutant with conspicuously altered phenotypes at leaves and delayed flowering from the T2 T-DNA tagging pool. Gene rescue results showed that the T-DNA inserted in the upstream region of At3g22970, which was annotated to encode an expressed protein, containing DUF506 domain. Besides, the transcripts of
At3g22970 were down-regulation in R1-2. Therefore, we want to find the
relation between the phenotype of the mutant and At3g22970. Using Northern blotting, it was shown that At3g22970 transcript levels are controlled by the circadian clock. In the overexpression lines, with delayed flowering phenotype, the transcripts were reduced compared with the WT. It was indicated that
Materials and Methods
Materials:
Arabidopsis thaliana ecotype Columbia was used for all experiments, and
was grown in an environmental growth chamber under a photoperiod regime of 16h light/8h dark or continuous light. Escherichia coli DH5α and Agrobacterium
tumefaciens strain GV3101 were used in this research.
Methods:
T-DNA tagging screening
The T-DNA tagging lines were generated in a Columbia background with the construct of pPZP202:SK:BAR via floral dipping Agrobacterium-mediated transformation procedure (Clough and Bent, 1998). For mutant screening, a T1 and T2 seeds were incubated at 4°C for 4 days before being sown on the water-soaked soil then transferred to the growth chamber. Herbicide BASTA (Glufosinate ammonium 0.25 g/L) was used to screen the T-DNA tagging lines. We screened for mutants displaying abnormal leaf phenotypes.
Southern blotting analysis
Genomic DNA was isolated from rosette leaves according to Dellaporta (1993). DNA was digested with restriction enzymes, separated on a 0.8% (w/v) agarose gel, transferred to a nylon membrane (Immobilon-Ny+, Millipore) and hybridized at 65°C with 32P -radiolabeled BAR or SK fragment as probe from the pPZP202:SK:BAR T-DNA tagging construct. The membrane was washed and analyzed with the phosphoimgaer.
Total RNA was isolated from the fully expanded leaves using REzol reagent (PROtech Technologies) as described by the manufacturer. 5-10 µg of RNA was electrophoresed on a 1.0% formaldehyde agarose gel and then transferred to a nylon membrane (Immobilon-Ny+) hybridized at 65°C with gene specific 32P-radiolabeled cDNA fragment as probe. The membrane was washed and analyzed with the phosphoimgaer.
IPCR (inverse PCR) identification of the T-DNA flanking sequence
For 1 µg genomic DNA was digested with HindIII or XbaI and purified. 100 ng DNA was used for ligation in a 100 µL reaction at 16°C overnight. 1 µL of the ligation mixture was used for IPCR. The IPCR consisted of 1X PCR buffer, 0.2 mM each dNTP, 0.5 µM each primer, 2.5 units Taq Polymerase. Thermal cycling conditions consisted of an initial denaturation at 95°C for 5 min followed by 30 to 40 cycles of 94°C for 15 sec, annealing for 30 sec, and 72°C for 2.5 min. The PCR products were subjected to nested PCR for increasing the specificity. Primer sets used for IPCR are shown as following:
IPCR Primer sequence Annealing
temperature 35s-pro-out-0 TGCCCTTTGGTCTTCTGAGACTGTATCT 1st Bar-fw-1 GGGAACTGGCATGACGTGGGTTTCT 60°C 35s-pro-out-1 GTGTCGTGCTCCACCATGTTGACCTGCA 2nd 35s-ter-out-1 CCCAAGAATATCCCAAAGCGAGTACACAACT 60°C LB-0 CCGCACCGATCGCCCTTCCCAACAG 3rd 35s-ter-out-1 CCCAAGAATATCCCAAAGCGAGTACACAACT 60°C LB-1 ATTCAGTACATTAAAAACGTCCGCAAT 4th 35s-ter-out-1 CCCAAGAATATCCCAAAGCGAGTACACAACT 60°C
RT-PCR (reverse transcription-PCR)
The cDNA synthesis was performed with 1 µg of total RNA using Ready-to-go RT-PCR kit (Amersham international, Buckinghamshire, England, UK) combined with the oligo dT primer (18 mer). For the 50 µL RT reaction, 1 µL of the reaction was used for further PCR. PCR consisted of 1X PCR buffer, 0.2 mM each dNTP, 0.5µM each primer, 2.5 unites Taq Polymerase. Thermal cycling conditions consisted of an initial denaturation at 94°C for 3 min followed by 24 to 35 cycles of 94°C for 15 sec, annealing for 20 sec, and 72°C for 40 sec. Primer sets used for the RT-PCR are shown as following:
Genes Primer sequence Annealing
temperature Product size Fw GATCTTTGCCGGAAAACAATT GGAGGA TGGT UBQ10 Rv CGACTTGTCATTAGAAAGAAA GAGATAACAGG 60°C 489bp Fw ATGCCATTTACGATGAAGATCC AA At3g22970 Rv TTCATATCGTCTTTTCGTTTCA CC 60 °C 506bp
For RNA loading control, the exponential range of amplification was determined according to the 24 PCR cycles used for UBQ10, and 28 cycles used for At3g22970. 5 out of 25 µL PCR products were analyzed via electrophoresis.
Plasmid constructs
The 35SDE::At3g22970 construction:
stop codon (TAA) by primers:
Primer sequence Annealing
temperature Product size Fw ATGCCATTTACGATGAAGATCCAA Rv GGTACCTTAGGGCTTTTCCTTAAAGAGTGAA 60 °C 1296bp
The PCR amplified fragment was cloned into yT&A vector for sequencing then digested with KpnI then subcloned into the binary vector pBIB-HYG (Becker, 1990). The expression of At3g22970 was drived by dual enhancers of the CaMV 35S promoter.
35SDE:: At3g22970-GFP construct
The At3g22970 cDNA was cloned by RT-PCR using primers are shown as following:
Primer sequence Annealing
temperature Product size Fw TCTAGAATGCCATTTACGATGAAGATCCAA Rv GGATCCAGGGCTTTTCCTTAAAGAGTGAAG 60°C 1111 bp
The RT-PCR cDNA was cloned into yT&A vector for sequencing and digested with XbaI and BamHI to subclone into the 326-GFP vector (acquired from Prof. In-Whan Hwang at Pohang University of Science and Technology).
Plant transformation
Plasmids for plant transformation were transformed into Agrobacterium
tumefaciens GV3101 by electroporation. Agrobacterium cells containing each
plasmid were transformed into the Arabidopsis thaliana ecotype Columbia by the floral dipping method (Clough and Bent, 1998). Transgenic plants were selected by spraying seedlings at 7-, 9-, and 11-d after germination with a
solution of 0.4% BASTA herbicide (McDowell et al., 1998) or were selected by half-strength MS medium (Murashige and Skoog, 1962) containing 25 µg mL-1 hygromycin.
Transient expression into tobacco protoplasts
Transformation of protoplasts with GFP constructs was performed using optimized electroporation parameters. The DNA for transfection was prepared with the PurelinkTM HiPure Plasmid DNA Purification kit (Invitrogen). Protoplasts were transiently transfected with plasmid DNA. Protoplasts were prepared from leaves of tobacco.
The subcellular localization was analyzed using a Leica confocol laser scanning microscope (Leica TCS SP2, Heidelberg, Germany) equipped with the Ar lasers for GFP excitation and the primary beam splitting mirror 488/543/633 was used. The emitted light was detected as following: GFP, 505-520 nm; chlorophyll 650-700nm. Images were processed using Leica confocol software.
Results
1. Characterization of the R1-2 mutant
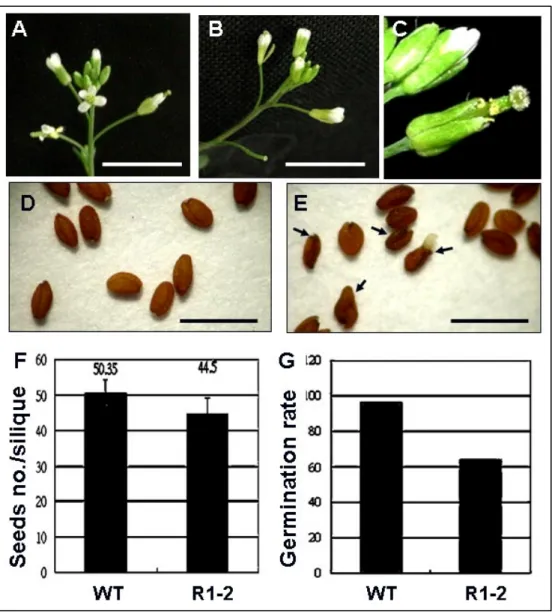
In a large-scale screening for Arabidopsis mutants displaying abnormally shaped leaves, a mutant named “R1-2”, which with curled leaves and delayed flowering phenotypes was obtained (Fig.1B-E). Compared with the wild type (WT), the floral organs of R1-2 were normal in shape but some with shorter stamens (Fig. 2A-C), with fewer seeds in siliques (Fig. 2E), and with reduced germination rate (64%) (Fig. 2F).
Under a long-day (LD, 16h light/8h dark) condition, the vegetative growth of R1-2 mutant was severely retarded, it took about 35 days to flowering compared with a 25 days needed for the WT. Consequently, plant size of the mutant was smaller at the early growth stage, but became comparable to that of the WT reaching/during to the flowering time (Fig.1C, D).
R1-2 showed a more obvious curled leaf phenotype under a continuous light than in a LD growth condition. During the continuous light, the rosette and cauline leaves displayed a more enhanced leaf margin serration (Fig.1D and Fig.2D).
To analyze the inheritance of the R1-2 mutant phenotype, R1-2 was crossed to the WT. None of the R1-2 phenotypes was observed in the F1 but were found in the F2 population (Fig.1C- E). Among thirty-six of the F2 seedlings, there were thirteen seedlings selected by herbicide BASTA. Twenty-three seedlings could survive after BASTA treatment, but only four seedlings showed R1-2 mutant phenotypes (homozygote). Therefore, it was suggested R1-2 was a recessive mutant.
2. Characterization and Isolation of the flanking sequence of T-DNA insertion by using Southern blotting and IPCR
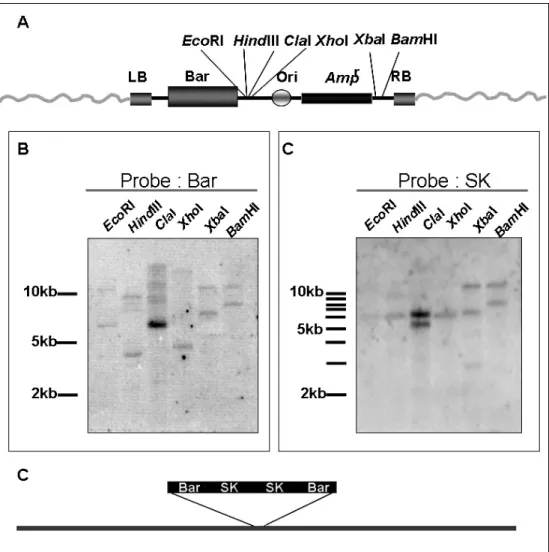
Different restriction enzymes (ex., EcoRI, HindIII, ClaI, XbaI, XhoI, and
BamHI) were used for R1-2 genomic DNA digestion and Southern blotting
analysis using the BAR or SK fragments of T-DNA as a probe (Fig. 3A). The Southern blotting results was shown that when probe with the BAR indicated there were two T-DNA insertions (Fig. 3B). However, using SK+ fragment (3 kb) as a probe, only one 6-kb hybridization signal was detected by EcoRI, HindIII and XbaI digestion (Fig. 3C). Therefore, the result revealed the two T-DNA was resulted in a right border (RB) to RB-linkage T-DNA. According to the results, we proposed that the model of two T-DNA inserted at same location in the R1-2 mutant (Fig. 3D).
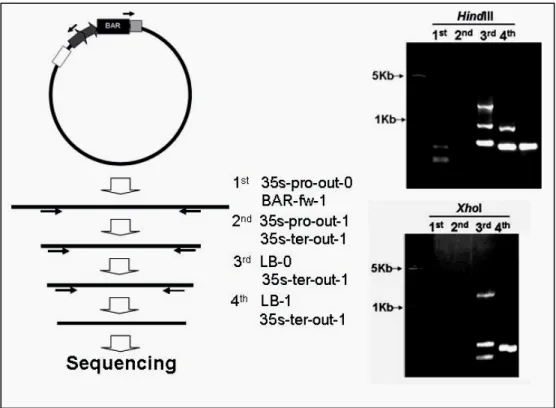
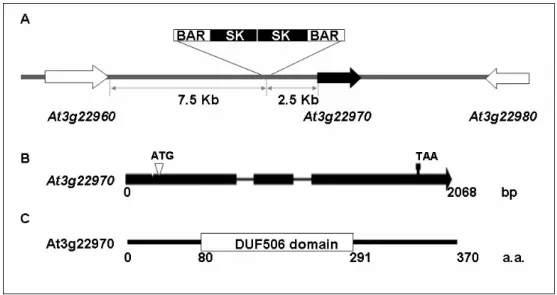
Southern results show that the HindIII and XbaI digestion with a smaller hybridization signals and these T-DNA flanking sequence could be rescued by inverse PCR (IPCR) (Fig. 4). When to sequence the 4th-nested IPCR/PCR product and analyzed by BLAST program of TAIR (http://www.arabidopsis.org/Blast/ index.jsp), showed that the T-DNA was tagged in a 2.5-Kb upstream of the At3g22970 gene (Fig. 5A).
3. Molecular characterization of the At3g22970 gene and DUF506 gene family
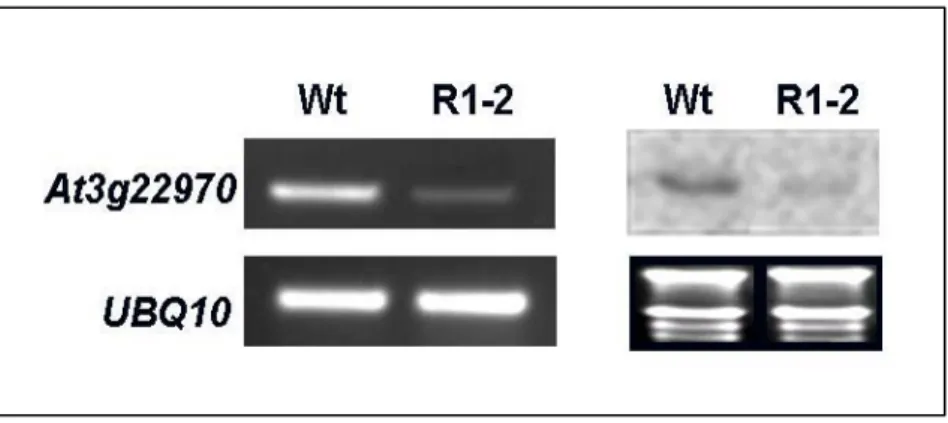
The T-DNA insertion site apart from the At3g22960 and At3g22980 gene were 7.5-kb and 11.2-Kb, respectively (Fig. 5A). Due to the insertion distance were greater than 5 kb, we therefore suggested that the interfered At3g22970 gene may cause to the R1-2 phenotype. The RT-PCR was used to analyze the expression of the At3g22970 in the WT and R1-2. In Figure 6, the result revealed that the At3g22970 was down-regulated in R1-2. Therefore, we
suggested that phenotypes of the R1-2 might be resulted in a down-regulation of the At3g22970 expression.
In TAIR, the genomic sequences of At3g22970 was proposed to be 2065 bp, consists three exons and two introns (Fig. 5B), encoding a protein of 370 amino acids. It is a protein containing a DUF506 domain and to be translocated into the chloroplast (Fig. 5C and Fig. 7). The putative chloroplast transit peptide was predicted by the TargetP program (http://www.cbs. dtu.dk/services/TargetP/) based on its N-terminal amino acid sequence of the
At3g22970 protein.
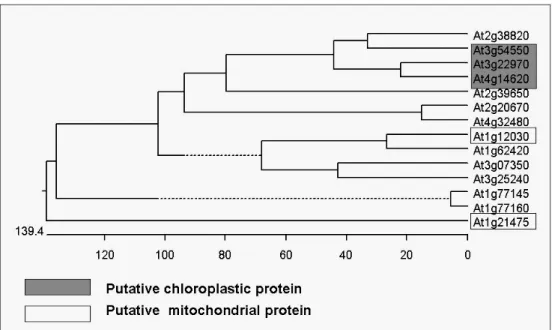
Searching on the database also revealed that the At3g22970 belongs to a gene family which containing a DUF506 domain, with 14 members in
Arabidopsis, and all shown with unknown function (Fig. 7). The DUF506
domain was defined by a domain found toward the C-terminus in a number of the uncharacterized plant proteins. The domain is strongly conserved (more than 30% sequence identity between most pairs of members), but flanked by a highly divergent regions including stretches of low-complexity sequence. The At3g22970 shared with the highest homology to the At4g14620 (52.5% amino acid identity).
4. Subcellular localization of the At3g22970 protein
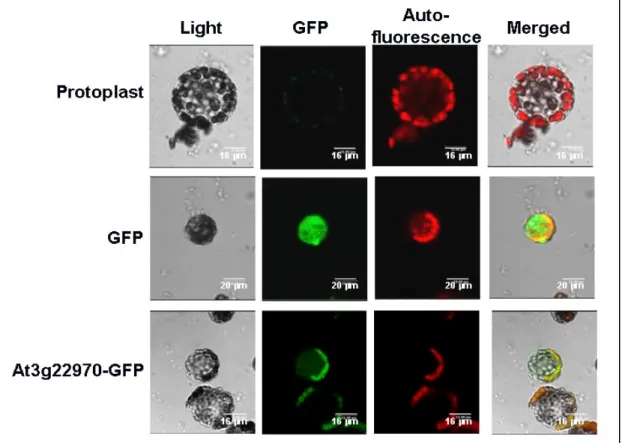
To confirm that the At3g22970 protein is translocated into the chloroplast, one GFP-coding sequence was fused to the 3’-end of At3g22970 coding sequence by using the p326-GFP vector. This p326-At3g22970-GFP vector was transiently expressed in the tobacco protoplasts. The recombinant GFP and the chlorophyll auto-florescent signals were colocalized in chloroplasts, as shown in the Fig. 8. which confirmed that the At3g22970 protein was translocated into the chloroplast.
5. Expression of the At3g22970 displays a rhythm with the light/dark cycles
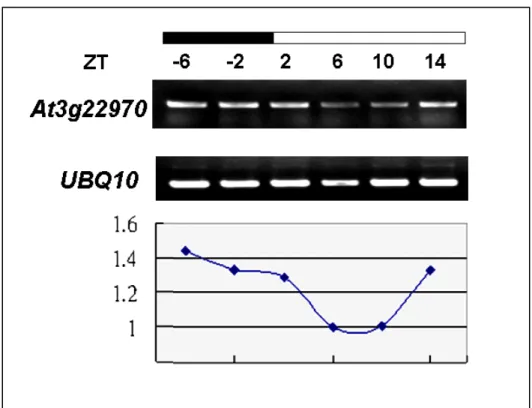
As shown in Fig. 9, one cis-acting regulatory element (CAAAtatatc) involved in the circadian control at 1829 bp upstream of the promoter region in the At3g22970 was predicted by the PlantCARE (http://oberon.fvms. ugent.be:8080/PlantCARE/index.html) and was also shown to have a circadian rhythm controlled in the microarray data-base in TAIR (http://www. arabidopsis.org/ servlets/TairObject?id=37402&type=locus). For these reasons, whether the At3g22970 transcript was fluctuated during the day-night cycle was examined by RT-PCR and Northern blotting. The young seedling was collected every 4 to 6h over a 24h or 48h period in a LD condition and used for RT-PCR and Northern blotting analysis. Sampling time was shown in hours as the Zeitgeber Time (ZT, Zerr et al., 1990), which is the number of hours after dawn at 6:00 Am (i.e., ZT-0). RT-PCR assay indicated that the At3g22970 transcript could be changed with time for 24h (Fig.10). Thus, Northern blotting was used to confirm that the change was rhythmic. The result revealed that the
At3g22970 transcript, indeed, has a cycled expression paten, with the highest
level at ZT -6h and ZT 18h, and the lowest level was at ZT 12 and ZT 36 (Fig.11).
6. Tissue-specific expression of the At3g22970
To correlate the function of At3g22970 genes with the abnormal leaf phenotype and flowering time, RT-PCR was conducted to analysis the tissue-specific expression patterns. Because the At3g22970 was shown to be expressed at the highest level at ZT 18 in a LD condition (Fig. 10 and Fig. 11), so the WT samples were harvested at ZT 18h. In Fig. 12, the At3g22970 transcripts could be detected in all of the tested tissues, ex. in the roots,
mature leaves, stems, flowers, and siliques, however, the highest expressed tissue was observed in the flowers.
7. Phenotypes of the At3g22970 T-DNA knock-out lines
The loss-of-function mutants were obtained from The Salk Institute Genome Analysis Laboratory (SIGnAL) in a T-DNA insertion loss/reduction- of-function pool (Sessions et al., 2002; Alonso et al., 2003), and three T-DNA insertion alleles of the At3g22970 (SALK-022456, SALK-085849 and SALK-085910) were analyzed (Fig.13A). Only one of the lines, SALK-085910, exhibited a delay-flowering phenotype, however, the leaves shown the same as the WT (Fig.13B). SALK-085910 is a heterozygote line and Northern blotting assay showed that expression of the At3g22970 in the T-DNA insertion line (SALK-085910) was less than WT but more than that in R1-2 (Fig.13C).
8. Over-expression of the At3g22970 in WT and R1-2
To further understand the functions of the At3g22970, the genomic clone was over-driven with the dual enhancers of CaMV 35S promoter. The 35SDE::At3g22970 construct was transformed into WT. For all of the 16 T1 independent lines, the leaves showed no without difference as these in WT. However, 5 out of the 16 lines showed the delayed flowering phenotype (Fig.14A). The At3g22970 expression was analyzed in these lines at ZT 18h by Northern blotting analysis and revealed that the At3g22970 was down-regulated as that of the R1-2 (Fig.14B), suggesting that the down-regulation of the At3g22970 expression in the transgene lines might result from co-suppression of both At3g22970 endogenous genes and transgene. The leaf phenotype of these transgene lines were not different from WT at 30 days, but these had weak margin serration at 40 days. Therefore, We
suggest that the phenotype of leaves may be caused by dosage-dependent effect.
On the other hand, when introduced the 35SDE::At3g22970 construct into the R1-2, only 3 independent T1 lines were obtained. Two of these T2 lines, grew very slowly than that of in the WT and R1-2 (Fig.15).
Discussion
1. Down-regulation of the At3g22970 in R1-2
In R1-2, T-DNA was inserted in 2.5 Kb upstream of the At3g22970 gene. Promoter was predicted by using Promoter Scan software (http://bimas.dcrt. nih.gov/molbio/proscan), that promoter region was covered from -1712 bp to -1962 bp and that TATA box was predicted by the PlantCARE (http://oberon.fvms. ugent.be:8080/PlantCARE/index.html) at 32 bp upstream of the transcription start site. Therefore, one possible reason is presumed that the T-DNA might affect the enhancer of At3g22970, a cis-element which highly increased the expression of At3g22970. It is reported that the enhancer might locate upstream or downstream of a gene with hundreds to thousands of base pairs away for effecting.
Second, from previous results, when introduced the 35SDE::At3g22970 into WT plant, five of sixteen transgenic plants showed delayed flowering phenotype and decreased the expression of At3g22970, indicated that the phenotype was caused by co-suppression of At3g22970 expression (Denli and Hannon, 2003; Szweykowska-Kulinska et al., 2003). Therefore, the phenotype may be also caused by co-suppression of At3g22970 expression in R1-2. Because two linked T-DNA had four 35S enhancers, the neighbor genes,
At3g22970, may be over-expressed and result in the down-regulation of At3g22970 in R1-2.
Third, recent studies have revealed that DNA (de)methylation and histone methylation are involved in the control of gene expression during development and differentiation (Bird, 1992 ; Martienssen et al., 1995). For example, the floral repressor gene FLOWERING LOCUS C (FLC) in Arabidopsis acquires methylation at H3K27 and H3K9 during an extended period of cold treatment
for floral induction (Bastow et al. 2004; Sung et al., 2004). In microarry data from the TAIR, the young seedlings were treated with 5-AC, a demethylating agent, and the At3g22970 could be induced after treatment. Thus, T-DNA may disrupt demethylation in R1-2 and then affect the At3g22970 expression.
Forth, T-DNA may affect the transcription factor binding to the promoter of
At3g22970, such as the circadian regulatory element, to disrupt the rhythmic
cycling of the At3g22970 transcription.
2. At3g22970 may play a role in seed development
The segregation ratio of F2 generation was not 3:1 (normal: abnormal), because the homozygote lines were less then expectation. It may be the reason that the R1-2 only produced 88% of seeds in one silique compared with that of WT(Fig. 2F). Among these seeds, many seeds were aborted and only 64% germinated (Fig. 2D, E). Thus, this might be the reason that only four homozygote seedlings were found among 36 of the F2 seedlings, suggested that the At3g22970 may play a role in seed development.
3. At3g22970 expression in wild-type plants and photoperiodic flowering
CCA1 (CIRCADIAN CLOCK ASSOCIATED 1) is an unusual DNA binding
protein in that it recognizes an asymmetric DNA sequence (the CCA1-binding site AAAAATCT or the evening element AAATATCT) and has only one Myb-like domain for DNA binding. Meanwhile, in the At3g22970 there are three CCA1-binding sites (AAAAATCT) in upstream of 1859 bp, 1798bp, and 498 bp in it promoter region. Analysis of the At3g22970 promoter by PlantCARE, it also revealed at -1829 bp also with a cis-acting regulatory element, CAAAtatatc, involved in the circadian control. Therefore, At3g22970 might be regulated by CCA1 and play a role in the circadian rhythm.
Previous physiological experiments implicated the circadian clock in photoperiodic time measurement, and genetic experiments in Arabidopsis have also identified mutants that disrupt both circadian clock-controlled responses and photoperiodic responses (Koornneef et al., 1998). The
At3g22970 gene may be involved in the flowering photoperiodic response,
because the flowering time in R1-2 mutants is delayed under long days (LD) condition. However, whether the At3g22970 expression patterns were different between plant growth under the LD and short days (SD, 12 light/12 dark) remain unclear. Therefore, to understand the relationship between circadian clock and the At3g22970 expression should be proceed by Northern analysis or RT-PCR in the WT and R1-2 under LD, SD, continuous light shift to dark period, and continuous dark shift to light period.
4. The leaves phenotype of R1-2, knock-out line (SALK-085910) and over-expression lines
It remain unclear which role AT3g22970 does play in leaf development. However, recent studies have revealed that many mutants with altered flowering time also disrupt the leaf development. For example, PIE1, an ISWI family gene, is required for FLC activation and floral repression in Arabidopsis. In the Ws genetic background, pie1-1 displays, in addition to early flowering, leaves that are slightly narrower and more serrated at the base (Noh et al., 2003).
We also find that the leaf phenotype of R1-2 is significant under constant light and high light intensity condition. Therefore, the leaf phenotype can be observed in the WT, R1-2, SALK_085910 and over-expression lines under LD, SD and LL and to find the relationship among the leaf phenotype, At3g22970 and light.
Photoperiod and temperature can alter the levels of active GAs by affecting specific steps in the biosynthetic pathway. Current evidence suggests that reduced activity of the GAs growth regulator pathway promotes meristematic activity, both in the natural context of KNOX function in the SAM and upon ectopic KNOX expression in Arabidopsis leaves (Hay et al., 2002). On the other hand, mutations that strongly reduce GA biosynthesis delay flowering under long-days and almost abolish flowering under short-days. Therefore, At3g22970 may affect the GAs biosynthetic pathway and result in curled leaves and delayed flowering phenotypes. We can use RT-PCR assay to study with GA synthesis gene in WT and R1-2 .
5. At3g22970 may play a role to control flowering time
When introduced the 35SDE::At3g22970 into WT plant, five of sixteen transgenic plants showed delayed flowering phenotype. Using Northern blotting analysis, the At3g22970 was down-regulated as seen in the R1-2. The correlation between delay flowering phenotype and reduced expression of
At3g22970 indicated that the phenotype was caused by co-suppression of At3g22970 expression (Denli and Hannon, 2003; Szweykowska-Kulinska et al.,
2003).
By RT-PCR assay, we also detected the expression of gene controlling flowering time, such as GI, CO, FT, LD, SOC1 and AP1. The results showed that CO and LD were down-regulated in R1-2 (data not show). The CO (CONSTANS) gene is found to encode a protein with similarity to GATA-1 type transcription factors and mediates between the circadian clock and the control of flowering in Arabidopsis (Putterill et al., 1995; Paula et al., 2001). The LD (luminidependens) gene encodes a glutamine-rich nuclear protein containing a possible homeodomain and promotes flowering by repressing FLC expression
(Lee et al., 1994; Sheldon et al., 2002). Recent studies have revealed that the
co and ld mutants are both late flowering under LD (Koornneef et al., 1991). To
understand the relationship between these gene and At3g22970, the expression of gene controlling flowering time should be detected by RT-PCR assay in WT, R1-2, SALK_085910 and over-expression lines with delayed flowering time phenotype under LD, SD or continuous light condition.
From previous results, At3g22970 may play a role to control flowering time.
Figure 1. The phenotype of R1-2
(A) WT and (B) R1-2 plants at 35 days in growth of LD (long day). Scale bar: 1 cm.
(C) In a T2 population, plants revealed the curled leaves and delayed the flowering time phenotype. A homozygote (left) and heterozygote (right) plants at 20-day-old under continuous light condition. Scale bar: 1 cm. (D) R1-2 plants at 30-day-old in growth under continuous light. Scale bar: 1
cm.
(E) The cauline leaves of R1-2 and WT at 30-day-old. The mutant with enhanced leaf margin serration compared with Wild-type under continuous light. Scale bar: 1 cm.
(F) A homozygote (left, arrow), heterozygote (mid), and WT (right) at 20-day-old under continuous light condition. Scale bar: 5 cm.
Figure 2. The phenotype of R1-2
The flowers in (A) WT and (B) R1-2. Scale bar: 1 cm. (C) R1-2 flowers with short stamens
The seeds in (D) WT and (E) R1-2. There are more abortive seeds (arrows) in R1-2 than WT. Scale bar: 1 mm.
(F) Seeds number in siliques of R1-2 and Wild-type. It was shown that the silique had fewer seeds in R1-2 than WT.
(G) A graph displaying percentage of seeds that could germinate. It was shown that only 64% of R1-2 seeds could germinate.
Figure 3. Southern blotting to analyze the T-DNA insertion numbers (A) The restriction enzyme site in the T-DNA, including EcoRI, HindIII, ClaI,
XhoI, XbaI, and BamHI.
Southern blotting analysis by BAR (B) or SK (C) fragments as a probe, respectively; shows that there were two T-DNA were linked together inserted in the R1-2.
(D) The model of two T-DNA linked together with SK fragments inserted in the DNA sequence in R1-2 mutant
Figure 4. IPCR to clone the T-DNA flanking sequence
Using different nested-PCR primers, the final T-DNA flanking sequence were amplified and sequenced.
Figure 5. Molecular cloning of the At3g22970
(A) In R1-2, the T-DNA is inserted between genes At3g22960 and At3g22970 (B) Schematic of At3g22970 gene. The genomic length is 2068 bp. There is
three exons (boxed) and two introns (lined) in At3g22970
(C) Schematic of At3g22970 protein. At3g22970 gene encodes a protein of 370 amino acids and contains the DUF506 domain from 80 a.a. to 291
Figure 6. The expression of At3g22970 in Wild-type and R1-2
RT-PCR and Northern blotting assay were performed on RNA from rosette leaves. The result was shown that the At3g22970 was down-regulated in the R1-2. The samples were collected at ZT 18h. UBQ10 was used as a control.
Figure 7. Phylogenetic analysis of genes with the DUF506 domain in
Arabidopsis.
A phylogenetic tree of the gene with DUF506 domain based on amino acid sequences. The plot was obtained by the Cluster Method of the MegAlign program (DNAstar, Madison, WI).
Figure 8. Confocal imaging for the colocalization of the chloroplast auto-fluorescence and recombinant At3g22970- GFP
Confocal fluorescence images of (A) protoplast, (B) transient expressing of p326-GFP, and (C) p326-At3g22970-GFP. From the left to the right, a bright-field, GFP fluorescence image, chlorophyll auto-fluorescence, and the merged confocal images, respectively. Bars represent 16 µm in (A) and (C) and 20µ m in (B).
Figure 9. The cis-elements related to the circadian in 3 kb upstream of
Figure 10. RT-PCR analysis of the At3g22970 gene expression during one day interval.
Seedlings of wild-type was collected every 4 h from ZT -6h to ZT 14h were used for RT-PCR analysis. UBQ10 was used as loading control. Results are presented as a proportion after normalization with UBQ10 signal.
Figure 11. Northern blotting analysis of the At3g22970 gene expression with a circadian rhythm
Seedlings of wild-type was collected every 6 h from ZT -6h to ZT 36h were used for Northern blotting analysis. Results are presented as proportion after normalization with RNA stained gel.
Figure 12. Tissue-specific expression of the At3g22970
RT-PCR was performed to analyze the At3g22970 gene expression in the roots (R), stems (St), rosette leaves (RL), cauline leaves (CL), flowers (F) and siliques (Si). The samples were collected at ZT 18h. UBQ10 was used as loading control.
Figure 13. The phenotypes of the At3g22970 T-DNA knock-out line (A) The T-DNA insertion site of SALK knock-out line
(B) SALK-085910, a At3g22970 T-DNA knock-out line, exhibited a delayed- flowering phenotype but with normal leaves phenotype as the WT.
(C) Northern blotting analysis the expression of At3g22970 in WT, R1-2 and the SALK- 085910 line. The samples were collected at ZT 18h.
Figure 14. The phenotype of over-expression At3g22970 in WT
(A) WT, R1-2, and the line with At3g22970 over-expressed in WT were growth in LD for 30 days. Scale bar: 1 cm.
(B) Northern blotting analysis the expression of At3g22970 in WT, R1-2, and the At3g22970 over-expressed in WT. The samples were collected at ZT 18h.
(C) The rosette leaf phenotype of WT, R1-2 and over-expressed lines at 40 days. Shown are the fifth rosette leaves. Scale bar: 1 cm.
Figure 15. The phenotype of over-expression At3g22970 in R1-2
WT, R1-2, the line with At3g22970 over-expressed in R1-2 grew in LD for 30 days. Scale bar: 1 cm.
Reference
Abe, M., Katsumata, H., Komeda, Y., and Takahashi, T. (2003). Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development 130, 635-643.
Altmann, T. (1998). Recent advances in brassinosteroid molecular genetics: Curr. Opin. Plant Biol. 1, 378–383.
Ambros, V. (2004). The functions of animal microRNAs. Nature 431, 350-355. Autran, D., Jonak, C., Belcram, K., Beemster, G.T., Kronenberger, J.,
Grandjean, O., Inze, D., and Traas, J. (2002). Cell numbers and leaf development in Arabidopsis: a functional analysis of the
STRUWWELPETER gene. EMBO J. 21, 6036-49.
Baima, S., Possenti, M., Matteucci, A., Wisman, E., Altamura, M.M., Ruberti, I., and Morelli, G. (2001). The Arabidopsis ATHB-8 HD-ZIP protein acts as a differentiation-promoting transcription factor of the vascular meristems. Plant Physiol. 126, 643–655.
Bartel, D.P. (2004). MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116, 281-297.
Barton, M.K., and Poethig, R.S. (1993). Formation of the shoot apical meristem in Arabidopsis thaliana: An analysis of development in the wild type and in the shoot meristemless mutant. Development 119, 823-831. Bastow, R., Mylne, J.S., Lister, C., Lippman, Z., Martienssen, R.A., and
Dean, C.. (2004). Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 427, 164–167.
Baum, S.F., and Bowman, J.L. (2003). Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 13, 1768–1774. Blázquez, M.A., Soowal, L.N., Lee, I., and Weigel, D. (1997). LEAFY expression and flower initiation in Arabidopsis. Development 124, 3835–3844.
Blázquez, M., Koornneef, M., and Putterill, J. (2001). Flowering on time: genes that regulate the floral transition. Workshop on the molecular basis of flowering time control. EMBO Rep. 2, 1078–1082.
Byrne, M.E., Barley, R., Curtis, M., Arroyo, J.M., Dunham, M., Hudson, A., and Martienssen, R.A. (2000). Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 408, 967-971. Byrne. M.E., Simorowski, J., and Martienssen, R.A. (2002). ASYMMETRIC
129, 1957-65.
Chien, J. C., and SUSSEX, I. M. (1996). Differential regulation of trichome formation on the abaxial and adaxial leaf surfaces by gibberellins and photoperiod in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 111,1321-1328.
Clark, S.E., Runnung, M.P., and Meyerowitz, E.M. (1993). CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development 119, 397-418.
Clark, S.E., Runnung, M.P., and Meyerowitz, E.M. (1995). CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same process as CLAVATA1. Development 121, 2057-2067.
Clough, S.J., and Bent, A.F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16, 735-43.
Denli, A.M., and Hannon, G.J. (2003). RNAi: an ever-growing puzzle. Trends Biochem. Sci. 28, 196 201.
Emery, J.F., Floyd, S.K., Alvarez, J., Eshed, Y., Hawker, N.P., Izhaki, A., Baum, S.F., and Bowman, J.L. (2003). Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr Biol. 13, 1768-1774. Eshed, Y., Baum, S.F., Perea, J.V., and Bowman, J.L. (2001). Establishmentof polarity in lateral organs of plants. Curr. Biol. 11, 1251-1260.
Eshed, Y., Izhaki, A., Baum, S.F., Floyd, S.K., and Bowman, J.L. (2004). Asymmetric leaf development and blade expansion in Arabidopsis are mediated by KANADI and YABBY activities. Development 131, 2997 3006.
FIetcher, J.C., Brand, U., Running, M.P., Simon, R., and Meyerowitz, E.M. (1999). Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283, 1911-1914.
Folkers, U., Kirik, V., Schobinger, U., Falk, S., Krishnakumar, S., Pollock, M.A., Oppenheimer, D.G., Day, I., Reddy, A.S., and Jürgens ,G. (2002). The cell morphogenesis gene ANGUSTIFOLIA encodes a CtBP/BARS-like protein and is involved in the control of the microtubule cytoskeleton. EMBO J. 21, 1280-1288.
Gendall, A.R., Levy, Y.Y., Wilson, A., and Dean, C. (2001). The
VERNALIZATION 2 gene mediates the epigenetic regulation of
vernalization in Arabidopsis. Cell 107, 525-535.
and Weigel, D. (2001). Evolution of floral meristem identity genes. Analysis of Lolium temulentum genes related to APETALA1 and LEAFY of
Arabidopsis. Plant Physiol. 125, 1788–1801.
Grandjean, O., Inzé, D., and Traas, J. (2002). Cell numbers and leaf development in Arabidopsis: a functional analysis of the
STRUWWELPETER gene. EMBO J. 21, 6036-6049.
Gaudin, V., Libault, M., Pouteau, S., Juul, T., Zhao, G., Lefebvre, D., and Grandjean, O. (2001). Mutations in LIKE HETEROCHROMATIN
PROTEIN 1 affect flowering time and plant architecture in Arabidopsis.
Development 128, 4847–4858.
Hay, A., Kaur, H., Phillips, A., Hedden, P., Hake, S., and Tsiantis, M. (2002). The gibberellin pathway mediates KNOTTED1-type homeobox function in plants with different body plans. Current Biology 12, 1557–1565.
Laufs, P., Grandjean, O., Jonak, C., Kiêu, K., and Traas, J. (1998). Cellular parameters of the shoot apical meristem in Arabidopsis. The Plant Cell 10, 1375-1389.
Lee, H., Xiong, L., Gong, Z., Ishitani, M., Stevenson, B., and Zhu, J.K. (2001). The Arabidopsis HOS1 gene negatively regulates cold signal transduction and encodes a RING finger protein that displays cold-regulated nucleo—cytoplasmic partitioning. Genes Dev. 15, 912-924. Lee, I., Aukerman, M.J., Gore, S.L., Lohman K.N., and Michaels, S.D.
(1994). Isolation of LUMINIDEPENDENS: a gene involved in the control of flowering time in Arabidopsis. Plant Cell 6, 75 83.
Lin, W., Shuai, B., and Springer, P.S. (2003). The Arabidopsis LATERAL
ORGAN BOUNDARIES-domain gene ASYMMETRIC LEAVES2 functions
in the repression of KNOX gene expression and in adaxial-abaxial patterning. Plant Cell 15, 2241–2252.
Long, J.A., Moan, E.I., Medford, J.I., and Barton, M.K. (1996). A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 379, 66-69.
Kerstetter, R.A., Bollman, K., Taylor, R.A., Bomblies, K., and Poethig, R.S. (2001). KANADI regulates organ polarity in Arabidopsis. Nature 411, 706-709.
Kidner, C.A., and Martienssen, R.A. (2004). Spatially restricted miRNA directs leaf polarity through ARGONAUTE1. Nature 428, 81-84.
Kim, G.T., Tsukaya, H., Saito, Y., and Uchimiya, H. (1999). Changes in the shapes of leaves and flowers upon overexpression of cytochrome P450 in Arabidopsis. Proc. Natl. Acad. Sci. USA 96, 9433-9437.
Kim, G.T., Shoda, K., Tsuge, T., Cho, K.H., Uchimiya, H., Yokoyama, R., Nishitani, K., and Tsukaya, H. (2002). The ANGUSTIFOLIA gene of
Arabidopsis, a plant CtBP gene, regulates leaf-cell expansion, the
arrangement of cortical microtubules in leaf cells and expression of a gene involved in cell-wall formation. EMBO J. 21, 1267-1279.
Koornneef, M., Hanhart, C.J., and van der Veen J.H. (1991). A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana.
Mol. Gen. Genet. 229, 57-66
Koornneef, M., H. Blankestijn-de Vries, Hanhart, C., Soppe, W., and Peeters, T. (1994). The phenotype of some late-flowering mutants is enhanced by a locus on chromosome 5 that is not effective in the
Landsberg erecta wild-type. Plant J. 6, 911-919.
Koornneef, M., Alonso-Blanco, C., Peeters, A.J.M., and Soppe, W. (1998). Genetic control of flowering time in Arabidopsis. Annu. Rev. Plant Physiol. 49, 345–370
Kotake, T., Takada, S., Nakahigashi, K., Ohto, M., and Goto, K. (2003).
Arabidopsis TERMINAL FLOWER 2 gene encodes a heterochromatin
protein 1 homolog and represses both FLOWERING LOCUS T to regulate flowering time and several floral homeotic genes. Plant Cell Physiol. 44, 555–564
Kumaran, M.K., Bowman, J.L., and Sundaresan, V. (2002). YABBY polarity genes mediate the repression of KNOX homeobox genes in Arabidopsis. Plant Cell 14, 2761–2770.
Kusaba, S., Fukumoto, M., Honda, C., Yamaguchi, I., Sakamoto, T., and Kano-Murakami, Y. (1998). Decreased GA1 content caused by the overexpression of OSH1 is accompanied by suppression of GA 20-oxidase gene expression. Plant Physiol. 117, 1179-84.
Kusaba, S., Kano-Murakami, Y., Matsuoka, M., Tamaoki, M., Sakamoto, T., Kerstetter, R.A., Bollman, K., Taylor, A., Bomblies, K., and Poethig, S. (2001). KANADI regulates organ polarity in Arabidopsis. Nature 411, 706-709.
Kuwabara, A., H. Tsukaya., and T. Nagaya. (2001). Identification of factors that cause heterophylly in Ludwigia arcuata Walt. (Onagraceae). Plant Biology 3, 98-105.
Kuwabara, A., Ikegami, K., Koshiba, T., and Nagata, T. (2003). Effects of ethylene and abscisic acid upon heterophylly in Ludwigia arcuata (Onagraceae). Planta 217, 880-887.
(1998). Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell. 6, 805-815.
McConnell, J.R., and Barton, M.K. (1998). Leaf polarity and meristem formation in Arabidopsis. Development 125, 2935-2942.
McConnell, J.R., Emery, J., Eshed, Y., Bao, N., Bowman, J., and Barton, M.K. (2001). Role of PHABULOSA and PHAVOLUTA in determining radial patterning in shoots. Nature 411, 709-713.
Michaels, S.D., and Amasino, R.M. (2001). Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. Plant Cell 13, 935-941.
Mockler, T., Yang, H., Yu, X., Parikh, D., Cheng, Y.C., Dolan, S., and Lin, C. (2003). Regulation of photoperiodic flowering by Arabidopsis photoreceptors. Proc. Natl. Acad. Sci. USA 100, 2140-2145.
Mouradov, A., Cremer, F., and Coupland, G. (2002). Control of flowering time: interacting pathways as a basis for diversity. Plant Cell 14, S111-130. Nath, U., Crawford, B.C., Carpenter, R., and Coen, E. (2003). Genetic
control of surface curvature. Science 299, 1404-1407.
Noh, Y.S., Amasino, R.M. (2003). PIE1, an ISWI family gene, is required for FLC activation and floral repression in Arabidopsis. Plant Cell. 15, 1671-82.
Ohashi-Ito, K., and Fukuda, H. (2003). HD-zip III homeobox genes that include a novel member, ZeHB-13 (Zinnia)/ATHB-15 (Arabidopsis), are involved in procambium and xylem cell differentiation. Plant Cell Physiol. 44, 1350-1358.
Olszewski, N, Sun, T.P., and Gubler, F. (2002). Gibberellin signaling: biosynthesis, catabolism, and response pathways. Plant Cell 14, S61-80. Palatnik, J.F., Allen, E., Wu, X., Schommer, C., Schwab, R., Carrington,
J.C., and Weigel, D. (2003). Control of leaf morphogenesis by microRNAs. Nature 425, 257–263.
Paula Suárez-López, Wheatley, K., Robson, F., Onouchi, H., Valverde, F. and Coupland, G. (2001) CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 410, 1116-1120. Putterill, J., Robson, F., Lee, K., Simon, R., and Coupland, G. (1995). The
CONSTANS gene of Arabidopsis promotes flowering and encodes a
protein showing similarities to zinc finger transcription factors. Cell 80, 847-857
M. (2001). KNOX homeodomain protein directly suppresses the expression of a gibberellin biosynthetic gene in the tobacco shoot apical meristem. Genes Dev. 15, 581-590.
Sawa, S., Watanabe, K., Goto, K., Liu, Y.G., Shibata, D., Kanaya, E., Morita, E.H., and Okada, K. (1999). FILAMENTOUS FLOWER, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev. 13, 1079-1088.
Schoof, H., Lenhard, M., Haecker, A., Mayer, K.F.X., Jurgens, G., and Laux, T. (2000). The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100, 635-644.
Sculthorpe, C.D. (1967) The Biology of Aquatic Vascular Plants. Arnold, London
Sheldon, C.C., Rouse, D.T., Finnegan, E.J., Peacock, W.J., and Dennis, E.S. (2000). The molecular basis of vernalization: the central role of
FLOWERING LOCUS C (FLC). Proc. Natl. Acad. Sci. USA 97,
3753–3758.
Sheldon, C.C., Conn, A.B., Dennis, E.S., and Peacock, W.J. (2002). Different regulatory regions are required for the vernalization-induced repression of FLOWERING LOCUS C and for the epigenetic maintenance of repression. Plant Cell 14, 2527–2537.
Siegfried, K.R., Eshed, Y., Baum, S.F., Otsuga, D., Drews, G.N., and Bowman, J.L. (1999). Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 126, 4117-4128.
Simpson, G.G., Gendall, A.R., and Dean, C. (1999). When to switch to flowering. Annu. Rev. Cell Dev. Biol. 15, 519–550.
Sung, S., and Amasino, R.M. (2004). Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 427, 159–164.
Sylvester, A.W., Smith, L., and Freeling, M. (1996). Aquisition of identity in the developing leaf. Annu. Rev. Cell Dev. Biol. 12, 257- 304.
Szweykowska-Kulinska, Z., Jarmolowski, A., and Figlerowicz, M. (2003) RNA interference and its role in the regulation of eucaryotic gene expression. Acta Biochim. Pol. 50, 217 229.
Telfer, A., Bollman, K.M., and Poethig, R.S. (1997). Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 124, 645-654.
Tsuge, T., Tsukaya, H., and Uchimiya, H. (1996). Two independent and polarized processes of cell elongation regulate leaf blade expansion in
Arabidopsis thaliana (L.) Heynh. Development 122, 1589–1600.
Tsukaya, H. (1995). Developmental genetics of leaf morphogenesis in dicotyledonous plants. J. Plant Res. 108, 407–416.
Yamagchi, I., and Fukumoto, M. (1998). Alteration of hormone levels in transgenic tobacco plants overexpressing the rice homeobox gene OSH1. Plant Physiology 116, 471–476.
Vollbrecht, E., Veit, B., Sinha, N., and Hake, S. (1991). The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 350, 241-243.
Yoshida, N., Yanai, Y., Chen, L., Kato, Y., Hiratsuka, J., Miwa, T., Sung, Z.R., and Takahashi, S. (2001). EMBRYONIC FLOWER2, a novel polycomb group protein homolog, mediates shoot development and flowering in Arabidopsis. Plant Cell 13, 2471–2481.
Zhang, H., and van Nocker S. (2002). The VERNALIZATION
INDEPENDENCE 4 gene encodes a novel regulator of FLOWERING LOCUS C. Plant J. 31, 663-673.
Appendixes
pPZP:SK:BAR vector
RB: T-DNA right border
35S: CaMV 35S promoter Bar: Basta-resistant gene
Nos: Terminator
MCS: Multiple Cloning Sites Ori: Replication Origin
Ampr: Ampicillin-resistant gene LB: T-DNA left border
Promoter cis-element prediction by the PlantCARE
(http://oberon.fvms. ugent.be:8080/PlantCARE/index.html)
Site Name Function Organism Strand Position + -1319, -1906 A-box sequence conserved in alpha-amylase
promoters
Oryza sativa
- -987, -991, -1444, -1594, -2230 AACA_motif involved in endosperm-specific
negative expression
Oryza sativa - -1522
+ -213 ABRE cis-acting element involved in the
abscisic acid responsiveness
Arabidopsis
thaliana - -111 ACA-motif part of gapA in (gapA-CMA1)
involved with light responsiveness
Arabidopsis thaliana
- -675
+ -113, -215 ACE cis-acting element involved in light
responsiveness
Petroselinum
crispum - -246, -341, -972, -1102, -2446
+ -2098 AE-box part of a module for light response Arabidopsis
thaliana - -691, -1376 AT1-motif part of a light responsive module Solanum
tuberosum
+ -1523, -2488
ATCT-motif part of a conserved DNA module involved in light responsiveness
Arabidopsis thaliana
- -1530
AuxRE part of an auxin-responsive element Glycine max + -2480 + -2054 AuxRR-core cis-acting regulatory element
involved in auxin responsiveness
Nicotiana
tabacum - -743 Box-W1 fungal elicitor responsive element Petroselinum
crispum
- -732, -1198
CATT-motif part of a li ght responsive element Zea mays + -2154 DRE cis-acting element involved in
dehydration, low-temp, salt stresses
Arabidopsis thaliana
- -238
EIRE elicitor-responsive element Nicotiana tabacum
- -1469
ELI-box3 elicitor-responsive element Brassica oleracea - -2075, -1259 ERE ethylene-responsive element Dianthus
caryophyllus
+ -507, -561, -1622, -2029
+ -213, -339 Pisum sativum
- -111, G-Box cis-acting regulatory element
aestivum G-box cis-acting regulatory element
involved in light responsiveness
Arabidopsis thaliana
+ -341
+ -2450 GA-motif part of a light responsive element Arabidopsis
thaliana - -1206, -1334 GAG-motif part of a light responsive element Arabidopsis
thaliana
- -47, -49, -69
+ -984, -1455, -2451 GATA-motif part of a light responsive element Arabidopsis
thaliana - -2280, + -187, -265 GC-motif enhancer-like element involved in
anoxic specific inducibility
Zea mays
- -110 + -24, -872 GCN4_motif cis-regulatory element involved in
endosperm expression
Oryza sativa
- -216, -1042, -1141, -2427
GT1-motif light responsive element Arabidopsis thaliana
+ -654
Gap-box part of a light responsive element Arabidopsis thaliana - -1340 + -32, -330, -899, -1777, -1928, -2480
I-box part of a light responsive element
Arabidopsis thaliana - -421, -1189, -1840 + -35, -2334 Pisum sativum - -651 LAMP-element part of a light responsive element
Spinacia oleracea - -1668, -2067 + -1946 LTR cis-acting element involved in
low-temperature responsiveness Hordeum vulgare - -237, -284, -1579, -1587, -1998, -2132 MBS MYB Binding Site Zea mays + -733
+ -1895, -2019 MRE MYB binding site involved in light
responsiveness Petroselinum crispum - -1565, -1578, -1597, -1602, -1607, -1670, -1681, -1721, -1980, -2048, -2063, -2073, -2201, -2227