電刺激大鼠側韁核對區辨性低頻操作式制約行為的影響 - 政大學術集成
70
0
0
全文
(2) 謝. 誌. 感謝口試委員郭博昭老師、賴桂珍老師與廖瑞銘老師的指導與補強,讓 本篇論文能更完整。感謝指導教授廖瑞銘老師嚴謹及細心的督促我對於論 文的種種細節,讓我對於做研究及撰寫學術論文有更深的了解。感謝楊幼 屏學姊不辭勞苦的幫忙一起收集實驗數據,黃鈺婷、陳俊宏幫忙訓練大白 鼠,有了大家的幫忙,實驗才能加快腳步。感謝楊幼屏學姊提供對於口試 的技巧;感謝江峰逵學長教導我關於動物行為的相關知識;感謝實驗室其. 立. 政 治 大. 他人在各方面的協助。感謝我的父母,讓我在沒有經濟壓力的狀況下,支. ‧ 國. 學. 持我完成研究所的工作。感謝為了我實驗犧牲的大白鼠們,有了你們的貢. ‧. 獻,本篇論文才能完成。最後感謝我的神,讓我順利完成此階段的學習,. Nat. n. al. er. io. sit. y. 將一切榮耀歸給祂。. Ch. engchi. I. i Un. v.
(3) 中文摘要 透過神經科學的研究,對於大腦的行為功能已有一定的認識,不同於以往的認識, 目前認為神經行為機制不只由單一腦區或單一神經化學系統所調控。深部大腦電刺激經 常被用來研究特定腦區的行為功能。但是,深部大腦電刺激的作用機制仍然不清楚。最 近幾年臨床研究發現,利用電刺激在側韁核成功的治療憂鬱症患者。然而,目前認為側 韁核與多巴胺系統互為負回饋作用,共同參與在動機行為的酬賞反應中。本實驗室先前 的研究顯示,破壞韁核造成區辨性低頻操作式制約行為 (簡稱 DRL 行為)學習的障礙, 然而,電刺激在側韁核造成 DRL 行為表現的結果還是未知的。所以,本實驗主要以電. 政 治 大. 刺激在側韁核觀察大鼠行為上的改變,探討側韁核在行為上參與的功能。實驗一的結果. 立. 顯示電刺激在側韁核並不影響自發性運動能力,在不同電流強度的刺激下也不會影響。. ‧ 國. 學. 實驗二的結果顯示電刺激在側韁核造成 DRL 15 秒的行為有類安非他命效果之行為表. ‧. 現,在高頻率電刺激有較顯著類安非他命的效果。實驗三的結果顯示電刺激在側韁核造. y. Nat. 成 DRL 15 秒的行為之影響,會被多巴胺受體抑制劑所抵消,而單獨注射巴胺受體抑制. er. io. sit. 劑並不影響 DRL 15 秒的行為。實驗四的結果顯示電刺激在側韁核造成 DRL 15 秒的行 為之影響,不會被正腎上腺素受體抑制劑所抵消。實驗五的結果顯示電刺激在側韁核造. al. n. iv n C 成 DRL 72 秒的行為之影響並不如 DRL h e15n 秒的行為顯著。實驗六的結果顯示電刺激在 gchi U. 側韁核並不會造成大鼠無法區辨酬賞的量。綜合而言,側韁核在動機行為的角色,是透 過影響多巴胺系統造成行為的改變。. 關鍵詞:深部大腦電刺激,區辨性低頻操作式制約(DRL)行為,側韁核,多巴胺受體抑 制劑,正腎上腺素受體抑制劑。. II.
(4) Abstract Behavioral function of the brain has been studied in neuroscience and progressively accumulated informative data to reveal the neurobehavioral mechanisms.. It is now realized that those underlying mechanisms of behaviors is. not as such simple as previous thought of limiting only in one locus of the brain or solely by one neurochemical system. The deep brain stimulation is usually used to study the behavioral function of specific brain regions. the deep brain stimulation is still unclear.. However, the mechanism of. The previous study has shown that. 政 治 大 It is proposed that an inhibitory role of LHb on the mibrain 立. electrical stimulation of the lateral habenula (LHb) successfully treated depression symptoms in the patients.. ‧ 國. 學. dopamine (DA) system which mediates the reward-related behavior. A previous study of this lab showed that lesion of habenula impaired the acquisition of differential. ‧. reinforcement of low-rate responding (DRL) behavior. But, the effect of LHb. y. To determine the functions of LHb. sit. Nat. stimulation on the DRL behavior is still unclear.. n. al. er. io. involving in the behavior, the electrical stimulation was applied in LHb to observe the behavioral change of rats.. i Un. v. The results of Experiment 1 showed that the LHb. Ch. e n g c hIni Experiment 2, the LHb stimulation. stimulation had no effect on locomotor activity.. was shown to affect DRL 15-s behavior, which effects were similar to those affected by amphetamine. Experiment 3 showed that the DA receptor antagonists reversed the effects of LHb stimulation, while experiment 4 showed that norepinephrine (NE) receptor antagonists had no reversal effect on DRL 15-s behavior.. In Experiment 5,. the amphetamine-like behavior induced by LHb stimulation had subtle effects on DRL 72-s behavior.. Experiment 6 showed that the LHb stimulation had no effect on a. discrimination task. These data suggest that the LHb modulating DRL behavior is DA-dependent. III.
(5) Key Words: deep brain stimulation, differential reinforcement of low-rate responding behavior, lateral habenula, dopamine receptor antagonists, norepinephrine receptor antagonists. 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. IV. i Un. v.
(6) Contents 謝誌…………………………………………………………………………….Ⅰ 中文摘要……………………………………………………………………….Ⅱ Abstract………………………………………………………………………..Ⅲ Contents…………………………………………………………………........Ⅴ List of Tables………………………………………………………………….Ⅶ. 政 治 大 Introduction……………………………………………………………………01 立. List of Figures…………………………………………………………..........Ⅷ. ‧ 國. 學. The application of deep brain stimulation in clinical and pre-clinical studies.……………………………………………………………………...02. ‧. The lateral habenulal………………………………………………………03. sit. y. Nat. The LHb and differential reinforcement of low-rate responding. er. io. behavior……………………………………………………………………..05. n. Aims and the rationaleaofl this study……………………………………...07 iv. n. C. hengchi U Materials and Methods……………………………………………………....09 Animals……………………………………………………………………..09 Apparatus…………………………………………………………………..09 DRL operant behavior………………………………………………….09 Locomotor activity………………………………………………………10 DRL behavioral training………………………………………………..10 Discrimination task……………………………………………………...11 Surgery……………………………………………………………………..12. Electrode preparation and habenula stimulation……………………….12 V.
(7) Drugs………………………………………………………………………..13 Procedures…………………………………………………………………13 Experiment 1…………………………………………………………….13 Experiment 2…………………………………………………………….14 Experiment 3…………………………………………………………….15 Experiment 4…………………………………………………………….16 Experiment 5…………………………………………………………….17 Experiment 6…………………………………………………………….17 Histology……………………………………………………………………17. 治 政 DRL behavioral data………………………………………………………18 大 立 Statistical analyses………………………………………………………..19 ‧ 國. 學. Results………………………………………………………………………...20. ‧. Experiment 1……………………………………………………………….20. y. Nat. Experiment 2……………………………………………………………….20. er. io. sit. Experiment 3……………………………………………………………….21 Experiment 4……………………………………………………………….23 a. n. iv l C n Experiment 5……………………………………………………………….24 hengchi U. Experiment 6……………………………………………………………….24 Discussion…………………………………………………………………….27 References……………………………………………………………………39. VI.
(8) List of Tables Table 1. The effects of lateral habenula (LHb) stimulation on DRL 72-s Behavior (Experiment 5)………………………………………45. Table 2. Within-session analyses of the effects of lateral habenula (LHb) stimulation on a reward discrimination task (Experiment 6)…………………………………………………..46 Number of choices made in each 10-trial block on discrimination task (Experiment 6)……………………………47. 立. 政 治 大. 學 ‧. ‧ 國 io. sit. y. Nat. n. al. er. Table 3. Ch. engchi. VII. i Un. v.
(9) List of Figures Figure 1. The electrode and brain stimulation in the rat……………….48. Figure 2. Histology of brain section with the LHb………………………49. Figure 3. The effects of electrical stimulation in LHb on locomotor activity (Experiment 1)…..………...…………………………50. Figure 4. The dose effects of amphetamine on the distance of locomotor activity (Experiment 1).……………...…………..51. Figure 5. The effects of LHb stimulation on DRL 15-s behavior as. 政 治 大 The effects of立 LHb stimulation on the MRE ratio of DRL-15. measured by the six dependent variables (Experiment 2).52. Figure 6. ‧ 國. Figure 7. 學. behavior (Experiment 2).…………………………………….53 The effects of SCH23390 and eticlopride on the alteration of. ‧. DRL 15-s behavior induced by LHb stimulation as. y. sit. The effects of SCH23390 and eticlopride on the alteration of. er. io. Figure 8. Nat. measured by the six dependent variables (Experiment 3).54. n. al DRL 15-s behavior induced by LHbi vstimulation as n. C. h eratio measured by MRE n g(Experiment c h i U 3)….………………..55 Figure 9. The effects of SCH23390 and eticlopride on DRL 15-s behavior, as measured by the six dependent variables (Experiment 3)……………………………………………..…56. Figure 10. The effects of SCH23390 and eticlopride on DRL 15-s behavior as measured by MRE ratio (Experiment 3).…...57. Figure 11. The effects of prazosin, yohimbine and propranolol on the alteration of DRL 15-s behavior induced by LHb stimulation, as measured by the six dependent variables (Experiment VIII.
(10) 4)……………………………………………………………….58 Figure 12 The effects of prazosin, yohimbine and propranolol on the alteration of DRL15-s behavior induced by LHb stimulation as measured by MRE ratio(Experiment 4).……………….59 Figure 13 The effects of electrical stimulation in LHb on a reward discrimination task, as measured by choice of large reward, omission rate and response latency(Experiment 6).…….60. 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. IX. i Un. v.
(11) Introduction While the issue of behavioral function of the brain has been heavily focused in neuroscience and progressively accumulated informative data to reveal the neurobehavioral mechanisms of various kinds of behavior from the past, it is now realized that those underlying mechanisms for the behavior may not be simply as previous thought of limiting only in one locus of the brain or solely by one neurochemical system. In terms of investigating the brain/behavior interaction, multiple research approaches that use the methods of lesion, stimulation and. 政 治 大 underlying neurobehavioral mechanisms. In another words, checking data 立. recording have to be taken in account for providing a better profile in elaborating the. ‧ 國. comprehensive conclusion can be made.. 學. consistently across different approaches on a specific issue is essential before a more Such that, also following the idea of neural. ‧. circuitry for behavioral function, it is important to examine a nucleus interacts with. y. sit. n. al. er. For instance the dopamine (DA) related reward motivation;. io. acceptable scale.. Nat. another that has been established (if not completely known) for its function with an. i Un. v. despite that a great deal of emphasis has been placed on behavior performance. Ch. engchi. driven by the increased DA neuronal activation, an alternative mechanism may have the active removal of an inhibitory control on DA neurons.. Accordingly, it is possible. that the neural processes of DA related function, from physiological to behavioral level, could be modulated or interacted by other nuclei.. A growing body of evidence. suggests that the habenula (Hb) plays a modulatory role on the mid brain DA systems (reviews see Hikosaka, 2010; Hikosaka et al., 2008).. To further test this proposition,. the present study was designed to investigate the role of lateral Hb involved in DA-related behavior by using the brain stimulation approach.. The use of brain. stimulation approach is set to be complimentary for a previous study using lesion 1.
(12) approach, both examining how the Hb influences the DA-related behavior.. The application of deep brain stimulation in clinical and pre-clinical studies Deep brain stimulation (DBS) is one of the therapeutic potential of neuromodulatory techniques today.. High frequency of DBS had been used in. treatment of neurological diseases and movement diseases (Gross et al., 2000; Wichmann et al., 2006). Such as, DBS of the subthalamic nucleus (STN) improved the symptoms of Parkinson’s disease (PD) (Perozzo et al., 2001), while DBS of the thalamus was used to treat epilepsy and Tourette’s syndrome (Zumsteg et al., 2006;. 政 治 大. Maciunas et al., 2007). In recent years, DBS had also been used in the treatment of. 立. Mayberg and associates (2005) demonstrated that DBS of the. 學. ‧ 國. depression patients.. subgenual cingulate white matter successfully treated depression symptoms in six Several other brain areas have been targeted to test the potential. ‧. patients.. sit. y. Nat. effectiveness of DBS treatment for depressed patients or animal models, including the. io. er. lateral habenula nucleus (LHb) (Li et al., 2011), the anterior limb of the anterior internal capsule (Gutman et al., 2009), the nucleus accumbens (NAc) (Schlaepfer et. al. n. iv n C al., 2008) and the thalamic peduncle h(Jimenez i U2005). e n g c eth al.,. Despite its clinical. application, it is unfortunately that the neural mechanism of DBS is still unknown. In terms of DBS of STN changed various neurotransmitter systems, previous studies showed that bilateral DBS of STN produced a decreasing effect on the firing rate of 5-hydroxytryptamine (5-HT) neurons (Temel et al., 2007).. It was also showed. that unilateral DBS of the left STN increased glutamate levels in the STN (Lee et al., 2007). In addition, GABAergic neurons are suggested to play an important role in the high frequency stimulation (Feuerstein et al., 2011). DBS of slices adopted from the striatum was demonstrated to increase extracellular GABA levels in vitro (Li et al., 2004), which results are consistent to an in vivo study showing DBS of the striatum 2.
(13) enhanced local GABA outflow in freely moving rats (Hiller et al., 2007). Thus, it is likely that the DBS mechanism would cause the changes of neurochemical transmission leading to functional alteration. However, it is still unclear about the neurochemical changes are dependent on the brain area where the DBS is applied. Pharmacological test administered in combining with DBS can be used to encounter this issue.. The lateral habenula There is a growing body of evidence indicating a strong correlation between the. 政 治 大. depression state and the hyperactivity in the habenula, especially in the LHb area. 立. (Sartorius and Henn, 2007; Hikosaka et al., 2008). In combining with the. ‧ 國. 學. aforementioned literature addressing DBS induced neurochemical effects, it is. ‧. reasonable inferred that the habenula is involved in the control of emotion and. sit. y. Nat. motivation. The shift of neural activity of habenular may alter the individual’s. io. er. behavioral function and mental state. Thus, to verify the neuroanatomy and neurophysiological function of the habenula may lead a further understanding of the. al. n. iv n C neurobehavioral mechanisms of emotion motivation which serve as the h e nand gchi U fundamental processes for the cognitive and higher order function.. The habenula complex was one of the areas of the brain and both hemispheres were symmetrical. The habenula complex formed a part of the cross-talk between limbic forebrain and some important ascending modulatory pathways.. The habenula. complex situated at the caudal end of the dorsal diencephalon and projected many axons to the brainstem, including the 5-HT neurons and DA neurons.. The habenula. complex was divided into two distinct nuclei, the medial (MHb) and the lateral (LHb) habenular nucleus.. Each nucleus had very different afferent and efferent. connections (Geisler et al., 2008). Whereas the MHb contained cholinergic neurons, 3.
(14) and substance P neurons, the LHb contained glutamatergic and only sparse GABAergic neurons (see a review by Lecourtier et al., 2007). The literature review below is focused only in the rodent animals, mainly in considering the rat is the subject used in this study.. In addition, the LHb rather than the MHb is attended because the. former Hb subarea is of particular interest to be manipulated in this study. In the rat, the LHb received GABAergic inputs from the entopeduncular nucleus, the lateral hypothalamus and the lateral preoptic area. the entopeduncular nucleus.. Also, a cholinergic inputted in. A number of studies have shown that the LHb afferent. fibers from the medial frontal cortex, the ventrolateral septum, the diagonal band of. 政 治 大. Broca, the nucleus accumbens, the ventral tegmental area (VTA) and median raphé. 立. The efferent fibers of LHb projected to the raphé nucleus (medial and. 學. ‧ 國. nucleus.. dorsal subareas), the hypothalamus (lateral, dorsomedial and posterior nuclei), VTA,. ‧. the substantia nigra (SN), thalamic nuclei (mediodorsal, central medial, ventromedial. sit. y. Nat. subareas), the parafascicular nucleus, the locus coeruleus, the nucleus accumbens,. io. er. the dorsolateral tegmental nucleus and the supramammillary nucleus (Sutherland, 1982; Geisler et al., 2008). The LHb involved in two neural circuits at least, such as. n. al. 5-HT and DA systems.. Ch. engchi. i Un. v. Despites that the LHb had been studied in flourish by the anatomical approach, how the behavioral function of the LHb remains unclear. Using neurophysiological approach, electrical stimulation of the LHb was reported to inhibit firing of almost all the midbrain DA neurons, up to 97% of tested neurons (Ji and Shepard, 2007). Hikosaka and his colleaugues (2008) demonstrated that LHb neurons encoded the negative reward signaling in contrast to the DA neurons mediating the positive reward process.. Moreover, LHb neurons activity were excited by no-reward-predicting. stimuli, but inhibited by reward-predicting stimuli (Hikosaka et al., 2007). That study also reported that the mesolimbic dopamine neurons exhibited a reverse response 4.
(15) compare with the above phenomenon.. In a more recent study, the LHb neurons are. indicate to represent mirror-reversed phasic effects with DA neurons in the reward-related behavior (Bromberg-Martin et al., 2010a).. Furthermore, it has been. further demonstrated that the LHb is involved in reward prediction error and negative reward processes (Bromberg-Martin et al., 2010b). While a progress has been made to delineate the LHb function by neurophysiological approach at cellular level, behavioral (in vivo) measurement at the system level is now getting attended by conducting behavioral test the free-moving animals with the manipulation in the LHb. Recently, several studies intended to examine how the LHb would affect the behavior. 政 治 大. of reward motivation. Electrical stimulation of the LHb was reported to attenuate the. 立. positive reward-associated reinforcement (Friedman, et al., 2011). And the. ‧ 國. 學. electrolytic lesions of the habenula attenuated brain stimulation reward (Morissette et. ‧. al., 2008). The LHb neurons were activated by no reward responses on. sit. y. Nat. reward-oriented eye movement behavior (Bromberg-Martin et al., 2010a). From In. io. er. these data, the role of LHb involved in the reward motivation is still in vague.. considering that the behavioral tasks used to test the reward motivation are mostly. al. n. iv n C established on the basis of operant h conditioning paradigm, e n g c h i U it is then worthy to challenge a more critical issue regarding to how the LHb affect the operant conditioned behavior.. The LHb and differential reinforcement of low-rate responding behavior Among different types of operant behaviors maintained by distinctive schedules of reinforcement, the present study particularly employing the differential reinforcement of low-rate responding (DRL) behavior for two reasons described below. First, from a previous study done in this laboratory (Chiang, 2006), it was found that lesion of habenula produced impairment on the acquisition of DRL behavior but not 5.
(16) the fixed-interval (FI) typed behavior. In complimentary to the aforementioned data collected from the subjects under LHb lesion, the present study aimed to test how DRL behavior would be influenced by the LHb stimulation.. And, surprisingly, the. electrical stimulation of LHb on DRL behavior had not been investigated.. Second,. the DRL behavior, characterized by self control and timing perception as for its behavioral components (see below), can be used a behavioral task to measuring the motivation and cogitative-like processes simultaneously.. This task is widely used to. screen clinical drugs treated for the psychiatric disorders, for instance, the DRL 72 sec behavioral task applied in testing antidepressant drugs or treatments (O’Donnell et al.,. 政 治 大. 2005). Thus, using DRL behavior task in the present study was aimed to investigate. 立. the role of LHb involved in reward-related motivation or cognitive-like effect by.. ‧ 國. 學. The DRL operant behavior was initially developed from an idea to combine the. ‧. response ratio and time interval schedules (Skinner, 1938). Operant behavior. sit. y. Nat. maintained on the DRL schedule of reinforcement had been characterized as In other words, the. io. er. exhibiting temporal regulation as well as behavioral inhibition.. animal subject trained on this schedule of reinforcement are required to inhibit or. al. n. iv n C withhold lever pressing for a minimum period of time in order to obtain a h especified ngchi U reinforcer. Any premature response leads not only to non-reinforcement consequence but also to re-setting of the time requirement to its full interval. This task has been widely used in psychopharmacology to study the relationship between the drug and behavior (reviews see Liao, 2009; Sanger and Jackson, 1989). Previous studies had shown that acute treatment of amphetamine disrupts DRL behavioral performance by increasing the number of responses and decreasing the number of reinforcers acquired (e.g. Liao and Cheng, 2005; Liao, 2009). A tendency of increasing in burst responding was observed after treatment of amphetamine. Accordingly, amphetamine caused rats to respond more with shorter IRT’s leading to 6.
(17) a leftward shift in the IRT frequency distribution.. And, amphetamine induced leftward. shift is in parallel with that of the saline control, suggesting an internal timing “clock” being speeded up by drug.. All these behavioral changes in the DRL task caused by. amphetamine have been argued to be mediated by drug produced the enhancement of dopamine release in the brain (Liao, 2009). Moreover, the DRL behavioral alteration induced by amphetamine can be partially reversed by DA receptor antagonists (Cheng & Liao, 2007). Therefore, operant behavior maintained on DRL behavior affected by amphetamine is DA dependent. Also, it is the reinforcement contingency that serves a basic component of reward motivation to lead the subject. 政 治 大. performs on a DRL behavior. Following this presumption, it is worthy to elucidate the. 立. neural mechanisms of reward- or DA-related motivation behavior via the examination. ‧ 國. 學. of DRL behavior in the subject with experimental manipulation of a specific brain area. Nat. sit. y. ‧. including the LHb as for a particular interest for this study.. io. er. Aims and the rationale of this study. Based on the literature reviewed above, it was hypothesized that the LHb could. al. n. iv n C play an influential role in the DA dependent behaviors. h e n greward-related chi U. The DRL. behavioral task was used as the major measurement of DA dependent reward-related behaviors. In considering the importance of the interval applied in DRL task, there are two intervals (15 sec and 72 sec) set in the DRL behavioral tasks employed in the stusy, namely a DRL 15-s and a DRL 72-s task.. To encounter the inconsistent. results caused by different parameters of LHb stimulation applied in the previous reports, the first aim of this study tested by two experiments (Experiment 1 and Experiment 2) was set to verify the parameters of intensity, frequency, and duration for LHb stimulation that could effectively alter the locomotor activity or DRL 15-s task. After building a model of DRL 15-s behavioral changed by LHb stimulation, the 7.
(18) second aim of this study as challenged by Experiment 3 was set to verify whether DA was involved such a behavioral alteration by LHb stimulation.. Selective DA D1 and. D2 receptor antagonists (SCH23390 and eticlopride, receptively) were administered in combining with LHb stimulation to examine pharmacologically to see if a DA dependent reversal effect is existed.. The third aim of this study was set to see if a. noradrenergic dependent reversal effect is existed in DRL 15-s behavioral alteration by LHb stimulation.. Experiment 4, thus, was conducted by pharmacological. treatments of norepinephrine (NE) α1, α2 and β receptor antagonists (prazosin, yohimbine and propranolol, respectively) given in combining with the LHb stimulation on DRL 15-s behavior.. 政 治 大. The forth aim of this study was set to examine whether the. 立. DRL behavior affected by LHb stimulation would be depended by the length of interval. ‧ 國. 學. applied in its reinforcement schedule. Accordingly, Experiment 5 tested the effects of Finally, regarding to the fifth aim, the. ‧. LHb stimulation on a DRL 72-s behavioral task.. sit. y. Nat. effects of LHb stimulation were tested in discrimination task which required the rat’s. io. er. ability to differentiate the different magnitudes of reward in a two-lever operant chamber (Experiment 6). Together, the results of this study were expected to answer. al. n. iv n C how the LHb is involved in the reward-related as measured by DRL h e n g cmotivation hi U behavior in the subject with the activated LHb by electrical stimulation.. 8.
(19) Materials and methods Subjects Male (300~350 g) Wistar rats were the subjects obtain from the BioLACO Taiwan Co.,Ltd.. All animals were housed individually and allowed ad libitum access to food. and water. The colony was maintained on a 12-hour light / dark cycle with lights on 08:00 AM, where the room temperature was kept at 22 ± 2 °C. The subjects were treated by a water restriction regimen before the DRL behavioral experiment, and a food restriction regimen before a discrimination behavioral experiment. The water. 政 治 大 The food restriction was conducted by gradually reduced the daily food 立. restriction was conducted by gradually reducing the daily access of water to 5 minutes every day.. ‧ 國. 學. intake of 15 g lab chow, which led the subjects body weight remained in about 85 % of free-feeding of body weight.. All experiments were conducted following the regulation. ‧. by the animal use and care committee of National Cheng-Chi University.. sit. n. al. er. io. DRL operant behavior. y. Nat. Apparatus. Ch. engchi. i Un. v. Six operant conditioning chambers (Med-Associates, Inc.; St Albans Vermont) were used.. Each chamber (30 cm х 20 cm х 25 cm) has one press lever, one house. light and a liquid dispenser as controlled by solenoid value. The chamber’s floor was formed by 18 stainless steels (5 mm of diameter) separated in 11 mm. The liquid dispenser supplied the water reinforcer based on behavioral contingency set up in each experiment, and each reinforcement supply was 0.04 ml of tab water. Each chamber was placed in a separate wooden soundproof box with a fan to provide ventilation and white noise.. All chambers were connected to a microcomputer that. controlled the behavioral program and data collection. The raw data were collected 9.
(20) from the inter-responses times (IRT), each generated from a lever press. The raw data were then reduced and calculated into seven variables including total responses, reinforced responses, non-reinforced responses, burst responses, peak rate, peak time and modified response efficiency (MRE). See Cheng and Liao (2007) for additional details.. Locomotor activity Four black acrylic boxes (45 x 45 x 36 cm each) were assembled and used to measure the locomotor activity.. A charge coupled device (CCD) camera was set up. 政 治 大. 52 cm high from the bottom and located in the center of the four-box assembly.. 立. (mm) that a subject traveled in each of the four boxes.. 學. ‧ 國. Controlled by a desk-top computer, the CCD simultaneously recorded the distance. ‧ sit. y. Nat. DRL behavioral training. io. er. After manually shaping, the water-deprived rat first learned to press the lever for water reward under a fixed-ratio 1 (FR-1) schedule. When the rat pressed the lever. al. n. iv n C over 40 times in one daily session of the DRL schedule of reinforcement was h30 e nmin, gchi U then introduced.. In which, a reinforcer was delivered contingent upon a lever press if. the time had elapsed since the previous press over the DRL interval set up. Premature responses led to a non-reinforcement contingency and a resetting of the interval delay, as indexed by a non-reinforced response. reinforced or not, reset the delay timer.. Each lever press, whether. Two intervals, 15 sec and 72 sec, were. chosen for the DRL behaviors applied in the present study.. A total of five groups of. the rats received the DRL behavioral training, in which four groups were trained on the DRL 15-s task and one group was trained on the DRL 72-s task (see details below). For the DRL 15-s behavioral training, after the stabilized performance on FR-1 10.
(21) schedule, the rat was directly subjected to respond for the DRL 15-sec schedule of reinforcement. The subjects reached a steady baseline of DRL 15-s after 25 training sessions (each of 30 min). A criteria to determine the stable baseline level of DRL 15-s behavioral performance was referred by the MRE ratio equal or greater than 0.45 (Cheng et al., 2008).. In the DRL 72-s behavioral training, after the initial lever-press. training, the subjects were trained on a DRL 18-s schedule for eleven sessions (each of 60 min).. Then, the interval of schedule requirement was increased to 72 sec.. The subject was run in DRL 72-sec behavioral training for 55 days to reach a stable baseline level. However, to avoid the potential of extinction on operant responding,. 政 治 大. in the DRL 72-s training, the subject was run with longer daily sessions from the. 立. beginning. Such that, within the 55 days of DRL 72-sec behavioral training, the 3 hr. ‧ 國. 學. training session run for the first 3 days, 2 hr training session run for the next 14 days, The criterion for the. ‧. and the regular 1 hr training session run for the rest of 38 days.. sit. y. Nat. definition of a stable baseline for DRL 72-s task was determined by both the total. io. er. responses less than 70 and the reinforced responses greater than 12, which performance was also less than 10% variation in the response rate for three. al. n. iv n C consecutive sessions (Zhang et al.,h 2006). The manipulation of LHb stimulation and engchi U pharmacological treatment was only conducted in the rat with a stable performance on either DRL 15-s or DRL 72-s behavioral task.. Discrimination task A behavioral testing of simple discrimination of different amount of reward was conducted in four operant chambers, each chamber (30.5 x 24 x 21 cm) enclosed in sound-attenuating boxes (Med-Associates, St. Albans, VT., USA). Each box was equipped with a fan to provide ventilation and to mask extraneous noise. operant chamber had two retractable levers on the front wall. 11. Each. The holes where the.
(22) levers extended were symmetrically 1 cm from the left and right side walls, and 10 cm from the floor.. The food reinforcement (45 mg; Bioserv, Frenchtown, NJ) was. delivered via a pellet dispenser to a food receptacle (2 x 2.5 cm) set in the middle of two levers and 6.5 cm above the floor.. The operant chamber was illuminated by a. single 100 mA house light located in the top-center of the wall opposite the levers. The lever choices and reaction latency were recorded by a desk-top computer connected to the chambers via an interface.. Surgery. 政 治 大. Before a surgery conducted in a stereotaxic instrument (Stoelting Co.), the rat. 立. was anesthetized by intraperitoneal injection (i.p.) of Zoletil 50 (Virbac, Carros, France). ‧ 國. 學. in a dose of 1 mg/kg. The subject was implanted with the homemade electrode.. ‧. The electrode was implanted into the left lateral habenula in coordinated from the. sit. y. Nat. bregma, AP = -3.6 mm, ML = +0.7 mm, and DV = -4.8 mm (Paxinos & Watson, 2007). After the electrode placement, the electrode was fixed to the skull with acrylic dental. er. io. post-surgery with a l Penicillin (20,000IU) wasiinjected v n C(i.m.) 0.2 ml and intramuscular injection to reduce the potential occurrence of wound U hen i h gc. n. cement to secure its patency.. infection. The subject was allowed one week to recover from surgery.. Electrode preparation and habenula stimulation The electrode was a bipolar self-designed stimulating electrode with two stainless steel wires (A-M System, Sequim, WA, USA) in a guide cannula (0.33 mm inner diameter, 0.63 mm outer diameter and 10 mm length). The stainless steel wire was 0.02 mm inter diameter for the stainless steel itself and 0.045 mm outer diameter as measured with the coated insulating materials.. The stainless steel wires were. extended 1 mm from the one end of guide cannula, and where the other end was 12.
(23) welded into a two channel bases as for connecting the electrical wires from the stimulator. The electrode was implanted into the lateral habenula of the left hemisphere for all the subjects received the lateral habenula stimulation. The subjects, thus, received habenula stimulation unilaterally.. And, the lateral habenula stimulation was. given for 15 min each time and conducted right before the behavioral session.. The. parameters of LHb stimulation current were tested for 0.05 mA and 0.1 mA with a 0.5 ms spike-duration. The stimulation frequencies were manipulated at 10 Hz and 100 Hz.. A diagram illustrating these parameters is shown Fig 1C.. 立. Drugs. 政 治 大. ‧ 國. 學. D-amphetamine sulfate (Sigma-Aldrich, St. Louis, USA), SCH23390. ‧. hydrochloride (Tocris, Ellisville, MO, USA), eticlopride hydrochloride (Tocris, Ellisville,. y. sit. dissolved in 0.9 % saline.. Prazosin hydrochloride (Research Biochemicals. io. er. Nat. MO, USA) and yohimbine hydrochloride (Sigma-Aldrich, St. Louis, USA) were. International, RBI, Natick, MA, USA) and propranolol hydrochloride (Sigma-Aldrich, St.. al. n. iv n C Louis, USA) were dissolved in distilled All drugs were administered by h ewater. ngchi U. intraperitoneal injection in a volume of 1 ml/kg, and the injection was conducted at 20 min before behavioral measurement.. Procedures Experiment 1: the effects of LHb stimulation on locomotor activity The rat was gently handled by experimenter for two weeks before receiving the locomotor activity tests and the LHb stimulation.. A group of the rats (n = 12) was. repeatedly used to test the locomotor activity conducted at 0, 10 and 20 min after the end of LHb stimulation. These subjects were further divided into three subgroups (n 13.
(24) = 4 each) which received a specific treatment of stimulation current intensity: 0 mA, 0.05 mA or 0.1 mA. These three tests were conducted every other day.. The. subjects were then remained in the colony for one week, during which a daily 15 min of experimenter’s holding was given.. Subsequently, a dose effect of amphetamine. on locomotor activity was evaluated.. For drug dosing treatment, the rat was. repeatedly injected amphetamine at doses of 0, 1, and 2 mg/kg under a Latin square design.. Three injection days were administered every other day. The locomotor. activity was conducted for 15 min after injection. The data of each locomotor test were collected in three 5 min blocks that offered the within-session analysis.. 政 治 大. Experiment 2: the effects of LHb stimulation on DRL 15-s behavior. 立. A group of rats (n = 12) was initially recruited and used to test the effects of LHb. ‧ 國. 學. stimulation on DRL 15-s behavior. When the stable maintenance levels were. ‧. reached, the subject received a surgery of electrode implantation. Following the. sit. y. Nat. surgical recovery, the rats were re-trained on DRL 15-s behavior for one week. The. io. er. rats were then subjected to test the effects of LHb stimulation on DRL 15-s behavior. The parameters for electrical stimulation frequencies (0, 10 and 100 Hz) were. al. n. iv n C manipulated and each test given in h a separate day. UThe aforementioned treatments engchi were conducted in a sequential order of the sham stimulation (sham S), LHb stimulation given at 10 Hz (LHb S 10 Hz) and LHb stimulation given at 100 Hz (LHb S 100 Hz).. Behavioral data completely collected after a successful LHb stimulation. were obtained by 12 subjects for 0 Hz treatment, 9 subjects for 10 Hz treatment and 9 subjects for 100 Hz treatment.. Missing data were due to the incompleted LHb. stimulation caused by the electrode implantation being damaged or dropped. The DRL 15-s test session lasted for 15 min.. On the day after the LHb stimulation test,. the rats were only run in DRL 15-s task without the manipulation of electrical stimulation. 14.
(25) Experiment 3: the effects of DA antagonists on DRL 15-s behavioral alterations induced by LHb stimulation A group of rats (n = 12) was initially recruited and used to test whether DA antagonists would reverse DRL 15-s behavioral alterations induced by LHb stimulation or not. The subjects with the implanted electrode were ensured with a stable performance on DRL 15-s task before the pharmacological tests.. Referred by. the results from the experiments described above, the parameters of the LHb stimulation conduced in this experiment were set by 900 sec of duration, 100 Hz of frequency, and 0.05 mA of intensity.. Doses tested for D1 and D2 receptor. 政 治 大. antagonists, respectively, were 0, 0.003 and 0.01 mg/kg of SCH23390 and 0, 0.01,. 立. and 0.03 mg/kg of eticlopride. Each rat received an injection prior to a stimulation. ‧ 國. 學. treatment in an experimental day, which was followed by a solely DRL 15-s re-running Accordingly, under a within-subject design applied in this experiment,. ‧. daily session.. y. Nat. the rat was repeatedly tested with six treatments including 1) saline vehicle with sham. er. io. sit. stimulation initialized as “veh + sham S”, 2) saline vehicle with LHb stimulation initialized as “veh + LHb S”, 3) SCH23390 of 0.003 mg/kg with LHb stimulation. al. n. iv n C initialized as “SCH 0.003 + LHb S”, h 4) SCH23390 of U e n g c h i 0.01 mg/kg with a LHb. stimulation initialized as “SCH 0.01 + LHb S”, 5) eticlopride of 0.01 mg/kg with LHb stimulation initialized as “eti 0.01 + LHb S”, and 6) eticlopride of 0.03 mg/kg with LHb stimulation initialized as “eti 0.03 + LHb S”. Behavioral data completely collected after a successful LHb stimulation were obtained by 12, 8, 8, 6, 8 and 6 rats for the aforementioned treatments, correspondingly.. And, the rat was tested DRL 15-s for. 15 min immediately after the completion of each of aforementioned treatments. In order to better elucidating the interaction between DA drugs and LHb stimulation, a drug treatment with sham stimulation should be taken into account with the treatments described above.. An additional experiment was conducted, by 15.
(26) recruiting naïve rats (n = 12), to examine the dose effects of SCH23390 and eticlopride on DRL 15-s behavior.. The experimental protocols of DRL 15-s training. and electrode implantation were same as those described above.. The rats then. received four treatments in sequence: 1) saline vehicle with sham stimulation initialized as “veh + sham S”, 2) saline vehicle with LHb stimulation initialized as “veh + LHb S”, 3) SCH23390 of 0.01 mg/kg with sham stimulation initialized as “SCH 0.01 + sham S”, and 4) eticlopride of 0.03 mg/kg with sham stimulation initialized as “eti 0.03 + sham S”. Behavioral data completely collected after a successful treatments were obtained by 12, 8, 7 and 7 rats for the aforementioned treatments, correspondingly.. 立. 政 治 大. Experiment 4: the effects of NE antagonists on DRL 15-s behavioral alterations. ‧ 國. 學. induced by LHb stimulation. ‧. A group of rats (n = 12) was initially recruited and used to test the effects of NE The. sit. y. Nat. antagonists on DRL 15-s behavioral alterations induced by LHb stimulation.. io. er. experimental design and protocols applied in this experiment were similar to those described in Experiment 3, except selective NE receptor agonists evaluated in this. al. n. iv n C The dosing treatments hincluded i Uand1 mg/kg for prazosin, 0, 0.5, e n g c0,h0.5,. experiment.. and1 mg/kg for yohimbine, and 0, 10, 20 mg/kg for propranolol. Thus, the rat was repeatedly tested with eight treatments including 1) vehicle with sham stimulation initialized as “veh + sham S”, 2) vehicle with LHb stimulation initialized as “veh + LHb S”, 3) prazosin of 0.5 mg/kg with LHb stimulation initialized as “pra 0.5 + LHb S”, 4) prazosin of 1 mg/kg with a LHb stimulation initialized as “pra 1.0 + LHb S”, 5) yohimbine of 0.5 mg/kg with LHb stimulation initialized as “yoh 0.5 + LHb S”, 6) yohimbine of 0.1 mg/kg with LHb stimulation initialized as “yoh 1.0 + LHb S”, 7) propranolol of 10 mg/kg with LHb stimulation initialized as “pro 10 + LHb S” and 8) propranolol of 20 mg/kg with LHb stimulation initialized as “pro 20 + LHb S”. 16.
(27) Behavioral data completely collected after a successful LHb stimulation were obtained by 12, 8, 8, 7, 7, 7, 7 and 7 rats for the aforementioned treatments, correspondingly. Experiment 5: the effects of LHb stimulation on DRL 72-s behavior A group of rats (n = 10) was used to test if LHb stimulation would affect operant behavior maintained on a DRL 72-s schedule.. The rats had initially trained in DRL. 72-s task and were run through a series of drug dosing tests (SNC80, amphetamine, GBR12909 and fluoxetine) for another project of this lab. To begin this experiment, the subjects were run in DRL 72-s for 48 sessions to ensure their stable performance on this behavioral task. Subsequently, the rat was implanted with the electrode. 政 治 The大subject was re-run in DRL 72-s. aimed to the left LHb via the stereotaxic surgery.. 立. LHb stimulation was the same as those described above.. The protocols set for. 學. ‧ 國. task for 48 sessions before the LHb stimulation manipulated.. ‧. Experiment 6: the effects of LHb stimulation on discrimination task. y. Nat. A group of rats (n = 4) was used to test whether the LHb stimulation could affect. er. io. sit. the rat’s ability to discriminate different magnitudes of reward in a discrimination task run in a two-lever operant chamber. The procedure of discrimination task consisted. al. n. iv n C of four blocks of 12 trials, two forcedhchoice trials and e n g c h i Uten free choice trials for each block.. In which, a single response made on one of the two levers immediately led to. a delivery of four pellets, whereas a press of the other lever caused a delivery of one pellet. The subject ran through this discrimination procedure in about 60 min daily. The subject reached the baseline of discriminate task by making 85 % of choices of large reward after one week training.. Subsequently, the rats were received LHb. stimulation with different current (0.05 mA and 0.1 mA) and tested by discrimination task in every other day.. Histology 17.
(28) At the conclusion of the experiments, the rat was anesthetized and transcardially perfused with PBS followed by 24 % paraformaldehyde. The brain was removed and immersed in preservation solution (100 ml 24 % paraformaldehyde, 165 ml distilled water, 12.2 g sucrose, 1 tablet of phosphate buffered saline) for 48 hours. The brain was then frozen on dry ice and slices (40 µm sections) using a cryostat microtome. Sections were mounted on glass slides (coated with 0.5 % gelatin), stained with Cresyl violet and subsequently examined under a microscope to verify the placement of the electrode.. DRL behavioral data. 立. 政 治 大. The DRL behavioral data were based on the IRT’s collected from each session. ‧ 國. 學. for each rat. In addition to categorizing the IRT’s into the total responses, reinforced. The IRT data of DRL 15-s behavior. sit. y. Nat. for both qualitative and quantitative analyses.. ‧. responses, and non-reinforced responses, a plot of IRT distribution curve was made. io. er. were plotted into a distribution with response frequencies for 30 consecutive 1 sec time bins, while the IRT data of DRL 72-s behavior would be plotted in to a distribution. al. n. iv n C with response frequencies for 144 consecutive 1 secUtime bins. hengch i. A bimodal IRT. distribution was reliably shown at the baseline stage as well as the control condition. Quantitative analyses of the IRT distribution included burst responses, peak time, peak rate, and MRE ratio.. The burst responses were the summed frequencies of. IRT less than 2 sec. The peak rate and peak time were calculated from the de-burst IRT’s, in which a moving average based on four consecutive 1 sec bins was applied to smooth the distribution. With the maximum frequencies of a 4 sec epoch identified, the peak time was the mean value of the IRT’s that fell within those four bins. The peak rate was calculated by the summed frequencies of those four bins divided by four. The MRE ratio, indexed the response efficiency, was calculated from 18.
(29) reinforced responses divided by de-burst responses: [number of reinforced responses ÷ (number of total responses – number of burst responses)].. Statistical analyses The data for each dependent variable were subjected to analyses of variance (ANOVA) following the experimental design used. the mean with the standard error (mean ± sem).. All the results are presented as Locomotor activity data of. Experiment 1 were analyzed by a three-way ANOVA for the factors of stimulation current, blocks and time.. In which, the stimulation current was on a between-subject. 政 治 大. design, while the blocks and time were the within-subject repeated measures.. 立. DRL. behavioral data collected from Experiment 2 to Experiment 5 were analyzed by a. ‧ 國. 學. one-way repeated ANOVA on each of dependent variables including the total. ‧. responses, reinforced responses, non-reinforced responses, burst responses, peak. sit. y. Nat. rate, peak time, and MRE ratio. The data of discrimination behavior in Experiment 7. io. er. were first analyzed by a one-way repeated ANOVA on the stimulation frequency. And, a two-way repeated ANOVA was further conduced for a within-session analysis. al. n. iv n C on the factors of stimulation frequency block. Statistical significance was h eand ngchi U. determined by p < 0.05. When the main effect was significantly yielded from ANOVA, the post hoc test was run by the use of Scheffe's method.. Simple main effect. comparisons were conducted when a significant interaction was revealed.. Statistical. analyses were conducted using commercial software (Statistica version 5.5, Statsoft, Tulsa, OK, USA).. 19.
(30) Results Experiment 1: the effects of LHb stimulation on locomotor activity Figure 3. shows the effects of electrical stimulation in LHb on locomotor activity. The results of a three-way ANOVA only revealed a significant main effects of block, F(2, 81). = 94.5, p < 0.001. Post hoc tests following the block main effect confirmed that. traveling distance in first block was significantly longer than that of the second and third blocks (both p < 0.001). And, a significant difference was detected in between the second and third blocks (p < 0.001).. 政 治 大 The results of a two-way ANOVA revealed significant main effects of dose 立. Figure 4. shows the dose effects of amphetamine on the distance of locomotor activity.. ‧ 國. interaction.. 學. (F(2, 99) = 15.98, p < 0.001) and block (F(2, 99) = 54.14, p < 0.001), but no significant Post hoc tests following the dose main effect revealed that traveling. ‧. distance was significantly increased by given at 1 mg/kg and 2 mg/kg (both p < 0.001). y. sit. n. al. er. Post hoc tests following the block main effect confirmed that traveling. io. mg/kg.. Nat. treatments, but no significant difference in between treatments of 1 mg/kg and 2. i Un. v. distance in first block was significantly longer than that of the second and third blocks. Ch. engchi. (both p < 0.001), but no significant difference in between the second and third blocks.. Experiment 2: the effects of LHb stimulation on DRL 15-s behavior Figure 5. presents the effects of LHb stimulation on DRL 15-s behavior, as measured by total responses, reinforced responses, non-reinforced responses, burst responses, peak time and peak rate.. LHb stimulation increased the total responses. only in a marginal significance, F(2, 27) = 3.26, p = 0.053 (Fig. 5A). LHb stimulation significantly decreased the reinforced responses, F(2, 27) = 5.98, p < 0.01 (Fig. 5B). Post hoc tests revealed the significant decreases of reinforced responses by low 20.
(31) frequency (p < 0.05) and high frequency (p < 0.01) treatments.. LHb stimulation. significantly increased the non-reinforced responses, F(2, 27) = 5.41, p < 0.01 (Fig. 5C). Post hoc tests showed that the non-reinforced responses were significantly increased by low frequency (p < 0.05) and high frequency (p < 0.001) treatments. LHb stimulation produced a marginally significant increment on the burst responses, F(2, 27) = 3.20, p = 0.056 (Fig. 5D). LHb stimulation significantly decreased the peak time, F(2, 27) = 6.11, p < 0.01 (Fig. 5E). Post hoc tests showed that the peak time was significantly decreased by either low frequency (p < 0.05) or high frequency (p < 0.01) treatment.. In contrast to the five dependent variables described above, the peak rate. 政 治 大. was not significantly affected by LHb stimulation (Fig. 5F).. 立. behavior, and the IRT distribution curves are plotted.. 學. ‧ 國. Figure 6. presents the effects of LHb stimulation on the MRE ratio of DRL-15 LHb stimulation significantly. ‧. decreased the MRE ratio, F(2, 27) = 6.71, p < 0.01 (Fig. 6A). Post hoc tests showed. The electrical stimulation of LHb produced a. io. leftward shift on the IRT distribution (Fig. 6B).. n. al. Ch. engchi. er. frequency (p < 0.001) treatment.. sit. y. Nat. that MRE ratio was significantly decreased by either low frequency (p < 0.05) or high. i Un. v. Experiment 3: the effects of DA antagonists on DRL 15-s behavioral alterations induced by LHb stimulation Figure 7. presents the effects of SCH23390 and eticlopride on the alteration of DRL 15-s behavior induced by LHb stimulation, as measured by the six dependent variables.. The results of ANOVA revealed that co-adminstration drugs with LHb. stimulation significantly affected the total responses, F(5, 41) = 4.22, p < 0.01 (Fig. 7A), non-reinforced responses, F(5, 41) = 3.67, p < 0.01 (Fig. 7C) and burst responses, F(5, 41) = 4.27, p < 0.01 (Fig. 7D). In comparing to the control, post hoc tests on total responses revealed a significant increment by LHb stimulation (p < 0.01). Such an 21.
(32) effect was significantly reversed by the higher dose of SCH23390 or eticlopride, whereas no reversal effect was detected for the lower dose of either drug.. The. results of post hoc comparisons on non-reinforced responses and burst responses were similar to those of total responses described above.. In contrast to the three. dependent variables described above, the reinforced responses, peak time and peak rate were not significantly affected by co-adminstration drugs with LHb stimulation. Figure 8. presents the effects of SCH23390 and eticlopride on the alteration of DRL 15-s behavior induced by LHb stimulation as measured by MRE ratio, and the IRT distribution curves are plotted. The results of ANOVA revealed that. 政 治 大. co-adminstration drugs with LHb stimulation significantly affected the MRE ratio, F(5, 41). 立. = 2.68, p < 0.05 (Fig. 8A). In comparing to the control, post hoc tests revealed a. ‧ 國. 學. significant decrement on the MRE ratios by LHb stimulation (p < 0.05). Such an. ‧. effect was significantly reversed by the higher dose of SCH23390 or eticlopride,. sit. y. Nat. whereas no reversal effect was detected for the lower dose of either drug.. io. er. Figure 9. presents the effects of SCH23390 and eticlopride on DRL 15-s behavior, as measured by the six dependent variables. The results of ANOVA revealed that. al. n. iv n C experimental treatments of drug administrations h e n g c hand i ULHb stimulation significantly affected the total responses, F(3, 28) = 6.45, p < 0.01 (Fig.9A), non-reinforced. responses, F(3, 28) = 6.77, p < 0.01 (Fig.9C), burst responses, F(3, 28) = 7.07, p < 0.01 (Fig.9D) and peak rate, F(3, 28) = 3.52, p < 0.05 (Fig.9F). In comparing to the control, post hoc tests on total responses revealed a significant increment by LHb stimulation (p < 0.01). Such an effect was not significantly detected for the dose treatment of SCH23390 or eticlopride. The results of post hoc comparisons on non-reinforced responses and burst responses were similar to those of total responses described above.. Post hoc tests on peak rate revealed a significant increment by SCH23390 (p. < 0.05), and no such an effect for LHb stimulation or eticlopride. 22. In contrast to those.
(33) four dependent variables described above, the measures of reinforced responses and peak time were not significantly affected by drug treatments or LHb stimulation (Fig.9B and 9E). Figure 10. presents the effects of SCH23390 and eticlopride on DRL 15-s behavior as measured by MRE ratio, and IRT distribution curves are plotted. The results of ANOVA revealed a significant treatment effect on the MRE ratio, F(3, 28) = 4.44, p < 0.05 (Fig.10A). Post hoc tests revealed only a significant decrement by LHb stimulation (p < 0.05).. 政 治 大. Experiment 4: the effects of NE antagonists on DRL 15-s behavioral alterations. 立. induced by LHb stimulation. ‧ 國. 學. Figure 11. presents the effects of prazosin, yohimbine and propranolol on the. ‧. alteration of DRL 15-s behavior induced by LHb stimulation, as measured by the six. sit. y. Nat. dependent variables. The results of ANOVA revealed that co-adminstration drugs. io. er. with LHb stimulation significantly affected the total responses, F(7, 55) = 7.63, p < 0.001 (Fig.11A), reinforced responses, F(7, 55) = 2.75, p < 0.05 (Fig.11B), non-reinforced. al. n. iv n C responses, F(7, 55) = 6.13, p < 0.001 h (Fig.11C), burst U e n g c h i responses, F(7, 55) = 5.94, p < 0.001 (Fig.11D) and peak rate, F(7, 55) = 4.50, p < 0.001 (Fig.11F). In comparing to the control, post hoc tests on total responses revealed a significant increment by LHb stimulation (p < 0.01). Such an effect was significantly reversed by prazosin and propranolol given at the higher dose of, but not for the lower dose of either drug.. No. significant reversal effect was produced by yohimbine. The results of post hoc comparisons on non-reinforced responses were similar to those of total responses described above.. Regarding to the reinforced responses, post hoc tests revealed. only a significant decrement by the both doses of yohimbine given in combining with LHb stimulation.. Post hoc tests on burst responses showed a significant increment 23.
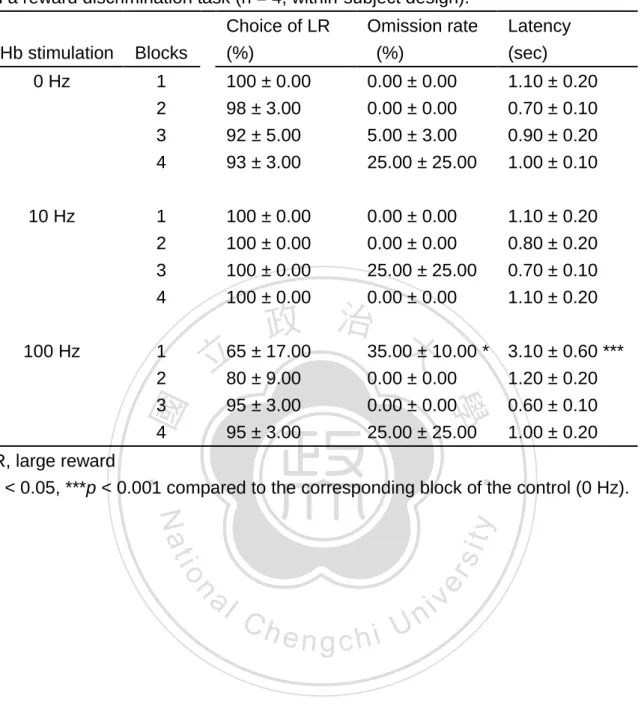
(34) by LHb stimulation (p < 0.001). Such an effect was reversed by prazosin and propranolol, but not by yohimbine. Post hoc tests on peak rate revealed a significant increment by the both doses of prazosin (p < 0.05) and yohimbine (p < 0.001) with LHb stimulation. Figure 12. presents the effects of prazosin, yohimbine and propranolol on the alteration of DRL15-s behavior induced by LHb stimulation as measured by MRE ratio, and IRT plots are shwon.. The results of ANOVA revealed a significant treatment. effect on the MRE ratio, F(7, 55) = 5.26, p < 0.001 (Fig.12A). Post hoc tests on the MRE ratio revealed a significant increment by LHb stimulation (p < 0.05). Such an. 政 治 大. effect was not significantly reversed by any drug treatment back to the control level.. 立. ‧ 國. 學. Experiment 5: the effects of LHb stimulation on DRL 72-s behavior. Paired-samples t-test on the burst. sit. y. Nat. dependent variables is presented in Table 1.. ‧. The effects of LHb stimulation on the DRL 72-s behavior as measured by the six. io. er. responses revealed a significant increment by LHb stimulation (p < 0.05). On peak time revealed a significant decrement by LHb stimulation (p < 0.05). None of other. al. n. iv n C behavioral measurements was significantly by LHb stimulation (Table 1). h e n gaffected chi U. Experiment 6: the effects of LHb stimulation on a reward discrimination task Figure 13. presents the effects of electrical stimulation in LHb on a reward discrimination task, as measured by choice of large reward, omission rate and response latency.. The results of ANOVA revealed that LHb stimulation significantly. affected the choice of large reward, F(2, 45) = 6.90, p < 0.01 (Fig.13A), omission rate, F(2, 45) = 3.20, p < 0.05 (Fig.13B) and response latency, F(2, 458) = 4.44, p < 0.05 (Fig.13C). In comparing to the control, post hoc tests that the higher, but not the low, frequency of LHb stimulation significantly decreased the choice of large reward and 24.
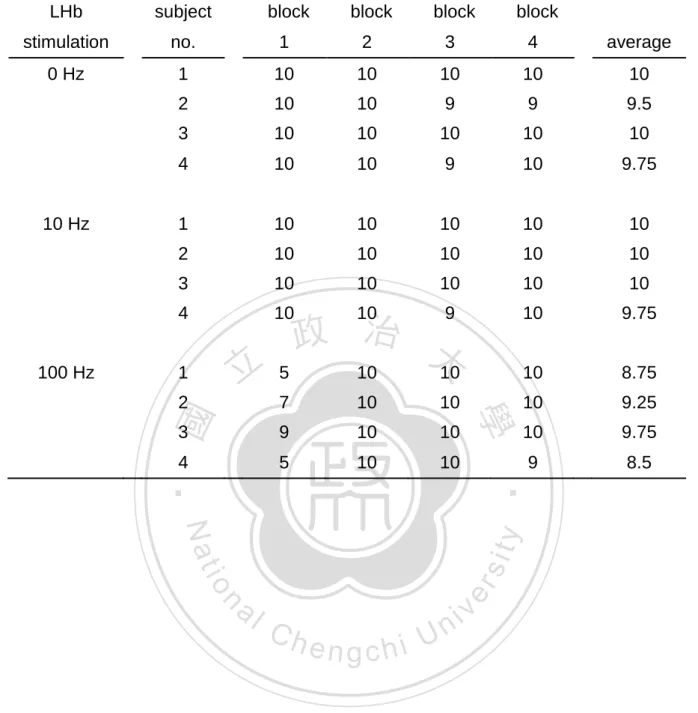
(35) increased the omission rate and response latency (all p < 0.05). Within-session analyses of the effects of lateral habenula (LHb) stimulation on a reward discrimination task are presented in Table 2.. Regarding to the choice of large. reward, the results of a two-way ANOVA revealed a significant main effect of frequency, F(2, 36) = 9.52, p < 0.01, and a significant frequency-by-block interaction, F(6, 36). = 2.49, p < 0.05. Post hoc tests following the frequency main effect revealed only. a significantly decrement by the high frequency treatment (p < 0.05). From simple main effect comparisons, the trend of a lower choice of large reward rate under the 100 Hz LHb stimulation was not significantly confirmed over all four blocks in. 政 治 大. comparing to the corresponding blocks of the control (p > 0.05). Regarding the. 立. 學. ‧ 國. omission rate, the results of a two-way ANOVA significantly yielded main effects of frequency (F(2, 36) = 8.6, p < 0.05) and block (F(3, 36) = 10.4, p < 0.01) and its interaction, Post hoc tests following the frequency main effect were. ‧. F(6, 36) = 7.43, p < 0.001.. sit. y. Nat. resulted by the significant increment by high frequency treatment (p < 0.05). Post. io. er. hoc tests following the block main effect confirmed that the omission rate in the first block was significantly higher than those in the second (p < 0.001), the third (p < 0.05). al. n. iv n C and the fourth (p < 0.01) blocks. Simple effect comparisons revealed a h e nmain gchi U. significant increment in the first block under high frequency treatment (p < 0.05), but no such an increment occurred in the other three blocks.. In terms of the response. latency, the results of a two-way ANOVA revealed significant main effects of the frequency, F(2, 449) = 11.49, p < 0.05, and the block, F(3, 449) = 14.93, p < 0.001. And the frequency-by-block interaction was significantly confirmed, F(6, 449) = 7.43, p < 0.001.. In comparing to the control, post hoc tests on the frequency factor revealed a. significantly increment by the high frequency treatment (p < 0.05). Post hoc tests following the block main effect confirmed that the response latency in the first block was significantly more than the second (p < 0.001), the third (p < 0.001) and the fourth 25.
(36) (p < 0.01) blocks,. In comparing to the control, simple main effect on response. latency revealed a significant increment by the first block in high frequency treatment (p < 0.001). In addition to the first block in high frequency treatment, there was no any significantly affected between frequencies and blocks. (The results of post hoc tests and simple main effect comparisons for the measure of response latency revealed a similar pattern to those described on the omission rate.). 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. 26. i Un. v.
(37) Discussion The present study mainly investigated whether the LHb stimulation would affect operant behavior maintained on DRL schedule of reinforcement.. Experiment 1. showed that the LHb stimulation as manipulated by two currents (mA) and tested at three different time points after stimulation did not affect locomotor activity.. The. lower current of LHb stimulation given immediately before behavioral test was then set for the follow-up experiments.. Two frequencies (Hz) of LHb stimulation was. tested on DRL 15-s behavior in Experiment 2, which found a significant Such a 政 治 大 behavioral change was pharmacologically reversed by SCH23390 and eticlopride as 立 frequency-dependent effect of LHb stimulation on DRL 15-s behavior.. ‧ 國. In Experiment 4, prazosin or propranolol. 學. revealed by the results of Experiment 3.. was shown to produce a similar, but subtle, reversal effect on LHb stimulation induced. ‧. DRL 15-s behavioral changes.. In Experiment 5, the LHb stimulation was. sit. y. Nat. demonstrated to affect a DRL 72-s behavior, but with less degree of influence (on two. n. al. er. io. out of seven behavior measurements) as compared to that of DRL 15-s task.. i Un. v. Experiment 6 confirmed that the LHb stimulation could affect the ability of. Ch. engchi. discrimination, which effectiveness was significant only in the first block (out of four).. No effects on locomotor activity by LHb stimulation In the first part of Experiment 1, the LHb stimulation did not affect on locomotor activity.. This part of results was in consistent to a previous report by Friedman and. associates (2010) showing that the traveling distance produced by LHb stimulation rat was not different from that by sham-operated rat.. However, there was a report. showing the locomotion was affected by LHb stimulation. The locomotor activity was increased in an animal model of depression by LHb stimulation given in chronic, 30 27.
(38) min every day for 28 days (Meng et al., 2011).. In that study, the rat treated by that. repeatedly exposed to a set of chronic mild stressors for 4 consecutive weeks was then subjected under a animal model of depression.. In addition, the LHb stimulation. was conducted in a long-term fashion before behavioral testing.. Thus, the difference. between these two studies and in comparing the present study, in terms of the effects of LHb stimulation on locomotion, could be attributed to different experimental protocols used among these studies.. A question may be raised in concerning the. negative results of LHb stimulation on locomotion in this study.. That is, it might be. due to the subjects were insensitive to any experimental treatment that is related to. 政 治 大. brain DA. Accordingly, the same rats were tested in the second part of Experiment 1,. 立. which results clearly showed those subjects were significantly affected by. ‧ 國. 學. amphetamine. The acute injection of amphetamine increased the locomotor activity. ‧. is a well established model to test the general motor function modulated by brain DA. sit. y. Nat. (e.g. Cole, 1978). Combining the results from the first and second parts of. io. er. Experiment 1, it is indicated that the subjects could be sensitively affected by drug treatment agonizing brain DA systems to increase locomotion. Thus, the negative. al. n. iv n C results of LHb stimulation applied inhthis experiment U e n g c h i implied that the intensities of LHb stimulation applied in this study would not affect the general motor function. However, whether the DA level was altered by the LHb stimulation or not has not been examined in this study, which is essential before making a conclusive remark.. DRL 15-s behavior affected by LHb stimulation Experiment 2 showed that the LHb stimulation significantly affected DRL 15-s behavior in a frequency-dependent manner.. Furthermore, these behavioral changes. by LHb stimulation are similar to those induced amphetamine on DRL behavior with interval set in a range of 10 to 20 sec (Liao, 2009). A previous study of this lab 28.
(39) showed that amphetamine increased the total responses, non-reinforced responses and burst responses, but decreased the reinforced responses and peak time (Cheng and Liao, 2007). With the similarity of DRL 15-s behavioral effects produced by the LHb stimulation and amphetamine treatment, there might be a common mechanism was shared for these two treatments.. In terms of neural substrates, with. amphetamine pharmacologically acting as a DA agonist, it was then inferred that the behavioral alterations induced by LHb stimulation was modulated by DA related mechanisms. This was a rationale to carry out Experiment 3 and Experiment 4 (see the relevant discussion below).. 政 治 大. The unilateral, but not bilateral, LHb stimulation was applied in the present and. 立. induced significant behavioral changes on DRL 15-s behavior.. A question may be. ‧ 國. 學. asked: how did the unilateral LHb stimulation on one side of the brain adequately A recent study addressed a strong. ‧. affect the Hb and then change the behavior?. sit. y. Nat. connectivity between the Hb of left and right hemispheres causes a reliable influence. io. er. each other via its connecting commissure in terms of anatomy (Kim, 2009). A few recent studies demonstrated significant behavioral effects also induced by the. al. n. iv n C application of unilateral stimulation (Friedman 2010, 2011; Li et al., 2011; Meng h e n g cethal., i U et al., 2011), which may support the aforementioned anatomical argument. So far, most (if not all) studies conducting with LHb stimulation may be due to a technical consideration. That is, the distance between the left and right hemispheres of habenula was about 1.4 mm.. This distance was not allowed to make the bilateral. implantation of two electrodes in the Hb.. Even by the manipulation of unilateral. stimulation in the LHb, behavioral changes on DRL 15-s behavior were significantly and reliably observed throughout this study. The results of DRL 15–s behavioral alterations produced by the present LHb stimulation are worthy to discuss.. According to a series of studies done by Hikosaka 29.
數據


相關文件
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
By kinematic constraints, we mean two kinds of constraints imposing on the 4-momenta of the invisible particles: the mass shell constraints and the measured missing transverse
Table 3 Numerical results for Cadzow, FIHT, PGD, DRI and our proposed pMAP on the noisy signal recovery experiment, including iterations (Iter), CPU time in seconds (Time), root of
• elearning pilot scheme (Four True Light Schools): WIFI construction, iPad procurement, elearning school visit and teacher training, English starts the elearning lesson.. 2012 •
Analysed by type of courses being taken and school sector of the graduates, among the 2014/15 S6 graduates who were pursuing full-time study, graduates of government schools
Microphone and 600 ohm line conduits shall be mechanically and electrically connected to receptacle boxes and electrically grounded to the audio system ground point.. Lines in
Comparing mouth area images of two different people might be deceptive because of different facial features such as the lips thickness, skin texture or teeth structure..
