行政院國家科學委員會專題研究計畫 期中進度報告
食用菇類誘導人類白血病 U937 細胞分化、熟成作用、蛋白
質體學分析及其對鼠皮下轉植 CT26 腫瘤之抑制作用(2/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2313-B-002-024- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學食品科技研究所 計畫主持人: 張鴻民 計畫參與人員: 鄭芳琪、張琬晴、簡佳怡 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 5 月 23 日
行政院國家科學委員會補助專題研究計畫成果報告
食用菇類誘導人類白血病U937 細胞分化、成熟作用、蛋白質體學分 析及其對鼠皮下轉植CT26 腫瘤之抑制作用 (2/3) 計畫類別:個別型計畫 計畫編號: NSC93-2313-B-002-047 執行期限:92/08/01- 93/07/31 計畫主持人:張鴻民 計畫參與人員:鄭芳琪、張琬晴、簡佳怡 執行單位:台灣大學食品科技研究所 中華民國九十三年五月二十一日中文摘要
由去年度之研究成果得知,由柳松菇Agrocybe aegerita (AA)與鴻喜菇 Hypsizigus marmoreus (HM) 所分離而得之蛋白質具有明顯 U937 細胞生長抑制作用與誘導分
化成熟作用,本年度為以硫胺分離蛋白質區分後,餵食皮下轉殖大腸癌細胞 (CT-26)之 Balb/c 鼠,觀察此 polysaccharide-bound proteins (PBP) 於 100 或 200 mg/kg bw/day 劑量下對腫瘤之抑制作用,並與 5-FU (5-fluorouracil) group (正控制組) (10 mg 5-FU/kg bw/day)做比較。結果顯示, PBP 具有明顯 a dose-dependent 抑癌能力。與 控制組(PBS)比較,餵食 200 mg PBP/kg bw/day 組可表現出 39-44 % (by 52 % in 5-FU group)之抑制腫瘤作用,且其脾臟細胞 數 (3.2-2.9×108 cells/spleen) 明顯高於控制 組 (2.4×108 cells/spleen) 以及 5-FU (1.7×108 cells/spleen) 組,顯示 PBS 之脾臟細胞活 化作用。餵食PBS 組老鼠(100 and 200 mg HMPBP/kg bw/day)之血清總 IgG 量約
為5.1 mg/mL,明顯高於控制組之 3.1 mg/mL。而為時 PBS 組之體重、脾臟重與肝、
腎功能皆正常,說明此菇類口服劑量並無毒性。此二種菇類含多醣蛋白質為一優 良抑癌劑。
ABSTRACT Protein fractions from dietary mushrooms of Agrocybe aegerita (AA) and
Hypsizigus marmoreus (HM) were examined to have cytotoxicity against human leukemic U937
previously. To further study the antitumor effect of AA and HM proteins through immunomodulation, CT26-bearing Balb/c mice were orally administrated with
polysaccharide-bound proteins (PBP) at 100 or 200 mg/kg bw/day dose level and the inhibition of tumor was compared with control group and 5-FU (5-fluorouracil) group (positive control) (10 mg 5-FU/kg bw/day). Results showed that PBP from those mushrooms displayed significant antitumor activity at a dose-dependent manner. Mice orally administrated 200 mg PBP/kg bw/day showed marked reduction in tumor weight by 39-44 % (by 52 % in 5-FU group), in comparison to the control group. In addition, numbers of spleen cell from mice fed with PSP (3.2-2.9×108 cells/spleen) were much higher than that of control (2.4×108 cells/spleen) and 5-FU (1.7×108 cells/spleen) groups, revealing the apparent activation of splenocytes by PSP. Total IgG content in the plasma from mice fed with 100 and 200 mg HMPBP/kg bw/day reached a value of about 5.1 mg/mL, comparing to only 3.1 mg/mL in control group, revealing the immunity enhancement by PBP. No significant changes were observed in body weight, spleen weight and renal and hepatic functions in mice orally administrated with PBP throughout the experiment. PBP of AA and HM appear to be good candidates of cancer chemotherapy. KEY WORDS:•polysaccharide-bound protein•immunity•antitumor•Agrocybe aegerita•
INTRODUCTION
Chemoprevention is one of the most promising areas in cancer research. The majority of chemical compounds, which are cytotoxic to cancer cells, are often toxic to normal cells (1). Hence, discovery and identification of novel drugs, without severe side effects, to supplement the chemotherapy and radiotherapy of cancers and various infectious diseases become an important goal in biomedical research. Biologically active ingredients from mushrooms, fungi, yeasts, algae, lichens, herbs and plants attracted more attentions recently in the biochemical and medical applications for immunomodulatory and antitumor abilities (2-3).
Mushrooms have been proved to be good sources of almost all essential amino acids while compared with common vegetables (4), and are known for medicinaland therapeutic values, diversity of bioactive compounds and various physiological properties such as
immunological enhancement, maintenance of homeostasis, and regulation of biorhythm (5-10). Shon and Nam (10) revealed that Phellinus linteus, Phellinus igniarius, and Agrocybe
cylindracea had antimutagenic activity against direct- and indirect-acting mutagens and also
showed chemopreventive activity as inducers of phase II drug-metabolizing enzymes and glutathione in murine hepatoma cell culture. Protein-bound polysaccharidesoriginating from some edible mushrooms were claimed to have immunomodulating and immunoenhancing effects by inhibiting cancer growth and metastasis. Besides, they also increasedhost resistance to bacterial, viral and parasitic infections and therefore, were suggested as oral
supplements in health food stores (6-12). However, despite the widespread use of mushrooms and/or mushroom extracts for potential health benefits based on theories that enhance immune function by in vitro studies, there was surprising paucity of epidemiologic and experimental studies that address the biological activities after oral administration to animals or humans (13).
Previously, protein fractions from dietary mushrooms of Agrocybe aegerita (AA) and
Hypsizigus marmoreus (HM) were isolated to stimulate peripheral blood mononuclear cells
(MNC) for the preparation of conditioned medium (CM), which was then used to incubate with human myeloid U937 cells to observe the growth inhibition (MNC-CM method) on U937 cells (14-15). Interestingly, high levels of cytokines such as interleukine 1-β (IL-1β), tumor necrosis factor-α (TNF-α), and granulocyte macrophage-colony stimulating factor (GM-CSF) in MNC-CM prepared with PBP of Agrocybe aegerita were considered to be responsible for the antitumor and differentiation-inducing effects of U937 cells (14). Further pharmacological investigations on the antitumor effects of oral administration of mushroom proteins using an in
vivo model were examined in this study. First, polysaccharide-bound proteins (PBP) were
prepared and administrated to tumor-bearing mice to observe the tumor growth; then, splenocytes were collected for the mitogen-stimulated proliferation. Finally, antibody level in plasma and renal and hepatic functions in mice were determined.
MATERIALS AND METHODS
PBP preparation. Fresh AA and HM, purchased from a local supermarket, were blended
in a cycle blender (Osterizer, Mexican) with 2 parts (1:2) cold (4°C) distilled water for 1 min. Stirring in a chilling room for 24 h was conducted in a magnetic stirrer to extract proteins, followed by centrifugation (12000 × g, 4°C, 30 min) to obtain the supernatant, which was treated overnight with 100 % saturated ammonium sulfate to obtain precipitates. Subsequent dialysis at 4°C against distilled water was performed to obtain the water-soluble portions, which were free-dried for protein and polysaccharide assays and dissolved in PBS
(phosphate-buffered saline, 8 g NaCl/1.15 g Na2HPO4/0.2 g KH2PO4/0.2 g KCl/L) prior to use.
Experimental animals and experimental design. Male Balb/c mice (5 wk old) obtained
from Laboratory Animal Center, School of Medicine, National Taiwan University were randomly divided into test and control groups and housed in microisolator cages for 1 wk before
experiment. These mice had free access to water and to chow diet (LabDiet®, Brentwood, USA) and were housed in a room maintained at 23 ± 2°C with 60-70 % relative humidity. The room lights were on for 12 h/d starting at 0700 h. Mice were treated according to the ethics
guide-lines of the Animal Center, Taiwan University. The experimental protocol was approved by the Animal Studies Committee of Taiwan University.
For the in vivo study of cell-mediated immune response, 6-wk-old mice (about 19 g) were divided into six tumor-bearing mice groups and one normal group, each group comprised of 10
mice. CT26 is a murine colon cancer cell line obtained from American Type Culture Collection (Rockville, Maryland, USA). Mice were injected subcutaneously (sc) with 1 x 106 CT26 cells in 100 µL PBS to examine the effect of PBP from mushrooms and 5-FU on immune functions. Normal and control (tumor-bearing mice) group received PBS solution. 5-FU group was intraperitoneally (ip) administrated with 5-FU at 10 mg/kg bw/day. Mushroom proteins oral intake groups were administrated with AA PBP or HM PBP at 100 or 200 mg/kg bw/day dose levels. AA 100 group and AA 200 group, AA PBP 100 and 200 mg/kg bw/day, respectively; HM 100 group and HM 200 group, HM PBP 100 and 200 mg/kg bw/day, respectively. The animals were sacrificed 24 h after the last dose at 20th day. After inoculation for 10 days, tumor growth was monitored daily by measuring two perpendicular tumor diameters with calipers. Tumor size was calculated according to the following formula (16-17):
Tumor size (mm3) = 0.5 x (the major axis) x (the minor axis)2 mm3
Tumor inhibition ratio (%) = (1 - Tumor weight of experimental group/Tumor weight of control group) x 100 %
Formulas are adopted in all experiments in this paper. Statistical analysis was performed using repeated measures analysis of variance (ANOVA).
Body weight, spleen weight and cellularity. Atthe end of study, mice were sacrificed
via ethyl esterasphyxiation.Body weight and tumor weight were determined for each mouse. Spleens were removed aseptically, weighted and placed in RPMI 1640 (Gibco BRL,
Gaitherburg, MD, USA) medium supplemented with 10% FBS (Hyclone, Logan, USA) and 1% glutamine and pressed through a sterile 70 µm Falcon cell strainer to remove connective tissues (one spleen per strainer). Suspended splenocytes in medium were counted under a light microscope and were used in in vitro proliferation procedures.
Quantification of leukocyte and IgG. For leukocyte determination, blood (10-20 µL) collected from mice by orbital vascular plexus in capillary before and every two days during the experiment for up to 20 days was stained with Methyl blue (Sigma, St. Louis, MO, USA) and leukocytes were counted under a light microscope. For IgG determination, blood (about 1 mL) was collected by a cardiac puncture in a 0.5 M EDTA-rinsed syringe (Terumo, Elkton, MD, USA). After centrifugation (10,000 g; 20 min), blood plasma was collected and stored at -80°C until use. Mouse serum IgG (pg/mL) was quantified with a commercial ELISA kit (ICN, CA, USA) by a sandwich ELISA method (y= 0.3145 + 2.205/[1+(X/100.5)-0.9263], x = logarithmic value of IgG concentration (ng/mL); y= absorbance at 450 nm; R2 =0.999). Plates were read in a Precision microplate reader (Emax, Molecular Devices Co. USA) at a wavelength of 450 nm.
Lymphocyte proliferation assay. Single cell suspension prepared aseptically from fresh
splenocytes were suspended at a concentration of 5×106 cells/mL in medium containing RPMI 1640 supplemented with 10% FBS and 1% glutamine, and studies were carried out to evaluate the effects of AA PBP, HM PBP, and 5-FU on the proliferation of mouse lymphocytes in the absence or presence of 20 µg/mL PHA (phytohemagglutinin) (Sigma, St. Louis, MO, USA). The
spleen cell suspensions were incubated for 72 h at 37°C in a 5% humidified CO2 incubator. Cell proliferation was evaluated by using MTT tetrazolium assay as described by (18).
Evaluation of hepatic and renal dysfunction. Circulating levels of blood urea nitrogen
(BUN), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) were measured in plasma samples of all experimental animals after feeding period. Concentrations of BUN, AST, and ALT were analyzed by using a commercial kit (Randox Laboratories Ltd., Crumlin, Co. Antrim, U.K.) according to manufacturer's instructions of Automatic Analyzer (Chemwell, FLA, USA). Results are expressed as mg/dL or IU/L.
Protein and polysaccharide assays. Protein level in lyophilized powder was
determined by the Bradford method (19) using the Bio-Rad protein assay dye reagent. Bovine serum albumen (0.05-1.0 mg/mL) was used to construct the standard curve (R2=0.999). Phenol-sulfuric acid method was applied to quantify the polysaccharide content in powder using glucose (20-500 µg/mL) to construct the standard curve (R2=0.997). Triplicate samples each were analyzed twice.
RESULTS
Freeze-dried powders from mushrooms were dissolved in de-ionized water (1 mg/mL) and the protein and polysaccharide content were determined. It was found that AA powder contained about 570 µg/mL protein and 420 µg/mL polysaccharide, and HM powder contained
about 510 µg/mL protein and 485 µg/mL polysaccharide. SDS-PAGE was conducted to prove that polysaccharides were covalently bound to proteins through Coomassie Brilland Blue R-250 staining and PAS (periodic acid-Schiff) staining in the previous study (20).
Effect of AA and HM PBP on body weight and leukocyte number. PBP administration
was started in Balb/c mice after implantation of the CT26 cells, and thereafter, all test groups gained weight, from about 19 to 22 g/mouse during the feeding period, and no death occurred until terminal sacrifice. There were no significantly different in body weightsbetween mice administrated with PBP and those fed control diet over a periodof 20 days (data not shown). However, reduction in body weights of the mice in 5-FU group during the feeding period was observed. 5-FU induced body weight loss, from initial about 19 to about 18 g/mouse, after 12 days administration in this experiment (data not shown) is consistent with previous reports (21-23). In contrast, the control and 5-FU administrated mice were smaller and less active than the PBP supplemented mice. They appeared to be weaker and their hairs were apparently sparse at the end of feeding period.
Compared to the number (8.7 ± 0.5 × 103 cells/µL) of leukocyte in orbital vascular plexus at o day (before implantation of CT26 cell), oral administration of AA PBP at a dosage of 100 or 200 mg/kg bw/day exhibited a significant (p < 0.05) increase in leukocytes number by about 29 % at the end of feeding period, while only about 18 % increase in leukocytes number was determined in the HM PBP group at the corresponding dosage, in CT26-implanted mice (data
not shown). Significant reduction (p < 0.05) in leukocytes number (myelotoxicity) in 5-FU group was observed, in comparison with the PBP supplemented group and control group (data not shown), as was indicated by Kimura and Okuda (21-22).
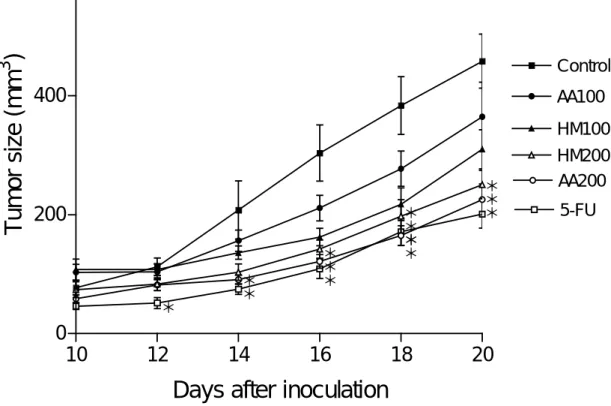
Antitumor activity of AA PBP and HM PBP. Tumor appeared at the site of the
inoculation and the volume progressively grew, which was measured from the 10th day after the implantation treatment. It was observed that 5-FU, AA PBP and HM PBP strongly suppressed tumor growth (Fig. 1). Increase in tumor size in control mice reached about 460 mm3 after 20 days of inoculation, while that in AA200 and HM200 group was about 220 and 240 mm3, respectively, revealing the significant (p < 0.05) inhibition on tumor growth by the intake of BPB. The inhibition ratio of tumor growth with oral administration of AA and HM was dose-dependent.
Tumor weights from sacrificed mice of 9-wk old were significantly (p < 0.05) reduced at the administration of 5-FU and AA PBP and HM PBP at 200 mg/kg bw/day dose level (Table 1). Tumor weight from control group of 0.62 g reduced to 0.38 and 0.35 g in HM200 and AA 200 group, respectively. Tumor inhibition ratio of AA200, HM200, and 5-FU group was 43.6, 38.7 and 51.6%, respectively. In addition, the net body weights of AA 200, HM100, and HM200 were higher than that of control and 5-FU administrated mice.
Effects of AAPBP and HM PBP on spleen weight and spleen cell number. The
spleen weight of CT26-bearing mice (6.5-7.3 g) was significantlygreater than that of the normal mice (3.7 g) (p < 0.05). In CT26-bearing mice, no effect (p > 0.05) in the spleen weight was
observed among 5-FU, any dose of AA and HM administration and control (normal saline-treated) mice (Table 2).
As regards lymphoid organ cellularity, the total number of spleen leukocytes was
significant (p < 0.05) increased at any dose of AA and HM administrated and control mice than that of the normal mice (Table 2). A significant (p < 0.05) increase in AA100 and HM200 groups and decrease in 5-FU and normal groups was also recorded as compared with control mice.
PHA-induced lymphoproliferative responses of spleen cells. Lymphoproliferative
responses of immune cells from spleens were examined in CT26-bearing mice treated with AAPBP and HMPBP as a nonspecific immunostimulating agent. Proliferation of spleen cells obtained from AAPBP, HMPBP, and 5-FU treated as well as the nontreated control groups of sacrificed CT26-implanted mice and stimulated by PHA was detected by MTT colorimetric assay. In the presence of optimal dose of PHA (10 µg/ml), oral administration of AA PBP and HM PBP did not show any suppressive effect on splenocytes even at high doses. The spleen cells from AA100 and AA200 administrated mice exhibited significantly (p < 0.05) higher proliferative activity than the proliferation noted in control mice (Fig. 2). Spleen cells obtained from HM PBP administrated mice slightly proliferated in response to PHA as compared with the control mice. 5-FU slightly depressed lymphoproliferative responses of spleen cells may be due to the toxic effect.
level of serum IgG at HMPBP administrated groups showed significant (p < 0.05) difference from the control mice (3.2 ± 0.3 mg/mL) (Fig. 3). The mean value of IgG was 3.3 ± 0.5 and 3.7 ± 0.3 mg/mL in AA100 and AA200, respectively. Administration of AAPBP at 200 mg/kg/day dose level (AA200) resulted in slight augmentation of IgG production as compared with the control mice. The mean value of IgG showed a tendency to be increased in HM100, HM200 and AA200 administrated mice versus normal mice.
Cardiac trunk blood was collected and stored at -80°C for the determination of BUN, AST and ALT. No significant difference (p > 0.05) in BUN, AST and ALT levels was observed in cardiac trunk blood of any treatment as compared with the normal and control mice (Table 3).
DISCUSSION
Oral administration of AA PBP and HM PBP at 100-200 mg/kg/day dose level induced 28-29 and 17-18% increase in leukocytes, respectively. Tumor size, tumor weight and tumor growth rate were significantly (P < 0.05) reduced at intraperitoneal administration of 5-FU and oral administration of AA PBP and HM PBP at 200 mg/kg/day dose level (Fig. 1 and Table 1). That is, AA and HM PBP (at 200 mg/kg/day dose level) can use as a therapeutic agent against a murine tumor model of colon carcinoma C26 in tumor-bearing mice and inhibit tumor growth via oral route. Chang et al. (14) indicated that high levels of cytokines such as IL-1β (1000 pg/mL ), TNF-α (1200 pg/mL), and GM-CSF (310 pg/mL) in MNC-CM prepared with proteins of
Agrocybe aegerita were detected and responsible for the antitumor effect of U937 cells.
In CT26-bearing mice, administrated of AA PBP and HM PBP exhibited an increase of the spleen leukocytes as compared with control mice (Table 2). Gendron et al. (24) revealed that leukocytes from peripheral immune tissues, including peripheral blood and spleen, are contributing to the immune response. Therefore, following administration of AA PBP and HM PBP all the CT26-implanted mice have equivalent increases in spleen leukocyte numbers, suggesting that the polysaccharide-bound proteins from AA and HM are contributing to immune activation.
Concanavalin A (Con A) and PHA are applied as T-cell specific stimulants and lipopolysaccharide (LPS) as a B-cell specific stimulant respectively (25). Normal human peripheral blood lymphocytes and spleen cells while stimulated by PHA and Con A produced T-cell growth factor (26-27). T-cell growth factor was considered relating to the clinical course of the disease and promotes the growth of adult T-cell leukemia cells in vivo (28). AA PBP
enhanced PHA-induced spleen cell proliferation, suggesting that it is a potent activator for T-cell proliferation and can be a candidate immuno-stimulator to improve immune responses.
The deterioration of the liver and kidney functions is assessed by elevated BUN, AST and ALT (29). Hence, the similar level (P > 0.05) of AST, ALT and BUN level in blood of any
treatment from the normal and control mice (Table 2), indicating that hepatic and renal dysfunction is not appeared while oral administration of AA and HM.
Regarding the effects on cancer chemotherapy of 5-FU, the combined effect of carp (Cyprinus carpio L.) extract or chitosan was used for the prevention of gastrointestinal toxicity, immunotoxicity, and myeloxicity in sarcoma 180-bearing ICR mice (21-22). In this study, no death occurred in AA PBP and HM PBP administrated mice, none of the AA and HM caused side effects such as reduction in body weight, myelotoxicity, immunotoxicity (reduction in spleen weight and spleen cell numbers) or plasma biochemistry profile that are usually caused by cancer chemotherapy drugs (30) revealed that oral intake of AA PBP and HM PBP at 100-200 mg/kg are harmless to mice and do not induce lethal death. Moreover, the AA PBP and HM PBP at 200 mg/kg/day dose level showed the same tumor inhibition ability with 5-FU. It has been reported that 5-FU causes immunosuppression and immunotoxicity as well as a reduction in the weights of the spleen and thymus (21-22, 31-32). Therefore, the immune stimulation and other beneficial effects for reducing the incidence of cancer reveal that the
polysaccharide-bound proteins of AA and HM are good candidates of cancer chemotherapy drugs; daily intake of Agrocybe aegerita and Hypsizigus marmoreus extracts may have potential for prevention of human cancers.
Although the present results yielded an incomplete picture on the effects of the AA PBP and HM PBP on the immune system and HM PBP can stimulate the production of nonspecific IgG antibodies, we also demonstrated that in the presence of mitogen (PHA) there was a potential immune response. The mechanism whereby the mushroom PBP modulates the
immune system remains unclear. Hence, further works to identify the active ingredientsin AA and HM that are responsible for the observed antitumor activityand immune stimulation are under study.
Acknowledgement
Financial support for this study from the National Science Council of the Republic of China under Grant NSC-93-2313-B-002-024 is greatly appreciated.
LITERATURE CITED
1. Kim, H. M., Han, S. B., Oh, G. T., Kim, Y. H., Hong, N. D. & Yoo, I.D. (1996) Stimulation of humoral and cell mediated immunity by polysaccharide from mushroom Phellinus linteus. Int. J. Immunopharmacol. 18: 295–303.
2. Ooi, V. E. C. & Liu, F. (2000) Immunomodulation and anticancer activity of polysaccharide protein complexes. Curr. Med. Chem. 7: 715–729.
3. Chen, Y. Y. & Chang, H. M. 2004. Antiproliferative and differentiating effects of polysaccharide fraction from fu-ling (Poria cocos) on human leukemic U937 and HL-60 cells. Food Chem. Toxicol. 42:759-769.
4. Mattila, P., Salo-Vaananen, P., Konko, K., Aro, H. & Jalava, T. (2002) Basic composition and amino acid contents of mushrooms cultivated in Finland. J. Agric. Food Chem. 50: 6419-6422.
5. Breene, W. M. (1990) Nutritional and medicinal value of specialty mushrooms. J. Food Protect. 53: 883-894.
6. Gan, L., Zhang, D. H., Yang, X. L. & Xu, H. B. (2004) Immunomodulation and antitumor activity by a polysaccharide–protein complex from Lycium barbarum. Int. Immunopharmacol. 4: 563-569.
7. Jong, S. C. & Birmingham, J. M. (1993) Medicinaland therapeutic value of the Shiitake mushroom. Adv.Appl. Microbiol. 39: 153-184.
8. Matsunaga, K., Morita, I. & Oguchi, (1987) Y. Restoration of immune responsiveness by a biological response modifier, PSK, in aged mice bearing syngeneic transplantable tumor. J. Clin. Lab. Immunol. 24: 143-149.
9. Matsunaga, K., Iijima, H., Aota, M., Oguchi, Y., Fujii, T., Yoshikumi, C. & Nomoto, K. (1992) Enhancementof effector cell activities in mice bearing syngeneic plasmacytomaX5563 by biological response modifier, PSK. J.Clin. Lab. Immunol. 37: 21-37.
10. Shon, Y. H & Nam, K. S. (2001) Antimutagenicity and induction of anticarcinogenic phase II enzymes by basidiomycetes. J. Ethnopharmacol. 77:103-109.
11. Cui, J. & Chisti, Y. (2003) Polysaccharopeptides of Coriolus versicolor: physiological activity, uses, and production. Biotechnol. Adv. 21: 109-122.
12. Wu, D., Han, S. N., Bronson, R. T., Smith, D. E. & Meydani, S. N. (1998) Dietary supplementation with mushroom-derived protein-bound glucan does not enhance immune function in young and old mice. J. Nutr. 128: 193-197.
13. Borchers, A. T., Keen, C. L. & Gershwin, M. E. (2004) Mushrooms, tumors, and immunity: an update. Exp Biol Med (Maywood). 229: 393-406.
14. Chang, H. M.; Hsu, L. L.; Chang, J. H.; Chen, Y. J. (2004) Antiproliferation and differentiation of human leukemic U937 cells induced by proteins from Agrocybe aegerita and Hypsizigus mamoreus. Food Chem. Toxicol. Submission.
leukemic U937 cells induced by proteins from dietary mushroom of Agrocybe aegerita. J Proteome Res. Accepted.
16. Makino, M., Shoji, H., Takemoto, D., Honboh, T., Nakamura, S., Kurayoshi, K. & Kaibara, N. (2001) Comparative study between daily and 5-days-a –week administration of oral 5-fluorouracil chemotherapy in mice: determining the superior regimen. Cancer Chemother. Pharmacol. 48: 370-374.
17. Takaku, T., Kimura, Y. & Okuda, H. (2001) Isolation of an antitumor compound from Agricus
Blazei Murill and its mechanism of action. J. Nutr. 131: 1409-1413.
18. Gerlier, D. & Thomasset, N. (1986) Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 94: 57-63.
19. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 72: 248-254.
20. Huang, Z. J. (2003) Proteomic analysis of human leukemic U937 cells following treatment with agrocybe aegerita protein extracts. Master Thesis of Graduate Institute of Food Science and Technology, National Taiwan University, Taipei, Taiwan.
21. Kimura, Y. & Okuda, H. (1999a) Prevention by carp extract of myelotoxicity and gastrointestinal toxicity induced by 5-fluorouracil without loss of antitumor activity in mice. J. Ethnopharmacol. 68: 39-45.
22. Kimura, Y. & Okuda H. (1999b) Prevention by chitosan of myelotoxicity, gastrointestinal toxicity and immunocompetent organic toxicity induced by 5-fluorouracil without loss of antitumor activity in mice. Jpn. J. Cancer Res. 90: 765-774.
23. Yamamoto, S., Kurebayashi, J., Kurosumi, M., Kunisue, J., Otsuki, T., Tanaka, K., & Sonoo, H. (2001) Combined effects of docetaxel and fluoropyrimidines on tumor growth and expression of interleukin-6 and thymidine phosphorylase in breast cancer xenografts. Cancer Chemother. Pharmacol. 48: 283-288.
24. Gendron, A., Teitelbaum, J., Cossette, C., Nuara, S., Dumont, M., Geadah, D., du Souich, P. & Kouassi, E. (2002) Temporal effects of left versus right middle cerebral artery occlusion on spleen lymphocyte subsets and mitogenic response in Wistar rats. Brain Res. 955: 85-97.
25. Nakamura, A., Nagai, K., Suzuki, S., Ando, K. & Tamura, G. (1986) A novel method of screening for immunomodulating substances, establishment of an assay system and its application to culture broths of microorganisms. J. Antibiot. 39: 1148-1154.
26. Gillis, S., Baker, P. E., Ruscetti, F. W. & Smith, K. A. (1978a) Long-term culture of human antigen-specific cytotoxic T-cell lines. J. Exp. Med.148: 1093-1098.
27. Gillis, S., Ferm, M. M., Ou, W. & Smith, K. A. (1978b) T cell growth factor: parameters of production and a quantitative microassay for activity. J. Immunol. 120: 2027-2032. 28. Tsuda, H. & Takatsuki, K. (1983) Correlation of aberrant proliferation with T-cell growth
factor in adult T-cell leukemia cells. Hematol. Oncol. 1: 177-185.
29. Sener, G., Kacmaz, A., User, Y., Ozkan, S., Tilki, M. & Yegen, B. C. (2003) Melatonin ameliorates oxidative organ damage induced by acute intra-abdominal compartment syndrome in rats. J. Pineal. Res. 35: 163-168.
30. Takaku, T., Kimura, Y. & Okuda, H. (2001) Isolation of an antitumor compound from
Agaricus blazei Murill and its mechanism of action. J. Nutr. 131: 1409-1413.
31. Inoue, T., Anderson, T. D., Hayes, T. J. & Horii, I. (1996) Comparative immunotoxicity assessment of N4-Trimethoxybenzoyl-5'-deoxy-5- fluorocytidine (Ro 09-1390) and 5'-deoxy-5-fluorouridine (5'-DFUR) in BDF1 mice. J. Toxicol. Sci. 21: 29-39.
32. Itoh, H., Ito, H., Amano, H. & Noda, H. (1994) Inhibitory action of a (1-->6)-beta-D-glucan-protein complex (F III-2-b) isolated from Agaricus blazei Murill ("himematsutake") on Meth A fibrosarcoma-bearing mice and its antitumor mechanism. Jpn. J. Pharmacol. 66: 265-271.
10 12 14 16 18 20 0 200 400 600 AA100 HM200 HM100 AA200 Control 5-FU
Days after inoculation
T
u
m
o
r s
iz
e
(m
m
3)
FIGURE 1 Changes in tumor size (mm3) of CT26-implanted Balb/c mice orally administrated with polysaccharide-bound proteins from Agrocybe aegerita and Hypsizigus marmareu. The tumor volume was determined by direct measurement with calipers and calculated by the formula [length (mm)×width (mm)2/2]. Values are means ± SEM, n=10. 5-FU, mice intraperitoneally administrated with 5-FU (10 mg/kg body weight/day); control, mice orally administrated with Saline solution. HM100 and HM200, 100 and 200 mgHM/kg body weight/day, respectively. AA100 and AA200, 100 and 200 mgAA/kg body weight/day, respectively.
*Significantly different at the 0.05 levels compared to control.
* * * * * * * * * * * * * *
N C HM100 HM200 AA100 AA200 5-FU 0.0 0.2 0.4 0.6 0.8 1.0
A
b
so
rb
an
ce
(
5
70
n
m
)
Treatment
FIGURE 2 Comparison of PHA-stimulated proliferations of spleen cells from CT26-implanted
mice orally administrated with polysaccharide-bound proteins from Hypsizigus marmareus and
Agrocybe aegerita. Values are means ± SEM, n=10. 5-FU, mice intraperitoneally administrated
with 5-FU (10 mg/kg body weight/day); control, mice orally administrated with Saline solution. HM100 and HM200, 100 and 200 mg HM/kg body weight/day, respectively. AA100 and AA200, 100 and 200 mg/kg body weight/day, respectively. Means without a common letter differ, p<0.05. e cd de abc ab a bc
N C HM100 HM200 AA100 AA200 5-FU 0 1 2 3 4 5 6 7
Treatment
C
o
nce
n
tr
at
io
n
of
I
g
G
(
m
g
/m
L
)
FIGURE 3 Comparison of IgG content in EDTA-plasma of CT26-implanted mice orally
administrated with polysaccharide-bound proteins from Hypsizigus marmareus and Agrocybe
aegerita. Values are means ± SEM, n=10. 5-FU, mice intraperitoneally administrated with 5-FU
(10 mg/kg body weight/day); control, mice orally administrated with Saline solution. HM100 and HM200, 100 and 200 mg HM/kg body weight/day, respectively. AA100 and AA200, 100 and 200 mg/kg body weight/day, respectively. Means without a common letter differ, p<0.05.
a a bc b bc c bc
Table 1
Comparison of net body weight and tumor weight change in CT26-implanted mice orally administrated with polysaccharide-bound proteins from Hypsizigus marmareus and Agrocybe
aegerita after feeding period 1
Group of mice Body weight (g) Tumor weight (g) Net body weight (g) Control 2 20.53 ± 0.62 bc 0.62 ± 0.33 a 19.91 ± 0.29 bc HM100 22.76 ± 3.11 ab 0.49 ± 0.10 ab 22.27 ± 2.94 a HM200 23.44 ± 2.36 ab 0.38 ± 0.11 b 23.07 ± 2.35 a AA100 22.14 ± 1.66 a 0.60 ± 0.23 a 21.54 ± 1.77 ab AA200 22.70 ± 0.91 ab 0.35 ± 0.10 b 22.35 ± 0.82 a 5-FU 3 18.86 ± 1.36 c 0.30 ± 0.12 b 18.56 ± 1.35 c 1 Values are means ± SEM, n=10. Mean values in the same column with a letter in common do
not differ significantly (p > 0.05). 2 Saline group.
Table 2
Comparison of spleen weight and spleen cell numbers in CT26-implanted mice orally administrated with 5-FU and polysaccharide-bound proteins from Hypsizigus marmareus and
Agrocybe aegerita 1
Group of mice Weight of spleen (mg/g of bw) Number of spleen cells (x 108/spleen)
Normal 3.7 ± 0.1 b 1.8 ± 0.2 c Control 2 6.5 ± 0.7 a 2.4 ± 0.2 b HM100 6.6 ± 0.4 a 2.9 ± 0.3 ab HM200 6.3 ± 0.3 a 3.1 ± 0.1 a AA100 7.3 ± 0.2 a 3.2 ± 0.2 a AA200 7.0 ± 0.3 a 3.0 ± 0.2 ab 5-FU 3 6.9 ± 0.7 a 1.7 ± 0.2 c
1 Values are means ± SEM, n=10. Mean values in the same column with a letter in common do not differ significantly (p > 0.05).
2 Saline group.
Table 3
Plasma biochemistry profiles in CT26-implanted Balb/c mice after feeding period 1 Group of mice (IU/L) ALT 2 AST(IU/L) 3 (mg/dL) BUN 4
Normal 31.90 ± 3.63 110.80 ± 17.48 19.25 ± 2.28 Control 26.00 ± 3.07 131.75 ± 18.47 21.81 ± 5.98 HM100 27.38 ± 4.35 122.00 ± 23.20 15.17 ± 1.80 HM200 22.00 ± 2.57 118.00 ± 23.14 15.60 ± 1.48 AA100 22.63 ± 1.78 125.50 ± 15.40 19.85 ± 3.13 AA200 24.33 ± 2.91 126.00 ± 20.41 16.02 ± 2.77 5-FU 26.25 ± 2.04 109.30 ± 14.34 16.50 ± 1.79 1 Values are means ± SEM, n=10. Mean values in the same column with a letter in
common do not differ significantly (p > 0.05). 2 Alanine aminotransferase
3 Aspartate aminotransferase 4 Blood urea nitrogen